
Long Non-Coding RNA ANRIL as a Potential Biomarker of Chemosensitivity and Clinical Outcomes in Osteosarcoma

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Results
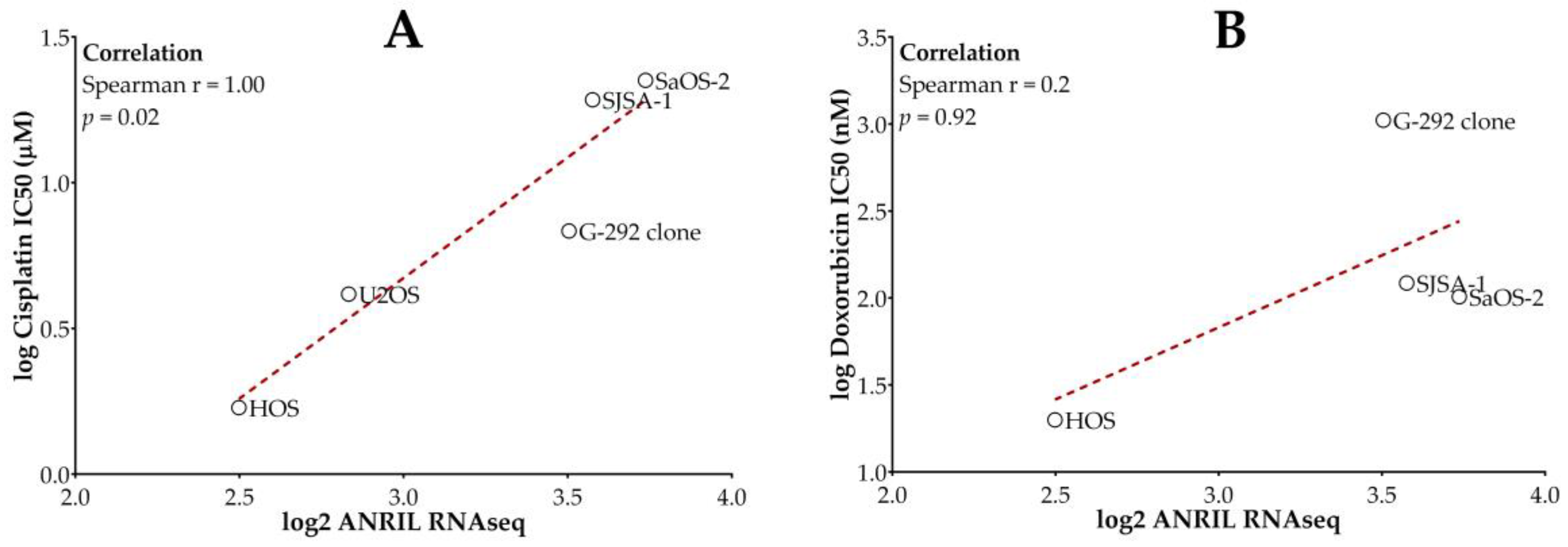
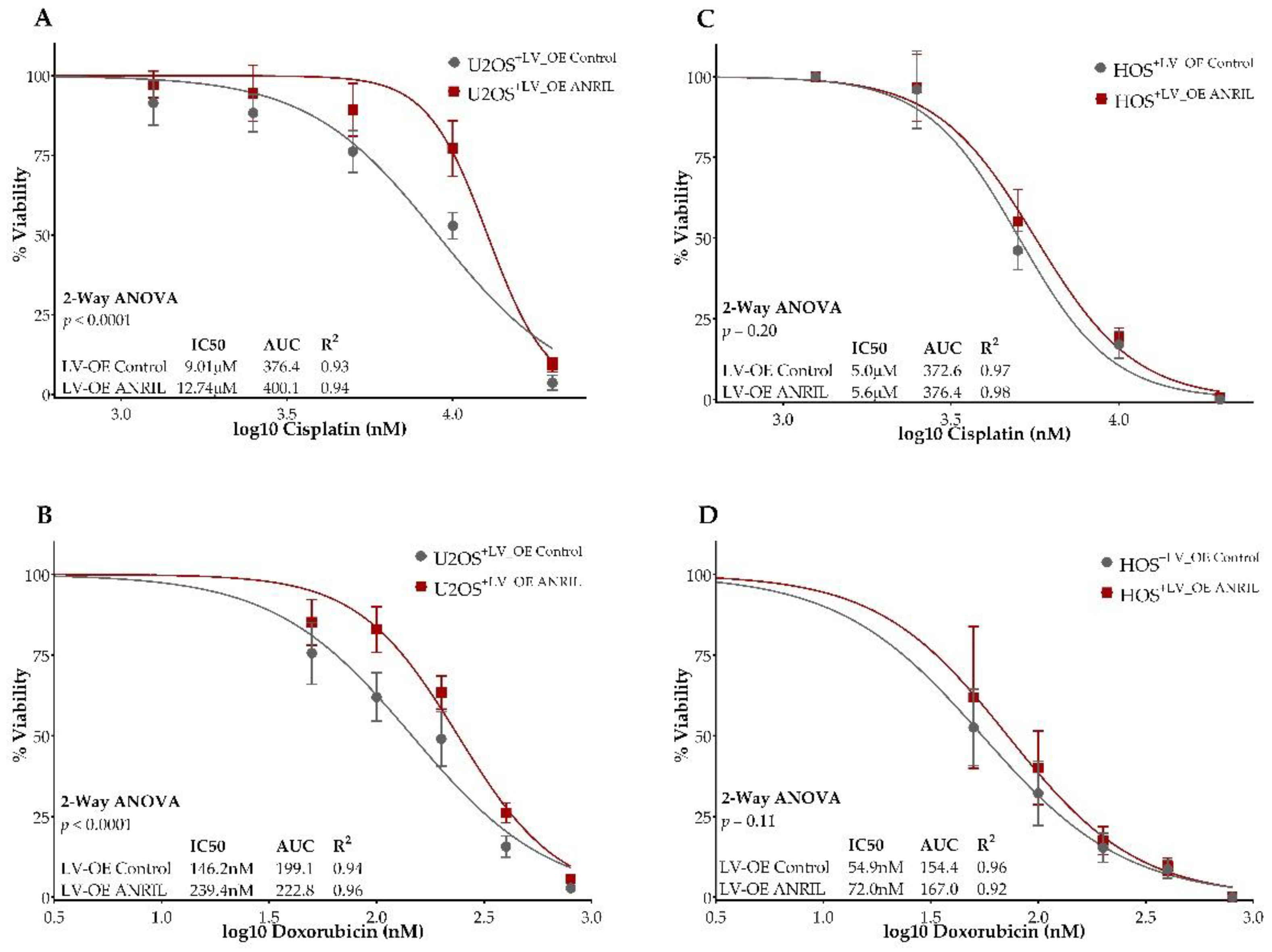
2.1. Higher ANRIL Expression Level Is Correlated with Cisplatin and Doxorubicin Resistance in Osteosarcoma Cell Lines
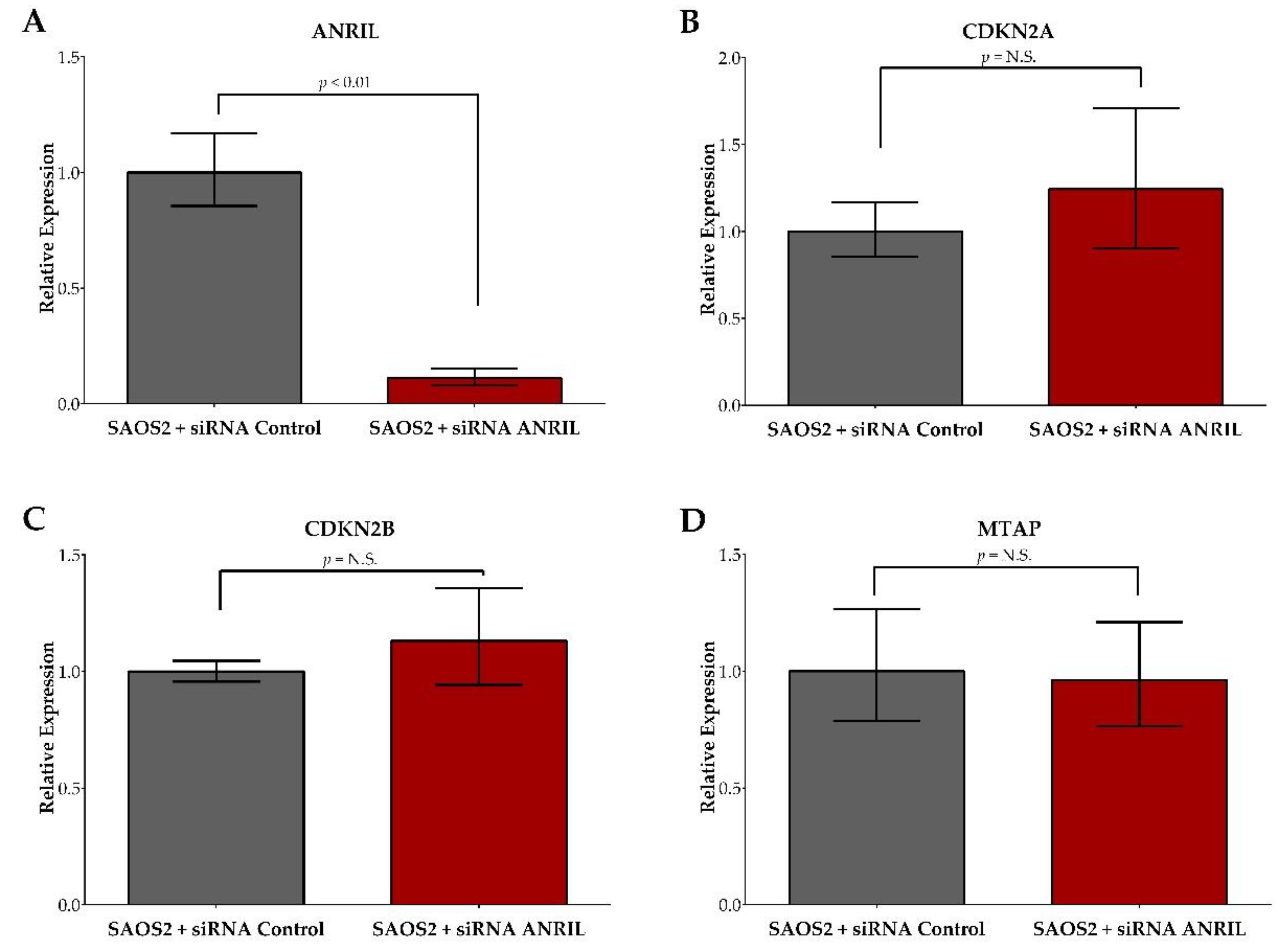
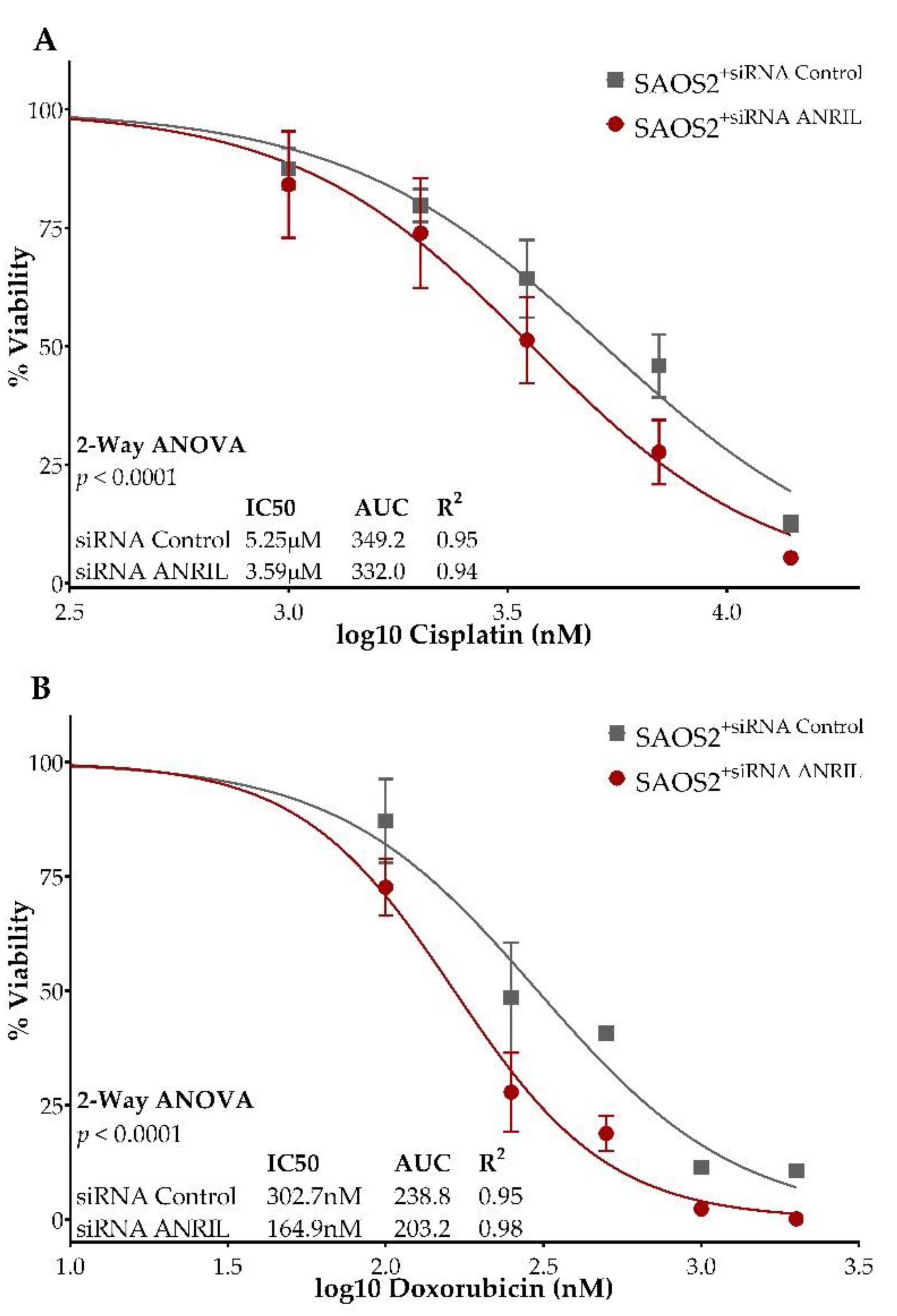
2.2. ANRIL Knockdown in SAOS2 and Impact on Cell Viability during Cisplatin and Doxorubicin Treatment
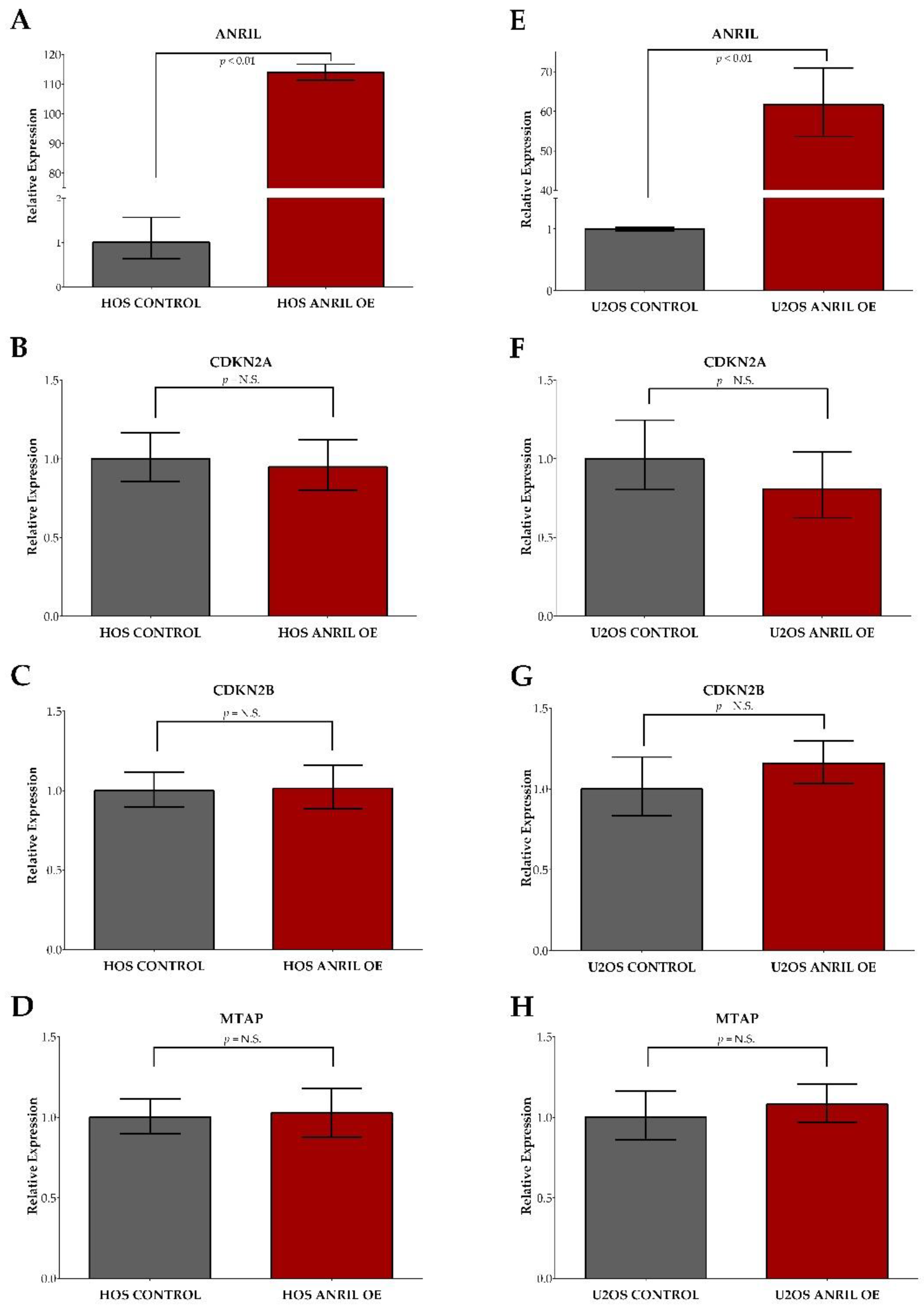
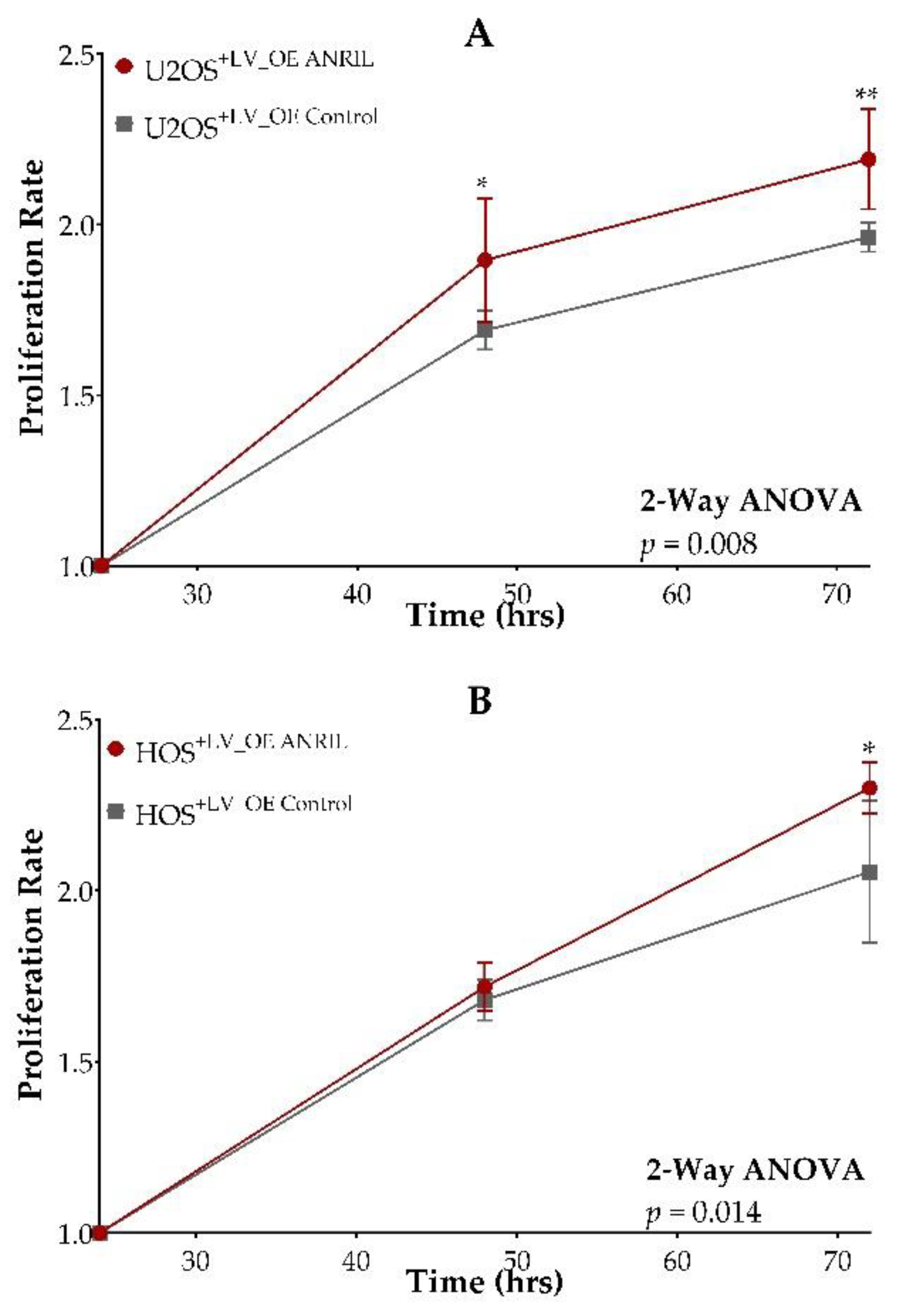
2.3. ANRIL Overexpression in HOS and U2OS and Impact on Cell Viability during Cisplatin and Doxorubicin Treatment
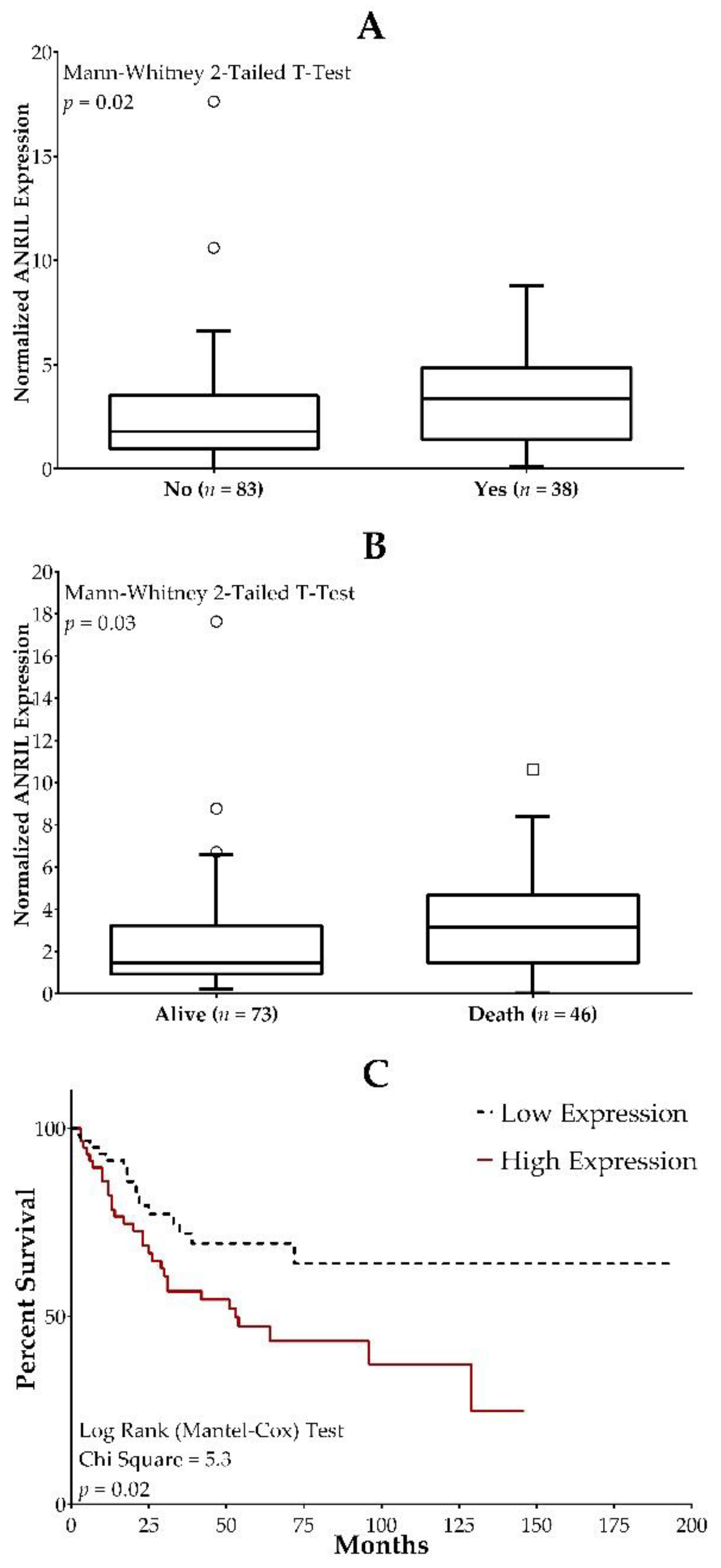
2.4. ANRIL Expression in Osteosarcoma Patients and Association with Clinical Outcome Measures
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Lines
4.3. lncRNA ANRIL Knockdown in SAOS2 Cells
4.4. lncRNA ANRIL Overexpression in HOS and U2OS Cells
4.5. lncRNA and Gene Expression Using Quantitative Real-Time PCR
4.6. Cell Viability and Drug Response Curves in Knockdown and Overexpression Models following Cisplatin and Doxorubicin Exposure
4.7. ANRIL Expression and Clinical Outcomes Assessment
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Friebele, J.C.; Peck, J.; Pan, X.; Abdel-Rasoul, M.; Mayerson, J.L. Osteosarcoma: A Meta-Analysis and Review of the Literature. Am. J. Orthop. 2015, 44, 547–553. [Google Scholar] [PubMed]
- Mirabello, L.; Troisi, R.J.; Savage, S.A. Osteosarcoma incidence and survival rates from 1973 to 2004: Data from the Surveillance, Epidemiology, and End Results Program. Cancer Interdisciplanary Int. J. Am. Cancer Soc. 2009, 115, 1531–1543. [Google Scholar] [CrossRef] [Green Version]
- Kager, L.; Tamamyan, G.; Bielack, S. Novel insights and therapeutic interventions for pediatric osteosarcoma. Future Oncol. 2017, 13, 357–368. [Google Scholar] [CrossRef]
- Sami, S.H.; Rafati, A.H.; Hodjat, P. Tissue necrosis after chemotherapy in osteosarcoma as the important prognostic factor. Saudi Med. J. 2008, 29, 1124–1129. [Google Scholar]
- Taran, S.J.; Taran, R.; Malipatil, N.B. Pediatric Osteosarcoma: An Updated Review. Indian J. Med. Paediatr. Oncol. 2017, 38, 33–43. [Google Scholar] [CrossRef] [Green Version]
- Durnali, A.G.; Turkoz, F.P.; Yukruk, F.A.; Tokluoglu, S.; Yazici, O.K.; Demirci, A.; Bal, O.; Buyukbas, S.G.; Esbah, O.; Oksuzoglu, B.; et al. Outcomes of Adolescent and Adult Patients with Lung Metastatic Osteosarcoma and Comparison of Synchronous and Metachronous Lung Metastatic Groups. PLoS ONE 2016, 11, e0152621. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Xie, L.; Guo, W. Neoadjuvant Chemotherapy Followed by Delayed Surgery: Is it Necessary for All Patients with Nonmetastatic High-Grade Pelvic Osteosarcoma? Clin. Orthop. Relat. Res. 2018, 476, 2177–2186. [Google Scholar] [CrossRef]
- Smeland, S.; Bielack, S.S.; Whelan, J.; Bernstein, M.; Hogendoorn, P.; Krailo, M.D.; Gorlick, R.; Janeway, K.A.; Ingleby, F.C.; Anninga, J.; et al. Survival and prognosis with osteosarcoma: Outcomes in more than 2000 patients in the EURAMOS-1 (European and American Osteosarcoma Study) cohort. Eur. J. Cancer. 2019, 109, 36–50. [Google Scholar] [CrossRef] [Green Version]
- Misaghi, A.; Goldin, A.; Awad, M.; Kulidjian, A.A. Osteosarcoma: A comprehensive review. SICOT J. 2018, 4, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirabello, L.; Zhu, B.; Koster, R.; Karlins, E.; Dean, M.; Yeager, M.; Gianferante, M.; Spector, L.; Morton, L.M.; Karyadi, D.; et al. Frequency of Pathogenic Germline Variants in Cancer-Susceptibility Genes in Patients with Osteosarcoma. JAMA Oncol. 2020, 6, 724–734. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ashana, A.O.; Moretti, V.M.; Lackman, R.D. The relation of tumour necrosis and survival in patients with osteosarcoma. Int. Orthop. 2011, 35, 1847–1853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanafy, E.; Al Jabri, A.; Gadelkarim, G.; Dasaq, A.; Nazim, F.; Al Pakrah, M. Tumor histopathological response to neoadjuvant chemotherapy in childhood solid malignancies: Is it still impressive? J. Investig. Med. 2018, 66, 289–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whelan, J.S.; Bielack, S.S.; Marina, N.; Smeland, S.; Jovic, G.; Hook, J.M.; Krailo, M.; Anninga, J.; Butterfass-Bahloul, T.; Böhling, T.; et al. EURAMOS-1, an international randomised study for osteosarcoma: Results from pre-randomisation treatment. Ann. Oncol. 2015, 26, 407–414. [Google Scholar] [CrossRef]
- Huang, Y.; Ling, A.; Pareek, S.; Huang, R.S. Oncogene or tumor suppressor? Long noncoding RNAs role in patient’s prognosis varies depending on disease type. Transl. Res. 2021, 230, 98–110. [Google Scholar] [CrossRef]
- Jarroux, J.; Morillon, A.; Pinskaya, M. History, Discovery, and Classification of lncRNAs. Adv. Exp. Med. Biol. 2017, 1008, 1–46. [Google Scholar]
- Kopp, F.; Mendell, J.T. Functional Classification and Experimental Dissection of Long Noncoding RNAs. Cell 2018, 172, 393–407. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.W.; Huang, K.; Yang, C.; Kang, C.S. Non-coding RNAs as regulators in epigenetics (Review). Oncol. Rep. 2017, 37, 3–9. [Google Scholar] [CrossRef] [Green Version]
- Lanzillotti, C.; De Mattei, M.; Mazziotta, C.; Taraballi, F.; Rotondo, J.C.; Tognon, M.; Martini, F. Long Non-coding RNAs and MicroRNAs Interplay in Osteogenic Differentiation of Mesenchymal Stem Cells. Front. Cell Dev. Biol. 2021, 9, 646032. [Google Scholar] [CrossRef]
- Moonmuang, S.; Chaiyawat, P.; Jantrapirom, S.; Pruksakorn, D.; Lo Piccolo, L. Circulating Long Non-Coding RNAs as Novel Potential Biomarkers for Osteogenic Sarcoma. Cancers 2021, 13, 4214. [Google Scholar] [CrossRef] [PubMed]
- Ghafouri-Fard, S.; Shirvani-Farsani, Z.; Hussen, B.M.; Taheri, M. The critical roles of lncRNAs in the development of osteosarcoma. Biomed. Pharmacother. 2021, 135, 111217. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Gao, L.; Ma, X.; Huang, J.-J.; Chen, J.; Zeng, L.; Jr., C.R.A.; Zou, C.; Chen, Z.-S. Long non-coding RNAs regulate drug resistance in cancer. Mol. Cancer 2020, 19, 54. [Google Scholar] [CrossRef]
- Chen, Q.N.; Wei, C.C.; Wang, Z.X.; Sun, M. Long non-coding RNAs in anti-cancer drug resistance. Oncotarget 2017, 8, 1925–1936. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Lü, M.; Zhou, Y.; Xu, L.; Jiang, Y.; Liu, Y.; Li, X.; Song, M. Role of Long Non-Coding RNAs in the Chemoresistance of Gastric Cancer: A Systematic Review. OncoTargets Ther. 2021, 14, 503–518. [Google Scholar] [CrossRef]
- Nath, A.; Geeleher, P.; Huang, R.S. Long non-coding RNA transcriptome of uncharacterized samples can be accurately imputed using protein-coding genes. Brief. Bioinform. 2020, 21, 637–648. [Google Scholar] [CrossRef]
- Nath, A.; Lau, E.Y.T.; Lee, A.M.; Geeleher, P.; Cho, W.C.S.; Huang, R.S. Discovering long noncoding RNA predictors of anticancer drug sensitivity beyond protein-coding genes. Proc. Natl. Acad. Sci. USA 2019, 116, 22020–22029. [Google Scholar] [CrossRef]
- Nath, A.; Huang, R.S. Emerging role of long non-coding RNAs in cancer precision medicine. Mol. Cell Oncol. 2020, 7, 1684130. [Google Scholar] [CrossRef]
- Pasmant, E.; Laurendeau, I.; Héron, D.; Vidaud, M.; Vidaud, D.; Bièche, I. Characterization of a germ-line deletion, including the entire INK4/ARF locus, in a melanoma-neural system tumor family: Identification of ANRIL, an antisense noncoding RNA whose expression coclusters with ARF. Cancer Res. 2007, 67, 3963–3969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.T.; Zhu, P.Q.; Luo, H.L.; Zhang, Y.; Hao, T.F.; Xia, G.F.; Zhu, Z.M.; Qiu, C. Long noncoding RNA ANRIL: A potential novel prognostic marker in cancer: A meta-analysis. Minerva Med. 2016, 107, 77–83. [Google Scholar] [PubMed]
- Li, Z.; Yu, X.; Shen, J. ANRIL: A pivotal tumor suppressor long non-coding RNA in human cancers. Tumour Biol. 2016, 37, 5657–5661. [Google Scholar] [CrossRef] [PubMed]
- Lou, N.; Liu, G.; Pan, Y. Long noncoding RNA ANRIL as a novel biomarker in human cancer. Future Oncol. 2020, 16, 2981–2995. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Bi, C.; Zhao, Q.; Ding, X.; Bian, C.; Wang, H.; Wang, T.; Liu, H. Knockdown long non-coding RNA ANRIL inhibits proliferation, migration and invasion of HepG2 cells by down-regulation of miR-191. BMC Cancer 2018, 18, 919. [Google Scholar] [CrossRef] [PubMed]
- Lan, W.-G.; Xu, D.-H.; Xu, C.; Ding, C.-L.; Ning, F.-L.; Zhou, Y.-L.; Ma, L.-B.; Liu, C.-M.; Chang-Ling, D. Silencing of long non-coding RNA ANRIL inhibits the development of multidrug resistance in gastric cancer cells. Oncol. Rep. 2016, 36, 263–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Cheng, N.; Luo, J. Downregulation of lncRNA ANRIL represses tumorigenicity and enhances cisplatin-induced cytotoxicity via regulating microRNA let-7a in nasopharyngeal carcinoma. J. Biochem Mol. Toxicol. 2017, 31, e21904. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Zhu, Y. Effect of lncRNA ANRIL knockdown on proliferation and cisplatin chemoresistance of osteosarcoma cells in vitro. Pathol. Res. Pract. 2019, 215, 931–938. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, G.; Cheng, Z.; Dai, L.; Jia, L.; Jing, X.; Wang, H.; Zhang, R.; Liu, M.; Jiang, T.; et al. Knockdown of lncRNA ANRIL inhibits the development of cisplatin resistance by upregulating miR-98 in lung cancer cells. Oncol. Rep. 2020, 44, 1025–1036. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Liao, Y.; Xiong, W.; Zhang, Y.; Zhou, Y.; Yang, Y. Hypoxia-induced lncRNA ANRIL promotes cisplatin resistance in retinoblastoma cells through regulating ABCG2 expression. Clin. Exp. Pharmacol Physiol. 2020, 47, 1049–1057. [Google Scholar] [CrossRef]
- Ferretti, V.A.; León, I.E. Long Non-coding RNAs in Cisplatin Resistance in Osteosarcoma. Curr. Treat. Options Oncol. 2021, 22, 41. [Google Scholar] [CrossRef]
- Yang, W.; Soares, J.; Greninger, P.; Edelman, E.; Lightfoot, H.; Forbes, S.; Sridhar, R.; Futreal, P.A.; Haber, D.; Stratton, M.; et al. Genomics of Drug Sensitivity in Cancer (GDSC): A resource for therapeutic biomarker discovery in cancer cells. Nucleic Acids Res. 2013, 41, D955–D961. [Google Scholar] [CrossRef] [Green Version]
- Jathar, S.; Kumar, V.; Srivastava, J.; Tripathi, V. Technological Developments in lncRNA Biology. Adv. Exp. Med. Biol. 2017, 1008, 283–323. [Google Scholar]
- Aguilo, F.; Zhou, M.M.; Walsh, M.J. Long noncoding RNA, polycomb, and the ghosts haunting INK4b-ARF-INK4a expression. Cancer Res. 2011, 71, 5365–5369. [Google Scholar] [CrossRef] [Green Version]
- Kong, Y.; Hsieh, C.H.; Alonso, L.C. A lncRNA at the CDKN2A/B Locus with Roles in Cancer and Metabolic Disease. Front. Endocrinol 2018, 9, 405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, G.; Mathur, R.; Hu, X.; Liu, Y.; Zhang, X.; Peng, G.; Lu, X. Long non-coding RNA ANRIL (CDKN2B-AS) is induced by the ATM-E2F1 signaling pathway. Cell Signal. 2013, 25, 1086–1095. [Google Scholar] [CrossRef] [Green Version]
- Guan, H.; Mei, Y.; Mi, Y.; Li, C.; Sun, X.; Zhao, X.; Liu, J.; Cao, W.; Li, Y.; Wang, Y. Downregulation of lncRNA ANRIL suppresses growth and metastasis in human osteosarcoma cells. OncoTargets Ther. 2018, 11, 4893–4899. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Liu, G.; Yuan, D.; Dai, J.; Cui, Y.; Tang, X. Long non-coding RNA ANRIL is associated with a poor prognosis of osteosarcoma and promotes tumorigenesis via PI3K/Akt pathway. J. Bone Oncol. 2018, 11, 51–55. [Google Scholar] [CrossRef]
- Qiu, J.J.; Lin, Y.Y.; Ding, J.X.; Feng, W.W.; Jin, H.Y.; Hua, K.Q. Long non-coding RNA ANRIL predicts poor prognosis and promotes invasion/metastasis in serous ovarian cancer. Int. J. Oncol. 2015, 46, 2497–2505. [Google Scholar] [CrossRef] [Green Version]
- Huang, M.-D.; Chen, W.-M.; Qi, F.-Z.; Xia, R.; Sun, M.; Xu, T.-P.; Yin, L.; Zhang, E.-B.; De, W.; Shu, Y.-Q. Long non-coding RNA ANRIL is upregulated in hepatocellular carcinoma and regulates cell proliferation by epigenetic silencing of KLF2. J. Hematol. Oncol. 2015, 8, 57. [Google Scholar] [CrossRef] [Green Version]
- Cheng, S.; Huang, T.; Li, P.; Zhang, W.; Wang, Z.; Chen, Y. Long non-coding RNA ANRIL promotes the proliferation, migration and invasion of human osteosarcoma cells. Exp. Ther. Med. 2017, 14, 5121–5125. [Google Scholar] [CrossRef]
- Naemura, M.; Tsunoda, T.; Inoue, Y.; Okamoto, H.; Shirasawa, S.; Kotake, Y. ANRIL regulates the proliferation of human colorectal cancer cells in both two- and three-dimensional culture. Mol. Cell. Biochem. 2016, 412, 141–146. [Google Scholar] [CrossRef]
- Zou, Z.W.; Ma, C.; Medoro, L.; Chen, L.; Wang, B.; Gupta, R.; Liu, T.; Yang, X.Z.; Chen, T.T.; Wang, R.Z.; et al. LncRNA ANRIL is up-regulated in nasopharyngeal carcinoma and promotes the cancer progression via increasing proliferation, reprograming cell glucose metabolism and inducing side-population stem-like cancer cells. Oncotarget 2016, 7, 61741–61754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Chen, Q.; Zhang, X.; Yang, J.; Lin, K.; Ji, C.; Xu, A.; Yang, L.; Miao, L. Long noncoding RNA ANRIL promotes the malignant progression of cholangiocarcinoma by epigenetically repressing ERRFI1 expression. Cancer Sci. 2020, 111, 2297–2309. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.Y.; Liu, Y.J.; He, Y.; Chen, P. Down-regulation of long non-coding RNA ANRIL inhibits the proliferation, migration and invasion of cervical cancer cells. Cancer Biomark. 2018, 23, 243–253. [Google Scholar] [CrossRef]
- Miao, J.T.; Gao, J.H.; Chen, Y.Q.; Chen, H.; Meng, H.Y.; Lou, G. LncRNA ANRIL affects the sensitivity of ovarian cancer to cisplatin via regulation of let-7a/HMGA2 axis. Biosci. Rep. 2019, 39, BSR20182101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Ding, L.; Li, Y.; Ren, J.; Shi, G.; Wang, Y.; Zhao, S.; Ni, Y.; Hou, Y. Midkine derived from cancer-associated fibroblasts promotes cisplatin-resistance via up-regulation of the expression of lncRNA ANRIL in tumour cells. Sci. Rep. 2017, 7, 16231. [Google Scholar] [CrossRef] [Green Version]
- Xu, R.; Mao, Y.; Chen, K.; He, W.; Shi, W.; Han, Y. The long noncoding RNA ANRIL acts as an oncogene and contributes to paclitaxel resistance of lung adenocarcinoma A549 cells. Oncotarget. 2017, 8, 39177–39184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotake, Y.; Nakagawa, T.; Kitagawa, K.; Suzuki, S.; Liu, N.; Xiong, Y. Long non-coding RNA ANRIL is required for the PRC2 recruitment to and silencing of p15(INK4B) tumor suppressor gene. Oncogene 2011, 30, 1956–1962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yap, K.L.; Li, S.; Muñoz-Cabello, A.M.; Raguz, S.; Zeng, L.; Mujtaba, S.; Gil, J.; Walsh, M.J.; Zhou, M.-M. Molecular interplay of the noncoding RNA ANRIL and methylated histone H3 lysine 27 by polycomb CBX7 in transcriptional silencing of INK4a. Mol. Cell. 2010, 38, 662–674. [Google Scholar] [CrossRef] [Green Version]
- Burd, C.E.; Jeck, W.R.; Liu, Y.; Sanoff, H.K.; Wang, Z.; Sharpless, N.E. Expression of linear and novel circular forms of an INK4/ARF-associated non-coding RNA correlates with atherosclerosis risk. PLoS Genet. 2010, 6, e1001233. [Google Scholar] [CrossRef] [PubMed]
- Holdt, L.M.; Beutner, F.; Scholz, M.; Gielen, S.; Gábel, G.; Bergert, H.; Schuler, G.; Thiery, J.; Teupser, D. ANRIL expression is associated with atherosclerosis risk at chromosome 9p21. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 620–627. [Google Scholar] [CrossRef] [Green Version]
- Cunnington, M.S.; Santibanez Koref, M.; Mayosi, B.M.; Burn, J.; Keavney, B. Chromosome 9p21 SNPs Associated with Multiple Disease Phenotypes Correlate with ANRIL Expression. PLoS Genet. 2010, 6, e1000899. [Google Scholar] [CrossRef] [Green Version]
- Jarinova, O.; Stewart, A.F.; Roberts, R.; Wells, G.; Lau, P.; Naing, T.; Buerki, C.; McLean, B.W.; Cook, R.C.; Parker, J.S.; et al. Functional analysis of the chromosome 9p21.3 coronary artery disease risk locus. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 1671–1677. [Google Scholar] [CrossRef] [Green Version]
- Motterle, A.; Pu, X.; Wood, H.; Xiao, Q.; Gor, S.; Ng, F.L.; Chan, K.; Cross, F.; Shohreh, B.; Poston, R.N.; et al. Functional analyses of coronary artery disease associated variation on chromosome 9p21 in vascular smooth muscle cells. Hum. Mol. Genet. 2012, 21, 4021–4029. [Google Scholar] [CrossRef] [Green Version]
- Zhi, L.; Liu, D.; Wu, S.G.; Li, T.; Zhao, G.; Zhao, B.; Li, M. Association of common variants in MTAP with susceptibility and overall survival of osteosarcoma: A two-stage population-based study in HAN Chinese. J. Cancer 2016, 7, 2179–2186. [Google Scholar] [CrossRef] [Green Version]
- García-Castellano, J.M.; Villanueva, A.; Healey, J.H.; Sowers, R.; Cordon-Cardo, C.; Huvos, A.; Bertino, J.R.; Meyers, P.; Gorlick, R. Methylthioadenosine phosphorylase gene deletions are common in osteosarcoma. Clin. Cancer Res. 2002, 8, 782–787. [Google Scholar] [PubMed]
- Lubin, M.; Lubin, A. Selective killing of tumors deficient in methylthioadenosine phosphorylase: A novel strategy. PLoS ONE 2009, 4, e5735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, F.; Pfeiffer, R.M.; Bhattacharjee, S.; Han, S.S.; Taylor, P.R.; Berndt, S.; Yang, H.; Sigurdson, A.J.; Toro, J.; Mirabello, L.; et al. Common genetic variants in the 9p21 region and their associations with multiple tumours. Br. J. Cancer 2013, 108, 1378–1386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Zheng, Z.P.; Li, H.; Zhang, H.Q.; Ma, F.Q. ANRIL is associated with the survival rate of patients with colorectal cancer, and affects cell migration and invasion In Vitro. Mol. Med. Rep. 2016, 14, 1714–1720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Sun, G.; Zhang, H.; Tian, J.; Li, Y. Long non-coding RNA ANRIL indicates a poor prognosis of cervical cancer and promotes carcinogenesis via PI3K/Akt pathways. Biomed. Pharmacother. 2017, 85, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Gu, Z.T.; Chen, W.H.; Cao, K.J. Increased expression of the long non-coding RNA ANRIL promotes lung cancer cell metastasis and correlates with poor prognosis. Diagn. Pathol. 2015, 10, 14. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Shen, E.-D.; Liao, M.-M.; Hu, Y.-B.; Wu, K.; Yang, P.; Zhou, L.; Chen, W.-D. Expression and mechanisms of long non-coding RNA genes MEG3 and ANRIL in gallbladder cancer. Tumour Biol. 2016, 37, 9875–9886. [Google Scholar] [CrossRef]
- Yin, Y.; Yang, W.; Zhang, L.; Liu, K.; Luo, Z. Long non-coding RNA ANRIL and its target microRNAs (microRNA-34a, microRNA-125a and microRNA-186) relate to risk stratification and prognosis in multiple myeloma. Hematology 2021, 26, 160–169. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Kim, H.Y. Statistical notes for clinical researchers: Two-way analysis of variance (ANOVA)-exploring possible interaction between factors. Restor. Dent. Endod. 2014, 39, 143–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McHugh, M.L. The chi-square test of independence. Biochem Med. 2013, 23, 143–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplan, E.L.; Meier, P. Nonparametric Estimation from Incomplete Observations. J. Am. Stat. Assoc. 1958, 53, 457–481. [Google Scholar] [CrossRef]
- Bland, J.M.; Altman, D.G. The logrank test. BMJ 2004, 328, 1073. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | PCR Primer Sequence |
|---|---|
| ANRIL | F: 5′-CTCTCATCTGATCTCCGTCCT-3′ |
| R: 5′-TCACATCCAAGACAGCAAGT-3′ | |
| CDKN2A | F: 5′-GTGCCACATTCGCTAAGTG-3′ |
| R: 5′-ACCCTGTCCCTCAAATCCT-3′ | |
| CDKN2B | F: 5′-ATGCGTTCACTCCAATGTCT-3′ |
| R: 5′-CTCCACTTTGTCCTCAGTCTTC-3′ | |
| MTAP | F: 5′–GCAGCCATGCTACTTTAATGTC-3′ |
| R: 5′–GCTTACTGCTCACTACCATACC–3′ | |
| GAPDH | F: 5′-GAACATCATCCCTGCCTCTAC-3′ |
| R: 5′-CCTGCTTCACCACCTTCTT-3′ | |
| ACTB (β-Actin) | F: 5′-GTGGCCGAGGACTTTGATT-3′ |
| R: 5′-TTTAGGATGGCAAGGGACTTC-3′ |
| TARGET Osteosarcoma Patient Dataset | |
|---|---|
| Clinical Characteristic | n (%) or Mean (Range) |
| Sex | n= 86 |
| Males | 50 (58%) |
| Females | 36 (42%) |
| Age at Diagnosis, Mean (Range) | 15 (8–32) |
| <14 years of age | 35 (41%) |
| ≥14 years of age | 51 (59%) |
| Metastases at Diagnosis | n= 86 |
| Yes | 21 (24%) |
| No | 65 (76%) |
| Overall Survival in Months | 126 (0–487) |
| <36 months | 73 (86%) |
| ≥36 months | 12 (14%) |
| % Necrosis | n= 43 |
| <91% | 19 (44%) |
| ≥91% | 24 (56%) |
| Death | n= 86 |
| Yes | 28 (33%) |
| No | 58 (67%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, A.M.; Ferdjallah, A.; Moore, E.; Kim, D.C.; Nath, A.; Greengard, E.; Huang, R.S. Long Non-Coding RNA ANRIL as a Potential Biomarker of Chemosensitivity and Clinical Outcomes in Osteosarcoma. Int. J. Mol. Sci. 2021, 22, 11168. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011168
Lee AM, Ferdjallah A, Moore E, Kim DC, Nath A, Greengard E, Huang RS. Long Non-Coding RNA ANRIL as a Potential Biomarker of Chemosensitivity and Clinical Outcomes in Osteosarcoma. International Journal of Molecular Sciences. 2021; 22(20):11168. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011168
Chicago/Turabian StyleLee, Adam M., Asmaa Ferdjallah, Elise Moore, Daniel C. Kim, Aritro Nath, Emily Greengard, and R. Stephanie Huang. 2021. "Long Non-Coding RNA ANRIL as a Potential Biomarker of Chemosensitivity and Clinical Outcomes in Osteosarcoma" International Journal of Molecular Sciences 22, no. 20: 11168. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011168