
Functions of PPR Proteins in Plant Growth and Development

School of Life Sciences, Qufu Normal University, Qufu 273165, China
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2021, 22(20), 11274; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011274
Submission received: 16 September 2021
/
Revised: 14 October 2021
/
Accepted: 16 October 2021
/
Published: 19 October 2021
(This article belongs to the Special Issue Research Progress in RNA-Binding Proteins)
Abstract
:Pentatricopeptide repeat (PPR) proteins form a large protein family in land plants, with hundreds of different members in angiosperms. In the last decade, a number of studies have shown that PPR proteins are sequence-specific RNA-binding proteins involved in multiple aspects of plant organellar RNA processing, and perform numerous functions in plants throughout their life cycle. Recently, computational and structural studies have provided new insights into the working mechanisms of PPR proteins in RNA recognition and cytidine deamination. In this review, we summarized the research progress on the functions of PPR proteins in plant growth and development, with a particular focus on their effects on cytoplasmic male sterility, stress responses, and seed development. We also documented the molecular mechanisms of PPR proteins in mediating RNA processing in plant mitochondria and chloroplasts.
1. Introduction
Pentatricopeptide repeat (PPR) proteins are characterized by the presence of tandem arrays of a degenerate 35-amino-acid repeat motif, PPR motif [1]. Based on the types of motif and their arrangement, PPR proteins are divided into two classes, P and PLS. P-class proteins only contain canonical P-motifs with 35 amino acids, whereas PLS-class proteins consist of P-, L- (35 or 36 amino acids), and S- (31 or 32 amino acids) motifs forming tandemly repeated PLS triplets [2]. Many of the PLS-class proteins are carboxyl terminally extended with highly conserved E, E+, or DYW domains. Thus, PLS-class proteins can be further divided into PLS, E, E+, and DYW subclasses according to the domains identified in carboxyl terminal [3].
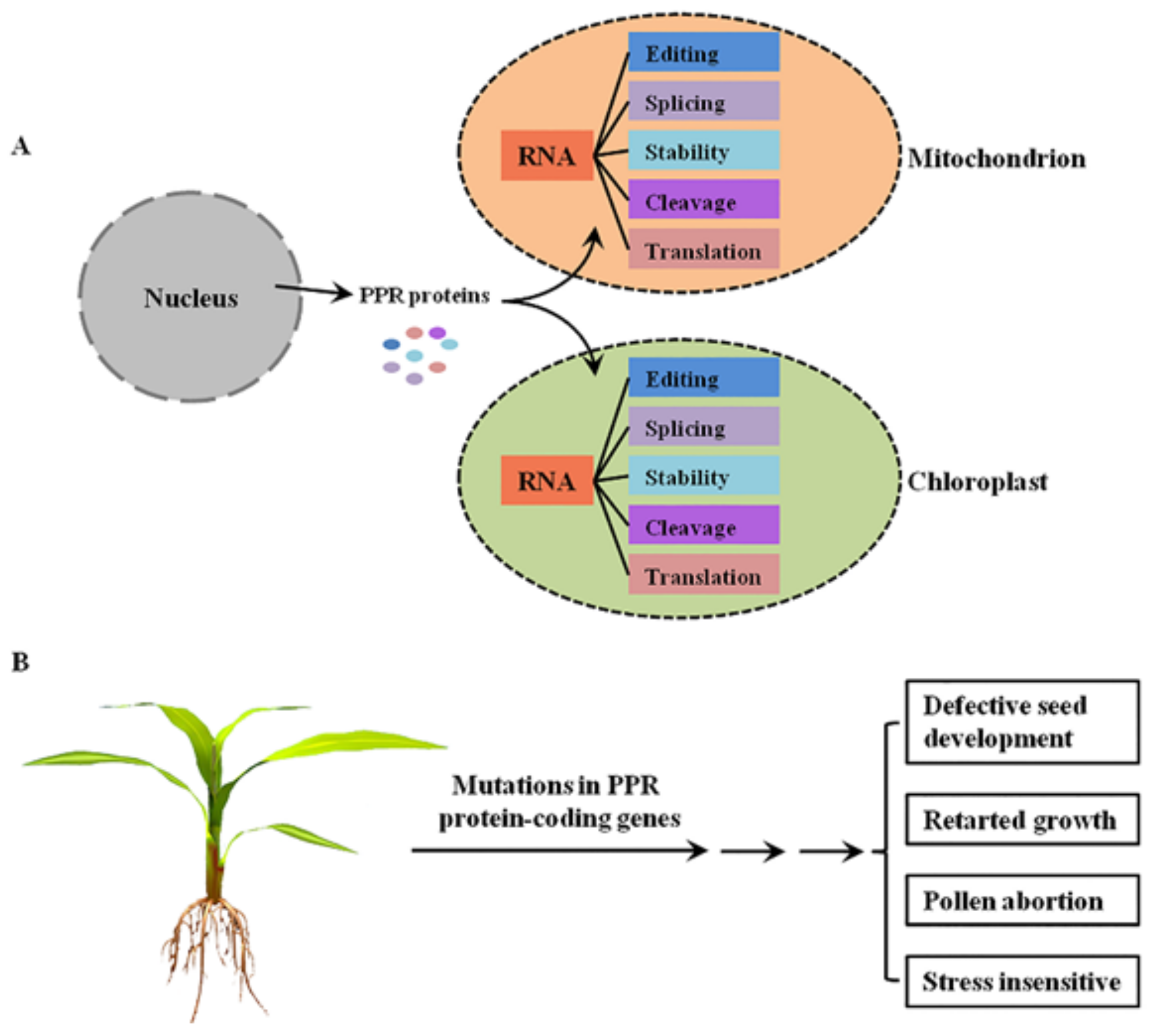
PPR proteins are sequence-specific RNA-binding proteins that are mostly localized to mitochondria and/or chloroplasts, where they are involved in RNA post-transcriptional processing [4] (Figure 1A). In general, the P-class PPR proteins mediate diverse aspects of RNA processing in plant organelles, while the PLS-class PPR proteins mainly function in RNA editing [5]. Mutations in these PPR protein-coding genes lead to the dysfunction of mitochondria and/or chloroplasts, thereby resulting in growth retardation, pollen abortion, and seed development defects in plants [4], indicating the important roles of PPR proteins in plant growth and development (Figure 1B).
2. Functions of PPR Proteins in Cytoplasmic Male Sterility
Cytoplasmic male sterility (CMS) is a maternally inherited trait that presents a defect in the production of viable pollen. CMS is widespread in higher plants, and has been widely used in the production of hybrid seeds and utilization of heterosis in many crop species [6,7]. Plant CMS is usually caused by mutations, rearrangements, or recombinations of mitochondrial DNA, and in many instances, male fertility can be restored specifically by restorer-of-fertility (Rf) genes in the nuclear genome [8,9]. To date, more than ten Rf genes have been cloned and functionally characterized in various crop species, and the majority of them were found to encode PPR protein, including Rf1a [10], Rf1b [11], Rf3 [12], Rf4 [13], Rf5 [14], Rf6 [15] in rice; Rfo [16], PPR-B [17], RsRf3-4 [18], Rfk1 [19] in radish; Rf1 [20], Rf2 [21] in sorghum; Rfp [22], Rfn [23] in rapeseed; Rf-PPR592 [24] in petunia; BrRfp1 [25] in Chinese cabbage; and Rfm1 [26] in barley. With the exception of the sorghum Rf1 [20] and the barely Rfm1 [26], the PPR-type Rf genes encode PPR proteins belonging to P class.
Members of P-class PPR proteins mostly function in various post-transcriptional process of organellar RNAs, such as RNA splicing, RNA stabilization, RNA cleavage, and translation [4,5]. Proteins encoded by Rf genes usually target mitochondria and act as fertility restorers by suppressing the production of mitochondrial CMS-inducing proteins [6,11]; however, the exact molecular mechanisms underlying fertility restoration by RF proteins is presently unclear. The PPR-type RF proteins have been proposed to rescue fertility by regulating the expression of CMS-conferring genes through the similar molecular mechanisms as that of other PPR proteins. In most of the CMS systems, PPR-type RF proteins bind to the mitochondrial CMS-conferring transcripts and promote their cleavage or degradation [27]. For example, rice RF1A and RF1B proteins, respectively encoded by Rf1a and Rf1b genes, have been considered to restore CMS by processing mitochondrial orf79 transcript via different mechanisms. RF1A directly binds to and cleaves the atp6-orf79 transcript at the intercistronic region, whereas RF1B promotes the rapid degradation of the atp6-orf79 transcript [10,11]. The protein encoded by rice Rf4 gene was reported to suppress WA352-mediated male sterility by reducing WA352 transcript levels [13]. Two rice RF proteins, Rfp and Rfn, are involved in transcript cleavage of orf224 and orf222, respectively [22,23].
In some CMS systems, PPR-type RF proteins restore the male fertility by impeding the translation or post-translational processing of mitochondrial CMS-inducing proteins. Studies on radish PPR-B revealed that PPR-B protein does not function through cleavage or degradation of the orf138 mRNA, but rather block its translation by inhibiting either the association with or the progression of mitochondrial ribosomes on the orf138 mRNA [17]. Similarly, rice RF3 protein does not affect the abundance of WA352 transcript but suppresses the accumulation of WA352 protein [12]. In addition, Rf1 from sorghum and Rfm1 from barely encode PPR proteins belonging to PLS class. As PLS-class PPR proteins almost exclusively play a role in RNA editing, it is possible that sorghum Rf1 and barely Rfm1 restore pollen fertility by editing S-orf transcripts or other target RNAs [8,26].
CMS was not only an ideal model system to study the interaction between mitochondrial and nuclear genomes but also a useful genetic tool for breeding to exploit hybrid vigor in crops [27]. Although PPR proteins are involved in the restoration of male fertility, functions of most PPR proteins are still obscure. Therefore, exploring the functions of PPR proteins will contribute to understanding the CMS mechanism and improving molecular breeding in crops.
3. Functions of PPR Proteins in Plant Responses to Biotic and Abiotic Stresses
In previous studies, PPR genes were found to change their expression patterns under biotic and abiotic stresses and regulate growth in many plants. For instance, in Arabidopsis, 11 PPR proteins have been shown to respond to biotic or abiotic stresses. SOAR1, a cytosol-nucleus dual-localized PPR protein, is involved in ABA signaling and tolerance to drought, salt, and cold stress [28]. GUN1 is a chloroplast-located PPR protein, the gun1 mutant is defective in response to norflurazon, lincomycin, and high-light treatments [29], and it also exhibits a more susceptible phenotype to photooxidative stress caused during the de-etiolation [30]. In addition, nine mitochondria-located PPR proteins, MED11/LOI1 [31], PPR40 [32], ABO5 [33], PGN [34], AHG11 [35], SLG1 [36], SLO2 [37], PPR96 [38], and POCO1 [39] were reported to participate in responses to many abiotic or biotic stresses. In rice, two chloroplast-located PPR proteins, OsV4 and TCD10, are required for chloroplast development at early seedling stage under cold stress [40,41]. The chloroplast-located PPR protein WSL affects chloroplast development and abiotic stress response in rice, and the wsl mutant displays chlorotic striations early in development and enhanced sensitivity to ABA, salt, and sugar [42]. Recently, a mitochondrial PPR protein OsNBL3 was found to be involved in response to biotic or abiotic stresses. The nbl3 mutant exhibits growth retardation, leaf wilting, and premature senescence, and it shows enhanced resistance against fungal and bacterial pathogens and to salt stress [43].
A few of the PPR proteins involved in plant responses to biotic and abiotic stresses have been shown to play roles in post-transcriptional processing of RNA in mitochondria and chloroplasts. In Arabidopsis, PGN participates in editing of mitochondrial cox2 and nad6 [34]. SLG1 and AHG11 are involved in editing of mitochondrial nad3 and nad4, respectively [35,36]. SLO2 was found to be required for mitochondrial RNA editing at multiple sites [37]. In rice, WSL functions in the splicing of chloroplast rpl2 [42]. OsNBL3 takes part in the splicing of mitochondrial nad5 [43]. Although POCO1 encodes a mitochondria-localized P-class PPR protein, multiple RNA editing defects were identified in poco1 mutant, which suggest that POCO1 may be required for RNA editing [39].
Under normal and stress conditions, retrograde signaling from organelles plays vital roles in coordinating the expression of nuclear and organellar genes, and in regulating plant growth and development. Arabidopsis GUN1 is identified to be a central integrator of chloroplast retrograde signaling pathways. In 2016, Tadini et al. revealed that GUN1 controls accumulation of the chloroplast ribosomal protein S1 at the protein level and interacts with proteins involved in chloroplast protein homeostasis [44]. MORF2 is an essential component of RNA editosome and is required for editing at almost all sites in chloroplast RNAs [45]. Unlike typical PPR proteins, GUN1 does not appear to bind to RNAs. However recently, GUN1 was found to physically interact with MORF2 to regulate the editing efficiency of multiple sites within chloroplast RNAs and modulate the activity of the nucleus-encoded chloroplast RNA polymerase, particularly during retrograde signaling [46,47], indicating that GUN1 is important for chloroplast RNA metabolism and chloroplast-to-nucleus retrograde communication. The mitochondrial PPR protein LOI1 of Arabidopsis was reported to be involved in RNA editing of mitochondrial transcripts cox3, nad4, and ccb203, and regulate biosynthesis of isoprenoids, metabolites known to affect defense gene expression in response to wounding and pathogen infection [31]. The loi1 mutant has decreased sensitivity to two inhibitors of isoprenoid synthesis, fungal phytotoxin lovastatin, and herbicide clomazone, showing the indirect effects of retrograde signaling from mitochondria to the cytoplasm to evoke alteration of the mevalonate pathway [48]. Therefore, these results reveal a putative link between organellar RNA processing and plant responses to environmental stresses.
4. Functions of PPR Proteins in Seed Development
In angiosperm plants, seed development starts with double fertilization of egg and central cells with two sperm cells, which leads to the formation of a diploid embryo and a triploid endosperm, and develops into mature seeds comprising three structures: maternal coat, embryo, and endosperm [49]. Development of embryo and endosperm is well correlated and regulated by numerous distinct proteins involved in many important physiological processes [50], including cell growth, RNA transcription and post-transcriptional processing, etc.
In recent years, more and more genetic and biochemical studies have shown that PPR proteins play important roles in seed development of higher plants, and loss-of-function of these PPR proteins usually leads to defects in embryogenesis and/or endosperm development [4,51]. According to the phenotypic expression, seed mutants can be divided into four major classes: empty pericarp (emp), embryo specific (emb), defective kernel (dek), and small kernel (smk). A detailed summary of the maize and Arabidopsis seed mutants caused by the functional defects of PPR proteins is provided in Table 1.
Most PPR proteins identified to date are targeted to mitochondria and/or chloroplasts [4]. The disruption of PPR proteins localized to chloroplasts usually results in the emb mutants that are defective in embryogenesis, but relatively normal in endosperm development. For instance, PPR8522 [94] and EMB-7L [95] in maize and GRP23 [103] in Arabidopsis are necessary for embryogenesis, and their mutations lead to arrested embryo development at the transition stage, resulting in an embryo-lethal phenotype. For mitochondrion-targeted PPR proteins, their disruptions mostly cause diverse seed development mutants, including smk, dek, and emp, with different degrees of defects in embryo and endosperm. The loss-of-function of SMK1 [65], SMK4 [66], SMK6 [67], ZmSMK9 [68], PPR2263 [69], and MPPR6 [70] in maize and PPR19 [97] in Arabidopsis arrests both embryo and endosperm development, resulting in smk phenotypes. Some characterized mitochondrion-targeted PPR proteins, such as DEK2 [52], DEK10 [53], DEK35 [54], DEK36 [55], DEK37 [56], DEK39 [57], DEK40 [58], DEK41/43 [59,60], DEK46 [61], and DEK53 [62] in maize, are necessary for seed development, and their disruptions result in dek mutants with arrested development of both the embryo and endosperm at an early stage. Meanwhile, many PPR proteins are targeted to mitochondria and function in development of both embryo and endosperm, and mutations in their encoding genes arrest embryo and endosperm development at early stages and even result in embryo lethality. For example, Emp5 encodes a mitochondrion-targeted DYW-type PPR protein, the emp5 mutants exhibit abortion of embryo and endosperm development at early stages in maize [74]. Loss-of-function of the mitochondrial P-type PPR protein EMP10 severely disturbs embryo and endosperm development, resulting in empty pericarp or papery seeds in maize [78]. Additionally, the P-type protein PPR5 was recently identified as a regulator required for endosperm development in rice, the ppr5 mutants develop small starch grains [106].
5. Mechanisms of PPR Protein-Mediated RNA Processing in Plant Organelles
Mostly, above-mentioned functions of PPR proteins in plant growth, development and stress responses are caused by the disturbed organellar gene expression pattern [107]. As a large family of RNA-binding proteins, some PPR proteins have shown to be required for post-transcriptional RNA processing events in plant mitochondria and chloroplasts [4]. These observations lead to an increasing interest towards understanding the nature and details of PPR protein-mediated RNA processing, and some progress has been made in the last decade [108], in particular, the mechanisms of PPR proteins involved in C-to-U RNA editing and in group II intron splicing.
5.1. Molecular Mechanisms of PPR Proteins in C-to-U RNA Editing
C-to-U RNA editing is highly prevalent in plant organelles, which converts cytidines to uridines at specific sites in transcripts of chloroplasts and plant mitochondria. The chemical nature of C-to-U RNA editing is through a site-specific cytidine deamination reaction [109]. To date, many E/E+ or DYW-subclass PPR proteins have been characterized as C-to-U RNA editing factors for one, several or multiple editing sites in plant organelles [84,85]. The DYW domain of some PPR proteins contains a conserved zinc-binding motif (HxE(x)nCxxC) in common with cytidine deaminase [110], and thus it has been proposed as the best candidate to catalyze deamination. Experimental support for this hypothesis came from mutagenesis experiments that modified the deaminase signature sequence. Mutation of several residues in cytidine deaminase signature of DYW1 significantly decreases the zinc-binding capacity and abolishes the editing of ndhD-1 [111]. The glutamate residue is required for the cytidine deaminase-catalyzed reactions. Mutagenesis of glutamate residue in the zinc-binding motif of OTP84 and CREF7 leads to loss of their editing activity at their cognate editing sites [112]. Recently, the DYW-PPR proteins called PPR56 and PPR65 from Physcomitrella patens were shown to drive C-to-U RNA editing individually at their corresponding targets in E. coli [113], and the purified recombinant PPR65 was found to exhibit the function of editing at its target site in synthetic RNAs [114], indicating a single DYW-deaminase domain-containing PPR protein is sufficient for editing its defined RNA targets within the bacterial cells and in vitro. In 2021, Takenaka et al. analyzed the structures of a DYW domain, they found that it contains a cytidine deaminase domain and a characteristic DYW motif, with catalytic and structural zinc atoms, respectively. Moreover, the DYW motif is stabilized by a zinc atom and functions in the control of zinc-mediated catalysis by altering the coordination setting around the catalytic zinc atom [115]. These results reinforce the hypothesis that the DYW domain functions as cytidine deaminase that carries out C-to-U RNA editing in plant organelles.
E and E+-type PPR proteins lack the DYW domain that contributes to the catalytic reaction of C-to-U RNA editing, and their exact function in the RNA-editing process remains unclear. Some studies rather suggest that they can form protein complexes with a DYW-type protein for a complete editing event. The first of these PPR editing factors to be identified is an E-type PPR protein CRR2. To edit chloroplast ndhD-2 site, CRR2 physically interacts with DYW1 that carries very few PPR motifs, no canonical E and E+ domains but end with a DYW domain [116]. Where, in which CRR2 acts as an editing site recognition factor, while DYW1 has key features of an editing deaminase [111]. DYW2 is a DYW-type PPR protein recently shown to be required for the editing of more than 100 sites in mitochondria and chloroplasts [117], and almost all of the DYW2 dependent editing sites are targets of E+-type PPR proteins, such as PGN, MEF37, OTP90 [118], etc. The functional association and physical interactions of DYW2 and E+-type PPR proteins suggest that DYW2 complements missing DYW domains in E+-type PPR proteins [117,118,119].
C-to-U RNA editing in plant organelles is performed by an editosome composed of RNA and protein factors. In addition to PPR proteins, several other protein factors were shown to be part of RNA editosome, and some of these protein factors were found to physically interact with each other or with PPR proteins. Previous studies indicate that the multiple organellar RNA editing factors (MORFs) can interact with the PLS-class PPR proteins and participate in RNA editing. MORF8 interacts with the DYW-type proteins RARE1 and MEF10 to participate in the C-to-U RNA editing in Arabidopsis chloroplast accD and mitochondrial nad2 transcripts, respectively [120,121]. ZmMORF8 interacts with DYW-type proteins EMP5 and EMP21, and are together required for editing at multiple sites in maize mitochondria [85]. In addition, other MORF proteins have been shown to be involved in C-to-U RNA editing in plant mitochondria or chloroplasts, and physically interact with PLS-class PPR proteins [91,122]. RNA binding activity has been reported for PPR proteins, and a PPR-RNA recognition code has been elucidated and experimentally validated [123]. The PLS-class PPR proteins recognize and bind to RNA sequences upstream of the edited cytidine residues, whereas the MORF proteins mediate their RNA-binding activity [124]. For instance, MORF9 binding induced significant compressed conformational changes of PLS-class PPR protein, leading to the increase in the RNA-binding activity of PPR proteins [125].
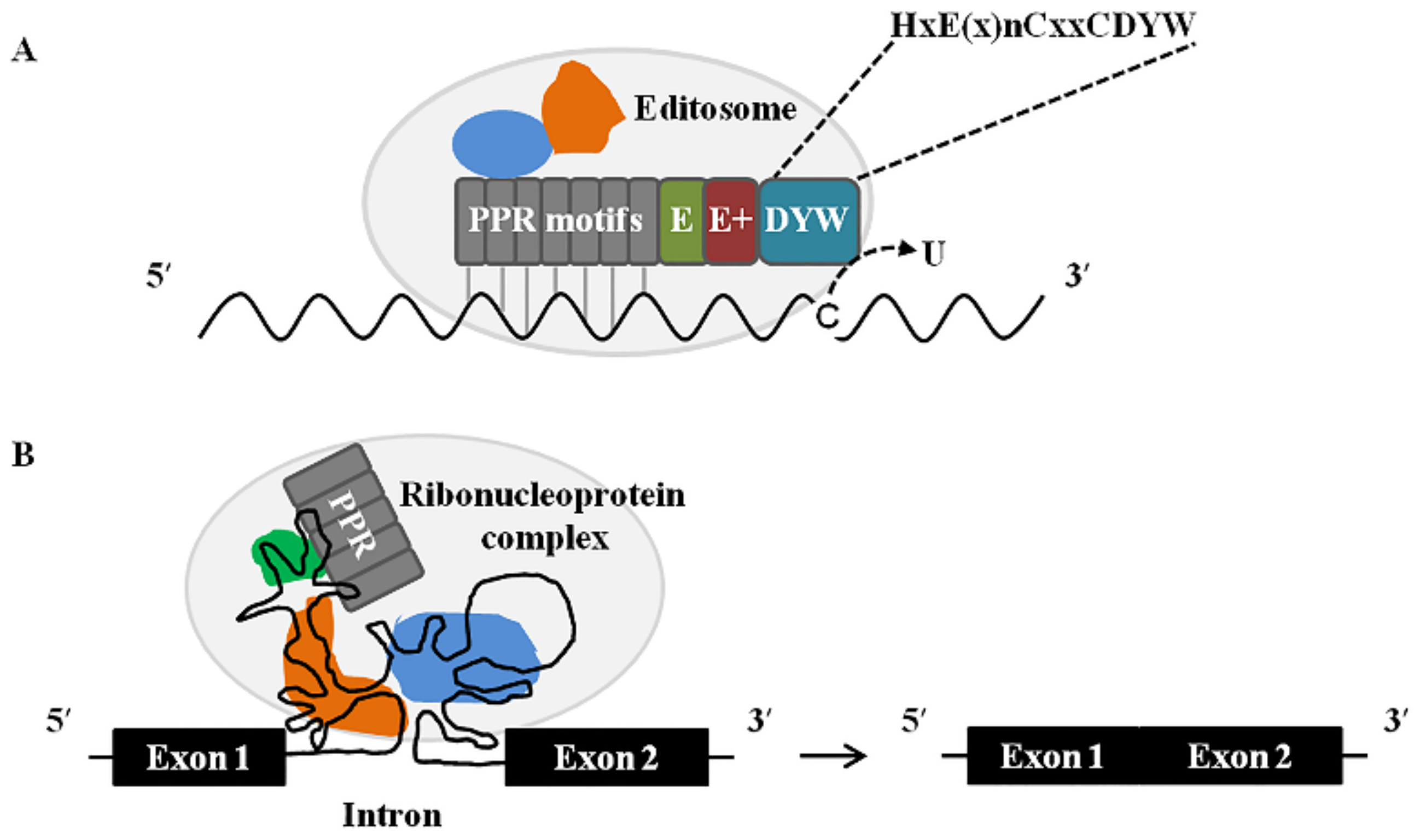
These data indicate that PPR proteins serve as the primary recognition factors that single out specific cytidines to be converted into uridines in organellar transcripts of higher plants. These data also highlight that the DYW domain is the likely cytidine deaminase performing the cytidine deamination reaction (Figure 2A).
5.2. Molecular Mechanisms of PPR Proteins in Group II Intron Splicing
Group II introns commonly exist in organellar genomes of land plants, with more than 20 in the mitochondrial and chloroplast genomes of Arabidopsis, rice, and maize. Because of the loss of the ability to undergo self-splicing, the splicing of organellar group II introns in higher plants relies on various protein cofactors coming from different families and encoded by nuclear and organellar genomes [126,127]. So far, some P-class PPR proteins have been shown to be required for the splicing of plant mitochondrial and chloroplast introns [4], such as ABO5, MTL1, TANG2, OTP439, OTP43, and SLO3 in Arabidopsis [33,96,128,129,130], DEK2, DEK35, DEK37, EMP8, EMP10, EMP16, PPR14, EMP24, EMP25, and PPR-SMR1 in maize [52,54,56,76,78,82,89,93,131], as well as OsNPPR1, OsNPPR3, FLO10, and PPR5 in rice [106,132,133,134]. These PPR proteins are involved in the splicing of one, several or multiple introns in plant mitochondria or chloroplasts. For example, DEK37 [56] in maize, MID1 [104] in Arabidopsis, and FLO10 [133] in rice are specifically involved in the splicing of a single intron in mitochondria or chloroplasts. EMP8 [76] and PPR14 [89] participate in the splicing of several mitochondrial introns in maize, while PPR-SMR1 in maize is required for the splicing of nearly 75% of the mitochondrial introns [93]. Besides, some PLS-class PPR proteins that are usually characterized to function in RNA editing, are also implicated in RNA splicing. SLO4 was reported to affect nad2 intron 1 splicing in Arabidopsis [135]. PpPPR43 influences the splicing efficiency of cox1 intron 3 in Physcomitrella patens [136].
Although some PPR proteins acted as splicing cofactors in plant organelles have been identified, how these PPR proteins function in the splicing of plant organellar group II introns is not well understood. The recruitment of PPR proteins and other splicing factors is generally required for the splicing of a group II intron in plant organelles. In chloroplasts, the PPR proteins PPR4 and EMB2654 [101,137,138], the CRM (chloroplast RNA splicing and ribosome maturation) protein CAF2 [139], the PTH (peptidyl-tRNA hydrolase) protein CRS2 [140], and the DEAD-box RNA helicase protein RH3 [141] are required for the removal of intron 1 from rps12 pre-mRNA, and they are all found in a large ribonucleoprotein complex. Similarly, in mitochondria, three PPR proteins, MISF68 [142], ZmSMK9 [68], and PPR-SMR1 [93], and a ribosomal protein uL18-L1 [143] participate in the splicing of nad5 intron 1. In addition, the PPR protein PPR14 was reported to physically interact with a PPR protein PPR-SMR1 and a CRM-domain protein Zm-mCSF1, and they are required for the splicing of mitochondrial nad2 intron 3 in maize [89,93]. As members of RNA-binding protein families, several P-class PPR proteins have been found to bind to their dependent splicing introns. For example, the chloroplast-localized THA8 is required for the splicing of ycf3 intron 2 and trnA intron in maize [144], and its orthologues from Brachypodium distachyon can bind to ycf3 intron 2 in vitro through multiple purine-rich sequences distributed in intron [145]. PPR4 and EMB2654 are known to be involved in the splicing of the chloroplast rps12 intron 1 in maize and Arabidopsis, respectively [101,146]. RNA footprinting and EMSA experiments indicated that the Arabidopsis ortholog of PPR4 binds to a sequence near the 5′ end of rps12 intron 1, while EMB2654 binds to a sequence near the 3′ end of rps12 intron 1 [101,146]. By associating with their target intron, these PPR proteins and other splicing factors could form ribonucleoprotein complexes that help intron folding and stabilize their structure in an active form [146,147].
Taken together, these findings strongly suggest that PPR proteins participate in the splicing of group II intron in plant organelles might by binding to a specific RNA sequence in their target intron and interacting with other splicing factors to form a large complex that is required for the splicing performed (Figure 2B), but how these proteins collaborate or whether they act independently remains to be elucidated.
6. Conclusions and Perspectives
Although there is more evidence indicating that PPR proteins specifically take part in multiple aspects of RNA processing in plant organelles [4], only a few PPR proteins have been functionally characterized in contrast to the vast majority of PPR members. Moreover, computational and structural studies have provided new insights into RNA recognition and cytidine deamination reaction by PPR proteins in C-to-U RNA editing events [115,123,148]. However, little is known about the mechanistic details of PPR proteins involved in other RNA processing events. Therefore, there is a need to promote a more comprehensive understanding of PPR proteins. Firstly, why are PPR proteins particularly prevalent in land plants, and display diverse functions in organellar RNA processing? Secondly, how do PPR proteins interact with each other or other proteins to achieve their final functionality, and how do these interactions contribute to PPR activities in organellar RNA processing? Finally, what roles do PPR proteins play in the regulatory network of nuclear-cytoplasmic interaction in land plants? Maybe plenty of evolutionary analyses will provide deeper insight into their subtle classification and distribution. Functional identification of other PPR proteins and further analysis of co-expressed PPR genes will reveal whether they also play roles in organellar RNA processing or whether they are involved in other steps of RNA expression in plant organelles, and will allow us to define the regulatory network of nuclear-cytoplasmic interaction in land plants. Experimental identification of RNA targets of PPR proteins and crystal structure analyses of protein–RNA complexes are however necessary to elucidate the action mode of PPR proteins in detail. Besides, more attention should be paid to PPR interacting proteins that could directly affect the PPR protein effects, such as MORF proteins [47]. These studies will help to better clarify the mechanisms of PPR proteins, and also help to understand the details of plant organellar RNA processing.
Author Contributions
X.L. and Y.J. designed the outlines in this review and edited the manuscript. X.L. wrote the original manuscript. M.S., S.L. (Shijuan Liu), Q.T. and S.L. (Shihui Li) participated in the collection and collation of literatures. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (32070246), Qufu Normal University Doctoral Fund Project (BSQD20152493), and Shandong Province Higher Educational Science and Technology Program (J16LE09).
Conflicts of Interest
The authors declare that there are no conflicts of interest.
References
- Small, I.D.; Peeters, N. The PPR motif-a TPR-related motif prevalent in plant organellar proteins. Trends Biochem. Sci. 2000, 25, 45–47. [Google Scholar] [CrossRef]
- Lurin, C.; Andrés, C.; Aubourg, S.; Bellaoui, M.; Bitton, F.; Bruyère, C.; Caboche, M.; Debast, C.; Gualberto, J.; Hoffmann, B.; et al. Genome-wide analysis of Arabidopsis pentatricopeptide repeat proteins reveals their essential role in organelle biogenesis. Plant Cell 2004, 16, 2089–2103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, S.F.; Gutmann, B.; Zhong, X.; Ye, Y.T.; Fisher, M.F.; Bai, F.Q.; Castleden, I.; Song, Y.; Song, B.; Huang, J.Y.; et al. Redefining the structural motifs that determine RNA binding and RNA editing by pentatricopeptide repeat proteins in land plants. Plant J. 2016, 85, 532–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barkan, A.; Small, I. Pentatricopeptide repeat proteins in plants. Annu. Rev. Plant Biol. 2014, 65, 415–442. [Google Scholar] [CrossRef] [PubMed]
- Schmitz-Linneweber, C.; Small, I. Pentatricopeptide repeat proteins: A socket set for organelle gene expression. Trends Plant Sci. 2008, 13, 663–670. [Google Scholar] [CrossRef]
- Chase, C.D. Cytoplasmic male sterility: A window to the world of plant mitochondrial-nuclear interactions. Trends Genet. 2007, 23, 81–90. [Google Scholar] [CrossRef]
- Bohra, A.; Jha, U.C.; Adhimoolam, P.; Bisht, D.; Singh, N.P. Cytoplasmic male sterility (CMS) in hybrid breeding in field crops. Plant Cell Rep. 2016, 35, 967–993. [Google Scholar] [CrossRef]
- Linke, B.; Börner, T. Mitochondrial effects on flower and pollen development. Mitochondrion 2005, 5, 389–402. [Google Scholar] [CrossRef] [PubMed]
- Eckardt, N.A. Cytoplasmic male sterility and fertility restoration. Plant Cell 2006, 18, 515–517. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.H.; Zou, Y.J.; Li, X.Y.; Zhang, Q.Y.; Chen, L.T.; Wu, H.; Su, D.H.; Chen, Y.L.; Guo, J.X.; Luo, D.; et al. Cytoplasmic male sterility of rice with Boro II cytoplasm is caused by a cytotoxic peptide and is restored by two related PPR motif genes via distinct modes of mRNA silencing. Plant Cell 2006, 18, 676–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazama, T.; Nakamura, T.; Watanabe, M.; Sugita, M.; Toriyama, K. Suppression mechanism of mitochondrial ORF79 accumulation by Rf1 protein in BT-type cytoplasmic male sterile rice. Plant J. 2008, 55, 619–628. [Google Scholar] [CrossRef]
- Luo, D.P.; Xu, H.; Liu, Z.L.; Guo, J.X.; Li, H.Y.; Chen, L.T.; Fang, C.; Zhang, Q.Y.; Bai, M.; Yao, N.; et al. A detrimental mitochondrial-nuclear interaction causes cytoplasmic male sterility in rice. Nat. Genet. 2013, 45, 573–577. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Luo, D.; Zhou, D.; Zhang, Q.; Tian, D.; Zheng, X.; Chen, L.; Liu, Y.G. The rice restorer Rf4 for wild-abortive cytoplasmic male sterility encodes a mitochondrial-localized PPR protein that functions in reduction of WA352 transcripts. Mol. Plant 2014, 7, 1497–1500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.; Wang, K.; Huang, W.C.; Liu, G.; Gao, Y.; Wang, J.M.; Huang, Q.; Ji, Y.X.; Qin, X.J.; Wan, L.; et al. The rice pentatricopeptide repeat protein RF5 restores fertility in Hong-Lian cytoplasmic male-sterile lines via a complex with the glycine-rich protein GRP162. Plant Cell 2012, 24, 109–122. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.C.; Yu, C.C.; Hu, J.; Wang, L.L.; Dan, Z.W.; Zhou, W.; He, C.L.; Zeng, Y.F.; Yao, G.X.; Qi, J.Z.; et al. Pentatricopeptide-repeat family protein RF6 functions with hexokinase 6 to rescue rice cytoplasmic male sterility. Proc. Natl. Acad. Sci. USA 2015, 112, 14984–14989. [Google Scholar] [CrossRef] [Green Version]
- Brown, G.G.; Formanová, N.; Jin, H.; Wargachuk, R.; Dendy, C.; Patil, P.; Laforest, M.; Zhang, J.F.; Cheung, W.Y.; Landry, B.S. The radish Rfo restorer gene of Ogura cytoplasmic male sterility encodes a protein with multiple pentatricopeptide repeats. Plant J. 2003, 35, 262–272. [Google Scholar] [CrossRef]
- Uyttewaal, M.; Arnal, N.; Quadrado, M.; Martin-Canadell, A.; Vrielynck, N.; Hiard, S.; Gherbi, H.; Bendahmane, A.; Budar, F.; Mireau, H. Characterization of Raphanus sativus pentatricopeptide repeat proteins encoded by the fertility restorer locus for Ogura cytoplasmic male sterility. Plant Cell 2008, 20, 3331–3345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.W.; De Wang, C.D.; Mei, S.Y.; Gao, L.; Zhou, Y.; Wang, T. An insertion-deletion at a pentatricopeptide repeat locus linked to fertility transition to cytoplasmic male sterility in radish (Raphanus sativus L.). Mol. Breed. 2015, 35, 108–112. [Google Scholar] [CrossRef]
- Imai, R.; Koizuka, N.; Fujimoto, H.; Hayakawa, T.; Sakai, T.; Imamura, J. Delimitation of the fertility restorer locus Rfk1 to a 43-kb contig in Kosena radish (Raphanus sativus L.). Mol. Genet. Genom. 2003, 269, 388–394. [Google Scholar] [CrossRef]
- Klein, R.R.; Klein, P.E.; Mullet, J.E.; Minx, P.; Rooney, W.L.; Schertz, K.F. Fertility restorer locus Rf1 of sorghum (Sorghum bicolor L.) encodes a pentatricopeptide repeat protein not present in the colinear region of rice chromosome 12. Theor. Appl. Genet. 2005, 111, 994–1012. [Google Scholar] [CrossRef]
- Madugula, P.; Uttam, A.G.; Tonapi, V.A.; Ragimasalawada, M. Fine mapping of Rf2, a major locus controlling pollen fertility restoration in sorghum A1 cytoplasm, encodes a PPR gene and its validation through expression analysis. Plant Breed. 2018, 137, 148–161. [Google Scholar] [CrossRef]
- Liu, Z.; Yang, Z.; Wang, X.; Li, K.; An, H.; Liu, J.; Yang, G.S.; Fu, T.; Yi, B.; Hong, D.F. A mitochondria-targeted PPR protein restores pol cytoplasmic male sterility by reducing orf224 transcript levels in oilseed rape. Mol. Plant 2016, 9, 1082–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Dong, F.M.; Wang, X.; Wang, T.; Su, R.; Hong, D.F.; Yang, G.S. A pentatricopeptide repeat protein restores nap cytoplasmic male sterility in Brassica napus. J. Exp. Bot. 2017, 68, 4115–4123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillman, J.D.; Bentolila, S.; Hanson, M.R. The petunia restorer of fertility protein is part of a large mitochondrial complex that interacts with transcripts of the CMS-associated locus. Plant J. 2007, 49, 217–227. [Google Scholar] [CrossRef]
- Zhang, H.M.; Wu, J.Q.; Dai, Z.H.; Qin, M.L.; Hao, L.Y.; Ren, Y.J.; Li, Q.F.; Zhang, L.G. Allelism analysis of BrRfp locus in different restorer lines and map-based cloning of a fertility restorer gene, BrRfp1, for pol CMS in Chinese cabbage (Brassica rapa L.). Theor. Appl. Genet. 2017, 130, 539–547. [Google Scholar] [CrossRef]
- Rizzolatti, C.; Bury, P.; Tatara, E.; Pin, P.A.; Rodde, N.; Bergès, H.; Budar, F.; Mireau, H.; Gielen, J.J.L. Map-based cloning of the fertility restoration locus Rfm1 in cultivated barley (Hordeum vulgare). Euphytica 2017, 213, 276–287. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.T.; Liu, Y.G. Male sterility and fertility restoration in crops. Annu. Rev. Plant Biol. 2014, 65, 579–606. [Google Scholar] [CrossRef]
- Jiang, S.C.; Mei, C.; Liang, S.; Yu, Y.T.; Lu, K.; Wu, Z.; Wang, X.F.; Zhang, D.P. Crucial roles of the pentatricopeptide repeat protein SOAR1 in Arabidopsis response to drought, salt and cold stresses. Plant Mol. Biol. 2015, 88, 369–385. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.Z.; Chalvin, C.; Hoelscher, M.; Meyer, E.H.; Wu, X.N.; Bock, R. Control of retrograde signaling by rapid turnover of GENOMES UNCOUPLED1. Plant Physiol. 2018, 176, 2472–2495. [Google Scholar] [CrossRef] [Green Version]
- Mochizuki, N.; Susek, R.; Chory, J. An intracellular signal transduction pathway between the chloroplast and nucleus is involved in de-etiolation. Plant Physiol. 1996, 112, 1465–1469. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, K.; Suzuki, M.; Tang, J.W.; Nagata, N.; Ohyama, K.; Seki, H.; Kiuchi, R.; Kaneko, Y.; Nakazawa, M.; Matsui, M.; et al. LOVASTATIN INSENSITIVE 1, a novel pentatricopeptide repeat protein, is a potential regulatory factor of isoprenoid biosynthesis in Arabidopsis. Plant Cell Physiol. 2007, 48, 322–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zsigmond, L.; Rigoó, G.; Szarka, A.; Szeékely, G.; Ötvös, K.; Darula, Z.; Medzihradszky, K.F.; Koncz, C.; Koncz, Z.; Szabados, L. Arabidopsis PPR40 connects abiotic stress responses to mitochondrial electron transport. Plant Physiol. 2008, 146, 1721–1737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; He, J.N.; Chen, Z.Z.; Ren, X.Z.; Hong, X.H.; Gong, Z.Z. ABA overly-sensitive 5 (ABO5), encoding a pentatricopeptide repeat protein required for cis-splicing of mitochondrial nad2 intron 3, is involved in the abscisic acid response in Arabidopsis. Plant J. 2010, 63, 749–765. [Google Scholar] [CrossRef]
- Laluk, K.; AbuQamar, S.; Mengiste, T. The Arabidopsis mitochondria-localized pentatricopeptide repeat protein PGN functions in defense against necrotrophic fungi and abiotic stress tolerance. Plant Physiol. 2011, 156, 2053–2068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murayama, M.; Hayashi, S.; Nishimura, N.; Ishide, M.; Kobayashi, K.; Yagi, Y.; Asami, T.; Nakamura, T.; Shinozaki, K.; Hirayama, T. Isolation of Arabidopsis ahg11, a weak ABA hypersensitive mutant defective in nad4 RNA editing. J. Exp. Bot. 2012, 63, 5301–5310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, H.; Liu, D. Functional disruption of the pentatricopeptide protein SLG1 affects mitochondrial RNA editing, plant development, and responses to abiotic stresses in Arabidopsis. Plant J. 2012, 70, 432–444. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Dugardeyn, J.; Zhang, C.Y.; Mühlenbock, P.; Eastmond, P.J.; Valcke, R.; De Coninck, B.; Öden, S.; Karampelias, M.; Cammue, B.P.A.; et al. The Arabidopsis thaliana RNA editing factor SLO2, which affects the mitochondrial electron transport chain, participates in multiple stress and hormone responses. Mol. Plant 2014, 7, 290–310. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.M.; Zhao, J.Y.; Lu, P.P.; Chen, M.; Guo, C.H.; Xu, Z.S.; Ma, Y.Z. The E-subgroup pentatricopeptide repeat protein family in Arabidopsis thaliana and confirmation of the responsiveness PPR96 to abiotic stresses. Front. Plant Sci. 2016, 7, 1825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emami, H.; Kumar, A.; Kempken, F. Transcriptomic analysis of poco1, a mitochondrial pentatricopeptide repeat protein mutant in Arabidopsis thaliana. BMC Plant Biol. 2020, 20, 209. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.D.; Su, Q.Q.; Lin, D.Z.; Jiang, Q.; Xu, J.L.; Zhang, J.H.; Teng, S.; Dong, Y.J. The rice OsV4 encoding a novel pentatricopeptide repeat protein is required for chloroplast development during the early leaf stage under cold stress. J. Integr. Plant Biol. 2014, 56, 400–410. [Google Scholar] [CrossRef]
- Wu, L.L.; Wu, J.; Liu, Y.X.; Gong, X.D.; Xu, J.L.; Lin, D.Z.; Dong, Y.J. The rice pentatricopeptide repeat gene TCD10 is needed for chloroplast development under cold stress. Rice 2016, 9, 67–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, J.J.; Tan, Z.H.; Wu, F.Q.; Sheng, P.K.; Heng, Y.Q.; Wang, X.H.; Ren, Y.L.; Wang, J.L.; Guo, X.P.; Zhang, X.; et al. A novel chloroplast-localized pentatricopeptide repeat protein involved in splicing affects chloroplast development and abiotic stress response in rice. Mol. Plant 2014, 7, 1329–1349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, T.C.; Zhao, X.S.; Feng, H.J.; Qi, L.L.; Yang, J.; Peng, Y.L.; Zhao, W.S. OsNBL3, a mitochondrion-localized pentatricopeptide repeat protein, is involved in splicing nad5 intron 4 and its disruption causes lesion mimic phenotype with enhanced resistance to biotic and abiotic stresses. Plant Biotechnol. J. 2021. [Google Scholar] [CrossRef] [PubMed]
- Tadini, L.; Pesaresi, P.; Kleine, T.; Rossi, F.; Guljamow, A.; Sommer, F.; Mühlhaus, T.; Schroda, M.; Masiero, S.; Pribil, M.; et al. GUN1 controls accumulation of the plastid ribosomal protein S1 at the protein level and interacts with proteins involved in plastid protein homeostasis. Plant Physiol. 2016, 170, 1817–1830. [Google Scholar] [CrossRef] [Green Version]
- Takenaka, M.; Zehrmann, A.; Verbitskiy, D.; Kugelmann, M.; Hartel, B.; Brennicke, A. Multiple organellar RNA editing factor (MORF) family proteins are required for RNA editing in mitochondria and plastids of plants. Proc. Natl. Acad. Sci. USA 2012, 109, 5104–5109. [Google Scholar] [CrossRef] [Green Version]
- Tadini, L.; Peracchio, C.; Trotta, A.; Colombo, M.; Mancini, I.; Jeran, N.; Costa, A.; Faoro, F.; Marsoni, M.; Vannini, C.; et al. GUN1 influences the accumulation of NEP-dependent transcripts and chloroplast protein import in Arabidopsis cotyledons upon perturbation of chloroplast protein homeostasis. Plant J. 2020, 101, 1198–1220. [Google Scholar] [CrossRef]
- Zhao, X.B.; Huang, J.Y.; Chory, J. GUN1 interacts with MORF2 to regulate plastid RNA editing during retrograde signaling. Proc. Natl. Acad. Sci. USA 2019, 116, 10162–10167. [Google Scholar] [CrossRef] [Green Version]
- Verbitskiy, D.; Zehrmann, A.; van der Merwe, J.A.; Brennicke, A.; Takenaka, M. The PPR protein encoded by the LOVASTATIN INSENSITIVE1 gene is involved in RNA editing at three sites in mitochondria of Arabidopsis thaliana. Plant J. 2010, 61, 446–455. [Google Scholar] [CrossRef]
- Goldberg, R.B.; de Paiva, G.; Yadegari, R. Plant embryogenesis: Zygote to seed. Science 1994, 266, 605–614. [Google Scholar] [CrossRef]
- Linkies, A.; Graeber, K.; Knight, C.; Leubner-Metzger, G. The evolution of seeds. New Phytol. 2010, 186, 817–831. [Google Scholar] [CrossRef]
- Dai, D.W.; Ma, Z.Y.; Song, R.T. Maize kernel development. Mol. Breed. 2021, 41, 2. [Google Scholar] [CrossRef]
- Qi, W.W.; Yang, Y.; Feng, X.Z.; Zhang, M.L.; Song, R.T. Mitochondrial function and maize kernel development requires Dek2, a pentatricopeptide repeat protein involved in nad1 mRNA splicing. Genetics 2017, 205, 239–249. [Google Scholar] [CrossRef] [Green Version]
- Qi, W.W.; Tian, Z.R.; Lu, L.; Chen, X.Z.; Chen, X.Z.; Zhang, W.; Song, R. Editing of mitochondrial transcripts nad3 and cox2 by Dek10 is essential for mitochondrial function and maize plant development. Genetics 2017, 205, 1489–1501. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.Z.; Feng, F.; Qi, W.W.; Xu, L.M.; Yao, D.S.; Wang, Q.; Song, R.T. Dek35 encodes a PPR protein that affects cis-splicing of mitochondrial nad4 intron 1 and seed development in maize. Mol. Plant 2017, 10, 427–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.; Zhong, M.Y.; Shuai, B.L.; Song, J.D.; Zhang, J.; Han, L.; Ling, H.L.; Tang, Y.P.; Wang, G.F.; Song, R.T. E+ subgroup PPR protein defective kernel 36 is required for multiple mitochondrial transcripts editing and seed development in maize and Arabidopsis. New Phytol. 2017, 214, 1563–1578. [Google Scholar] [CrossRef] [Green Version]
- Dai, D.W.; Luan, S.C.; Chen, X.Z.; Wang, Q.; Feng, Y.; Zhu, C.G.; Qi, W.W.; Song, R.T. Maize Dek37 encodes a P-type PPR protein that affects cis-splicing of mitochondrial nad2 intron 1 and seed development. Genetics 2018, 208, 1069–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.J.; Gu, W.; Sun, S.L.; Chen, Z.L.; Chen, J.; Song, W.B.; Zhao, H.M.; Lai, J.S. Defective Kernel 39 encodes a PPR protein required for seed development in maize. J. Integr. Plant Biol. 2018, 60, 45–64. [Google Scholar] [CrossRef] [Green Version]
- Ren, R.C.; Lu, X.D.; Zhao, Y.J.; Wei, Y.M.; Wang, L.L.; Zhang, L.; Zhang, W.T.; Zhang, C.Y.; Zhang, X.S.; Zhao, X.Y. Pentatricopeptide repeat protein DEK40 is required for mitochondrial function and kernel development in maize. J. Exp. Bot. 2019, 70, 6163–6179. [Google Scholar] [CrossRef]
- Ren, R.C.; Wang, L.L.; Zhang, L.; Zhao, Y.J.; Wu, J.W.; Wei, Y.M.; Zhang, X.S.; Zhao, X.Y. DEK43 is a P-type pentatricopeptide repeat (PPR) protein responsible for the cis-splicing of nad4 in maize mitochondria. J. Integr. Plant Biol. 2020, 62, 299–313. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.G.; Jin, G.P.; Fang, P.; Zhang, Y.; Feng, X.Z.; Tang, Y.P.; Qi, W.W.; Song, R.T. Maize pentatricopeptide repeat protein DEK41 affects cis-splicing of mitochondrial nad4 intron 3 and is required for normal seed development. J. Exp. Bot. 2019, 70, 3795–3808. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.H.; Song, S.; Yang, Y.Z.; Lu, F.; Zhang, M.D.; Sun, F.; Jia, R.X.; Song, R.L.; Tan, B.C. DEK46 performs C-to-U editing of a specific site in mitochondrial nad7 introns that is critical for intron splicing and seed development in maize. Plant J. 2020, 103, 1767–1782. [Google Scholar] [CrossRef] [PubMed]
- Dai, D.W.; Jin, L.F.; Huo, Z.Z.; Yan, S.M.; Ma, Z.Y.; Qi, W.W.; Song, R.T. Maize pentatricopeptide repeat protein DEK53 is required for mitochondrial RNA editing at multiple sites and seed development. J. Exp. Bot. 2020, 71, 6246–6261. [Google Scholar] [CrossRef] [PubMed]
- Ren, R.C.; Yan, X.W.; Zhao, Y.J.; Wei, Y.M.; Lu, X.D.; Zang, J.; Wu, J.W.; Zheng, G.M.; Ding, X.H.; Zhang, X.S.; et al. The novel E-subgroup pentatricopeptide repeat protein DEK55 is responsible for RNA editing at multiple sites and for the splicing of nad1 and nad4 in maize. BMC Plant Biol. 2020, 20, 553. [Google Scholar] [CrossRef]
- Fan, K.J.; Peng, Y.X.; Ren, Z.J.; Li, D.L.; Zhen, S.H.; Hey, S.; Cui, Y.; Fu, J.J.; Gu, R.L.; Wang, J.H.; et al. Maize defective kernel605 encodes a canonical DYW-Type PPR protein that edits a conserved site of nad1 and is essential for seed nutritional quality. Plant Cell Physiol. 2020, 61, 1954–1966. [Google Scholar] [CrossRef]
- Li, X.J.; Zhang, Y.F.; Hou, M.M.; Sun, F.; Shen, Y.; Xiu, Z.H.; Wang, X.M.; Chen, Z.L.; Sun, S.S.M.; Small, I.; et al. Small kernel 1 encodes a pentatricopeptide repeat protein required for mitochondrial nad7 transcript editing and seed development in maize (Zea mays) and rice (Oryza sativa). Plant J. 2014, 79, 797–809. [Google Scholar] [CrossRef]
- Wang, H.C.; Sayyed, A.; Liu, X.Y.; Yang, Y.Z.; Sun, F.; Wang, Y.; Wang, M.D.; Tan, B.C. SMALL KERNEL4 is required for mitochondrial cox1 transcript editing and seed development in maize. J. Integr. Plant Biol. 2020, 62, 777–792. [Google Scholar] [CrossRef]
- Ding, S.; Liu, X.Y.; Wang, H.C.; Wang, Y.; Tang, J.J.; Yang, Y.Z.; Tan, B.C. SMK6 mediates the C-to-U editing at multiple sites in maize mitochondria. J. Plant Physiol. 2019, 240, 152992. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.Y.; Liu, M.; Xiao, Z.Y.; Ren, X.M.; Zhao, H.L.; Gong, D.M.; Liang, K.; Tan, Z.D.; Shao, Y.Q.; Qiu, F.Z. ZmSMK9, a pentatricopeptide repeat protein, is involved in the cis-splicing of nad5, kernel development and plant architecture in maize. Plant Sci. 2019, 288, 110205. [Google Scholar] [CrossRef]
- Sosso, D.; Mbelo, S.; Vernoud, V.; Gendrot, G.; Dedieu, A.; Chambrier, P.; Dauzat, M.; Heurtevin, L.; Guyon, V.; Takenaka, M.; et al. PPR2263, a DYW-subgroup pentatricopeptide repeat protein, is required for mitochondrial nad5 and cob transcript editing, mitochondrion biogenesis, and maize growth. Plant Cell 2012, 24, 676–691. [Google Scholar] [CrossRef] [Green Version]
- Manavski, N.; Guyon, V.; Meurer, J.; Wienand, U.; Brettschneider, R. An essential pentatricopeptide repeat protein facilitates 5′ maturation and translation initiation of rps3 mRNA in maize mitochondria. Plant Cell 2012, 24, 3087–3105. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.Z.; Ding, S.; Wang, Y.; Wang, H.C.; Liu, X.Y.; Sun, F.; Xu, C.H.; Liu, B.H.; Tan, B.C. PPR20 is required for the cis-splicing of mitochondrial nad2 intron 3 and seed development in maize. Plant Cell Physiol. 2020, 61, 370–380. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.F.; Suzuki, M.; Sun, F.; Tan, B.C. The mitochondrion-targeted PENTATRICOPEPTIDE REPEAT78 protein is required for nad5 mature mRNA stability and seed development in maize. Mol. Plant 2017, 10, 1321–1333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutierrez-Marcos, J.F.; Dal Pra, M.; Giulini, A.; Costa, L.M.; Gavazzi, G.; Cordelier, S.; Sellam, O.; Tatout, C.; Paul, W.; Perez, P.; et al. Empty pericarp4 encodes a mitochondrion-targeted pentatricopeptide repeat protein necessary for seed development and plant growth in maize. Plant Cell 2007, 19, 196–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.J.; Xiu, Z.H.; Meeley, R.; Tan, B.C. Empty pericarp5 encodes a pentatricopeptide repeat protein that is required for mitochondrial RNA editing and seed development in maize. Plant Cell 2013, 25, 868–883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, F.; Wang, X.M.; Bonnard, G.; Shen, Y.; Xiu, Z.H.; Li, X.J.; Gao, D.H.; Zhang, Z.H.; Tan, B.C. Empty pericarp7 encodes a mitochondrial E-subgroup pentatricopeptide repeat protein that is required for ccmFN editing, mitochondrial function and seed development in maize. Plant J. 2015, 84, 283–295. [Google Scholar] [CrossRef]
- Sun, F.; Zhang, X.Y.; Shen, Y.; Wang, H.C.; Liu, R.; Wang, X.M.; Gao, D.H.; Yang, Y.Z.; Liu, Y.W.; Tan, B.C. The pentatricopeptide repeat protein EMPTY PERICARP8 is required for the splicing of three mitochondrial introns and seed development in maize. Plant J. 2018, 95, 919–932. [Google Scholar] [CrossRef]
- Yang, Y.Z.; Ding, S.; Wang, H.C.; Sun, F.; Huang, W.L.; Song, S.; Xu, C.H.; Tan, B.C. The pentatricopeptide repeat protein EMP9 is required for mitochondrial ccmB and rps4 transcript editing, mitochondrial complex biogenesis and seed development in maize. New Phytol. 2017, 214, 782–795. [Google Scholar] [CrossRef] [Green Version]
- Cai, M.J.; Li, S.Z.; Sun, F.; Sun, Q.; Zhao, H.L.; Ren, X.M.; Zhao, Y.X.; Tan, B.C.; Zhang, Z.X.; Qiu, F.Z. Emp10 encodes a mitochondrial PPR protein that affects the cis-splicing of nad2 intron 1 and seed development in maize. Plant J. 2017, 91, 132–144. [Google Scholar] [CrossRef] [Green Version]
- Ren, X.M.; Pan, Z.Y.; Zhao, H.L.; Zhao, J.L.; Cai, M.J.; Li, J.; Zhang, Z.X.; Qiu, F.Z. EMPTY PERICARP11 serves as a factor for splicing of mitochondrial nad1 intron and is required to ensure proper seed development in maize. J. Exp. Bot. 2017, 68, 4571–4581. [Google Scholar] [CrossRef] [Green Version]
- Sun, F.; Xiu, Z.H.; Jiang, R.C.; Liu, Y.W.; Zhang, X.Y.; Yang, Y.Z.; Li, X.J.; Zhang, X.; Wang, Y.; Tan, B.C. The mitochondrial pentatricopeptide repeat protein EMP12 is involved in the splicing of three nad2 introns and seed development in maize. J. Exp. Bot. 2019, 70, 963–972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Lu, G.; Liu, L.; Raihan, M.S.; Xu, J.T.; Jian, L.M.; Zhao, L.X.; Tran, T.M.; Zhang, Q.H.; Liu, J.; et al. The kernel size-related quantitative trait locus qKW9 encodes a pentatricopeptide repeat protein that affects photosynthesis and grain filling. Plant Physiol. 2020, 183, 1696–1709. [Google Scholar] [CrossRef]
- Xiu, Z.H.; Sun, F.; Shen, Y.; Zhang, X.Y.; Jiang, R.C.; Bonnard, G.; Zhang, J.H.; Tan, B.C. EMPTY PERICARP16 is required for mitochondrial nad2 intron 4 cis-splicing, complex I assembly and seed development in maize. Plant J. 2016, 85, 507–519. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, X.Y.; Huang, Z.Q.; Li, Y.Y.; Yang, Y.Z.; Sayyed, A.; Sun, F.; Gu, Z.Q.; Wang, X.M.; Tan, B.C. PPR-DYW protein EMP17 is required for mitochondrial RNA editing, complex III biogenesis, and seed development in maize. Front. Plant Sci. 2021, 12, 693272. [Google Scholar]
- Li, X.L.; Huang, W.L.; Yang, H.H.; Jiang, R.C.; Sun, F.; Wang, H.C.; Zhao, J.; Xu, C.H.; Tan, B.C. EMP18 functions in mitochondrial atp6 and cox2 transcript editing and is essential to seed development in maize. New Phytol. 2019, 221, 896–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Liu, X.Y.; Yang, Y.Z.; Huang, J.; Sun, F.; Lin, J.S.; Gu, Z.Q.; Sayyed, A.; Xu, C.H.; Tan, B.C. Empty Pericarp21 encodes a novel PPR-DYW protein that is required for mitochondrial RNA editing at multiple sites, complexes I and V biogenesis, and seed development in maize. PLoS Genet. 2019, 15, e1008305. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.Z.; Ding, S.; Liu, X.Y.; Tang, J.J.; Wang, Y.; Sun, F.; Xu, C.H.; Tan, B.C. EMP32 is required for the cis-splicing of nad7 intron 2 and seed development in maize. RNA Biol. 2021, 18, 499–509. [Google Scholar] [CrossRef]
- Ren, Z.J.; Fan, K.J.; Fang, T.; Zhang, J.J.; Yang, L.; Wang, J.H.; Wang, G.Y.; Liu, Y.J. Maize Empty pericarp602 encodes a P-type PPR protein that is essential for seed development. Plant Cell Physiol. 2019, 60, 1734–1746. [Google Scholar] [CrossRef]
- Fan, K.J.; Ren, Z.J.; Zhang, X.F.; Liu, Y.; Fu, J.J.; Qi, C.L.; Tatar, W.; Rasmusson, A.G.; Wang, G.Y.; Liu, Y.J. The pentatricopeptide repeat protein EMP603 is required for the splicing of mitochondrial Nad1 intron 2 and seed development in maize. J. Exp. Bot. 2021, erab339. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.C.; Chen, Z.L.; Yang, Y.Z.; Sun, F.; Ding, S.; Li, X.L.; Xu, C.H.; Tan, B.C. PPR14 interacts with PPR-SMR1 and CRM protein Zm-mCSF1 to facilitate mitochondrial intron splicing in maize. Front. Plant Sci. 2020, 11, 732. [Google Scholar] [CrossRef]
- Liu, R.; Cao, S.K.; Sayyed, A.; Xu, C.H.; Sun, F.; Wang, X.M.; Tan, B.C. The mitochondrial pentatricopeptide repeat protein PPR18 is required for the cis-splicing of nad4 intron 1 and essential to seed development in maize. Int. J. Mol. Sci. 2020, 21, 4047. [Google Scholar]
- Liu, R.; Cao, S.K.; Sayyed, A.; Yang, H.H.; Zhao, J.; Wang, X.M.; Jia, R.X.; Sun, F.; Tan, B.C. The DYW-subgroup pentatricopeptide repeat protein PPR27 functions on editing of multiple mitochondrial transcripts and interacts with ZmMORF1 in maize. J. Exp. Bot. 2020, 71, 5495–5505. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.H.; Xiu, Z.H.; Wang, L.; Cao, S.K.; Li, X.L.; Sun, F.; Tan, B.C. Two pentatricopeptide repeat proteins are required for the splicing of nad5 introns in maize. Front. Plant Sci. 2020, 11, 732. [Google Scholar] [CrossRef]
- Chen, Z.L.; Wang, H.C.; Shen, J.Y.; Sun, F.; Wang, M.D.; Xu, C.H.; Tan, B.C. PPR-SMR1 is required for the splicing of multiple mitochondrial introns, interacts with Zm-mCSF1, and is essential for seed development in maize. J. Exp. Bot. 2019, 70, 5245–5258. [Google Scholar] [CrossRef]
- Sosso, D.; Canut, M.; Gendrot, G.; Dedieu, A.; Chambrier, P.; Barkan, A.; Consonni, G.; Rogowsky, P.M. PPR8522 encodes a chloroplast-targeted pentatricopeptide repeat protein necessary for maize embryogenesis and vegetative development. J. Exp. Bot. 2012, 63, 5843–5857. [Google Scholar] [CrossRef] [Green Version]
- Yuan, N.N.; Wang, J.C.; Zhou, Y.; An, D.; Xiao, Q.; Wang, W.Q.; Wu, Y.R. EMB-7L is required for embryogenesis and plant development in maize involved in RNA splicing of multiple chloroplast genes. Plant Sci. 2019, 287, 110203. [Google Scholar] [CrossRef] [PubMed]
- De Longevialle, A.F.; Meyer, E.H.; Andres, C.; Taylor, N.L.; Lurin, C.; Millar, A.H.; Small, I.D. The pentatricopeptide repeat gene OTP43 is required for trans-splicing of the mitochondrial nad1 intron 1 in Arabidopsis thaliana. Plant Cell 2007, 19, 3256–3265. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Han, J.H.; Park, Y.I.; Colas des Francs-Small, C.; Small, I.; Kang, H. The mitochondrial pentatricopeptide repeat protein PPR19 is involved in the stabilization of NADH dehydrogenase 1 transcripts and is crucial for mitochondrial function and Arabidopsis thaliana development. New Phytol. 2017, 215, 202–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Huang, J.Y.; Zhong, S.; Gu, H.Y.; He, S.; Qu, L.J. Novel DYW-type pentatricopeptide repeat (PPR) protein BLX controls mitochondrial RNA editing and splicing essential for early seed development of Arabidopsis. J. Genet. Genom. 2018, 45, 155–168. [Google Scholar] [CrossRef]
- Lu, Y.Q.; Li, C.; Wang, H.; Chen, H.; Berg, H.; Xia, Y.J. AtPPR2, an Arabidopsis pentatricopeptide repeat protein, binds to plastid 23S rRNA and plays an important role in the first mitotic division during gametogenesis and in cell proliferation during embryogenesis. Plant J. 2011, 67, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.W.; An, Y.Q.; Qi, Z.; Xiao, J.W. PPR protein early chloroplast development 2 is essential for chloroplast development at the early stage of Arabidopsis development. Plant Sci. 2021, 308, 110908. [Google Scholar] [CrossRef]
- Aryamanesh, N.; Ruwe, H.; Sanglard, L.V.P.; Eshraghi, L.; Bussell, J.D.; Howell, K.A.; Small, I.; Colas des Francs-Small, C. The pentatricopeptide repeat protein EMB2654 is essential for trans-splicing of a chloroplast small ribosomal subunit transcript. Plant Physiol. 2017, 173, 1164–1176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchetti, F.; Cainzos, M.; Shevtsov, S.; Córdoba, J.P.; Sultan, L.D.; Brennicke, A.; Takenaka, M.; Pagnussat, G.; Ostersetzer-Biran, O.; Zabaleta, E. Mitochondrial pentatricopeptide repeat protein, EMB2794, plays a pivotal role in NADH dehydrogenase subunit nad2 mRNA maturation in Arabidopsis thaliana. Plant Cell Physiol. 2020, 61, 1080–1094. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.H.; Liu, N.Y.; Tang, Z.S.; Liu, J.; Yang, W.C. Arabidopsis GLUTAMINE-RICH PROTEIN23 is essential for early embryogenesis and encodes a novel nuclear PPR motif protein that interacts with RNA polymerase II subunit III. Plant Cell 2006, 18, 815–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, P.; Wang, F.; Li, N.; Shi, D.Q.; Yang, W.C. Pentatricopeptide repeat protein MID1 modulates nad2 intron 1 splicing and Arabidopsis development. Sci Rep. 2020, 10, 2008. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xiao, J.W.; Li, Y.Q.; Su, B.D.; Xu, H.M.; Shan, X.Y.; Song, C.W.; Xie, J.B.; Li, R.L. PDM3, a pentatricopeptide repeat-containing protein, affects chloroplast development. J. Exp. Bot. 2017, 68, 5615–5627. [Google Scholar] [CrossRef]
- Zhang, L.; Qi, Y.Z.; Wu, M.M.; Zhao, L.; Zhao, Z.C.; Lei, C.L.; Hao, Y.Y.; Yu, X.W.; Sun, Y.L.; Zhang, X.; et al. Mitochondrion-targeted PENTATRICOPEPTIDE REPEAT5 is required for cis-splicing of nad4 intron 3 and endosperm development in rice. Crop J. 2021, 9, 282–296. [Google Scholar] [CrossRef]
- Leu, K.C.; Hsieh, M.H.; Wang, H.J.; Hsieh, H.L.; Jauh, G.Y. Distinct role of Arabidopsis mitochondrial P-type pentatricopeptide repeat protein-modulating editing protein, PPME, in nad1 RNA editing. RNA Biol. 2016, 13, 593–604. [Google Scholar] [CrossRef] [Green Version]
- Small, I.D.; Schallenberg-Rudinger, M.; Takenaka, M.; Mireau, H.; Ostersetzer-Biran, O. Plant organellar RNA editing: What 30 years of research has revealed. Plant J. 2020, 101, 1040–1056. [Google Scholar] [CrossRef] [PubMed]
- Blanc, V.; Litvak, S.; Araya, A. RNA editing in wheat mitochondria proceeds by a deamination mechanism. FEBS Lett. 1995, 373, 56–60. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, S.; Navaratnam, N.; Morrison, J.R.; Scott, J.; Taylor, W.R. Cytosine nucleoside/nucleotide deaminases and apolipoprotein B mRNA editing. Trends Biochem. Sci. 1994, 19, 105–106. [Google Scholar] [CrossRef]
- Boussardon, C.; Avon, A.; Kindgren, P.; Bond, C.S.; Challenor, M.; Lurin, C.; Small, I. The cytidine deaminase signature HxE(x)nCxxC of DYW1 binds zinc and is necessary for RNA editing of ndhD-1. New Phytol. 2014, 203, 1090–1095. [Google Scholar] [CrossRef]
- Hayes, M.L.; Dang, K.N.; Diaz, M.F.; Mulligan, R.M. A conserved glutamate residue in the C-terminal deaminase domain of pentatricopeptide repeat proteins is required for RNA editing activity. J. Biol. Chem. 2015, 290, 10136–10142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oldenkott, B.; Yang, Y.Y.; Lesch, E.; Knoop, V.; Schallenberg-Rüdinger, M. Plant-type pentatricopeptide repeat proteins with a DYW domain drive C-to-U RNA editing in Escherichia coli. Commun. Biol. 2019, 2, 85. [Google Scholar] [CrossRef] [PubMed]
- Hayes, M.L.; Santibanez, P.I. A plant pentatricopeptide repeat protein with a DYW-deaminase domain is sufficient for catalyzing C-to-U RNA editing in vitro. J. Biol. Chem. 2020, 295, 3497–3505. [Google Scholar] [CrossRef] [PubMed]
- Takenaka, M.; Takenaka, S.; Barthel, T.; Frink, B.; Haag, S.; Verbitskiy, D.; Oldenkott, B.; Schallenberg-Rüdinger, M.; Feiler, C.G.; Weiss, M.S.; et al. DYW domain structures imply an unusual regulation principle in plant organellar RNA editing catalysis. Nat. Catal. 2021, 4, 510–522. [Google Scholar] [CrossRef]
- Boussardon, C.; Salone, V.; Avon, A.; Berthomé, R.; Hammani, K.; Okuda, K.; Shikanai, T.; Small, I.; Lurin, C. Two interacting proteins are necessary for the editing of the NdhD-1 site in Arabidopsis plastids. Plant Cell 2012, 24, 3684–3694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andres-Colas, N.; Zhu, Q.; Takenaka, M.; De Rybel, B.; Weijers, D.; Van Der Straeten, D. Multiple PPR protein interactions are involved in the RNA editing system in Arabidopsis mitochondria and plastids. Proc. Natl. Acad. Sci. USA 2017, 114, 8883–8888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malbert, B.; Burger, M.; Lopez-Obando, M.; Baudry, K.; Launay-Avon, A.; Härtel, B.; Verbitskiy, D.; Jörg, A.; Berthomé, R.; Lurin, C.; et al. The analysis of the editing defects in the dyw2 mutant provides new clues for the prediction of RNA targets of Arabidopsis E+-class PPR proteins. Plants 2020, 9, 280. [Google Scholar] [CrossRef] [Green Version]
- Guillaumot, D.; Lopez-Obando, M.; Baudry, K.; Avon, A.; Rigaill, G.; Falcon de Longevialle, A.; Broche, B.; Takenaka, M.; Berthomé, R.; De Jaeger, G.; et al. Two interacting PPR proteins are major Arabidopsis editing factors in plastid and mitochondria. Proc. Natl. Acad. Sci. USA 2017, 114, 8877–8882. [Google Scholar] [CrossRef] [Green Version]
- Bentolila, S.; Heller, W.P.; Sun, T.; Babina, A.M.; Friso, G.; van Wijk, K.J.; Hanson, M.R. RIP1, a member of an Arabidopsis protein family, interacts with the protein RARE1 and broadly affects RNA editing. Proc. Natl. Acad. Sci. USA 2012, 109, e1453–e1461. [Google Scholar] [CrossRef] [Green Version]
- Härtel, B.; Zehrmann, A.; Verbitskiy, D.; van der Merwe, J.A.; Brennicke, A.; Takenaka, M. MEF10 is required for RNA editing at nad2-842 in mitochondria of Arabidopsis thaliana and interacts with MORF8. Plant Mol. Biol. 2013, 81, 337–346. [Google Scholar] [CrossRef]
- Brehme, N.; Bayer-Császár, E.; Glass, F.; Takenaka, M. The DYW subgroup PPR protein MEF35 targets RNA editing sites in the mitochondrial rpl16, nad4 and cob mRNAs in Arabidopsis thaliana. PLoS ONE 2015, 10, e0140680. [Google Scholar] [CrossRef] [Green Version]
- Barkan, A.; Rojas, M.; Fujii, S.; Yap, A.; Chong, Y.S.; Bond, C.S.; Small, I. A combinatorial amino acid code for RNA recognition by pentatricopeptide repeat proteins. PLoS Genet. 2012, 8, e1002910. [Google Scholar] [CrossRef]
- Huang, C.; Li, Z.R.; Yu, Q.B.; Ye, L.S.; Cui, Y.L.; Molloy, D.P.; Yang, Z.N. MORF2 tightly associates with MORF9 to regulate chloroplast RNA editing in Arabidopsis. Plant Sci. 2019, 278, 64–69. [Google Scholar] [CrossRef]
- Yan, J.J.; Zhang, Q.X.; Guan, Z.Y.; Wang, Q.; Li, L.; Ruan, F.Y.; Lin, R.C.; Zou, T.T.; Yin, P. MORF9 increases the RNA-binding activity of PLS-type pentatricopeptide repeat protein in plastid RNA editing. Nat. Plants 2017, 3, 17037. [Google Scholar] [CrossRef]
- Brown, G.G.; Colas des Francs-Small, C.; Ostersetzer-Biran, O. Group II intron splicing factors in plant mitochondria. Front. Plant Sci. 2014, 5, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitz-Linneweber, C.; Lampe, M.K.; Sultan, L.D.; Ostersetzer-Biran, O. Organellar maturases: A window into the evolution of the spliceosome. Biochim. Biophys. Acta BBA Bioenerg. 2015, 1847, 798–808. [Google Scholar] [CrossRef] [Green Version]
- Colas des Francs-Small, C.; Falcon de Longevialle, A.; Li, Y.H.; Lowe, E.; Tanz, S.K.; Smith, C.; Bevan, M.W.; Small, I. The pentatricopeptide repeat proteins TANG2 and ORGANELLE TRANSCRIPT PROCESSING439 are involved in the splicing of the multipartite nad5 transcript encoding a subunit of mitochondrial complex I. Plant Physiol. 2014, 165, 1409–1416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, W.Y.; Liao, J.C.; Chang, C.Y.; Harrison, T.; Boucher, C.; Hsieh, M.H. The SLOW GROWTH3 pentatricopeptide repeat protein is Required for the splicing of mitochondrial NADH dehydrogenase subunit7 intron 2 in Arabidopsis. Plant Physiol. 2015, 168, 490–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haïli, N.; Planchard, N.; Arnal, N.; Quadrado, M.; Vrielynck, N.; Dahan, J.; Colas des Francs-Small, C.; Mireau, H. The MTL1 pentatricopeptide repeat protein is required for both translation and splicing of the mitochondrial NADH DEHYDROGENASE SUBUNIT7 mRNA in Arabidopsis. Plant Physiol. 2016, 170, 354–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiu, Z.H.; Peng, L.; Wang, Y.; Yang, H.H.; Sun, F.; Wang, X.M.; Cao, S.K.; Jiang, R.C.; Wang, L.; Chen, B.Y.; et al. Empty pericarp24 and Empty pericarp25 are required for the splicing of mitochondrial introns, complex I assembly, and seed development in maize. Front. Plant Sci. 2020, 11, 608550. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.Y.; Wang, Y.L.; Wu, M.M.; Zhu, X.P.; Teng, X.; Sun, Y.L.; Zhu, J.P.; Zhang, Y.Y.; Jing, R.N.; Lei, J.; et al. The nuclear-localized PPR protein OsNPPR1 is important for mitochondrial function and endosperm development in rice. J. Exp. Bot. 2019, 70, 4705–4720. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.M.; Ren, Y.L.; Cai, M.H.; Wang, Y.L.; Zhu, S.S.; Zhu, J.P.; Hao, Y.Y.; Teng, X.; Zhu, X.P.; Jing, R.N.; et al. Rice FLOURY ENDOSPERM 10 encodes a pentatricopeptide repeat protein that is essential for the trans-splicing of mitochondrial nad1 intron 1 and endosperm development. New Phytol. 2020, 223, 736–750. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.Y.; Liu, L.L.; Yu, Y.F.; Zhu, J.P.; Gao, H.; Wang, Y.H.; Wan, J.M. Lose-of-function of a rice nucleolus-localized pentatricopeptide repeat protein is responsible for the floury endosperm14 mutant phenotypes. Rice 2019, 12, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weißenberger, S.; Soll, J.; Carrie, C. The PPR protein SLOW GROWTH 4 is involved in editing of nad4 and affects the splicing of nad2 intron 1. Plant Mol. Biol. 2016, 93, 355–368. [Google Scholar] [CrossRef]
- Ichinose, M.; Tasaki, E.; Sugita, C.; Sugita, M. A PPR-DYW protein is required for splicing of a group II intron of cox1 pre-mRNA in Physcomitrella patens. Plant J. 2012, 70, 271–278. [Google Scholar] [CrossRef]
- Schmitz-Linneweber, C.; Williams-Carrier, R.E.; Williams-Voelker, P.M.; Kroeger, T.S.; Vichas, A.; Barkan, A. A pentatricopeptide repeat protein facilitates the trans-splicing of the maize chloroplast rps12 pre-mRNA. Plant Cell 2006, 18, 2650–2663. [Google Scholar] [CrossRef] [Green Version]
- Tadini, L.; Ferrari, R.; Lehniger, M.K.; Mizzotti, C.; Moratti, F.; Resentini, F.; Colombo, M.; Costa, A.; Masiero, S.; Pesaresi, P. Trans-splicing of plastid rps12 transcripts, mediated by AtPPR4, is essential for embryo patterning in Arabidopsis thaliana. Planta 2018, 248, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Ostheimer, G.J.; Williams-Carrier, R.; Belcher, S.; Osborne, E.; Gierke, J.; Barkan, A. Group II intron splicing factors derived by diversification of an ancient RNA-binding domain. EMBO J. 2003, 22, 3919–3929. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, B.D.; Barkan, A. Recruitment of a peptidyl-tRNA hydrolase as a facilitator of group II intron splicing in chloroplasts. EMBO J. 2001, 20, 872–879. [Google Scholar] [CrossRef]
- Asakura, Y.; Galarneau, E.; Watkins, K.P.; Barkan, A.; van Wijk, K.J. Chloroplast RH3 DEAD Box RNA helicases in maize and Arabidopsis function in splicing of specific group II introns and affect chloroplast ribosome biogenesis. Plant Physiol. 2012, 159, 961–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.D.; Aubé, F.; Quadrado, M.; Dargel-Graffin, C.; Mireau, H. Three new pentatricopeptide repeat proteins facilitate the splicing of mitochondrial transcripts and complex I biogenesis in Arabidopsis. J. Exp. Bot. 2018, 69, 5131–5140. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Fourdin, R.; Quadrado, M.; Dargel-Graffin, C.; Tollete, D. Rerouting of ribosomal proteins into splicing in plant organelles. Proc. Natl. Acad. Sci. USA 2020, 117, 29979–29987. [Google Scholar] [CrossRef] [PubMed]
- Khrouchtchova, A.; Monde, R.A.; Barkan, A. A short PPR protein required for the splicing of specific group II introns in angiosperm chloroplasts. RNA 2012, 18, 1197–1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ke, J.Y.; Chen, R.Z.; Ban, T.; Zhou, X.E.; Gu, X.; Tan, M.H.E.; Chen, C.; Kang, Y.Y.; Brunzelle, J.S.; Zhu, J.K.; et al. Structural basis for RNA recognition by a dimeric PPR-protein complex. Nat. Struct. Mol. Biol. 2013, 20, 1377–1382. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Park, S.J.; Colas des Francs-Small, C.; Whitby, M.; Small, I.; Kang, H. The coordinated action of PPR4 and EMB 2654 on each intron half mediates trans-splicing of rps12 transcripts in plant chloroplasts. Plant J. 2019, 100, 1193–1207. [Google Scholar] [CrossRef]
- Fedorova, O.; Solem, A.; Pyle, A.M. Protein-facilitated folding of group II intron ribozymes. J. Mol. Biol. 2010, 397, 799–813. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.J.; Yao, Y.Y.; Hong, S.X.; Yang, Y.; Shen, C.C.; Zhang, Q.X.; Zhang, D.L.; Zou, T.T.; Yin, P. Delineation of pentatricopeptide repeat codes for target RNA prediction. Nucleic Acids Res. 2019, 47, 3728–3738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
The functions of PPR proteins in plants. (A) The molecular functions of PPR proteins in plant mitochondria and chloroplasts. PPR proteins are encoded by nuclear genes, translated in the cytoplasm, and then imported into mitochondrion or chloroplast to mediate multiple steps of RNA processing. (B) The main growing and developmental phenotypes of plant mutants of PPR protein-coding genes.
Figure 1.
The functions of PPR proteins in plants. (A) The molecular functions of PPR proteins in plant mitochondria and chloroplasts. PPR proteins are encoded by nuclear genes, translated in the cytoplasm, and then imported into mitochondrion or chloroplast to mediate multiple steps of RNA processing. (B) The main growing and developmental phenotypes of plant mutants of PPR protein-coding genes.

Figure 2.
Proposed model for the molecular mechanisms of PPR proteins in plant organellar RNA editing and RNA splicing. (A) A model for the involvement of a DYW-type PPR protein in C-to-U RNA editing. C-to-U RNA editing in plant mitochondria and chloroplasts is carried out by an editosome composed of proteins and RNA. PPR motifs bind to its target RNA according to the PPR-RNA recognition code [123], which can be influenced by other proteins (shown in blue and orange). The DYW domain containing a cytidine deaminase signature (listed as HxE(x)nCxxC) converts the target cytidine (C) to uridine (U). (B) A model for the involvement of a P-class PPR protein in intron splicing. The splicing of introns in plant mitochondria and chloroplasts is proposed to be mediated by a ribonucleoprotein complex, where PPR proteins and other RNA-binding proteins (shown in green, blue, and orange) bind to their target intron to help its folding and stabilizing in an active form.
Figure 2.
Proposed model for the molecular mechanisms of PPR proteins in plant organellar RNA editing and RNA splicing. (A) A model for the involvement of a DYW-type PPR protein in C-to-U RNA editing. C-to-U RNA editing in plant mitochondria and chloroplasts is carried out by an editosome composed of proteins and RNA. PPR motifs bind to its target RNA according to the PPR-RNA recognition code [123], which can be influenced by other proteins (shown in blue and orange). The DYW domain containing a cytidine deaminase signature (listed as HxE(x)nCxxC) converts the target cytidine (C) to uridine (U). (B) A model for the involvement of a P-class PPR protein in intron splicing. The splicing of introns in plant mitochondria and chloroplasts is proposed to be mediated by a ribonucleoprotein complex, where PPR proteins and other RNA-binding proteins (shown in green, blue, and orange) bind to their target intron to help its folding and stabilizing in an active form.


{kind=link}
{kind=link}
Table 1.
Selected functionally characterized PPR proteins essential for seed development in maize and Arabidopsis.
Table 1.
Selected functionally characterized PPR proteins essential for seed development in maize and Arabidopsis.
| Species | Subcellular Localization | Mutant Phenotype | Protein Name | PPR Class | Function(s) | References |
|---|---|---|---|---|---|---|
| Maize | Mitochondrion | dek | DEK2 | P | RNA splicing, nad1 intron 1 | [52] |
| Mitochondrion | dek | DEK10 | PLS | RNA editing, nad3-61, 62, and cox2-550 | [53] | |
| Mitochondrion | dek | DEK35 | P | RNA splicing, nad4 intron 1 | [54] | |
| Mitochondrion | dek | DEK36 | PLS | RNA editing, atp4-59, nad7-383, and ccmFN-302 | [55] | |
| Mitochondrion | dek | DEK37 | P | RNA splicing, nad2 intron 1 | [56] | |
| Mitochondrion | dek | DEK39 | PLS | RNA editing, nad3-247 and nad3-275 | [57] | |
| Mitochondrion | dek | DEK40 | PLS | RNA editing, cox3-314, nad2-26, and nad5-1916 | [58] | |
| Mitochondrion | dek | DEK41/ DEK43 | P | RNA splicing, nad4 intron 1 and 3 | [59,60] | |
| Mitochondrion | dek | DEK46 | PLS | RNA editing, D5-C22 of nad7 intron 3 and 4 | [61] | |
| Mitochondrion | dek | DEK53 | PLS | RNA editing, multiples sites | [62] | |
| Mitochondrion | dek | DEK55 | PLS | RNA splicing, nad4 intron 1 and 3;RNA editing, multiple sites | [63] | |
| Mitochondrion | dek | DEK605 | PLS | RNA editing, nad1-608 | [64] | |
| Mitochondrion | smk | SMK1 | PLS | RNA editing, nad7-836 | [65] | |
| Mitochondrion | smk | SMK4 | PLS | RNA editing, cox1-1489 | [66] | |
| Mitochondrion | smk | SMK6 | PLS | RNA editing, nad1-740, nad4L-110, nad7-739, and mttB-138, 139 | [67] | |
| Mitochondrion | smk | ZmSMK9 | P | RNA splicing, nad5 intron 1 and 4 | [68] | |
| Mitochondrion | smk | PPR2263 | PLS | RNA editing, nad5-1550 and cob-908 | [69] | |
| Mitochondrion | smk | MPPR6 | P | Translation, rps3 mRNA | [70] | |
| Mitochondrion | dek/smk | PPR20 | P | RNA splicing, nad2 intron 3 | [71] | |
| Mitochondrion | smk | PPR78 | P | RNA stabilization, nad5 mature mRNA | [72] | |
| Mitochondrion | emp | EMP4 | P | Expression of mitochondrial transcripts | [73] | |
| Mitochondrion | emp | EMP5 | PLS | RNA editing, multiple sites | [74] | |
| Mitochondrion | emp | EMP7 | PLS | RNA editing, ccmFN-1553 | [75] | |
| Mitochondrion | emp | EMP8 | P | RNA splicing, nad1 intron 4, nad2 intron 1, and nad4 intron 1 | [76] | |
| Mitochondrion | emp | EMP9 | PLS | RNA editing, ccmB-43 and rps4-335 | [77] | |
| Mitochondrion | emp | EMP10 | P | RNA splicing, nad2 intron 1 | [78] | |
| Mitochondrion | emp | EMP11 | P | RNA splicing, nad1 intron 1, 2, 3, and 4 | [79] | |
| Mitochondrion | emp | EMP12 | P | RNA splicing, nad2 intron 1, 2, and 4 | [80] | |
| Chloroplast | smk | qKW9 | PLS | RNA editing, NdhB-246 | [81] | |
| Mitochondrion | emp | EMP16 | P | RNA splicing, nad2 intron 4 | [82] | |
| Mitochondrion | emp | EMP17 | PLS | RNA editing, ccmFC-799 and nad2-677 | [83] | |
| Mitochondrion | emp | EMP18 | PLS | RNA editing, atp6-635 and cox2-449 | [84] | |
| Mitochondrion | emp | EMP21 | PLS | RNA editing, multiple sites | [85] | |
| Mitochondrion | emp | EMP32 | P | RNA splicing, nad7 intron 2 | [86] | |
| Mitochondrion | emp | EMP602 | P | RNA splicing, nad4 intron 1 and 3 | [87] | |
| Mitochondrion | emp | EMP603 | P | RNA splicing, nad1 intron 2 | [88] | |
| Mitochondrion | emp | PPR14 | P | RNA splicing, nad2 intron 3, nad7 intron 1 and 2 | [89] | |
| Mitochondrion | emp | PPR18 | P | RNA splicing, nad4 intron 1 | [90] | |
| Mitochondrion | emp | PPR27 | PLS | RNA editing, multiple sites | [91] | |
| Mitochondrion | emp | PPR101 | P | RNA splicing, nad5 intron 1 and 2 | [92] | |
| Mitochondrion | emp | PPR231 | P | RNA splicing, nad5 intron 1, 2, 3 and nad2 intron 3 | [92] | |
| Mitochondrion | emp | PPR-SMR1 | P | RNA splicing, multiple introns | [93] | |
| Chloroplast | emb | PPR8522 | P | RNA transcription, nearly all chloroplast-encoded genes | [94] | |
| Chloroplast | emb | EMB-7L | P | RNA splicing, multiple introns | [95] | |
| Arabidopsis | Mitochondrion | dek | OTP43 | P | RNA splicing, nad1 intron 1 | [96] |
| Mitochondrion | smk | PPR19 | P | RNA stabilization, nad1 mature mRNA | [97] | |
| Mitochondrion | emp | BLX | PLS | RNA editing, multiple sites; RNA splicing, nad1 intron 4 and nad2 intron 1 | [98] | |
| Chloroplast | emb | AtPPR2 | P | RNA translation | [99] | |
| Chloroplast | emb | ECD2 | P | RNA splicing, ndhA, ycf3 intron 1, rps12 intron 2 and clpp intron 2 | [100] | |
| Chloroplast | emb | EMB2654 | P | RNA splicing, rps12 intron 1 | [101] | |
| Mitochondrion | emb | EMB2794 | P | RNA splicing, nad2 intron 2 | [102] | |
| Nucleus | emb | GRP23 | P | RNA transcription | [103] | |
| Mitochondrion | emb | MID1 | P | RNA splicing, nad2 intron 1 | [104] | |
| Chloroplast | emb | PMD3 | P | RNA splicing, trnA, ndhB, and clpP-1 | [105] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, X.; Sun, M.; Liu, S.; Teng, Q.; Li, S.; Jiang, Y. Functions of PPR Proteins in Plant Growth and Development. Int. J. Mol. Sci. 2021, 22, 11274. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011274
AMA Style
Li X, Sun M, Liu S, Teng Q, Li S, Jiang Y. Functions of PPR Proteins in Plant Growth and Development. International Journal of Molecular Sciences. 2021; 22(20):11274. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011274
Chicago/Turabian StyleLi, Xiulan, Mengdi Sun, Shijuan Liu, Qian Teng, Shihui Li, and Yueshui Jiang. 2021. "Functions of PPR Proteins in Plant Growth and Development" International Journal of Molecular Sciences 22, no. 20: 11274. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011274
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.