
LncRNAs in the Regulation of Genes and Signaling Pathways through miRNA-Mediated and Other Mechanisms in Clear Cell Renal Cell Carcinoma

 , , , and
, , , and
Abstract
:1. Introduction
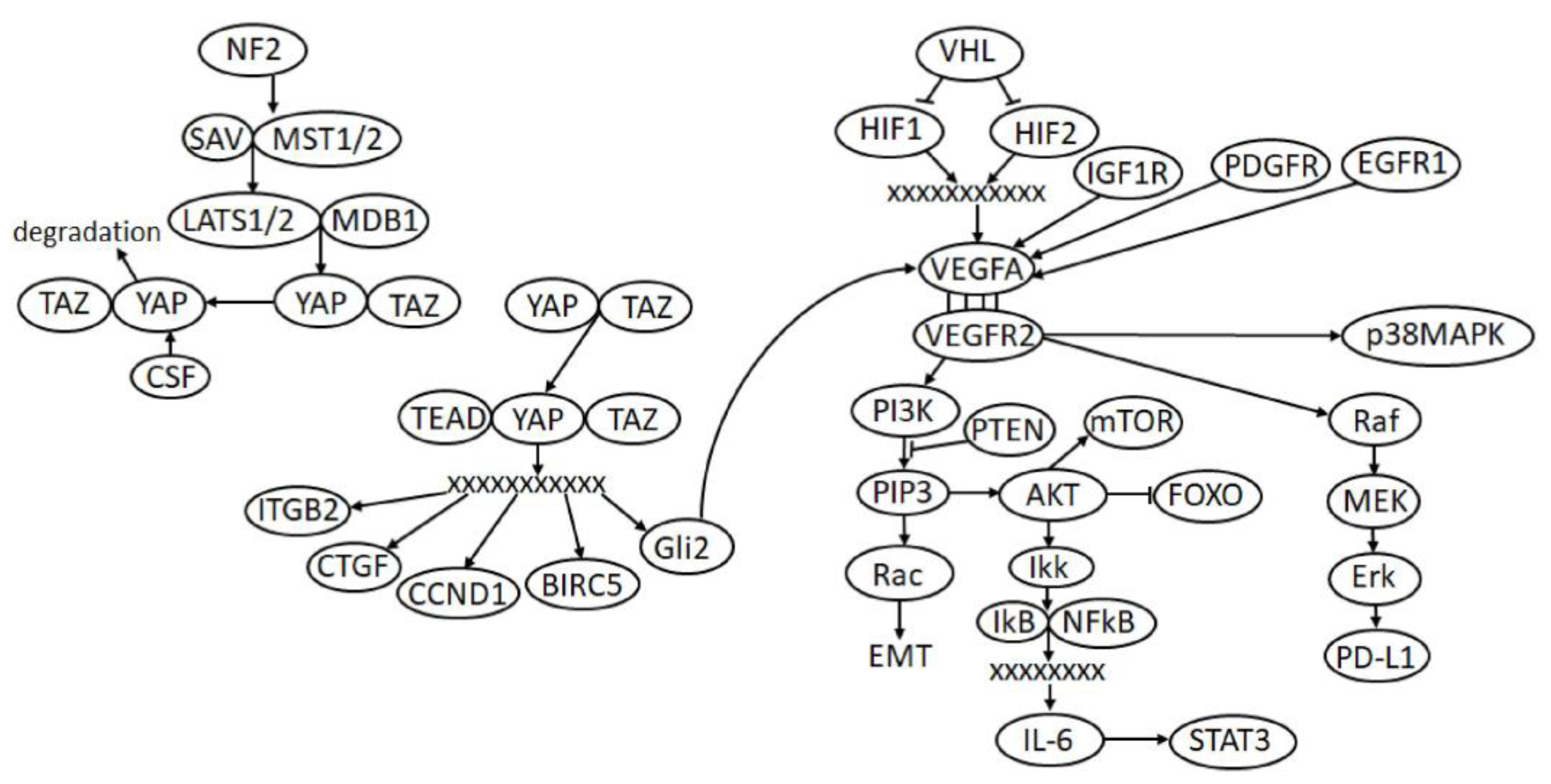
2. Signaling Pathways and Processes Significant in ccRCC
2.1. VHL/HIF/VEGF Pathway
2.2. PI3K/AKT/mTOR Pathway
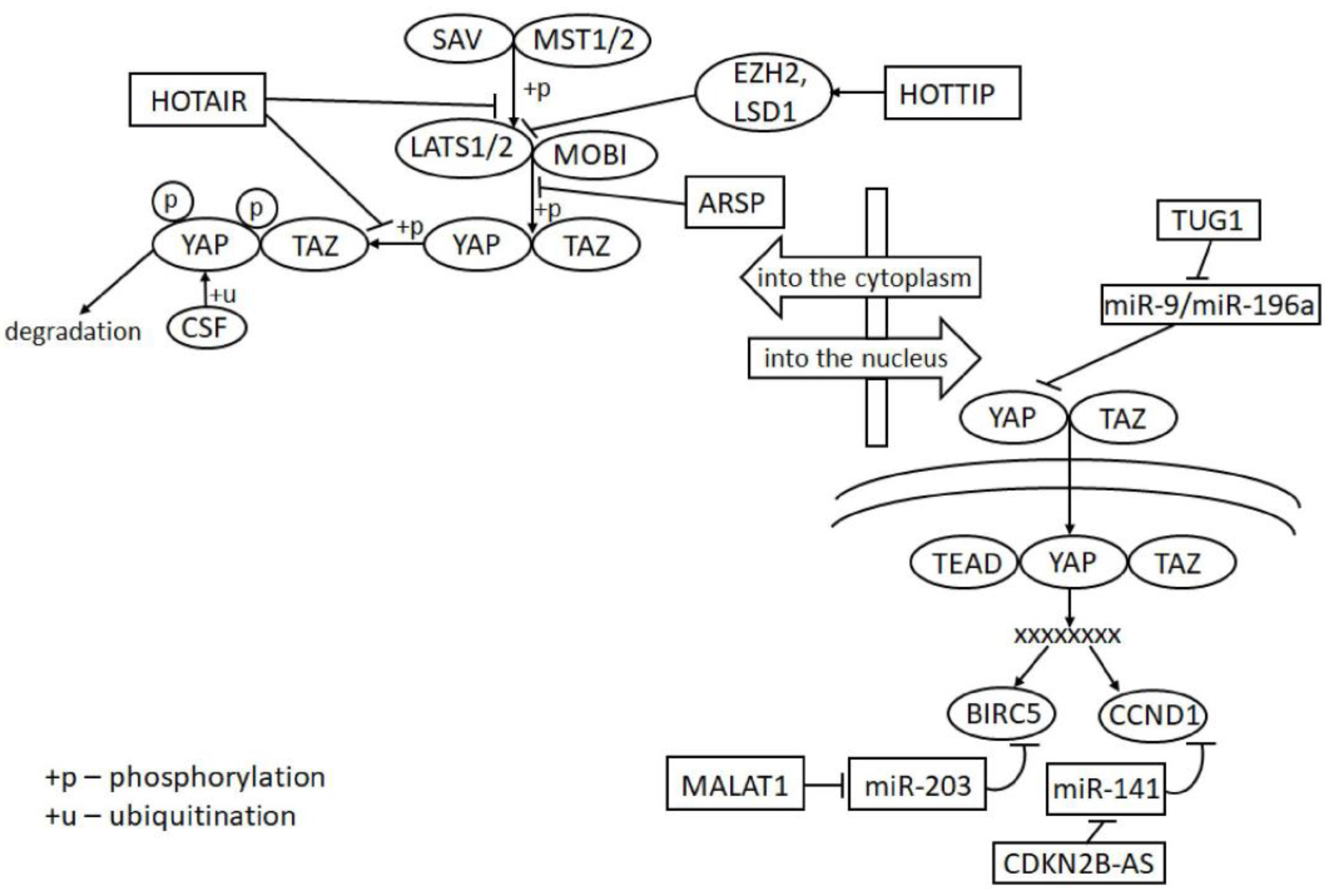
2.3. Hippo Signaling
2.4. Epithelial-Mesenchymal Transition (EMT)
2.5. Suppression of the Immune Response
3. Regulatory Function of LncRNAs via the ceRNA Mechanism in ccRCC
3.1. The LncRNAs Acting According to the ceRNA Model
3.2. Oncogenic LncRNA CDKN2B-AS1 in the ceRNA Model
3.3. Oncogenic LncRNA HCP5 in the ceRNA Model
3.4. Oncogenic LncRNA LINC00973 as an Immune Suppressor in the ceRNA Model
3.5. Oncogenic LncRNA LINC01094 in the ceRNA Model
3.6. Oncogenic LncRNA LOXL1-AS1 in the ceRNA Model
3.7. Oncogenic LncRNA PCGEM1 in the ceRNA Model
3.8. Oncogenic LncRNA SNHG5 in the ceRNA Model
3.9. Oncogenic LncRNA SNHG12 in the ceRNA Model
3.10. Oncogenic LncRNA SNHG16 in the ceRNA Model
3.11. Oncogenic LncRNA UCA1 in the ceRNA Model
3.12. Novel Suppressive LncRNA PENG in the ceRNA Model
3.13. Multiple Functions of LncRNAs in the ceRNA Model
3.14. Dual Features of Some LncRNAs and Their Targets or the Need to Verify Interactions for Them
4. Alternative Mechanisms of Regulation of the Expression of Protein-Coding Genes with the Participation of LncRNAs in ccRCC
4.1. SNHG12/SP1/CDCA3 in Transcription Regulation
4.2. MAGI2-AS3/HEY1/ACY1 in Transcription Regulation
4.3. HOTTIP/EZH2, LSD1/LATS2 in Chromatin Reorganization through Binding to the Gene Promoter Region
4.4. EGFR-AS1/EGFR in Binding to mRNA
4.5. MALAT1/Livin in Binding to Protein
4.6. Alternative Mechanisms of Action of LncRNAs
5. Effect of LncRNAs on Key Pathways and Processes in ccRCC
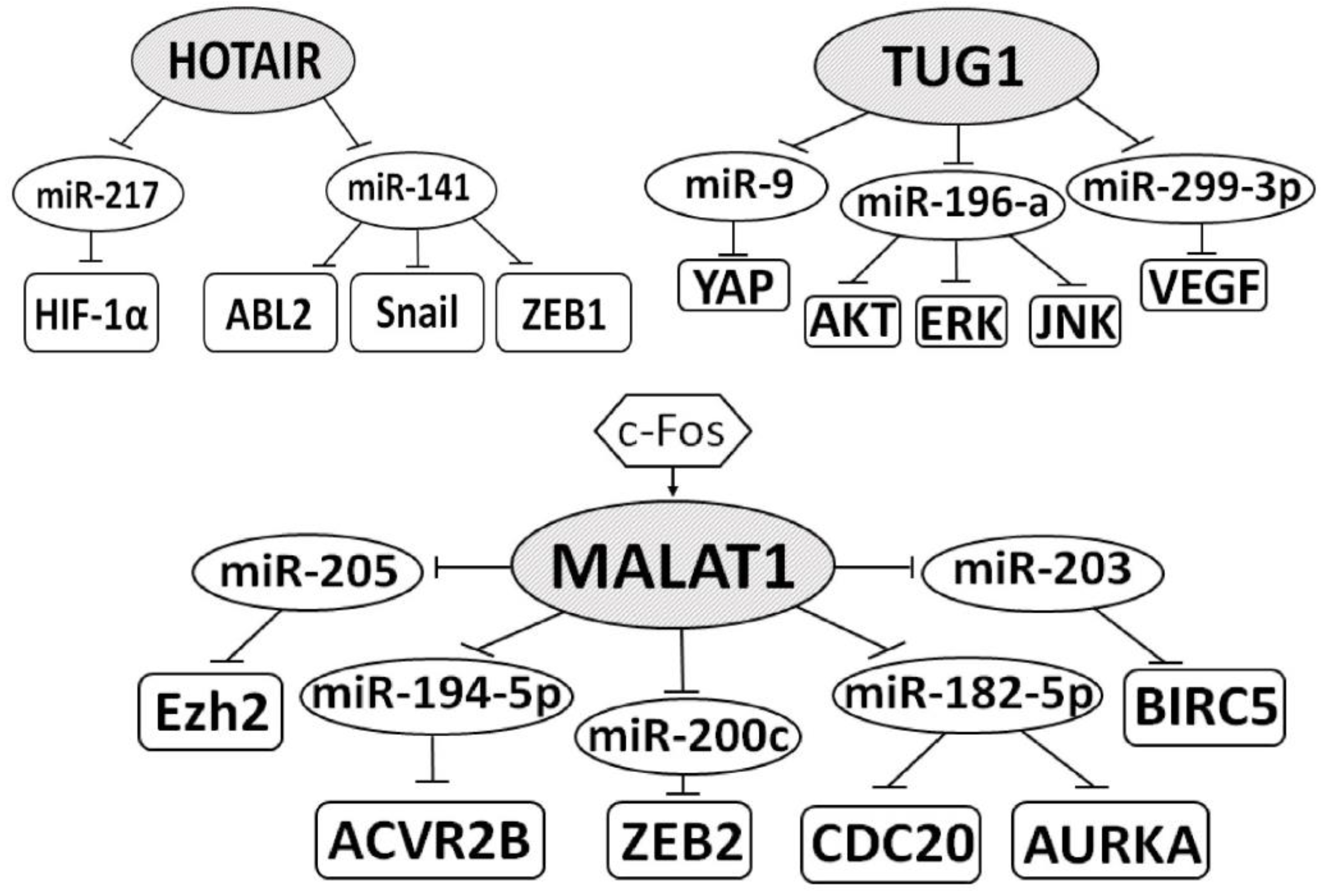
5.1. LncRNAs in VEGF Signaling
5.2. LncRNAs in PI3K/AKT/mTOR Signaling
5.3. LncRNAs in Hippo Signaling
5.4. LncRNAs in EMT Processes
5.5. LncRNAs in the Suppression of the Immune Response
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Linehan, W.M.; Ricketts, C.J. The Cancer Genome Atlas of renal cell carcinoma: Findings and clinical implications. Nat. Rev. Urol. 2019, 16, 539–552. [Google Scholar] [CrossRef]
- Tannir, N.M.; Pal, S.K.; Atkins, M.B. Second-Line Treatment Landscape for Renal Cell Carcinoma: A Comprehensive Review. Oncologist 2018, 23, 540–555. [Google Scholar] [CrossRef] [Green Version]
- Mori, K.; Mostafaei, H.; Miura, N.; Karakiewicz, P.I.; Luzzago, S.; Schmidinger, M.; Bruchbacher, A.; Pradere, B.; Egawa, S.; Shariat, S.F. Systemic therapy for metastatic renal cell carcinoma in the first-line setting: A systematic review and network meta-analysis. Cancer Immunol. Immunother. 2020, 70, 265–273. [Google Scholar] [CrossRef]
- Jenkins, R.W.; Barbie, D.A.; Flaherty, K.T. Mechanisms of resistance to immune checkpoint inhibitors. Br. J. Cancer 2018, 118, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Rosenzweig, S.A. Acquired Resistance to Drugs Targeting Tyrosine Kinases. Adv. Cancer Res. 2018, 138, 71–98. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Cheung, B.B.; Atmadibrata, B.; Marshall, G.M.; Dinger, M.; Liu, P.Y.; Liu, T. The regulatory role of long noncoding RNAs in cancer. Cancer Lett. 2017, 391, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Luo, G.; Chen, Q. Long noncoding RNAs as tumorigenic factors and therapeutic targets for renal cell carcinoma. Cancer Cell Int. 2021, 21, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tay, Y.; Rinn, J.; Pandolfi, P.P. The multilayered complexity of ceRNA crosstalk and competition. Nature 2014, 505, 344–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, R.; Wang, L.; Li, J.; Ma, L.; Wang, F.; Wang, Y. Integrated Analysis of a Competing Endogenous RNA Network Reveals a Prognostic Signature in Kidney Renal Papillary Cell Carcinoma. Front. Cell Dev. Biol. 2020, 8, 612924. [Google Scholar] [CrossRef]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA Hypothesis: The Rosetta Stone of a Hidden RNA Language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Wu, D.; Gao, L.; Liu, X.; Jin, Y.; Wang, D.; Wang, T.; Li, X. Competing endogenous RNA networks in human cancer: Hypothesis, validation, and perspectives. Oncotarget 2016, 7, 13479–13490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirata, H.; Hinoda, Y.; Shahryari, V.; Deng, G.; Nakajima, K.; Tabatabai, Z.L.; Ishii, N.; Dahiya, R. Long Noncoding RNA MALAT1 Promotes Aggressive Renal Cell Carcinoma through Ezh2 and Interacts with miR-205. Cancer Res. 2015, 75, 1322–1331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choueiri, T.K.; Jr, W.G.K. Targeting the HIF2-VEGF axis in renal cell carcinoma. Nat. Med. 2020, 26, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Dengler, V.L.; Galbraith, M.; Espinosa, J.M. Transcriptional regulation by hypoxia inducible factors. Crit. Rev. Biochem. Mol. Biol. 2013, 49, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uniacke, J.; Holterman, C.; Lachance, G.; Franovic, A.; Jacob, M.D.; Fabian, M.R.; Payette, J.; Holcik, M.; Pause, A.; Lee, S. An oxygen-regulated switch in the protein synthesis machinery. Nature 2012, 486, 126–129. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Roche, O.; Yan, M.S.; Finak, G.; Evans, A.J.; Metcalf, J.L.; Hast, E.B.; Hanna, S.C.; Wondergem, B.; Furge, K.A.; et al. Regulation of endocytosis via the oxygen-sensing pathway. Nat. Med. 2009, 15, 319–324. [Google Scholar] [CrossRef]
- Masoud, G.N.; Li, W. HIF-1α pathway: Role, regulation and intervention for cancer therapy. Acta Pharm. Sin. B 2015, 5, 378–389. [Google Scholar] [CrossRef] [Green Version]
- Raval, R.R.; Lau, K.W.; Tran, M.G.B.; Sowter, H.M.; Mandriota, S.J.; Li, J.-L.; Pugh, C.; Maxwell, P.; Harris, A.L.; Ratcliffe, P.J. Contrasting Properties of Hypoxia-Inducible Factor 1 (HIF-1) and HIF-2 in von Hippel-Lindau-Associated Renal Cell Carcinoma. Mol. Cell. Biol. 2005, 25, 5675–5686. [Google Scholar] [CrossRef] [Green Version]
- Roskoski, R. Vascular endothelial growth factor (VEGF) and VEGF receptor inhibitors in the treatment of renal cell carcinomas. Pharmacol. Res. 2017, 120, 116–132. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; German, P.; Bai, S.; Barnes, S.; Guo, W.; Qi, X.; Lou, H.; Liang, J.; Jonasch, E.; Mills, G.B.; et al. The PI3K/AKT Pathway and Renal Cell Carcinoma. J. Genet. Genom. 2015, 42, 343–353. [Google Scholar] [CrossRef] [Green Version]
- Miricescu, D.; Balan, D.G.; Tulin, A.; Stiru, O.; Vacaroiu, I.A.; Mihai, D.A.; Popa, C.C.; Papacocea, R.I.; Enyedi, M.; Sorin, N.A.; et al. PI3K/AKT/mTOR signalling pathway involvement in renal cell carcinoma pathogenesis (Review). Exp. Ther. Med. 2021, 21, 1–7. [Google Scholar] [CrossRef]
- Dizman, N.; Philip, E.J.; Pal, S.K. Genomic profiling in renal cell carcinoma. Nat. Rev. Nephrol. 2020, 16, 1–17. [Google Scholar] [CrossRef]
- Calses, P.C.; Crawford, J.J.; Lill, J.R.; Dey, A. Hippo Pathway in Cancer: Aberrant Regulation and Therapeutic Opportunities. Trends Cancer 2019, 5, 297–307. [Google Scholar] [CrossRef] [Green Version]
- Schütte, U.; Bisht, S.; Heukamp, L.; Kebschull, M.; Florin, A.; Haarmann, J.; Hoffmann, P.; Bendas, G.; Buettner, R.; Brossart, P.; et al. Hippo Signaling Mediates Proliferation, Invasiveness, and Metastatic Potential of Clear Cell Renal Cell Carcinoma. Transl. Oncol. 2014, 7, 309–321. [Google Scholar] [CrossRef] [Green Version]
- Mittal, R.; Jaiswal, P.; Goel, A. Survivin: A molecular biomarker in cancer. Indian J. Med. Res. 2015, 141, 389–397. [Google Scholar] [CrossRef]
- Rybarczyk, A.; Klacz, J.; Wronska, A.; Matuszewski, M.; Kmiec, Z.; Wierzbicki, P. Overexpression of the YAP1 oncogene in clear cell renal cell carcinoma is associated with poor outcome. Oncol. Rep. 2017, 38, 427–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.-H.; Ding, C.-K.C.; Sun, T.; Rupprecht, G.; Lin, C.-C.; Hsu, D.; Chi, J.-T. The Hippo Pathway Effector TAZ Regulates Ferroptosis in Renal Cell Carcinoma. Cell Rep. 2019, 28, 2501–2508.e4. [Google Scholar] [CrossRef] [PubMed]
- Piva, F.; Giulietti, M.; Santoni, M.; Occhipinti, G.; Scarpelli, M.; López-Beltrán, A.; Cheng, L.; Principato, G.; Montironi, R. Epithelial to Mesenchymal Transition in Renal Cell Carcinoma: Implications for Cancer Therapy. Mol. Diagn. Ther. 2016, 20, 111–117. [Google Scholar] [CrossRef]
- Yuan, H.; Meng, X.; Guo, W.; Cai, P.; Li, W.; Li, Q.; Wang, W.; Sun, Y.; Xu, Q.; Gu, Y. Transmembrane-Bound IL-15–Promoted Epithelial-Mesenchymal Transition in Renal Cancer Cells Requires the Src-Dependent Akt/GSK-3β/β-Catenin Pathway. Neoplasia 2015, 17, 410–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, B.; Shim, J.S. Targeting Epithelial–Mesenchymal Transition (EMT) to Overcome Drug Resistance in Cancer. Molecules 2016, 21, 965. [Google Scholar] [CrossRef] [Green Version]
- Landolt, L.; Eikrem, Ø; Strauss, P.; Scherer, A.; Lovett, D.H.; Beisland, C.; Finne, K.; Osman, T.; Ibrahim, M.M.; Gausdal, G.; et al. Clear Cell Renal Cell Carcinoma is linked to Epithelial-to-Mesenchymal Transition and to Fibrosis. Physiol. Rep. 2017, 5, e13305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikami, S.; Oya, M.; Mizuno, R.; Kosaka, T.; Katsube, K.-I.; Okada, Y. Invasion and metastasis of renal cell carcinoma. Med. Mol. Morphol. 2013, 47, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Marona, P.; Górka, J.; Kotlinowski, J.; Majka, M.; Jura, J.; Miekus, K. C-Met as a Key Factor Responsible for Sustaining Undifferentiated Phenotype and Therapy Resistance in Renal Carcinomas. Cells 2019, 8, 272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yakirevich, E.; Patel, N.R. Tumor mutational burden and immune signatures interplay in renal cell carcinoma. Ann. Transl. Med. 2020, 8, 269. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, X.; Zhang, C.; Zhang, H.; Huang, Y. LINC00973 is involved in cancer immune suppression through positive regulation of Siglec-15 in clear-cell renal cell carcinoma. Cancer Sci. 2020, 111, 3693–3704. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Hu, W.; Wang, Y.; An, Y.; Song, L.; Shang, P.; Yue, Z. Long non-coding RNA UCA1 promotes malignant phenotypes of renal cancer cells by modulating the miR-182-5p/DLL4 axis as a ceRNA. Mol. Cancer 2020, 19, 1–17. [Google Scholar] [CrossRef]
- Zhang, Y.-J.; Lu, C. Long non-coding RNA HCP5 promotes proliferation and metastasis of clear cell renal cell carcinoma via targeting miR-140-5p/IGF1R pathway. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 2965–2975. [Google Scholar]
- Dasgupta, P.; Kulkarni, P.; Majid, S.; Hashimoto, Y.; Shiina, M.; Shahryari, V.; Bhat, N.S.; Tabatabai, L.; Yamamura, S.; Saini, S.; et al. LncRNA CDKN2B-AS1/miR-141/cyclin D network regulates tumor progression and metastasis of renal cell carcinoma. Cell Death Dis. 2020, 11, 1–12. [Google Scholar] [CrossRef]
- Wang, L.; Zhu, X.; Song, X.; Xu, Y. Long noncoding RNA lung cancer associated transcript 1 promotes proliferation and invasion of clear cell renal cell carcinoma cells by negatively regulating miR-495-3p. J. Cell. Biochem. 2018, 119, 7599–7609. [Google Scholar] [CrossRef]
- Kulkarni, P.; Dasgupta, P.; Bhat, N.S.; Shahryari, V.; Shiina, M.; Hashimoto, Y.; Majid, S.; Deng, G.; Saini, S.; Tabatabai, Z.L.; et al. Elevated miR-182-5p Associates with Renal Cancer Cell Mitotic Arrest through Diminished MALAT-1 Expression. Mol. Cancer Res. 2018, 16, 1750–1760. [Google Scholar] [CrossRef] [Green Version]
- Chan, J.J.; Tay, Y. Noncoding RNA: RNA Regulatory Networks in Cancer. Int. J. Mol. Sci. 2018, 19, 1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, M.; Guo, H.; Chen, Y.; Li, L.; Zhang, L. DARS-AS1 promotes clear cell renal cell carcinoma by sequestering miR-194-5p to up-regulate DARS. Biomed. Pharmacother. 2020, 128, 110323. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhang, J.; Zhang, Z.; Feng, Z.; Wei, J.; Lu, J.; Fang, Y.; Liang, Y.; Cen, J.; Pan, Y.; et al. The putative tumor suppressor microRNA-30a-5p modulates clear cell renal cell carcinoma aggressiveness through repression of ZEB2. Cell Death Dis. 2017, 8, e2859. [Google Scholar] [CrossRef]
- Chiyomaru, T.; Fukuhara, S.; Saini, S.; Majid, S.; Deng, G.; Shahryari, V.; Chang, I.; Tanaka, Y.; Enokida, H.; Nakagawa, M.; et al. Long Non-coding RNA HOTAIR Is Targeted and Regulated by miR-141 in Human Cancer Cells. J. Biol. Chem. 2014, 289, 12550–12565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, Q.; Li, O.; Zheng, W.; Xiao, W.-Z.; Zhang, L.; Wu, D.; Cai, G.-Y.; He, J.C.; Chen, X.-M. LncRNA HOTAIR regulates HIF-1α/AXL signaling through inhibition of miR-217 in renal cell carcinoma. Cell Death Dis. 2017, 8, e2772. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Wu, G.; Zhang, Z.; Tang, Q.; Zheng, W.; Chen, X.; Chen, F.; Li, Q.; Che, X. Long non-coding RNA HOTTIP promotes renal cell carcinoma progression through the regulation of the miR-615/IGF-2 pathway. Int. J. Oncol. 2018, 53, 2278–2288. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Zhang, J.; Jin, J.; Yang, C.; Zhang, W.; Zhang, H.; Zheng, J.; Weng, Z. HOXA11-AS promotes the growth and invasion of renal cancer by sponging miR-146b-5p to upregulate MMP16 expression. J. Cell. Physiol. 2018, 233, 9611–9619. [Google Scholar] [CrossRef]
- Zhang, W.; Lu, Y.; Shi, H.; Li, X.; Zhang, Z.; Deng, X.; Yang, Y.; Wan, B. LncRNA ITGB2-AS1 promotes the progression of clear cell renal cell carcinoma by modulating miR-328-5p/HMGA1 axis. Hum. Cell 2021, 1–13. [Google Scholar] [CrossRef]
- Jin, Y.; Huang, R.; Xia, Y.; Huang, C.; Qiu, F.; Pu, J.; He, X.; Zhao, X. Long Noncoding RNAKIF9-AS1Regulates Transforming Growth Factor-β and Autophagy Signaling to Enhance Renal Cell Carcinoma Chemoresistance via microRNA-497-5p. DNA Cell Biol. 2020, 39, 1096–1103. [Google Scholar] [CrossRef]
- Deng, H.; Huang, C.; Wang, Y.; Jiang, H.; Peng, S.; Zhao, X. LINC00511 promotes the malignant phenotype of clear cell renal cell carcinoma by sponging microRNA-625 and thereby increasing cyclin D1 expression. Aging 2019, 11, 5975–5991. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhang, H.; Li, W.; Yan, Y.; Yao, X.; Gu, W. FOXM1-Activated LINC01094 Promotes Clear Cell Renal Cell Carcinoma Development via MicroRNA 224-5p/CHSY1. Mol. Cell. Biol. 2020, 40, e00357-19. [Google Scholar] [CrossRef] [PubMed]
- Qu, L.; Ding, J.; Chen, C.; Wu, Z.-J.; Liu, B.; Gao, Y.; Chen, W.; Liu, F.; Sun, W.; Li, X.-F.; et al. Exosome-Transmitted lncARSR Promotes Sunitinib Resistance in Renal Cancer by Acting as a Competing Endogenous RNA. Cancer Cell 2016, 29, 653–668. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Zhang, J. Long non-conding RNA LOXL1-AS1 sponges miR-589-5p to up-regulate CBX5 expression in renal cell carcinoma. Biosci. Rep. 2020, 40, BSR20200212. [Google Scholar] [CrossRef]
- Ye, Y.; Zhang, F.; Chen, Q.; Huang, Z.; Li, M. LncRNA MALAT1 modified progression of clear cell kidney carcinoma (KIRC) by regulation of miR-194-5p/ACVR2B signaling. Mol. Carcinog. 2018, 58, 279–292. [Google Scholar] [CrossRef]
- Xiao, H.; Tang, K.; Liu, P.; Chen, K.; Hu, J.; Zeng, J.; Xiao, W.; Yu, G.; Yao, W.; Zhou, H.; et al. LncRNA MALAT1 functions as a competing endogenous RNA to regulate ZEB2 expression by sponging miR-200s in clear cell kidney carcinoma. Oncotarget 2015, 6, 38005–38015. [Google Scholar] [CrossRef]
- Zhang, H.; Li, W.; Gu, W.; Yan, Y.; Yao, X.; Zheng, J. MALAT1 accelerates the development and progression of renal cell carcinoma by decreasing the expression of miR-203 and promoting the expression of BIRC5. Cell Prolif. 2019, 52, e12640. [Google Scholar] [CrossRef] [Green Version]
- Qu, Y.; Xiao, H.; Xiao, W.; Xiong, Z.; Hu, W.; Gao, Y.; Ru, Z.; Wang, C.; Bao, L.; Wang, K.; et al. Upregulation of MIAT Regulates LOXL2 Expression by Competitively Binding MiR-29c in Clear Cell Renal Cell Carcinoma. Cell. Physiol. Biochem. 2018, 48, 1075–1087. [Google Scholar] [CrossRef]
- Tao, M.; Zhou, Y.; Jin, Y.; Pu, J. Blocking lncRNA MIR155HG/miR-155-5p/-3p inhibits proliferation, invasion and migration of clear cell renal cell carcinoma. Pathol. Res. Pract. 2019, 216, 152803. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.; Miao, C.; Tian, Y.; Qin, Z.; Xue, J.; Xia, J.; Zhu, S.; Xu, A.; Yang, J.; Wang, Z. lncRNA MIR4435-2HG promoted clear cell renal cell carcinoma malignant progression via miR-513a-5p/KLF6 axis. J. Cell. Mol. Med. 2020, 24, 10013–10026. [Google Scholar] [CrossRef]
- Cai, X.; Zhang, X.; Mo, L.; Zhu, J.; Yu, H. LncRNA PCGEM1 promotes renal carcinoma progression by targeting miR-433-3p to regulate FGF2 expression. Cancer Biomarkers 2020, 27, 493–504. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Zhang, D.; Zhong, Z.; Zhang, W. lncRNA ROR promotes the progression of renal cell carcinoma through the miR-206/VEGF axis. Mol. Med. Rep. 2019, 20, 3782–3792. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Jin, W.; Song, Y.; Fei, X. LncRNA RP11-436H11.5, functioning as a competitive endogenous RNA, upregulates BCL-W expression by sponging miR-335-5p and promotes proliferation and invasion in renal cell carcinoma. Mol. Cancer 2017, 16, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Qu, Y.; Xiao, H.; Xiao, W.; Liu, J.; Gao, Y.; Li, M.; Liu, J. LncRNA SNHG3 promotes clear cell renal cell carcinoma proliferation and migration by upregulating TOP2A. Exp. Cell Res. 2019, 384, 111595. [Google Scholar] [CrossRef] [PubMed]
- Xiang, W.; Lv, L.; Zhou, G.; Wu, W.; Yuan, J.; Zhang, C.; Jiang, G. The lncRNA SNHG5-mediated miR-205-5p downregulation contributes to the progression of clear cell renal cell carcinoma by targeting ZEB1. Cancer Med. 2020, 9, 4251–4264. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Liang, H.; Zhou, J.; Wang, Y.; Liu, S.; Wang, X.; Su, L.; Kang, X. lncRNA small nucleolar RNA host gene 12 promotes renal cell carcinoma progression by modulating the miR-200c-5p/collagen type XI α1 chain pathway. Mol. Med. Rep. 2020, 22, 3677–3686. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.; Shuang, W.; Ye, D.; Zhang, W.; Yang, Z.; Fang, W.; Xu, H.; Gu, M.; Xu, W.; Guan, C. SNHG16 promotes cell proliferation and inhibits cell apoptosis via regulation of the miR-1303-p/STARD9 axis in clear cell renal cell carcinoma. Cell. Signal. 2021, 84, 110013. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Sun, D.; Yu, J.; Zhang, M.; Yi, C.; Yang, R.; Dan, B.; Li, A. Long noncoding RNA TUG1 promotes renal cell carcinoma cell proliferation, migration and invasion by downregulating microRNA-196a. Mol. Med. Rep. 2018, 18, 5791–5798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zheng, D.; Pan, L.; Dai, Y.; Cai, S.; Zhao, L.; Zhu, H. Knockdown of TUG1 by shRNA inhibited renal cell carcinoma formation by miR-299–3p/VEGF axis in vitro and in vivo. Eur. J. Pharmacol. 2019, 860, 172536. [Google Scholar] [CrossRef]
- Liu, S.; Yang, Y.; Wang, W.; Pan, X. Long noncoding RNA TUG1 promotes cell proliferation and migration of renal cell carcinoma via regulation of YAP. J. Cell. Biochem. 2018, 119, 9694–9706. [Google Scholar] [CrossRef]
- Dong, D.; Mu, Z.; Wei, N.; Sun, M.; Wang, W.; Xin, N.; Shao, Y.; Zhao, C. Long non-coding RNA ZFAS1 promotes proliferation and metastasis of clear cell renal cell carcinoma via targeting miR-10a/SKA1 pathway. Biomed. Pharmacother. 2019, 111, 917–925. [Google Scholar] [CrossRef]
- Song, E.-L.; Xing, L.; Wang, L.; Song, W.-T.; Li, D.-B.; Wang, Y.; Gu, Y.-W.; Liu, M.-M.; Ni, W.-J.; Zhang, P.; et al. LncRNA ADAMTS9-AS2 inhibits cell proliferation and decreases chemoresistance in clear cell renal cell carcinoma via the miR-27a-3p/FOXO1 axis. Aging 2019, 11, 5705–5725. [Google Scholar] [CrossRef]
- Liu, L.; Pang, X.; Shang, W.; Xie, H.; Feng, Y.; Feng, G. Long non-coding RNA GAS5 sensitizes renal cell carcinoma to sorafenib via miR-21/SOX5 pathway. Cell Cycle 2019, 18, 257–263. [Google Scholar] [CrossRef]
- Zeng, J.; Li, Y.; Wang, Y.; Xie, G.; Feng, Q.; Yang, Y.; Feng, J. lncRNA 00312 Attenuates Cell Proliferation and Invasion and Promotes Apoptosis in Renal Cell Carcinoma via miR-34a-5p/ASS1 Axis. Oxidative Med. Cell. Longev. 2020, 2020, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Qi, Y.; Ma, Y.; Peng, Z.; Wang, L.; Li, L.; Tang, Y.; He, J.; Zheng, J. Long noncoding RNA PENG upregulates PDZK1 expression by sponging miR-15b to suppress clear cell renal cell carcinoma cell proliferation. Oncogene 2020, 39, 4404–4420. [Google Scholar] [CrossRef]
- He, H.; Dai, J.; Zhuo, R.; Zhao, J.; Wang, H.; Sun, F.; Zhu, Y.; Xu, D. Study on the mechanism behind lncRNA MEG3 affecting clear cell renal cell carcinoma by regulating miR-7/ RASL11B signaling. J. Cell. Physiol. 2018, 233, 9503–9515. [Google Scholar] [CrossRef]
- Xue, S.; Wang, S.; Li, J.; Guan, H.; Jiang, S.; Guo, Y.; Li, Q. LncRNA NBAT1 suppresses cell proliferation and migration via miR-346/GSK-3β axis in renal carcinoma. IUBMB Life 2019, 71, 1720–1728. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Jia, Z.; Duan, R.; Yan, Z.; Jin, Z.; Yan, L.; Li, Q.; Yang, J. Long non-coding RNA XIST regulates miR-106b-5p/P21 axis to suppress tumor progression in renal cell carcinoma. Biochem. Biophys. Res. Commun. 2019, 510, 416–420. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Sun, J.; Liu, L.N.; Flies, D.B.; Nie, X.; Toki, M.; Zhang, J.; Song, C.; Zarr, M.; Zhou, X.; et al. Siglec-15 as an immune suppressor and potential target for normalization cancer immunotherapy. Nat. Med. 2019, 25, 656–666. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-H.; Lan, C.-T.; Chou, J.-F.; Tseng, T.-J.; Liao, W.-C. CHSY1 promotes aggressive phenotypes of hepatocellular carcinoma cells via activation of the hedgehog signaling pathway. Cancer Lett. 2017, 403, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Qian, J.; Luo, X.; Zhou, A.; Zhang, Z.; Fang, Q. CHSY1 promoted proliferation and suppressed apoptosis in colorectal cancer through regulation of the NFκB and/or caspase-3/7 signaling pathway. Oncol. Lett. 2018, 16, 6140–6146. [Google Scholar] [CrossRef]
- Boguslawska, J.; Kedzierska, H.; Poplawski, P.; Rybicka, B.; Tanski, Z.; Piekielko-Witkowska, A. Expression of Genes Involved in Cellular Adhesion and Extracellular Matrix Remodeling Correlates with Poor Survival of Patients with Renal Cancer. J. Urol. 2016, 195, 1892–1902. [Google Scholar] [CrossRef]
- Zheng, J.; Wang, L.; Peng, Z.; Yang, Y.; Feng, D.; He, J. Low level of PDZ domain containing 1 (PDZK1) predicts poor clinical outcome in patients with clear cell renal cell carcinoma. EBioMedicine 2016, 15, 62–72. [Google Scholar] [CrossRef] [Green Version]
- Zeng, X.; Hu, Z.; Ke, X.; Tang, H.; Wu, B.; Wei, X.; Liu, Z. Long noncoding RNA DLX6-AS1 promotes renal cell carcinoma progression via miR-26a/PTEN axis. Cell Cycle 2017, 16, 2212–2219. [Google Scholar] [CrossRef] [Green Version]
- Que, W.-C.; Qiu, H.-Q.; Cheng, Y.; Liu, M.-B.; Wu, C.-Y. PTEN in kidney cancer: A review and meta-analysis. Clin. Chim. Acta 2018, 480, 92–98. [Google Scholar] [CrossRef]
- Liu, X.; Gomez-Pinillos, A.; Loder, C.; Pau, E.C.D.S.; Qiao, R.; Unger, P.D.; Kurek, R.; Oddoux, C.; Melamed, J.; Gallagher, R.E.; et al. KLF6 Loss of Function in Human Prostate Cancer Progression Is Implicated in Resistance to Androgen Deprivation. Am. J. Pathol. 2012, 181, 1007–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, U.E.; Kocabayoglu, P.; Cheng, G.Z.; Ghiassi-Nejad, Z.; Muñoz, U.; Vetter, D.; Eckstein, D.A.; Hannivoort, R.A.; Walsh, M.J.; Friedman, S.L. GSK3β phosphorylation of the KLF6 tumor suppressor promotes its transactivation of p21. Oncogene 2012, 32, 4557–4564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Guo, D.-D.; Zheng, J.-Y.; Wu, Y.-A. Expression of KLF6-SV2 in colorectal cancer and its impact on proliferation and apoptosis. Eur. J. Cancer Prev. 2018, 27, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Cheng, G.; Huang, Z.; Bao, L.; Liu, J.; Wang, C.; Xiong, Z.; Zhou, L.; Xu, T.; Liu, D.; et al. Long noncoding RNA SNHG12 promotes tumour progression and sunitinib resistance by upregulating CDCA3 in renal cell carcinoma. Cell Death Dis. 2020, 11, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Wu, Q.; Zhang, Y.; Xiong, H.; Li, X.; Li, B.; Xie, W.; Zhang, L.; Xu, M.; Zhang, K.; et al. LncRNA LOC653786 promotes growth of RCC cells via upregulating FOXM1. Oncotarget 2018, 9, 12101–12111. [Google Scholar] [CrossRef]
- Zhai, W.; Zhu, R.; Ma, J.; Gong, D.; Zhang, H.; Zhang, J.; Chen, Y.; Huang, Y.; Zheng, J.; Xue, W. A positive feed-forward loop between LncRNA-URRCC and EGFL7/P-AKT/FOXO3 signaling promotes proliferation and metastasis of clear cell renal cell carcinoma. Mol. Cancer 2019, 18, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Peng, F.; Shi, X.; Meng, Y.; Dong, B.; Xu, G.; Hou, T.; Shi, Y.; Liu, T. Long non-coding RNA HOTTIP is upregulated in renal cell carcinoma and regulates cell growth and apoptosis by epigenetically silencing of LATS2. Biomed. Pharmacother. 2018, 105, 1133–1140. [Google Scholar] [CrossRef]
- Song, C.; Xiong, Y.; Liao, W.; Meng, L.; Yang, S.-X. Long noncoding RNA ATB participates in the development of renal cell carcinoma by downregulating p53 via binding to DNMT1. J. Cell. Physiol. 2018, 234, 12910–12917. [Google Scholar] [CrossRef]
- Shi, H.; Sun, Y.; He, M.; Yang, X.; Hamada, M.; Fukunaga, T.; Zhang, X.; Chang, C. Targeting the TR4 nuclear receptor-mediated lncTASR/AXL signaling with tretinoin increases the sunitinib sensitivity to better suppress the RCC progression. Oncogene 2019, 39, 530–545. [Google Scholar] [CrossRef]
- You, B.; Sun, Y.; Luo, J.; Wang, K.; Liu, Q.; Fang, R.; Liu, B.; Chou, F.; Wang, R.; Meng, J.; et al. Androgen receptor promotes renal cell carcinoma (RCC) vasculogenic mimicry (VM) via altering TWIST1 nonsense-mediated decay through lncRNA-TANAR. Oncogene 2021, 40, 1674–1689. [Google Scholar] [CrossRef]
- Wang, A.; Bao, Y.; Wu, Z.; Zhao, T.; Wang, D.; Shi, J.; Liu, B.; Sun, S.; Yang, F.; Wang, L.; et al. Long noncoding RNA EGFR-AS1 promotes cell growth and metastasis via affecting HuR mediated mRNA stability of EGFR in renal cancer. Cell Death Dis. 2019, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Ma, P.; Zhao, Y.; Li, B.; Jiang, S.; Xiong, H.; Wang, Z.; Wang, H.; Jin, X.; Liu, C. Biological function and mechanism of MALAT-1 in renal cell carcinoma proliferation and apoptosis: Role of the MALAT-1–Livin protein interaction. J. Physiol. Sci. 2016, 67, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Dong, B.; Zhang, J.; Zhai, W.; Xie, T.; Huang, B.; Huang, C.; Yao, X.; Zheng, J.; Che, J.; et al. The long noncoding RNA HOTAIR activates the Hippo pathway by directly binding to SAV1 in renal cell carcinoma. Oncotarget 2017, 8, 58654–58667. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Yang, F.; Wei, D.; Liu, B.; Chen, C.; Bao, Y.; Wu, Z.; Wu, D.; Tan, H.; Li, J.; et al. Long noncoding RNA-SRLR elicits intrinsic sorafenib resistance via evoking IL-6/STAT3 axis in renal cell carcinoma. Oncogene 2016, 36, 1965–1977. [Google Scholar] [CrossRef] [PubMed]
- Qu, L.; Wu, Z.; Li, Y.; Xu, Z.; Liu, B.; Liu, F.; Bao, Y.; Wu, D.; Liu, J.; Wang, A.; et al. A feed-forward loop between lncARSR and YAP activity promotes expansion of renal tumour-initiating cells. Nat. Commun. 2016, 7, 12692. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.-T.; Huang, H.; Huang, W.-P.; Hu, W.-L. LncRNA THOR promotes human renal cell carcinoma cell growth. Biochem. Biophys. Res. Commun. 2018, 501, 661–667. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.; Tang, Y.; Li, H.; Li, P.; Ye, Y.; Cen, J.; Gui, C.; Luo, J.; Cao, J.; Wei, J. N6-Methyladenosine Modification of LncRNA DUXAP9 Promotes Renal Cancer Cells Proliferation and Motility by Activating the PI3K/AKT Signaling Pathway. Front. Oncol. 2021, 11, 1363. [Google Scholar] [CrossRef]
- Wang, G.; Li, H.; Hou, Y. LncRNA MAGI2-AS3 inhibits tumor progression and angiogenesis by regulating ACY1 via interacting with transcription factor HEY1 in clear cell renal cell carcinoma. Cancer Gene Ther. 2021, 1–12. [Google Scholar] [CrossRef]
- Pu, Y.; Dong, Z.; Xia, Y.; Zhang, M.; Song, J.; Han, J.; Liu, H. LncRNA NONHSAT113026 represses renal cell carcinoma tumorigenesis through interacting with NF-κB/p50 and SLUG. Biomed. Pharmacother. 2019, 118, 109382. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, P.; Dasgupta, P.; Hashimoto, Y.; Shiina, M.; Shahryari, V.; Tabatabai, Z.L.; Yamamura, S.; Tanaka, Y.; Saini, S.; Dahiya, R.; et al. A lncRNA TCL6-miR-155 Interaction Regulates the Src-Akt-EMT Network to Mediate Kidney Cancer Progression and Metastasis. Cancer Res. 2021, 81, 1500–1512. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.-D.; Han, L.; Lee, H.; Zhuang, L.; Zhang, Y.; Baddour, J.; Nagrath, D.; Wood, C.G.; Gu, J.; Wu, X.; et al. Energy stress-induced lncRNA FILNC1 represses c-Myc-mediated energy metabolism and inhibits renal tumor development. Nat. Commun. 2017, 8, 1–13. [Google Scholar] [CrossRef]
- Zhai, W.; Sun, Y.; Guo, C.; Junhua, Z.; Wang, M.; Zheng, J.; Lin, W.; Huang, Q.; Li, G.; Zheng, J.; et al. LncRNA-SARCC suppresses renal cell carcinoma (RCC) progression via altering the androgen receptor (AR)/miRNA-143-3p signals. Cell Death Differ. 2017, 24, 1502–1517. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Wei, P.; Lv, W.; Han, X.; Yang, J.; Qin, S. Long noncoding RNA lnc-DILC stabilizes PTEN and suppresses clear cell renal cell carcinoma progression. Cell Biosci. 2019, 9, 1–13. [Google Scholar] [CrossRef]
- Yang, F.; Liu, C.; Zhao, G.; Ge, L.; Song, Y.; Chen, Z.; Liu, Z.; Hong, K.; Ma, L. Long non-coding RNA LINC01234 regulates proliferation, migration and invasion via HIF-2α pathways in clear cell renal cell carcinoma cells. PeerJ 2020, 8, e10149. [Google Scholar] [CrossRef]
- Yang, Y.; Dong, M.H.; Hu, H.M.; Min, Q.H.; Xiao, L. LncRNA FGD5-AS1/miR-5590-3p axis facilitates the proliferation and metastasis of renal cell carcinoma through ERK/AKT signalling. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 8756–8766. [Google Scholar] [CrossRef]
- Zhang, C.; Li, X.-Y.; Luo, Z.-Z.; Wu, T.-W.; Hu, H. Upregulation of LINC00982 inhibits cell proliferation and promotes cell apoptosis by regulating the activity of PI3K/AKT signaling pathway in renal cancer. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 1443–1450. [Google Scholar]
- Yue, G.; Chen, C.; Bai, L.; Wang, G.; Huang, Y.; Wang, Y.; Cui, H.; Xiao, Y. Knockdown of long noncoding RNA DLEU1 suppresses the progression of renal cell carcinoma by downregulating the Akt pathway. Mol. Med. Rep. 2019, 20, 4551–4557. [Google Scholar] [CrossRef]
- Zheng, Z.; Zhao, F.; Zhu, D.; Han, J.; Chen, H.; Cai, Y.; Chen, Z.; Xie, W. Long Non-Coding RNA LUCAT1 Promotes Proliferation and Invasion in Clear Cell Renal Cell Carcinoma Through AKT/GSK-3β Signaling Pathway. Cell. Physiol. Biochem. 2018, 48, 891–904. [Google Scholar] [CrossRef]
- Li, Z.; Ma, Z.; Xu, X. Long non-coding RNA MALAT1 correlates with cell viability and mobility by targeting miR-22-3p in renal cell carcinoma via the PI3K/Akt pathway. Oncol. Rep. 2018, 41, 1113–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Y.; Lu, J.; Chen, X.; Liang, C.; Luo, P.; Qin, C.; Zhang, J. Long non-coding RNA HOTTIP affects renal cell carcinoma progression by regulating autophagy via the PI3K/Akt/Atg13 signaling pathway. J. Cancer Res. Clin. Oncol. 2018, 145, 573–588. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Zhao, X.; Zhou, J.; Cheng, X.; Ye, Z.; Ji, Z. LncRNA TP73-AS1 Promotes Cell Proliferation and Inhibits Cell Apoptosis in Clear Cell Renal Cell Carcinoma Through Repressing KISS1 Expression and Inactivation of PI3K/Akt/mTOR Signaling Pathway. Cell. Physiol. Biochem. 2018, 48, 371–384. [Google Scholar] [CrossRef]
- Zhong, W.; Zhang, F.; Huang, C.; Lin, Y.; Huang, J. Identification of Epithelial–Mesenchymal Transition-Related lncRNA With Prognosis and Molecular Subtypes in Clear Cell Renal Cell Carcinoma. Front. Oncol. 2020, 10, 2619. [Google Scholar] [CrossRef] [PubMed]
- Zuo, S.; Wang, L.; Wen, Y.; Dai, G. Identification of a universal 6-lncRNA prognostic signature for three pathologic subtypes of renal cell carcinoma. J. Cell. Biochem. 2018, 120, 7375–7385. [Google Scholar] [CrossRef]
- Sourbier, C.; Liao, P.-J.; Ricketts, C.J.; Wei, D.; Yang, Y.; Baranes, S.M.; Gibbs, B.; Ohanjanian, L.; Krane, L.S.; Scroggins, B.T.; et al. Targeting loss of the Hippo signaling pathway in NF2-deficient papillary kidney cancers. Oncotarget 2018, 9, 10723–10733. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| LncRNA/miRNA/mRNA Axis | Effect on Pathogenesis | Refs. |
|---|---|---|
| Oncogenic lncRNAs | ||
| CDKN2B-AS1/miR-141/ CCND1, CCND2 | migration, invasion, and clonogenicity in vitro/in vivo, RAC1/pPXN p/w, EMT, poor survival | [38] |
| DARS-AS1/miR-194-5p/DARS | proliferation, apoptosis inhibition | [42] |
| DLEU2/miR-30a-5p/ZEB2 | EMT, migration, invasion, metastasis, short OS | [43] |
| HCP5/miR-140-5p/IGF1R | migration, invasion, metastasis, cell cycle, poor OS and PFS | [37] |
| HOTAIR/miR-141/ ABL2, Snail, ZEB1 | proliferation, invasion | [44] |
| HOTAIR/miR-217/HIF-1α | migration, EMT, AXL signaling | [45] |
| HOTTIP/miR-615/IGF-2 | decreased apoptosis, enhanced migration and invasion in vitro, metastasis, vascular invasion | [46] |
| HOXA11-AS/miR-146b-5p/ MMP16 | invasion, EMT, advanced stage, metastasis | [47] |
| ITGB2-AS1/miR-328-5p/HMGA1 | tumorigenesis in vitro/in vivo, poorer prognosis | [48] |
| KIF9-AS1/miR-497-5p/ SMAD3, ATG9A | decreased apoptosis, enhanced cell viability, resistance to sorafenib, TGF-β/autophagy p/w | [49] |
| LINC00511/miR-625/CCND1 | metastasis, cell cycle, short OS | [50] |
| LINC00973/miR-7109/Siglec-15 | ccRCC progression, cancer immune evasion | [35] |
| FOXM1→LINC01094/ miR-224-5p/CHSY1 | tumor cell growth, metastasis | [51] |
| lncARSR/miR-34, miR-449/ AXL, c-MET | promotion of sunitinib resistance, activation of STAT3, AKT, and ERK p/w | [52] |
| LOXL1-AS1/miR-589-5p/CBX5 | cell proliferation, migration | [53] |
| LUCAT1/miR-495-3p/SATB1 | proliferation and invasion in vitro/in vivo, metastasis, shorter OS | [39] |
| MALAT1/miR-182-5p/ CDC20, AURKA | progression in vitro/in vivo, cell cycle | [40] |
| MALAT1/miR-194-5p/ACVR2B | cell viability, proliferation, colony formation | [54] |
| MALAT1/miR-200c/ZEB2 | invasion and metastasis in vitro/in vivo, EMT | [55] |
| MALAT1/miR-203/BIRC5 | invasion and migration in vitro/in vivo, cell cycle | [56] |
| c-Fos→MALAT1/miR-205/Ezh2 | EMT, invasion, decreased apoptosis, shorter OS | [12] |
| MIAT/miR-29c/Loxl2 | proliferation and metastasis in vitro/in vivo, poor OS and DFS | [57] |
| MIR155HG/miR-155-5p(-3p)/ MMP2, MMP9 | invasion, migration | [58] |
| MIR4435-2HG/miR-513a-5p/KLF6 | proliferation in vitro/in vivo, invasion, metastasis | [59] |
| PCGEM1/miR-433-3p/FGF2 | cell proliferation, migration, apoptosis repression | [60] |
| ROR/miR-206/VEGF | migration, invasion, metastasis | [61] |
| RP11-436H11.5/miR-335-5p/ BCL-W | proliferation and invasion in vitro/in vivo, poor survival | [62] |
| SNHG3/miR-139-5p/TOP2A | proliferation and metastasis in vitro/in vivo, worse OS and DFS | [63] |
| SNHG5/miR-205-5p/ZEB1 | lymph node invasion, distant metastasis, EMT | [64] |
| SNHG12/miR-200c-5p/COL11A1 | invasion, suppression of apoptosis, poor survival | [65] |
| SNHG16/miR-1303-p/STARD9 | proliferation, suppression of apoptosis | [66] |
| TUG1/miR-196a/AKT, ERK, JNK | proliferation, migration, and invasion in vitro | [67] |
| TUG1/miR-299-3p/VEGF | invasion, migration, EMT | [68] |
| TUG1/miR-9/YAP1 | proliferation, migration, did not alter Hippo p/w or YAP1 protein distribution | [69] |
| UCA1/miR-182-5p/DLL4 | migration, tumorigenicity, Notch signaling | [36] |
| ZFAS1/miR-10a/SKA1 | migration, invasion, poor prognosis, shorter OS | [70] |
| Suppressive lncRNAs | ||
| ADAMTS9-AS2/miR-27a-3p/ FOXO1 | decreased proliferation, decreased chemoresistance | [71] |
| GAS5/miR-21/SOX5 | sensitivity to sorafenib in vitro/in vivo | [72] |
| lncRNA 00312/miR-34a-5p/ASS1 | apoptosis and inhibition of invasion in vitro, better OS | [73] |
| lncPENG/miR-15b/PDZK1 | proliferation inhibition in vitro/in vivo, better survival | [74] |
| MEG3/miR-7/RASL11B | apoptosis, G0/G1 arrest, migration, invasion inhibition | [75] |
| NBAT1/miR-346/GSK-3β | migration, invasion suppression, Wnt/β-catenin p/w | [76] |
| XIST/miR-106b-5p/P21 | G0/G1 arrest, tumor suppression in vitro/in vivo | [77] |
| LncRNA/Protein | Mechanisms of Action | Effect on Pathogenesis, Survival, Drug Resistance | Refs. |
|---|---|---|---|
| Oncogenic lncRNAs | |||
| Regulation of transcription | |||
| SNHG12/SP1/CDCA3↑ | binding to SP1 and preventing the ubiquitylation-dependent proteolysis of SP1; stabilized SP1 bound to the promoter of CDCA3 and increased CDCA3 expression | tumor progression and sunitinib resistance, promoting proliferation, migration, invasion in vivo | [88] |
| LOC653786/FOXM1↑ | enhancing the transcriptional activity of FOXM1 gene promoter | elevating the expression of FOXM1 downstream target genes cyclin D1 and cyclin B1, promoting cell cycle progression of RCC and its growth in vitro and in vivo | [89] |
| Chromatin reorganization | |||
| URRCC/EGFL7↑ | enhances the expression of EGFL7 by mediating histone H3 acetylation of EGFL7 promoter | activation of P-AKT signaling, and suppressing FOXO3, promotes proliferation and metastasis | [90] |
| HOTTIP/EZH2, LSD1/LATS2↓ | binding to the enhancer of zeste homolog 2 (EZH2) and lysine specific demethylase 1 (LSD1), and repressing LATS2 expression | cell growth and apoptosis inhibition | [91] |
| ATB/DNMT1↑/p53↓ | binding to DNMT1 and stabilizing its expression; promoting the binding of DNMT1 to p53 | promoted proliferative and migratory capacities but inhibited apoptosis | [92] |
| Binding to mRNA | |||
| TASR/AXL↑ | binding to the 5′-UTR of AXL mRNA and its stabilization | sunitinib resistance | [93] |
| AR/TANAR/TWIST1↑ | binding to the 5′-UTR of TWIST1 mRNA and inhibition of its nonsense-mediated mRNA decay (NMD) | vasculogenic mimicry and metastasis | [94] |
| EGFR-AS1/HuR/EGFR↑ | binding to EGFR mRNA and enhancing HuR-mediated mRNA stability | promoted cell proliferation and invasion in vitro and in vivo | [95] |
| Binding to protein | |||
| MALAT1/Livin↑ | binding to Livin and enhancing its stability | promotion of proliferation and metastasis | [96] |
| HOTAIR/SAV1↓ | binding to the SAV1 protein inhibits its interaction with MST1/2, activation of LATS1/2, and subsequent phosphorylation of YAP1 | promotion of RCC development and growth by activating the Hippo pathway through direct binding to the SAV1, as a result—promotion of YAP1 translocation to the nucleus | [97] |
| SRLR/NF-κB/IL-6↑ | binding to NF-κB | binding to NF-κB and promoting IL-6 transcription, leading to the activation of STAT3 and the development of sorafenib tolerance | [98] |
| ARSR/YAP1↑/ARSR | binding of lncARSR to YAP1 impedes LATS1-induced YAP1 phosphorylation and facilitates YAP1 nuclear translocation, reciprocally, YAP1/TEAD promotes lncARSR transcription | self-renewal, tumorigenicity and metastasis, tumor-initiating cells properties | [99] |
| THOR/IGF2BP1↑/ IGF2↑, GLI1↑, MYC↑ | THOR directly associates with insulin-like growth factor 2 mRNA-binding protein 1 (IGF2BP1) to promote mRNA stabilization of IGF2BP1-regulated genes, including IGF2, GLI1, and MYC | cell growth, viability, and proliferation | [100] |
| DUXAP9/IGF2BP2↑ | methylation at N6-adenosine, binding to IGF2BP2, which increases its stability | cell proliferation and motility, EMT, activate PI3K/AKT pathway and Snail expression | [101] |
| Suppressive lncRNAs | |||
| Regulation of transcription | |||
| MAGI2-AS3/HEY1/ACY1↑ | binding of MAGI2-AS3 with HEY1 and reducing the HEY1 enrichment at the ACY1 promoter region, increasing ACY1 gene transcription | overexpression of MAGI2-AS3 reduces ccRCC cell viability and migration, inhibits vessel-like tube formation of HUVECs in vitro, and represses tumor growth and angiogenesis in vivo | [102] |
| Binding to mRNA | |||
| NONHSAT 113026 (NOAT113026)/ (NF-κB/p50) ↓, SLUG↓ | binding to the 3′-UTR of mRNA for NF-κB/p50 and SLUG and reducing their expression | inhibits the ability of cell migration, invasion, proliferation, colony formation, EMT in vitro, tumorigenesis in vivo | [103] |
| TCL6/miR-155/STAU1/Src↓ | recruiting STAU1 and mediation of Src mRNA decay, the interaction between miR-155 and lncTCL6 attenuates this process | repress of cell proliferation and migration/invasion, EMT and induced cell-cycle arrest and apoptosis, inhibits Src-Akt-EMT network | [104] |
| Binding to protein | |||
| FILNC1/AUF1↓/ c-Myc↓ | interaction of FILNC1 with AUF1, a protein that binds c-Myc mRNA, and sequestering of AUF1 from the binding of c-Myc mRNA. This leads to suppression of the c-Myc protein expression | energy stress-induced apoptosis, inhibition of Warburg effect and tumor development | [105] |
| SARCC/AR↓/miR-143-3p↑ | binding and destabilizing AR protein, preventing AR movement from the cytoplasm to the nucleus, preventing AR from interacting with HSP90 | AR could directly decrease miR-143-3p, so, de-repressing miR-143-3p expression entails the expression inhibition of AKT, MMP-13, K-RAS, and P-ERK, attenuation of cell invasion, migration, and proliferation in vitro and in vivo | [106] |
| lnc-DILC/WWP2/USP11/PTEN↑ | repressing PTEN ubiquitination through blocking the interaction between PTEN and E3 ubiquitin ligase WWP2 and recruiting the deubiquitinase USP11 to PTEN. | inhibits cell proliferation, migration, and invasion | [107] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Braga, E.A.; Fridman, M.V.; Filippova, E.A.; Loginov, V.I.; Pronina, I.V.; Burdennyy, A.M.; Karpukhin, A.V.; Dmitriev, A.A.; Morozov, S.G. LncRNAs in the Regulation of Genes and Signaling Pathways through miRNA-Mediated and Other Mechanisms in Clear Cell Renal Cell Carcinoma. Int. J. Mol. Sci. 2021, 22, 11193. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011193
Braga EA, Fridman MV, Filippova EA, Loginov VI, Pronina IV, Burdennyy AM, Karpukhin AV, Dmitriev AA, Morozov SG. LncRNAs in the Regulation of Genes and Signaling Pathways through miRNA-Mediated and Other Mechanisms in Clear Cell Renal Cell Carcinoma. International Journal of Molecular Sciences. 2021; 22(20):11193. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011193
Chicago/Turabian StyleBraga, Eleonora A., Marina V. Fridman, Elena A. Filippova, Vitaly I. Loginov, Irina V. Pronina, Alexey M. Burdennyy, Alexander V. Karpukhin, Alexey A. Dmitriev, and Sergey G. Morozov. 2021. "LncRNAs in the Regulation of Genes and Signaling Pathways through miRNA-Mediated and Other Mechanisms in Clear Cell Renal Cell Carcinoma" International Journal of Molecular Sciences 22, no. 20: 11193. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011193