
The Expression and Localisation of G-Protein-Coupled Inwardly Rectifying Potassium (GIRK) Channels Is Differentially Altered in the Hippocampus of Two Mouse Models of Alzheimer’s Disease

 , , , and
, , , and 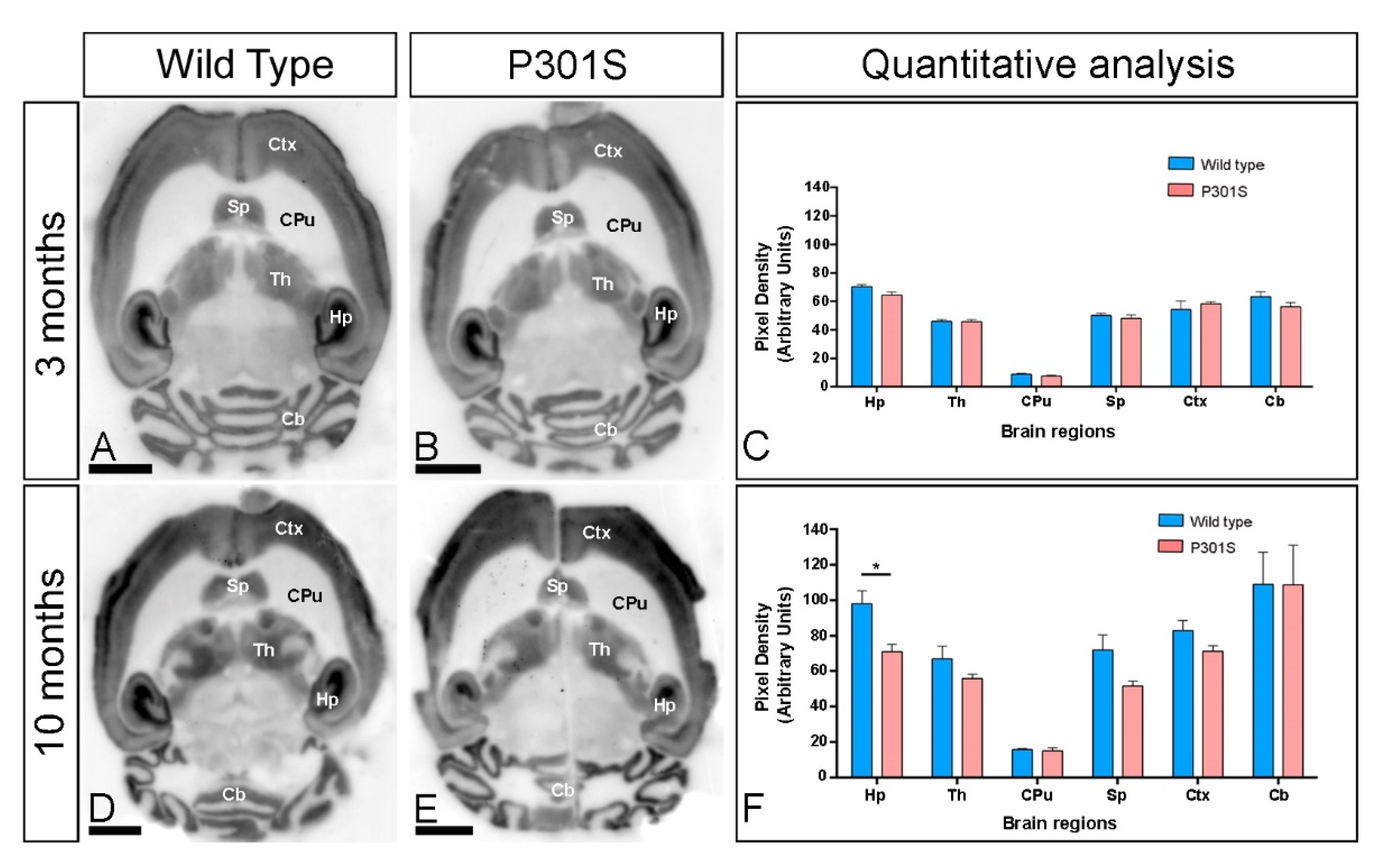{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Brain Expression of GIRK2 Channels in P301S Mice
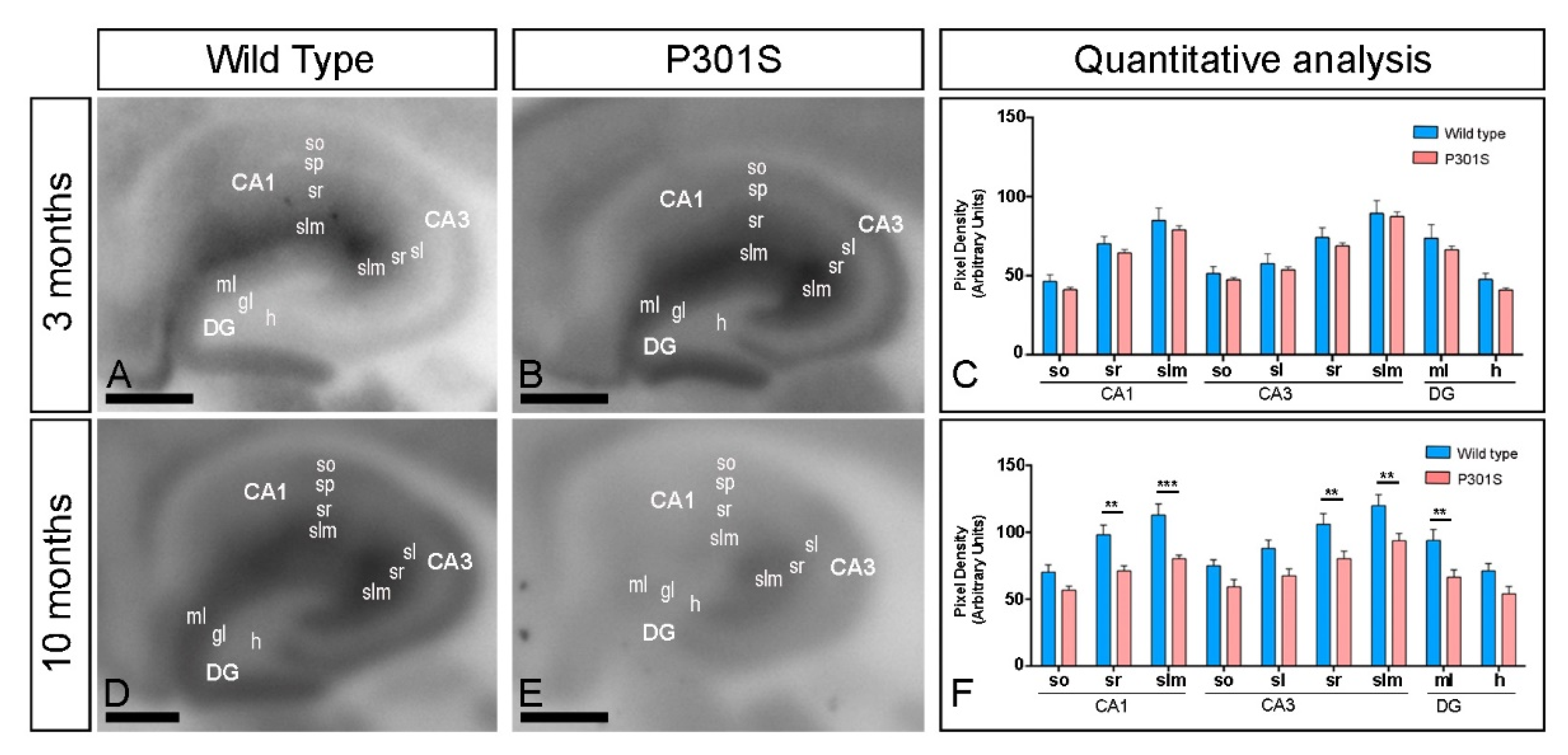
2.2. Laminar Expression of GIRK2 Channels in the Hippocampus of P301S Mice
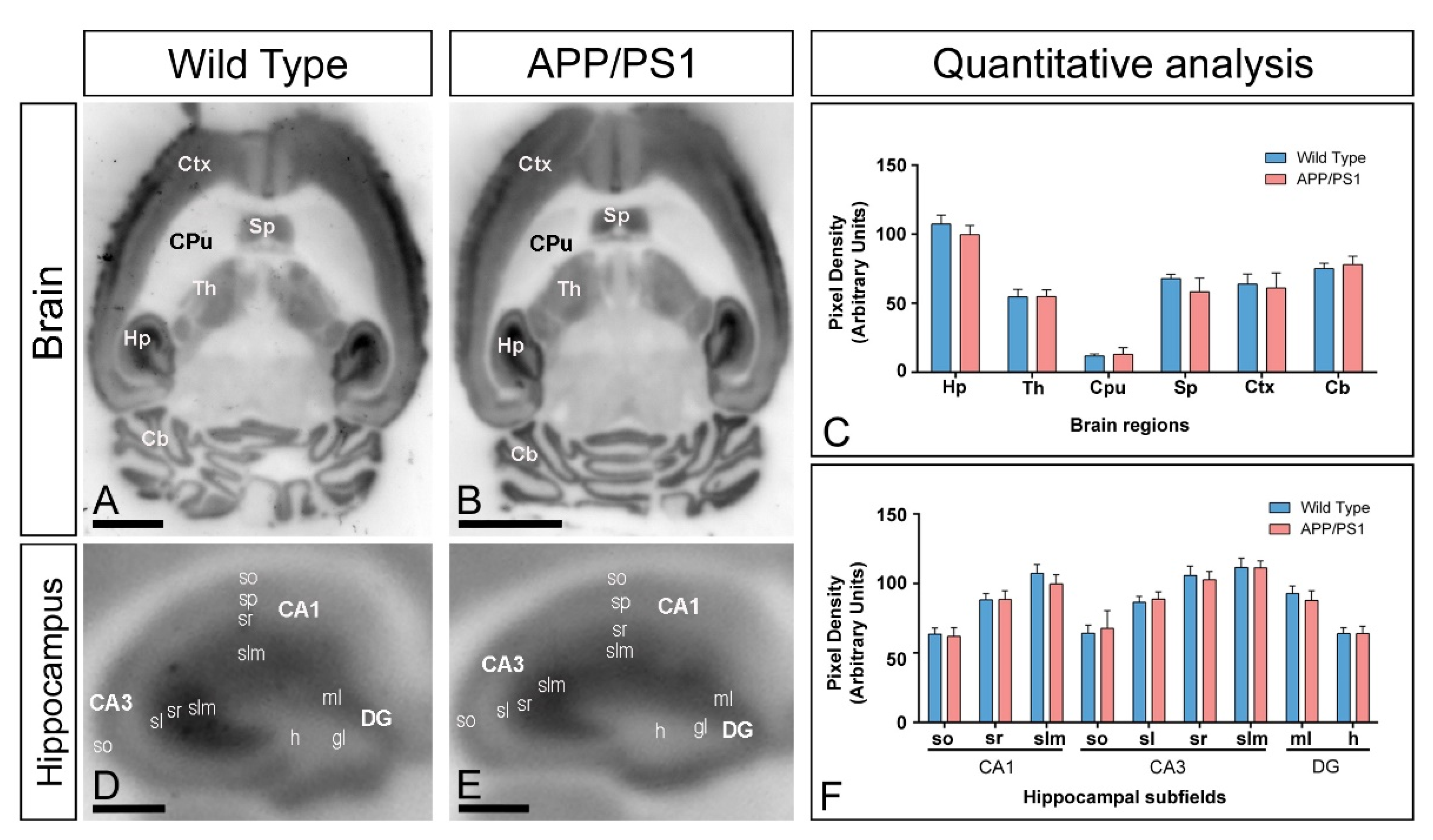
2.3. Similar Brain and Hippocampal Expression of GIRK2 Channels in APP/PS1 Mice
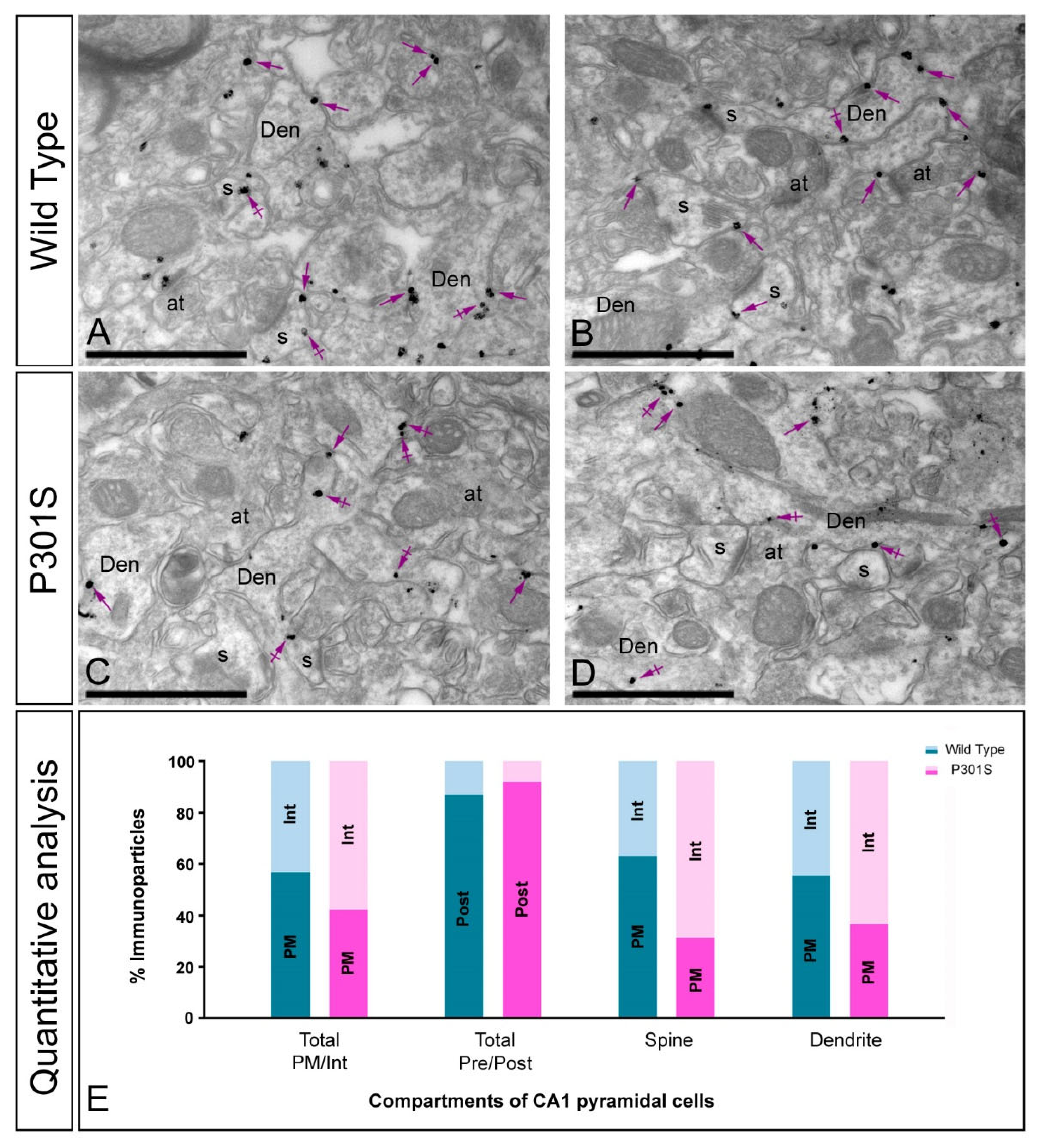
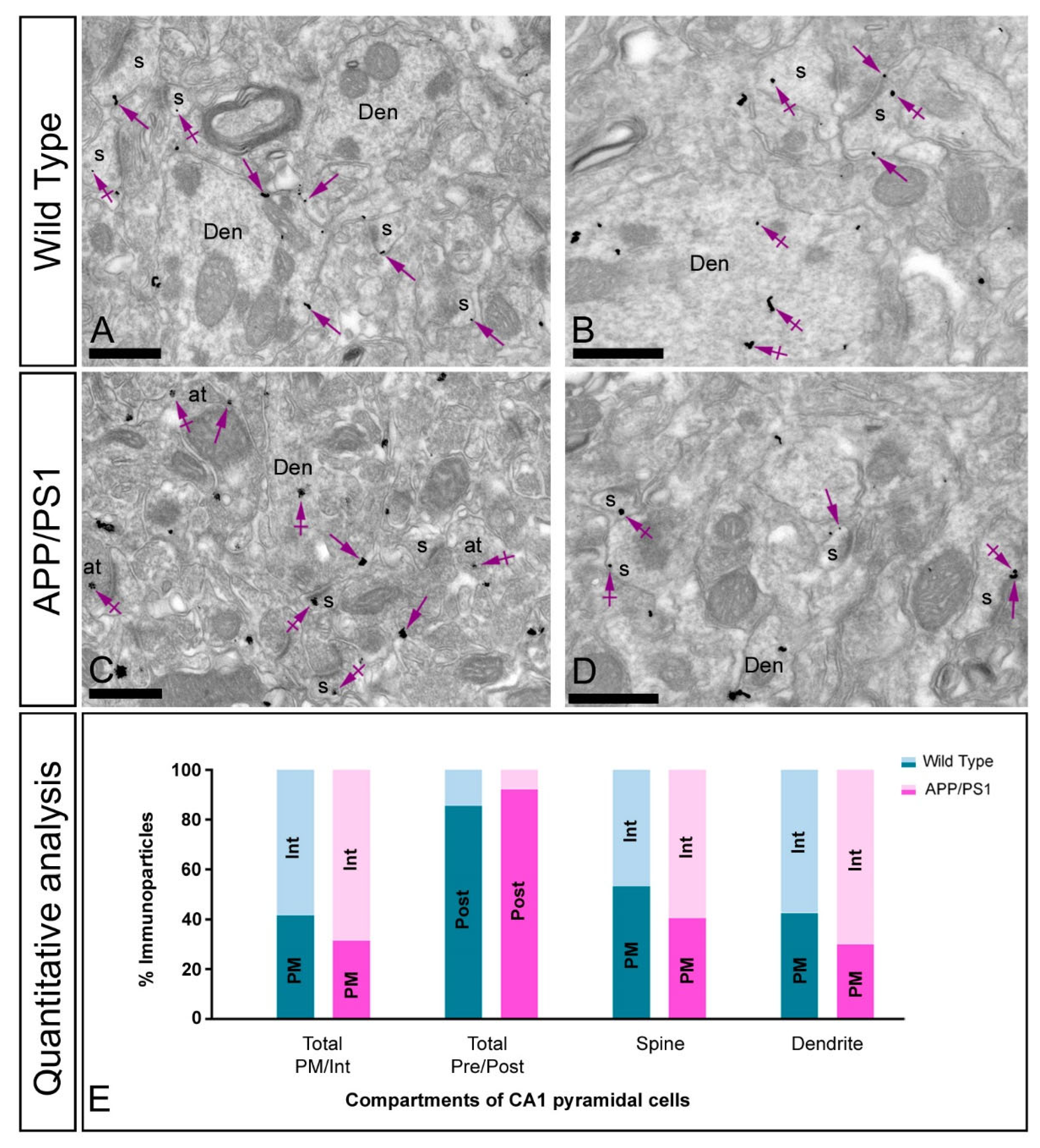
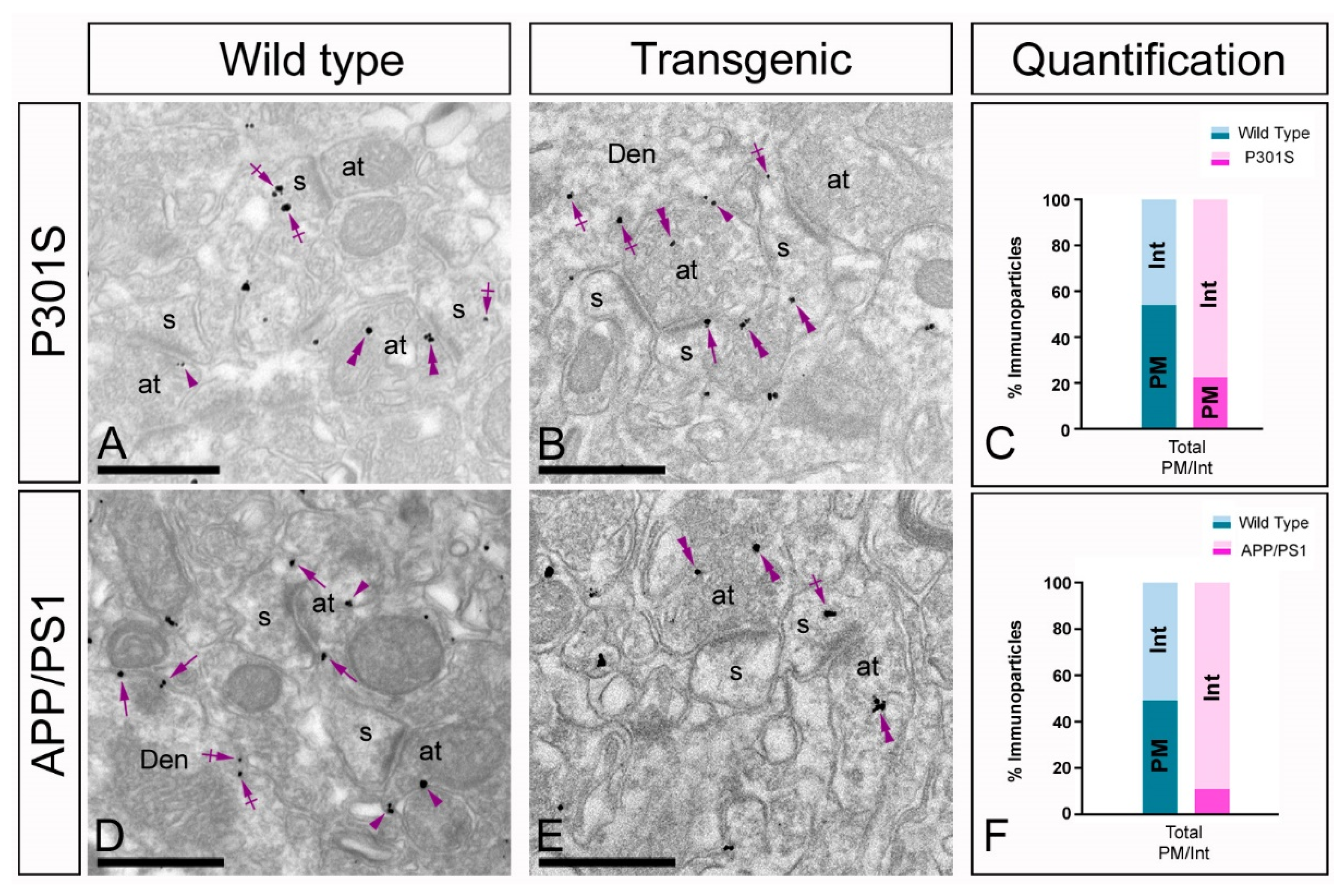
2.4. Altered Postsynaptic Localisation of GIRK2 in the Hippocampus of Transgenic Mice
2.5. Altered Presynaptic Localisation of GIRK2 in the Hippocampus of Transgenic Mice
3. Discussion
3.1. Differential Expression Pattern of GIRK Channels in Transgenic Mice
3.2. Altered Postsynaptic Localisation of GIRK Channels in Transgenic Mice
3.3. Altered Presynaptic Distribution of GIRK Channels in Transgenic Mice
4. Material and Methods
4.1. Animals
4.2. Antibodies and Chemicals
4.3. Histoblotting
4.4. Immunohistochemistry for Electron Microscopy
4.5. Quantification and Analysis of Pre-Embedding Immunogold Labelling
4.6. Controls
4.7. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hibino, H.; Inanobe, A.; Furutani, K.; Murakami, S.; Findlay, I.; Kurachi, Y. Inwardly Rectifying Potassium Channels: Their Structure, Function, and Physiological Roles. Physiol. Rev. 2010, 90, 291–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lüscher, C.; Slesinger, P.A. Emerging roles for G protein-gated inwardly rectifying potassium (GIRK) channels in health and disease. Nat. Rev. Neurosci. 2010, 11, 301–315. [Google Scholar] [CrossRef]
- Lüscher, C.; Jan, L.; Stoffel, M.; Malenka, R.C.; Nicoll, R.A. G Protein-Coupled Inwardly Rectifying K+ Channels (GIRKs) Mediate Postsynaptic but Not Presynaptic Transmitter Actions in Hippocampal Neurons. Neuron 1997, 19, 687–695. [Google Scholar] [CrossRef] [Green Version]
- Oldham, W.; Hamm, H.E. Heterotrimeric G protein activation by G-protein-coupled receptors. Nat. Rev. Mol. Cell Biol. 2008, 9, 60–71. [Google Scholar] [CrossRef]
- Thathiah, A.; de Strooper, B. The role of G protein-coupled receptors in the pathology of Alzheimer’s disease. Nat. Rev. Neurosci. 2011, 12, 73–87. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Carlsson, J.; Ambrogini, P.; Narváez, M.; Wydra, K.; Tarakanov, A.O.; Li, X.; Millón, C.; Ferraro, L.; Cuppini, R.; et al. Understanding the Role of GPCR Heteroreceptor Complexes in Modulating the Brain Networks in Health and Disease. Front. Cell. Neurosci. 2017, 11, 37. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, P.; Ramos, M.L.S.; Amaro, A.J.; Dias, R.; Vieira, S.I. Gi/o-Protein Coupled Receptors in the Aging Brain. Front. Aging Neurosci. 2019, 11, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayfield, J.; Blednov, Y.A.; Harris, R.A. Behavioral and Genetic Evidence for GIRK Channels in the CNS: Role in Physiology, Pathophysiology, and Drug Addiction. Int. Rev. Neurobiol. 2015, 123, 279–313. [Google Scholar]
- Walsh, K.B. Targeting GIRK Channels for the Development of New Therapeutic Agents. Front. Pharmacol. 2011, 2, 64. [Google Scholar] [CrossRef] [Green Version]
- Luján, R.; de Velasco, E.M.F.; Aguado, C.; Wickman, K. New insights into the therapeutic potential of Girk channels. Trends Neurosci. 2013, 37, 20–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeremic, D.; Sanchez-Rodriguez, I.; Jimenez-Diaz, L.; Navarro-Lopez, J.D. Therapeutic potential of targeting G protein-gated inwardly rectifying potassium (GIRK) channels in the central nervous system. Pharmacol. Ther. 2021, 223, 107808. [Google Scholar] [CrossRef]
- Zhao, Y.; Gameiro-Ros, I.; Glaaser, I.W.; Slesinger, P.A. Advances in Targeting GIRK Channels in Disease. Trends Pharmacol. Sci. 2021, 42, 203–215. [Google Scholar] [CrossRef]
- Yamada, M.; Inanobe, A.; Kurachi, Y. G protein regulation of potassium ion channels. Pharmacol. Rev. 1998, 50, 723–760. [Google Scholar] [PubMed]
- Karschin, C.; Dißmann, E.; Stühmer, W.; Karschin, A. IRK (1–3) and GIRK (1–4) Inwardly Rectifying K+ Channel mRNAs Are Differentially Expressed in the Adult Rat Brain. J. Neurosci. 1996, 16, 3559–3570. [Google Scholar] [CrossRef] [Green Version]
- Wickman, K.; Karschin, C.; Karschin, A.; Picciotto, M.R.; Clapham, D.E. Brain Localization and Behavioral Impact of the G-Protein-Gated K+Channel Subunit GIRK4. J. Neurosci. 2000, 20, 5608–5615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickman, K.; Pu, W.T.; Clapham, D.E. Structural characterization of the mouse Girk genes. Gene 2002, 284, 241–250. [Google Scholar] [CrossRef]
- Hearing, M.; Kotecki, L.; de Velasco, E.M.F.; Fajardo-Serrano, A.; Chung, H.J.; Luján, R.; Wickman, K. Repeated Cocaine Weakens GABAB-Girk Signaling in Layer 5/6 Pyramidal Neurons in the Prelimbic Cortex. Neuron 2013, 80, 159–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luján, R.; Aguado, C. Localization and Targeting of GIRK Channels in Mammalian Central Neurons. Int. Rev. Neurobiol. 2015, 123, 161–200. [Google Scholar] [CrossRef]
- Chung, H.J.; Ge, W.-P.; Qian, X.; Wiser, O.; Jan, Y.N.; Jan, L.Y. G protein-activated inwardly rectifying potassium channels mediate depotentiation of long-term potentiation. Proc. Natl. Acad. Sci. USA 2008, 106, 635–640. [Google Scholar] [CrossRef] [Green Version]
- Bloom, G.S. Amyloid-β and Tau. JAMA Neurol. 2014, 71, 505–508. [Google Scholar] [CrossRef] [Green Version]
- Nava-Mesa, M.O.; Jiménez-Díaz, L.; Yajeya, J.; Navarro-Lopez, J.D. Amyloid-β induces synaptic dysfunction through G protein-gated inwardly rectifying potassium channels in the fimbria-CA3 hippocampal synapse. Front. Cell. Neurosci. 2013, 7, 117. [Google Scholar] [CrossRef] [Green Version]
- Mayordomo-Cava, J.; Yajeya, J.; Navarro-Lopez, J.D.; Jiménez-Díaz, L. Amyloid-β (25-35) Modulates the Expression of GirK and KCNQ Channel Genes in the Hippocampus. PLoS ONE 2015, 10, e0134385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguado, C.; Luján, R. The histoblot technique: A reliable approach to analyse expression profile of proteins and to predict their molecular association. Neuromethods 2019, 144, 65–68. [Google Scholar]
- Fernández-Alacid, L.; Aguado, C.; Ciruela, F.; Martín, R.; Colón, J.; Cabañero, M.J.; Gassmann, M.; Watanabe, M.; Shigemoto, R.; Wickman, K.; et al. Subcellular compartment-specific molecular diversity of pre- and post-synaptic GABAB-activated GIRK channels in Purkinje cells. J. Neurochem. 2009, 110, 1363–1376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koyrakh, L.; Luján, R.; Colón, J.; Karschin, C.; Kurachi, Y.; Karschin, A.; Wickman, K. Molecular and Cellular Diversity of Neuronal G-Protein-Gated Potassium Channels. J. Neurosci. 2005, 25, 11468–11478. [Google Scholar] [CrossRef] [PubMed]
- Djebari, S.; Iborra-Lázaro, G.; Temprano-Carazo, S.; Sánchez-Rodríguez, I.; Nava-Mesa, M.O.; Múnera, A.; Gruart, A.; Delgado-García, J.M.; Jiménez-Díaz, L.; Navarro-López, J.D. G-Protein-Gated Inwardly Rectifying Potassium (Kir3/GIRK) Channels Govern Synaptic Plasticity That Supports Hippocampal-Dependent Cognitive Functions in Male Mice. J. Neurosci. 2021, 41, 7086–7102. [Google Scholar] [CrossRef]
- Belmonte, A.M.; Aguado, C.; Alfaro-Ruíz, R.; Moreno-Martínez, A.E.; de la Ossa, L.; Martinez-Hernandez, J.; Buisson, A.; Früh, S.; Bettler, B.; Shigemoto, R.; et al. Reduction in the neuronal surface of post and presynaptic GABA B receptors in the hippocampus in a mouse model of Alzheimer’s disease. Brain Pathol. 2019, 30, 554–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Rodríguez, I.; Temprano-Carazo, S.; Nájera, A.; Djebari, S.; Yajeya, J.; Gruart, A.; Delgado-García, J.M.; Jiménez-Díaz, L.; Navarro-López, J.D. Activation of G-protein-gated inwardly rectifying potassium (Kir3/GirK) channels rescues hippocampal functions in a mouse model of early amyloid-β pathology. Sci. Rep. 2017, 7, 14658. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Rodríguez, I.; Djebari, S.; Temprano-Carazo, S.; Vega-Avelaira, D.; Jiménez-Herrera, R.; Iborra-Lázaro, G.; Yajeya, J.; Jiménez-Díaz, L.; Navarro-López, J.D. Hippocampal long-term synaptic depression and memory deficits induced in early amyloidopathy are prevented by enhancing G-protein-gated inwardly rectifying potassium channel activity. J. Neurochem. 2019, 153, 362–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, T.; Ikeda, K.; Ichikawa, T.; Abe, S.; Togashi, S.; Kumanishi, T. Molecular Cloning of a Mouse G-Protein-Activated K+ Channel (mGIRK1) and Distinct Distributions of 3 GIRK (GIRK1, 2 and 3) mRNAs in Mouse Brain. Biochem. Biophys. Res. Commun. 1995, 208, 1166–1173. [Google Scholar] [CrossRef]
- Kulik, Á.; Vida, I.; Fukazawa, Y.; Guetg, N.; Kasugai, Y.; Marker, C.L.; Rigato, F.; Bettler, B.; Wickman, K.; Frotscher, M.; et al. Compartment-Dependent Colocalization of Kir3.2-Containing K+ Channels and GABAB Receptors in Hippocampal Pyramidal Cells. J. Neurosci. 2006, 26, 4289–4297. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Alacid, L.; Watanabe, M.; Molnár, E.; Wickman, K.; Luján, R. Developmental regulation of G protein-gated inwardly-rectifying K+ (GIRK/Kir3) channel subunits in the brain. Eur. J. Neurosci. 2011, 34, 1724–1736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, D.; Zerangue, N.; Raab-Graham, K.; Fried, S.R.; Jan, Y.N.; Jan, L. Diverse Trafficking Patterns Due to Multiple Traffic Motifs in G Protein-Activated Inwardly Rectifying Potassium Channels from Brain and Heart. Neuron 2002, 33, 715–729. [Google Scholar] [CrossRef] [Green Version]
- Hardy, J.; Selkoe, D.J. The amyloid hypothesis of Alzheimer’s disease: Progress and problems on the road to therapeutics. Science 2002, 297, 353–356. [Google Scholar] [CrossRef] [Green Version]
- Spires-Jones, T.L.; Hyman, B.T. The Intersection of Amyloid Beta and Tau at Synapses in Alzheimer’s Disease. Neuron 2014, 82, 756–771. [Google Scholar] [CrossRef] [Green Version]
- Ittner, A.; Ittner, L.M. Dendritic Tau in Alzheimer’s Disease. Neuron 2018, 99, 13–27. [Google Scholar] [CrossRef] [Green Version]
- Llorens-Martín, M.; Blazquez-Llorca, L.; Benavides-Piccione, R.; Rábano, A.; Hernandez, F.; Avila, J.; de Felipe, J.; Llorens-Martín, M. Selective alterations of neurons and circuits related to early memory loss in Alzheimer’s disease. Front. Neuroanat. 2014, 8. [Google Scholar] [CrossRef] [Green Version]
- Akyuz, E.; Villa, C.; Beker, M.; Elibol, B. Unraveling the Role of Inwardly Rectifying Potassium Channels in the Hippocampus of an Aβ(1–42)-Infused Rat Model of Alzheimer’s Disease. Biomedicines 2020, 8, 58. [Google Scholar] [CrossRef] [Green Version]
- Yoshiyama, Y.; Higuchi, M.; Zhang, B.; Huang, S.-M.; Iwata, N.; Saido, T.C.; Maeda, J.; Suhara, T.; Trojanowski, J.Q.; Lee, V.M.-Y. Synapse Loss and Microglial Activation Precede Tangles in a P301S Tauopathy Mouse Model. Neuron 2007, 53, 337–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jankowsky, J.L.; Fadale, D.J.; Anderson, J.; Xu, G.M.; Gonzales, V.; Jenkins, N.A.; Copeland, N.G.; Lee, M.; Younkin, L.H.; Wagner, S.L.; et al. Mutant presenilins specifically elevate the levels of the 42 residue β-amyloid peptide in vivo: Evidence for augmentation of a 42-specific γ secretase. Hum. Mol. Genet. 2003, 13, 159–170. [Google Scholar] [CrossRef] [Green Version]
- Luscher, C.; Ungless, M.A. The Mechanistic Classification of Addictive Drugs. PLoS Med. 2006, 3, e437. [Google Scholar] [CrossRef]
- Takigawa, T.; Alzheimer, C. G protein-activated inwardly rectifying K + (GIRK) currents in dendrites of rat neocortical pyramidal cells. J. Physiol. 1999, 517, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Fajardo-Serrano, A.; Wydeven, N.; Young, D.; Watanabe, M.; Shigemoto, R.; Martemyanov, K.A.; Wickman, K.; Luján, R. Association of Rgs7/Gβ5 complexes with Girk channels and GABAB receptors in hippocampal CA1 pyramidal neurons. Hippocampus 2013, 23, 1231–1245. [Google Scholar] [CrossRef] [Green Version]
- Martín-Belmonte, A.; Aguado, C.; Alfaro-Ruiz, R.; Albasanz, J.; Martín, M.; Moreno-Martínez, A.; Fukazawa, Y.; Luján, R. The Density of Group I mGlu5 Receptors Is Reduced along the Neuronal Surface of Hippocampal Cells in a Mouse Model of Alzheimer’s Disease. Int. J. Mol. Sci. 2021, 22, 5867. [Google Scholar] [CrossRef]
- Dolphin, A.C. G Protein Modulation of Voltage-Gated Calcium Channels. Pharmacol. Rev. 2003, 55, 607–627. [Google Scholar] [CrossRef]
- Ladera, C.; Godino, M.D.C.; Cabañero, M.J.; Torres, M.; Watanabe, M.; Luján, R.; Sánchez-Prieto, J. Pre-synaptic GABAB receptors inhibit glutamate release through GIRK channels in rat cerebral cortex. J. Neurochem. 2008, 107, 1506–1517. [Google Scholar] [CrossRef] [PubMed]
- Verret, L.; Mann, E.; Hang, G.B.; Barth, A.M.; Cobos, I.; Ho, K.; Devidze, N.; Masliah, E.; Kreitzer, A.C.; Mody, I.; et al. Inhibitory Interneuron Deficit Links Altered Network Activity and Cognitive Dysfunction in Alzheimer Model. Cell 2012, 149, 708–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.; McInnes, J.; Wierda, K.; Holt, M.; Herrmann, A.G.; Jackson, R.; Wang, Y.-C.; Swerts, J.; Beyens, J.; Miskiewicz, K.; et al. Tau association with synaptic vesicles causes presynaptic dysfunction. Nat. Commun. 2017, 8, 15295. [Google Scholar] [CrossRef]
- Dinamarca, M.C.; Raveh, A.; Schneider, A.; Fritzius, T.; Früh, S.; Rem, P.D.; Stawarski, M.; Lalanne, T.; Turecek, R.; Choo, M.; et al. Complex formation of APP with GABAB receptors links axonal trafficking to amyloidogenic processing. Nat. Commun. 2019, 10, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Rice, H.C.; de Malmazet, D.; Schreurs, A.; Frere, S.; Van Molle, I.; Volkov, A.N.; Creemers, E.; Vertkin, I.; Nys, J.; Ranaivoson, F.M.; et al. Secreted amyloid-β precursor protein functions as a GABABR1a ligand to modulate synaptic transmission. Science 2019, 363, eaao4827. [Google Scholar] [CrossRef]
- Ostrovskaya, O.; Xie, K.; Masuho, I.; Fajardo-Serrano, A.; Lujan, R.; Wickman, K.; A Martemyanov, K. RGS7/Gβ5/R7BP complex regulates synaptic plasticity and memory by modulating hippocampal GABABR-GIRK signaling. eLife 2014, 3, e02053. [Google Scholar] [CrossRef] [Green Version]
- Jankowsky, J.L.; Slunt, H.H.; Ratovitski, T.; Jenkins, N.A.; Copeland, N.G.; Borchelt, D.R. Co-expression of multiple transgenes in mouse CNS: A comparison of strategies. Biomol. Eng. 2001, 17, 157–165. [Google Scholar] [CrossRef]
- Garcia-Alloza, M.; Robbins, E.M.; Zhang-Nunes, S.X.; Purcell, S.M.; Betensky, R.A.; Raju, S.; Prada, C.; Greenberg, S.M.; Bacskai, B.J.; Frosch, M.P. Characterization of amyloid deposition in the APPswe/PS1dE9 mouse model of Alzheimer disease. Neurobiol. Dis. 2006, 24, 516–524. [Google Scholar] [CrossRef] [PubMed]
- Gimbel, D.A.; Nygaard, H.B.; Coffey, E.E.; Gunther, E.C.; Laurén, J.; Gimbel, Z.A.; Strittmatter, S. Memory Impairment in Transgenic Alzheimer Mice Requires Cellular Prion Protein. J. Neurosci. 2010, 30, 6367–6374. [Google Scholar] [CrossRef] [PubMed]
- Aguado, C.; Colón, J.; Ciruela, F.; Schlaudraff, F.; Cabañero, M.J.; Perry, C.; Watanabe, M.; Liss, B.; Wickman, K.; Luján, R. Cell type-specific subunit composition of G protein-gated potassium channels in the cerebellum. J. Neurochem. 2008, 105, 497–511. [Google Scholar] [CrossRef] [PubMed]
- Luján, R.; Nusser, Z.; Roberts, J.D.B.; Shigemoto, R.; Somogyi, P. Perisynaptic Location of Metabotropic Glutamate Receptors mGluR1 and mGluR5 on Dendrites and Dendritic Spines in the Rat Hippocampus. Eur. J. Neurosci. 1996, 8, 1488–1500. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alfaro-Ruiz, R.; Martín-Belmonte, A.; Aguado, C.; Hernández, F.; Moreno-Martínez, A.E.; Ávila, J.; Luján, R. The Expression and Localisation of G-Protein-Coupled Inwardly Rectifying Potassium (GIRK) Channels Is Differentially Altered in the Hippocampus of Two Mouse Models of Alzheimer’s Disease. Int. J. Mol. Sci. 2021, 22, 11106. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011106
Alfaro-Ruiz R, Martín-Belmonte A, Aguado C, Hernández F, Moreno-Martínez AE, Ávila J, Luján R. The Expression and Localisation of G-Protein-Coupled Inwardly Rectifying Potassium (GIRK) Channels Is Differentially Altered in the Hippocampus of Two Mouse Models of Alzheimer’s Disease. International Journal of Molecular Sciences. 2021; 22(20):11106. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011106
Chicago/Turabian StyleAlfaro-Ruiz, Rocío, Alejandro Martín-Belmonte, Carolina Aguado, Félix Hernández, Ana Esther Moreno-Martínez, Jesús Ávila, and Rafael Luján. 2021. "The Expression and Localisation of G-Protein-Coupled Inwardly Rectifying Potassium (GIRK) Channels Is Differentially Altered in the Hippocampus of Two Mouse Models of Alzheimer’s Disease" International Journal of Molecular Sciences 22, no. 20: 11106. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011106