
Fungicidal Activity and Mechanism of Action of Glabridin from Glycyrrhiza glabra L.


Abstract
:1. Introduction
2. Results and Discussion
2.1. In Vitro Fungicidal Activity
2.2. In Vivo Fungicidal Activity of Glabridin
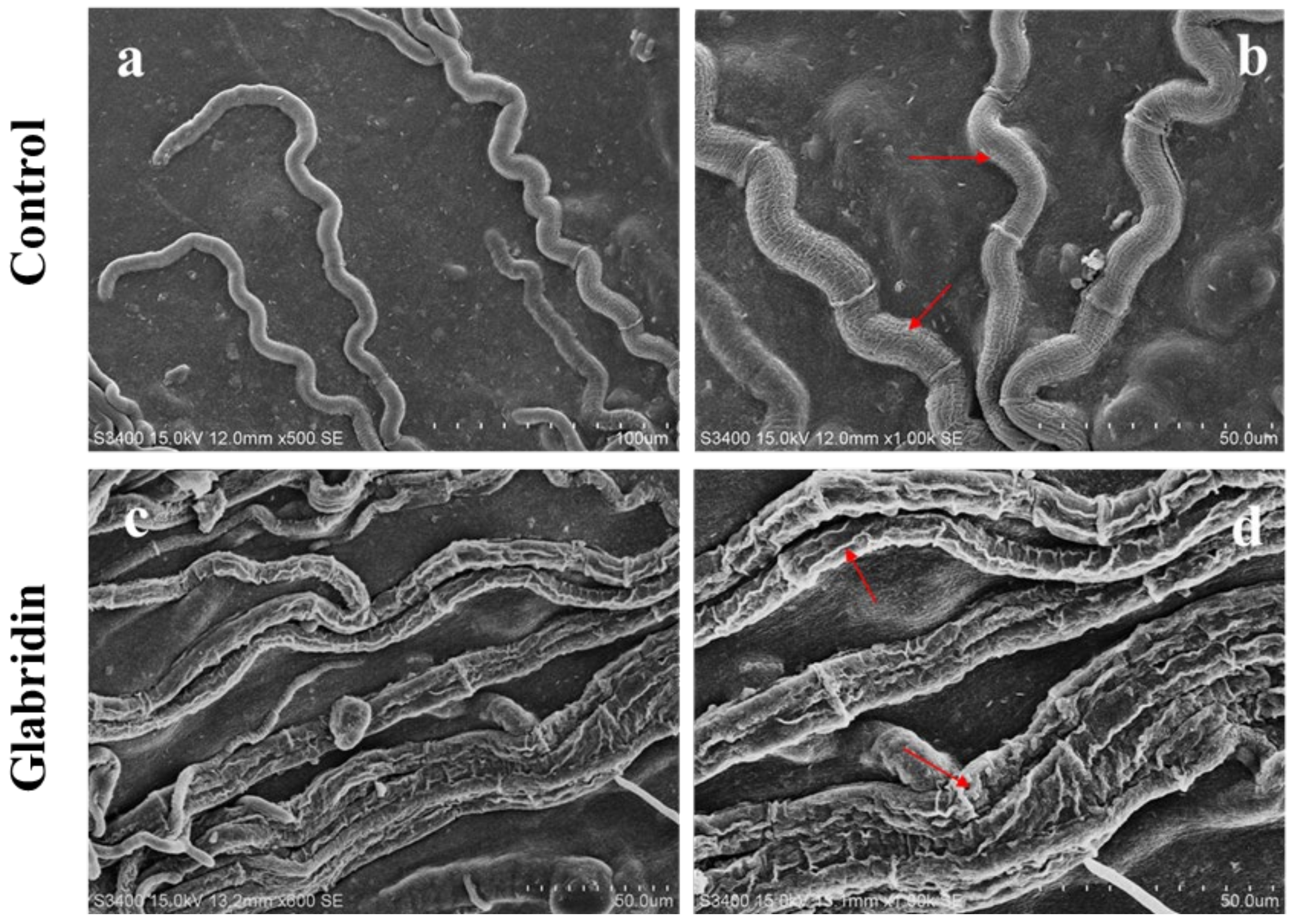
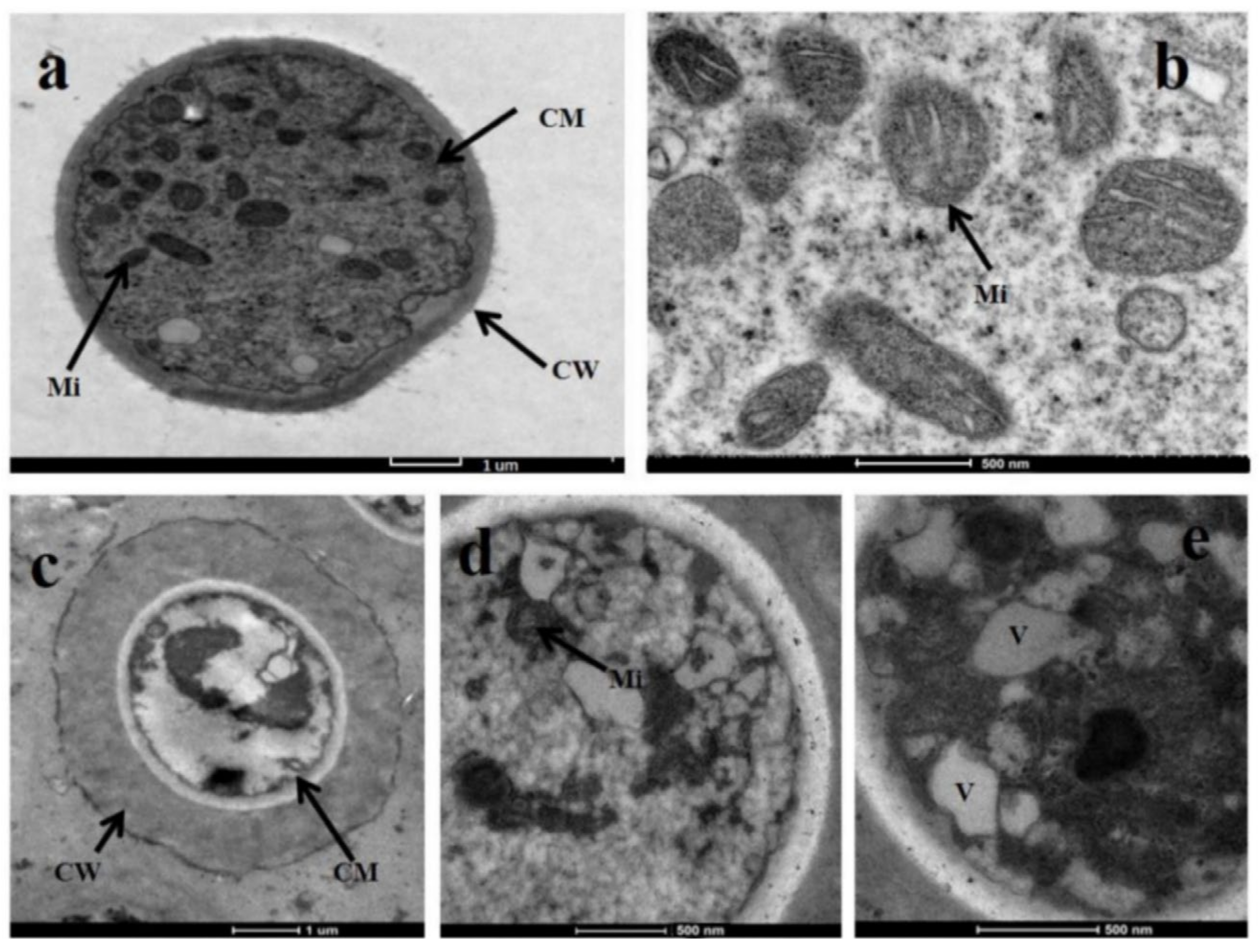
2.3. Effect of Glabridin on the Morphology of S. sclerotiorum
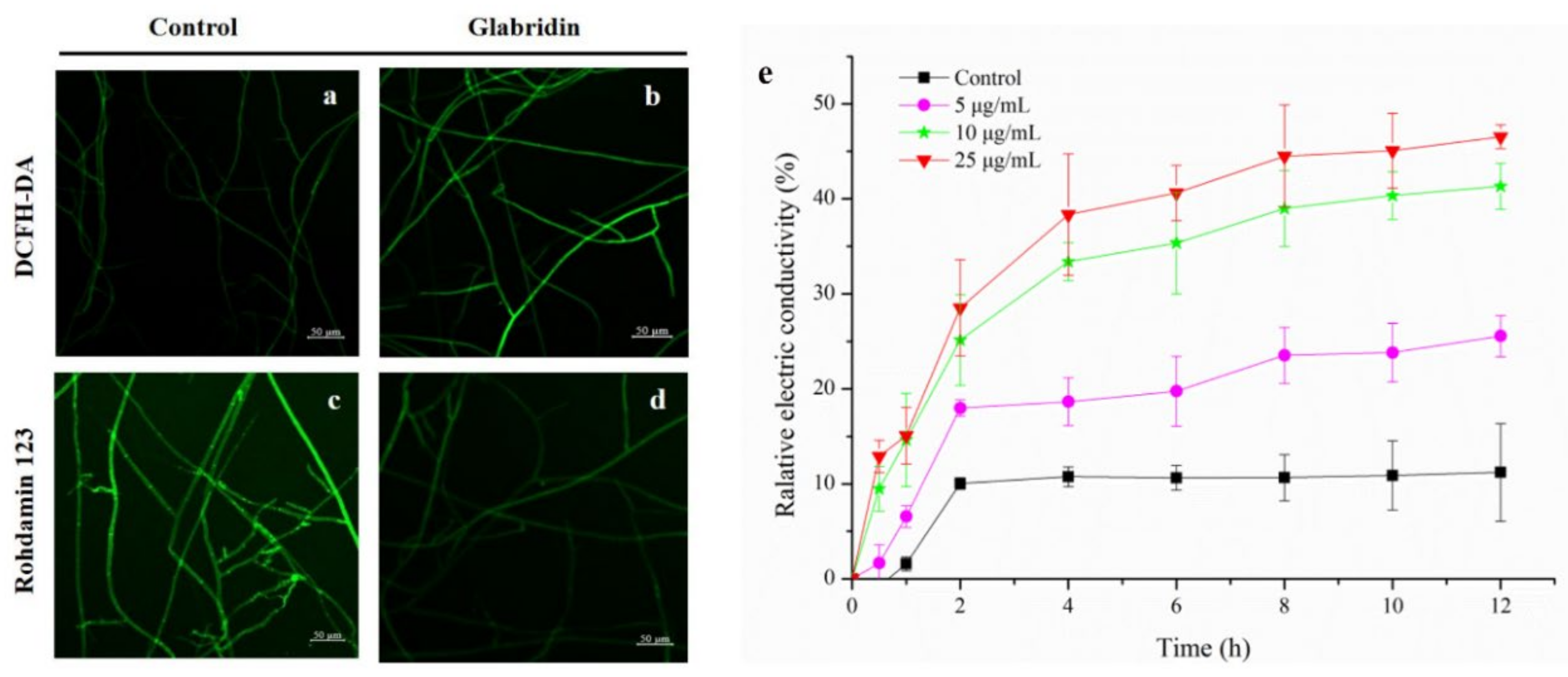
2.4. Effect of Glabridin on the Permeability of Cell Membrane
2.5. Effect of Glabridin on ROS Production of S. sclerotiorum
2.6. Effect of Glabridin on MMP of S. sclerotiorum
2.7. Transcriptomic Analysis
2.7.1. RNA- Sequencing, Assembly and Annotation
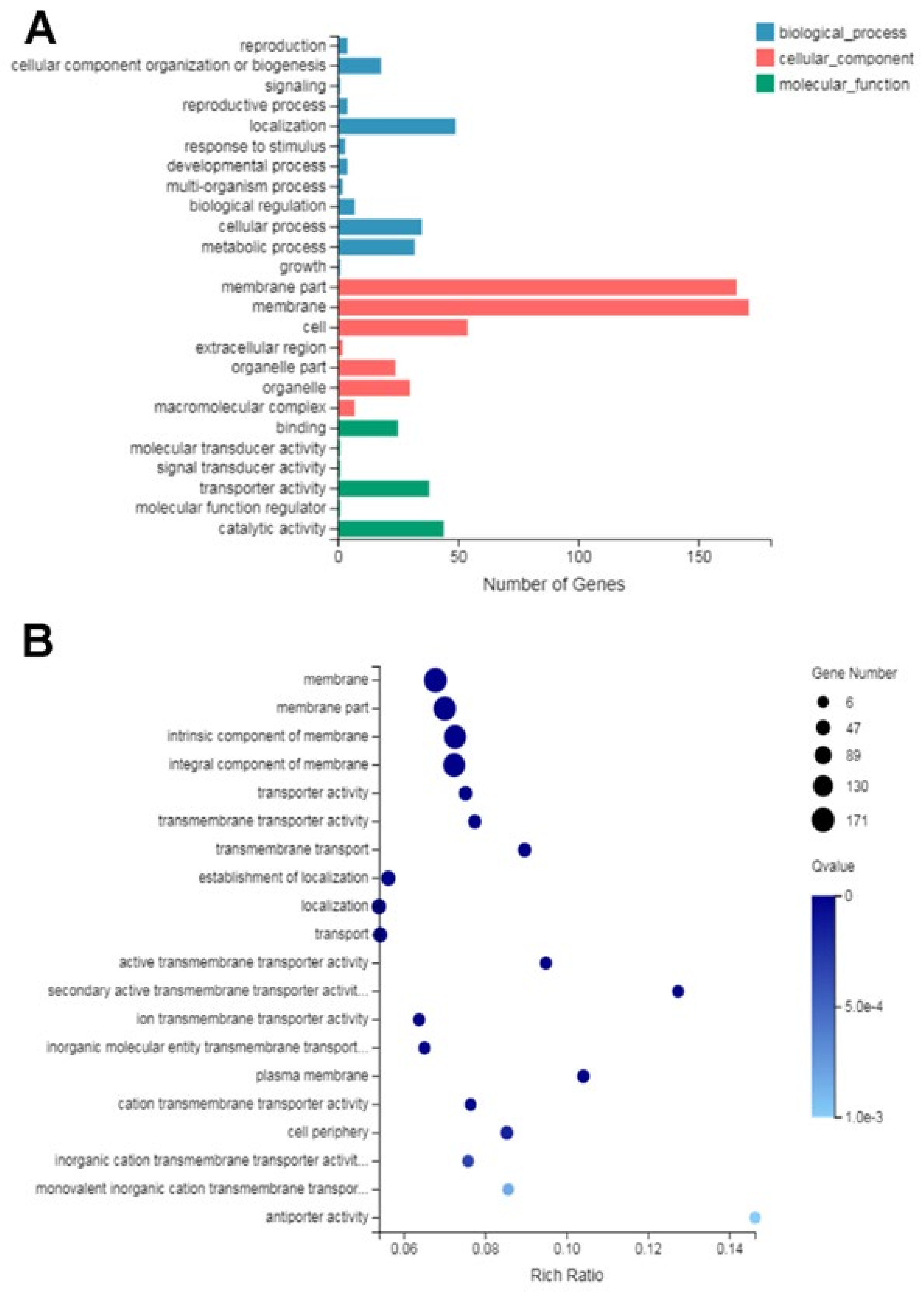
2.7.2. Gene Ontology (GO) Functional Analysis of DEGs
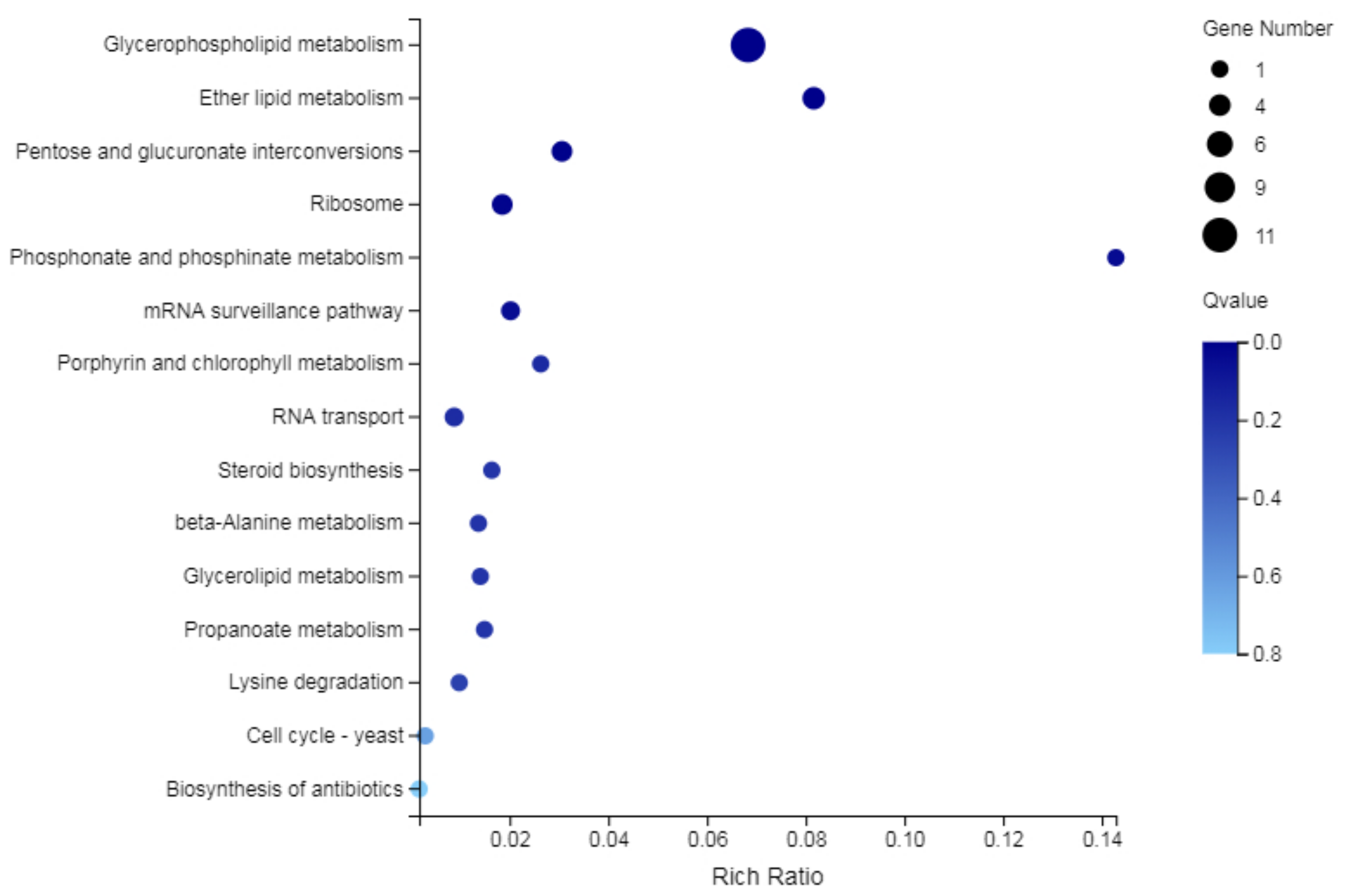
2.7.3. KEGG Functional Analysis of DEGs
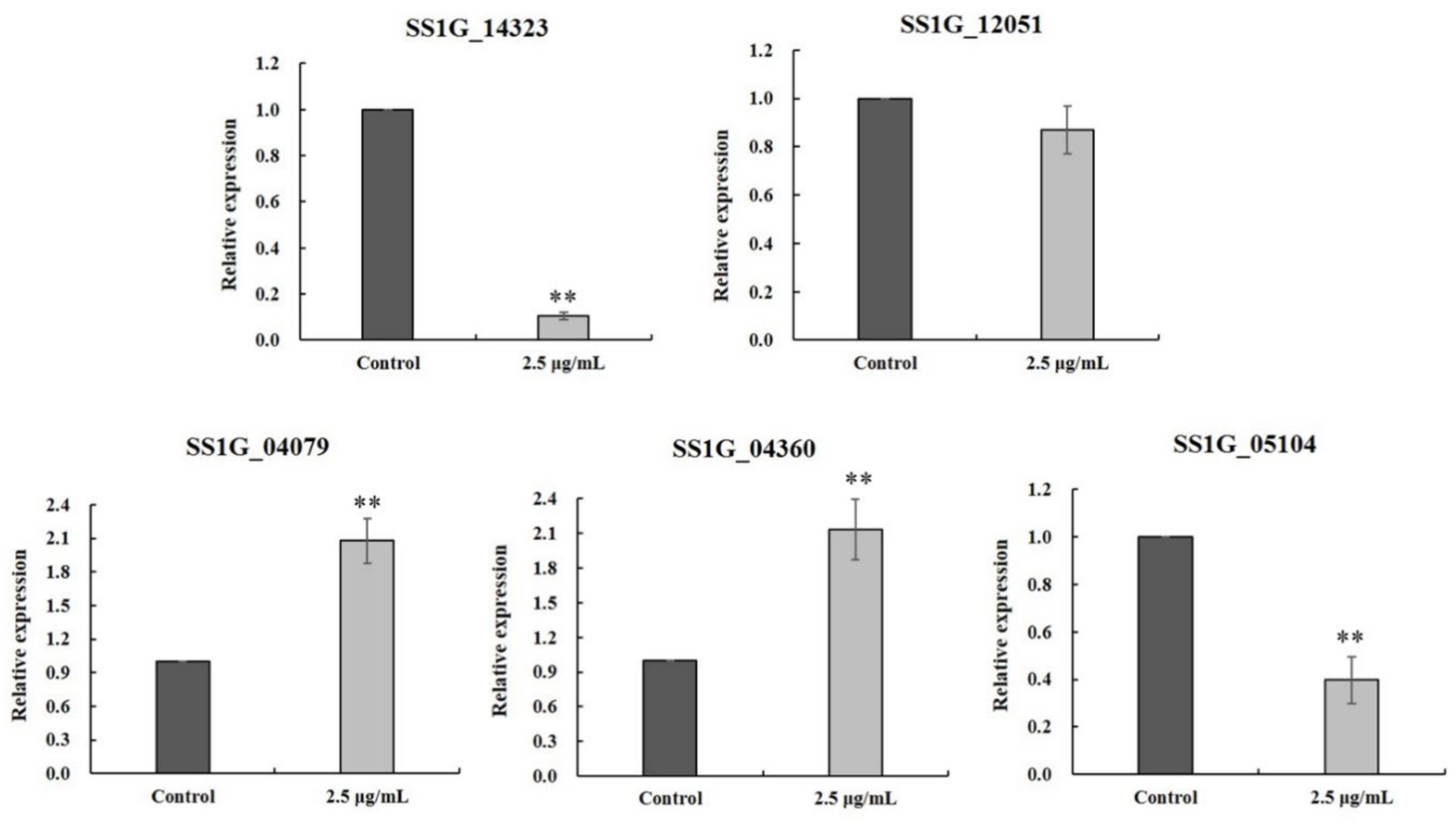
2.8. RT-qPCR Verification
2.9. Effect of Glabridin on Sclerotia Formation and Germination
3. Materials and Methods
3.1. Pathogenic Fungi and Reagents
3.2. In Vitro Antifungal Assay
3.3. In Vivo Fungicidal Assay
3.4. Transcriptomics Analysis
3.5. RT-qPCR Verification
3.6. Scanning Electron Microscopy (SEM) Observations
3.7. Transmission Electron Microscopy (TEM) Observations
3.8. Effect on Cell Membrane Permeability
3.9. Effect on the Reactive Oxygen Species (ROS)
3.10. Effect on the Mitochondrial Membrane Potential (MMP)
3.11. Effects on Sclerotia Formation and Germination
3.12. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gautam, A.K.; Sharma, S.; Avasthi, S.; Bhadauria, R. Diversity, pathogenicity and toxicology of A. niger: An important spoilage fungi. Res. J. Microbiol. 2011, 6, 270–280. [Google Scholar] [CrossRef]
- Isman, M.B.; Grieneisen, M.L. Botanical insecticide research: Many publications, limited useful data. Trends Plant Sci. 2014, 19, 140–145. [Google Scholar] [CrossRef]
- Jürgen, S. Modern Trends in Agrochemistry. Bioorg. Med. Chem. 2009, 17, 2019. [Google Scholar]
- Ghorbanpour, M.; Omidvari, M.; Abbaszadehdahaji, P.; Omidvar, R.; Kariman, K. Mechanisms underlying the protective effects of beneficial fungi against plant diseases. Biol. Control 2018, 117, 147–157. [Google Scholar] [CrossRef]
- Hejazi, I.I.; Khanam, R.; Mehdi, S.H.; Bhat, A.R.; Rizvi, M.M.A.; Islam, A.; Thakur, S.C.; Athar, F. New insights into the antioxidant and apoptotic potential of Glycyrrhiza glabra L. during hydrogen peroxide mediated oxidative stress: An in vitro and in silico evaluation. Biomed. Pharmacother. 2017, 94, 265–279. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yan, L.-T.; Yuan, E.-L.; Ding, H.-X.; Ye, H.-C.; Zhang, Z.-K.; Yan, C.; Liu, Y.-Q.; Feng, G. Antifungal activity of compounds extracted from Cortex Pseudolaricis against Colletotrichum gloeosporioides. J. Agric. Food Chem. 2014, 62, 4905–4910. [Google Scholar] [CrossRef]
- Zotte, A.D.; Celia, C.; Cullere, M.; Szendrő, Z.; Kovács, M.; Gerencsér, Z.; Bosco, A.D.; Giaccone, V.; Matics, Z. Effect of an in vivo and/or in-meat application of a liquorice (Glycyrrhiza glabra L.) extract on fattening rabbits live performance, carcass traits and meat quality. Anim. Feed. Sci. Technol. 2020, 260, 114333. [Google Scholar] [CrossRef]
- Pastorino, G.; Cornara, L.; Soares, S.; Rodrigues, F.; Oliveira, M.B.P.P. Liquorice (Glycyrrhiza glabra): A phytochemical and pharmacological review. Phytother. Res. 2018, 32, 2323–2339. [Google Scholar] [CrossRef] [PubMed]
- Naidu, K.C.; Lalam, R.; Bobbarala, V. Antimicrobial agents from Rubia cordifolia and Glycyrrhiza glabra against phytopathogens of Gossypium. Int. J. PharmTech Res. 2009, 1, 1512–1518. [Google Scholar]
- Schuster, C.; Konstantinidou-Doltsinis, S.; Schmitt, A. Glycyrrhiza glabra extract protects plants against important phytopathogenic fungi. Commun. Agric. Appl. Biol. Sci. 2010, 75, 531–540. [Google Scholar]
- Khan, A.; Ahmad, A.; Akhtar, F.; Yousuf, S.; Xess, I.; Khan, L.A.; Manzoor, N. Induction of oxidative stress as a possible mechanism of the antifungal action of three phenylpropanoids. FEMS Yeast Res. 2011, 11, 114–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.-J.; Jiang, Z.-Y.; Zhu, Q.; Zhong, G.-H. Discovery of β-carboline oxadiazole derivatives as fungicidal agents against rice sheath blight. J. Agric. Food Chem. 2018, 66, 9598–9607. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ren, X.; Guo, W.; Wang, D.; Han, L.; Feng, J. Oxidative stress and apoptosis of Gaeumannomyces graminis (Get) induced by carabrone. J. Agric. Food Chem. 2019, 67, 10448–10457. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Cheng, S.-T.; Wang, H.-Y.; Wu, J.-H.; Luo, Y.-M.; Wang, Q.; Wang, F.-X.; Xia, G.-X. iTRAQ-based proteomic analysis of defence responses triggered by the necrotrophic pathogen Rhizoctonia solani in cotton. J. Proteomics 2017, 152, 226–235. [Google Scholar] [CrossRef] [Green Version]
- Tang, L.; Chi, H.; Li, W.; Zhang, L.; Zhang, L.; Chen, L.; Zou, S.; Liu, H.; Liang, Y.; Yu, J.; et al. FgPsd2, a phosphatidylserine decarboxylase of Fusarium graminearum, regulates development and virulence. Fungal Genet. Biol. 2021, 146, 103483. [Google Scholar] [CrossRef]
- Yang, G.-Z.; Zhu, J.-K.; Yin, X.-D.; Yan, Y.-F.; Wang, Y.-L.; Shang, X.-F.; Liu, Y.-Q.; Zhao, Z.-M.; Peng, J.-W.; Liu, H. Design, synthesis, and antifungal evaluation of novel quinoline derivatives inspired from natural quinine alkaloids. J. Agric. Food Chem. 2019, 67, 11340–11353. [Google Scholar] [CrossRef]
- Zhao, Z.-M.; Shang, X.-F.; Lawoe, R.K.; Liu, Y.-Q.; Zhou, R.; Sun, Y.; Yan, Y.-F.; Li, J.-C.; Yang, G.-Z.; Yang, C.-J. Anti-phytopathogenic activity and the possible mechanisms of action of isoquinoline alkaloid sanguinarine. Pestic. Biochem. Physiol. 2019, 159, 51–58. [Google Scholar] [CrossRef]
- Wolf, J.B.W. Principles of transcriptome analysis and gene expression quantification: An RNA-seq tutorial. Mol. Ecol. Resour. 2013, 13, 559–572. [Google Scholar] [CrossRef]
- Zhang, X.; Wu, F.; Gu, N.; Yan, X.; Wang, K.; Dhanasekaran, S.; Gu, X.; Zhao, L.; Zhang, H. Postharvest biological control of Rhizopus rot and the mechanisms involved in induced disease resistance of peaches by Pichia membranefaciens. Postharvest Biol. Technol. 2020, 163, 111146. [Google Scholar] [CrossRef]
- Feng, G.; Zhang, X.-S.; Zhang, Z.-K.; Ye, H.-C.; Liu, Y.-Q.; Yang, G.-Z.; Chen, C.; Chen, M.; Yan, C.; Wang, L.-Y.; et al. Fungicidal activities of camptothecin semisynthetic derivatives against Colletotrichum gloeosporioides in vitro and in mango fruit. Postharvest Biol. Technol. 2019, 147, 139–147. [Google Scholar] [CrossRef]
- Li, J.-C.; Wang, R.-X.; Sun, Y.; Zhu, J.-K.; Hu, G.-F.; Wang, Y.-L.; Zhou, R.; Zhao, Z.-M.; Liu, Y.-Q.; Peng, J.-W.; et al. Design, synthesis and antifungal activity evaluation of isocryptolepine derivatives. Bioorg. Chem. 2019, 92, 103266. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sun, Y.; Zhang, Y.; Zhang, Y.X.; Han, L.; Zhang, X.; Feng, J. Sensitivity and biochemical characteristics of Sclerotinia sclerotiorum to propamidine. Pestic. Biochem. Physiol. 2017, 135, 82–88. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
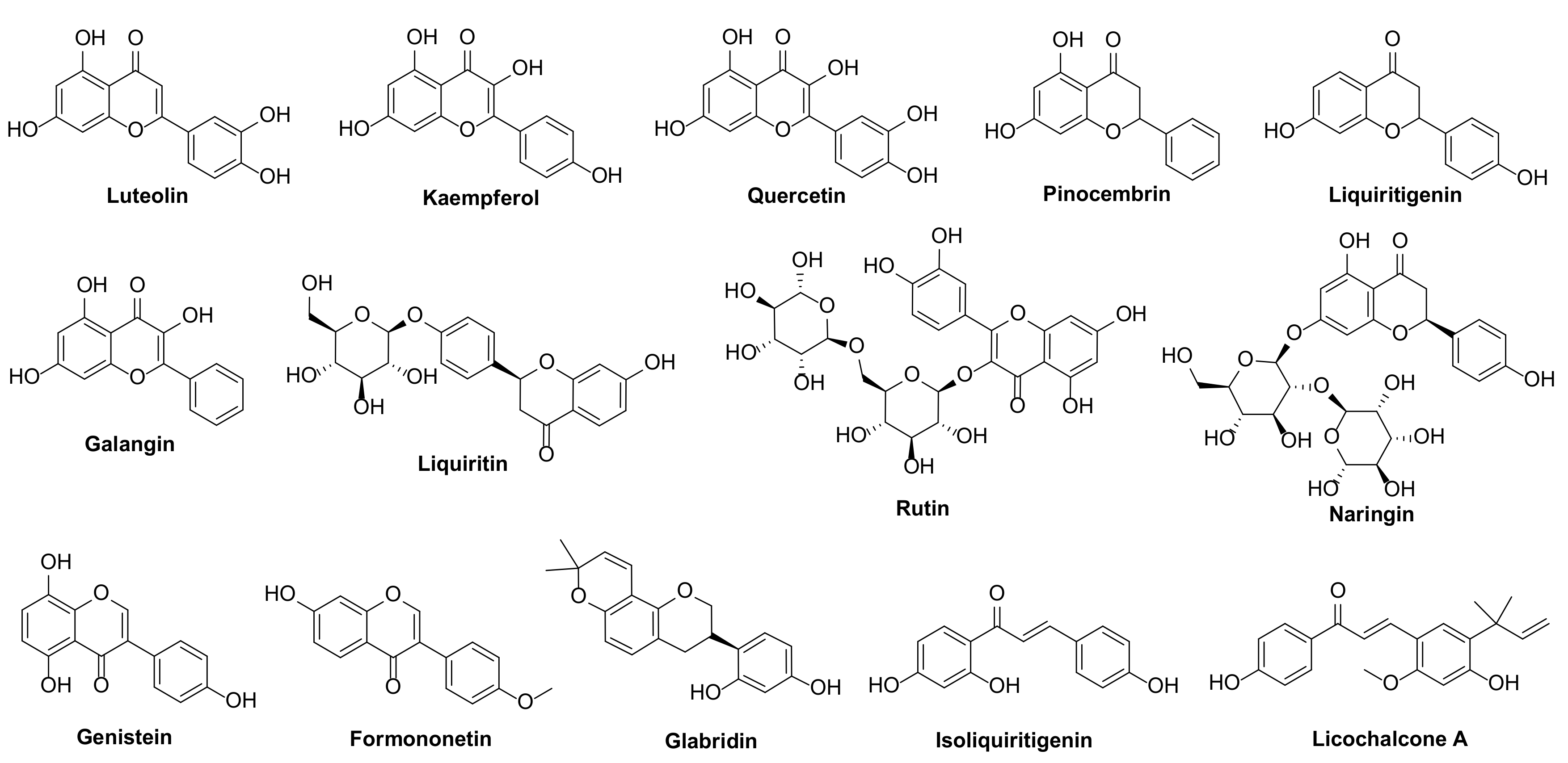
| Flavonoids | Inhibition Rate (%) ± SD | |||
|---|---|---|---|---|
| S. sclerotiorum | R. solani | F. graminearum | B. cinerea | |
| Luteolin | 35.20 ± 0.70 | NA | 4.63 ± 0.34 | 35.08 ± 1.69 |
| Kaempferol | NA | 26.24 ± 1.18 | 2.97 ± 0.37 | 16.02 ± 0.67 |
| Quercetin | 12.56 ± 0.35 | NA | 14.37 ± 0.29 | 32.78 ± 1.56 |
| Pinocembrin | 18.14 ± 1.25 | 51.88 ± 0.60 | 18.93 ± 1.54 | 31.65 ± 0.48 |
| Liquiritigenin | 30.02 ± 0.57 | 30.50 ± 0.41 | 53.26 ± 0.71 | 36.48 ± 1.57 |
| Galangin | NA | 10.90 ± 0.95 | 20.40 ± 1.20 | 13.92 ± 0.80 |
| Liquiritin | 1.91 ± 0.40 | 5.58 ± 0.68 | 2.39 ± 0.60 | 3.00 ± 0.54 |
| Rutin | 24.42 ± 0.32 | 22.80 ± 0.51 | 24.38 ± 0.43 | 37.79 ± 0.61 |
| Naringin | 7.59 ± 0.98 | 9.68 ± 1.17 | 2.34 ± 0.82 | 11.62 ± 0.98 |
| Genistein | 4.30 ± 0.69 | 4.30 ± 0.69 | 34.25 ± 1.48 | 21.10 ± 0.52 |
| Formononetin | NA | 2.87 ± 0.45 | 6.28 ± 0.80 | 6.63 ± 0.69 |
| Glabridin | 100 ± 0.00 | 100 ± 0.00 | 96.21 ± 0.76 | 93.12 ± 0.81 |
| Isoliquiritigenin | 31.71 ± 0.70 | 34.15 ± 0.85 | 55.29 ± 0.58 | 51.38 ± 0.44 |
| Licochalcone A | 76.10 ± 0.54 | 80.19 ± 0.54 | 18.58 ± 0.52 | 73.28 ± 0.64 |
| Azoxystrobin | 97.13 ± 0.47 | 100 ± 0.00 | 100 ± 0.00 | 100 ± 0.00 |
| Compounds | Pathogenic Fungi | Virulence Equation (Y = ax + b) | Correlation Coefficient (R2) | EC50 (µg/mL) |
|---|---|---|---|---|
| Glabrindin | B. cinerea | y = 0.6094x + 4.4290 | 0.8672 | 8.65 |
| S. sclerotiorum | y = 2.2922x + 2.7926 | 0.9295 | 6.78 | |
| F. graminearum | y = 1.0731x + 3.6297 | 0.9452 | 18.92 | |
| M. oryae | y = 1.4210x + 3.2215 | 0.8943 | 17.85 | |
| F. oxysporum | y = 0.6476x + 4.4204 | 0.9833 | 9.85 | |
| R. solani | y = 2.1669x + 2.4477 | 0.9699 | 15.06 | |
| M. melonis | y = 1.3578x + 3.4252 | 0.9717 | 14.45 | |
| P. capsici | y = 1.9803x + 2.1392 | 0.9845 | 27.84 | |
| T. cucumeris | y = 0.4647x − 1.0538 | 0.8924 | 8.33 | |
| C. gloeosporioides | y = 0.4889x − 0.7973 | 0.9545 | 44.97 | |
| Azoxystrobin | B. cinerea | - | - | >50 |
| S. sclerotiorum | - | - | >50 | |
| F. graminearum | y = 2.0532x − 8.8182 | 0.8882 | 27.43 | |
| M. oryae | y = 0.9405x − 3.6062 | 0.9641 | 12.04 | |
| F. oxysporum | y = 2.3424x − 10.265 | 0.9946 | 27.96 | |
| R. solani | y =1.4638x − 5.6362 | 0.9400 | 49.29 | |
| M. melonis | y = 2.0431x − 8.9002 | 0.9744 | 20.40 | |
| P. capsici | - | - | >50 | |
| T. cucumeris | y = 0.571x + 4.1224 | 0.9347 | 34.43 | |
| C.gloeosporioides | - | - | >50 |
| Compounds | Concentration (μg/mL) | Protective Effect | Curative Effect | ||
|---|---|---|---|---|---|
| Lesion Length (mm ± SD) | Control Efficacy (%) | Lesion Length (mm ± SD) | Control Efficacy (%) | ||
| Glabridin | 50 | 13.70 ± 0.96 | 23.75 | 17.65 ± 1.14 | 33.60 |
| 100 | 11.20 ± 0.89 | 38.91 | 11.82 ± 0.99 | 64.22 | |
| 200 | 9.30 ± 1.01 | 76.54 | 8.50 ± 1.07 | 81.63 | |
| Azoxystrobin | 25 | 8.10 ± 0.75 | 71.82 | 7.82 ± 0.42 | 85.21 |
| 50 | 7.00 ± 0.08 | 81.40 | 6.58 ± 0.44 | 91.69 | |
| 100 | 6.00 ± 0.10 | 90.13 | 6.28 ± 0.19 | 93.26 | |
| Control | - | 17.65 ± 1.70 | - | 24.05 ± 1.13 | - |
| Sample | Raw Reads (Mb) | Clean Reads (Mb) | Clean Bases (Gb) | Q20 (%) | Q30 (%) | Clean Reads Ratio (%) |
|---|---|---|---|---|---|---|
| CO1 | 49.08 | 44.1 | 6.61 | 97.19 | 89.34 | 89.84 |
| CO2 | 49.08 | 44.65 | 6.7 | 97.28 | 89.6 | 90.97 |
| CO3 | 47.33 | 43.16 | 6.47 | 97.51 | 90.11 | 91.19 |
| TR1 | 47.33 | 43.21 | 6.48 | 97.14 | 89.15 | 91.3 |
| TR2 | 47.33 | 42.81 | 6.42 | 97.22 | 89.44 | 90.46 |
| TR3 | 47.33 | 42.63 | 6.39 | 97.12 | 89.14 | 90.08 |
| CO summary | 145.49 | 131.91 | 19.78 | 97.33 | 89.68 | 90.67 |
| TR summary | 141.99 | 128.65 | 19.29 | 97.16 | 89.24 | 90.61 |
| Summary | 287.48 | 260.56 | 39.07 | 97.25 | 89.46 | 90.64 |
| Gene Symbol | FPKM (Control) | FPKM (Treat) | Log2FC | Annotation | Primer Sequences |
|---|---|---|---|---|---|
| SSIG_14322 | 23.42 | 7.45 | −1.66 | phosphatidylserine decarboxylase | F- GTGGTGGGAGTGATCCTTATATC R- GAGTGACAAGCAAAGCACAAG |
| SSIG_12051 | 4.06 | 1.91 | −1.04 | phosphatidylserine decarboxylase | F- CGGATTCACCACGACGATAAT R- GACTTCCACATGGACTCGTAAG |
| SSIG_05104 | 1.86 | 5.84 | 1.65 | phosphatidylserine decarboxylase | F- AGCAAATCAGGCTGGAGATAG R- GTCTGAGTAATAACCGTCGTCTT |
| SSIG_04079 | 8.86 | 18.70 | 1.12 | phosphatidylserine decarboxylase | F- AGCAAATCAGGCTGGAGATAG R- GTCTGAGTAATAACCGTCGTCTT |
| SSIG_04360 | 6.13 | 3.19 | −1.16 | phosphatidylserine decarboxylase | F- CCGCTGTCATCAGAACCTATT R- CTACTGCGCACCATACGATAA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, A.; Zhao, Z.; Zhang, S.; Zhang, Z.; Shi, Y. Fungicidal Activity and Mechanism of Action of Glabridin from Glycyrrhiza glabra L. Int. J. Mol. Sci. 2021, 22, 10966. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222010966
Li A, Zhao Z, Zhang S, Zhang Z, Shi Y. Fungicidal Activity and Mechanism of Action of Glabridin from Glycyrrhiza glabra L. International Journal of Molecular Sciences. 2021; 22(20):10966. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222010966
Chicago/Turabian StyleLi, Anping, Zhongmin Zhao, Shaoyong Zhang, Zhijun Zhang, and Yanping Shi. 2021. "Fungicidal Activity and Mechanism of Action of Glabridin from Glycyrrhiza glabra L." International Journal of Molecular Sciences 22, no. 20: 10966. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222010966