
What Do We Know about Classical and Non-Classical Progesterone Receptors in the Human Female Reproductive Tract? A Review

 , ,
, ,  and
and
Abstract
:1. Introduction
2. PGR Mechanisms of Action
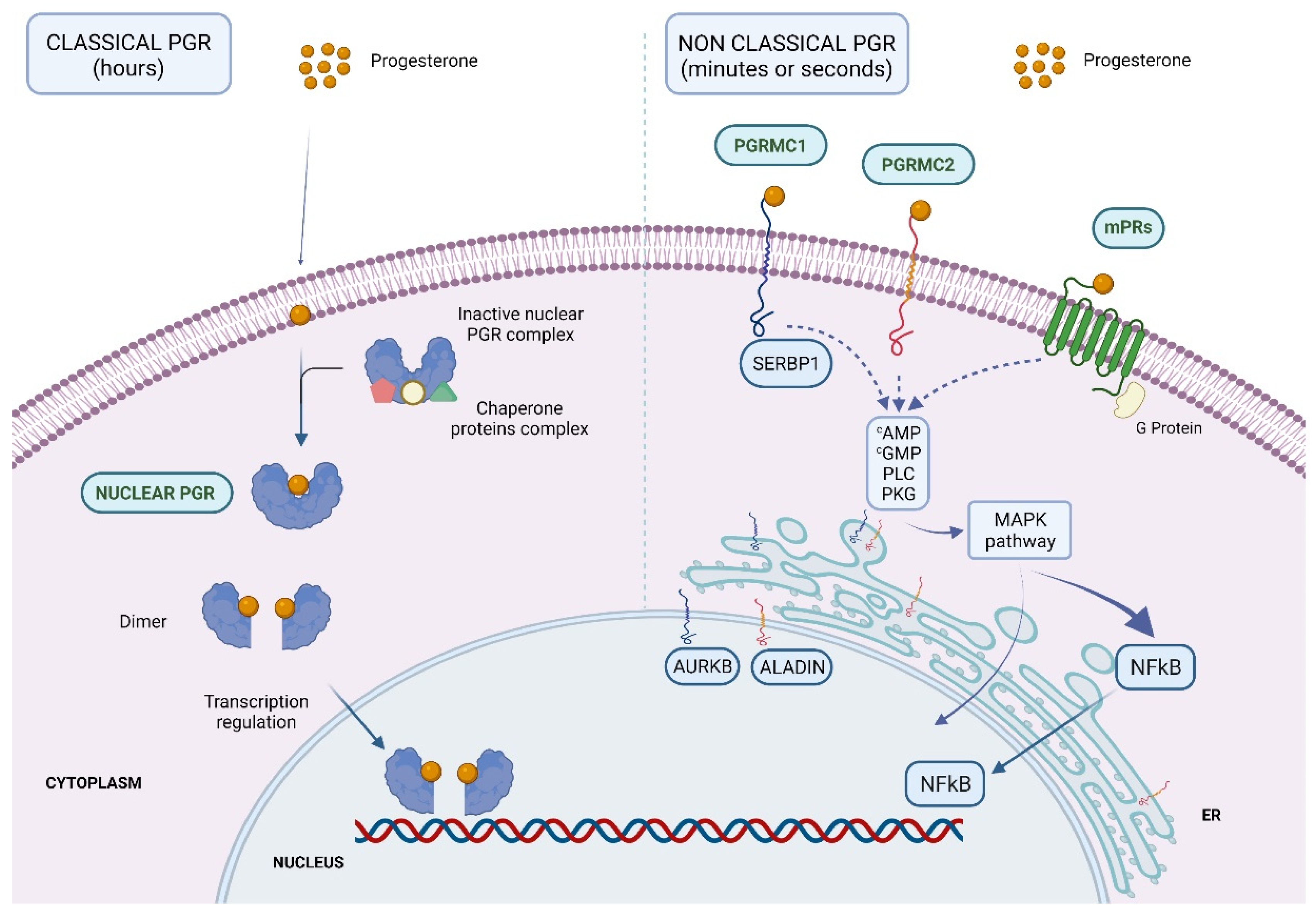
2.1. Classical PGR

2.2. Non-Classical PGR
3. PGR in the Ovary and Oviduct
3.1. Role of PGR in the Hypothalamic–Pituitary–Gonadal Axis
3.2. Role of PGR in Follicular Development and Recruitment
3.3. Role of PGR in Ovulation
3.4. Role of PGR in Oocyte Acquisition of Competence
3.5. Role of PGR in Oviductal Transport of COCs and Embryos
4. PGR in the Uterus
4.1. Role of PGR in the Endometrium
4.2. Role of PGR in the Myometrium
4.3. Role of PGR in the Cervix
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Taraborrelli, S. Physiology, production and action of progesterone. Acta Obstet. Gynecol. Scand. 2015, 94, 8–16. [Google Scholar] [CrossRef]
- Salazar, E.; Calzada, L. The role of progesterone in endometrial estradiol- and progesterone-receptor synthesis in women with menstrual disorders and habitual abortion. Gynecol. Endocrinol. 2007, 23, 222–225. [Google Scholar] [CrossRef]
- Szekeres-Bartho, J.; Wilczynski, J.R.; Basta, P.; Kalinka, J. Role of progesterone and progestin therapy in threatened abortion and preterm labour. Front. Biosci. 2008, 13, 1981–1990. [Google Scholar] [CrossRef] [Green Version]
- Graham, J.; Clarke, C. Physiological action of progesterone in target tissues. Endocr. Rev. 1997, 18, 502–519. [Google Scholar] [CrossRef] [Green Version]
- Conneely, O.M.; Mulac-Jericevic, B.; Arnett-Mansfield, R. Progesterone Signaling in Mammary Gland Development. In Progestins and the Mammary Gland. Ernst Schering Foundation Symposium Proceedings, vol 2007/1; Springer: Berlin/Heidelberg, Germany, 2008; pp. 175–185. [Google Scholar] [CrossRef]
- Loutradis, D.; Beretsos, P.; Arabatzi, E.; Anagnostou, E.; Drakakis, P. The role of steroid hormones in ART. J. Steroid Biochem. Mol. Biol. 2008, 112, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Yoshinaga, K. Progesterone and its downstream molecules as blastocyst implantation essential factors. Am. J. Reprod. Immunol. 2014, 72, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Morel, Y.; Roucher, F.; Plotton, I.; Goursaud, C.; Tardy, V.; Mallet, D. Evolution of steroids during pregnancy: Maternal, placental and fetal synthesis. Ann. Endocrinol. 2016, 77, 82–89. [Google Scholar] [CrossRef]
- Tamburrino, L.; Marchiani, S.; Muratori, M.; Luconi, M.; Baldi, E. Progesterone, spermatozoa and reproduction: An updated review. Mol. Cell. Endocrinol. 2020, 516, 110952. [Google Scholar] [CrossRef]
- Yin, P.; Lin, Z.; Cheng, Y.; Marsh, E.; Utsunomiya, H.; Ishikawa, H.; Xue, Q.; Reierstad, S.; Innes, J.; Thung, S.; et al. Progesterone receptor regulates Bcl-2 gene expression through direct binding to its promoter region in uterine leiomyoma cells. J. Clin. Endocrinol. Metab. 2007, 92, 4459–4466. [Google Scholar] [CrossRef] [Green Version]
- Bunch, K.; Tinnemore, D.; Huff, S.; Hoffer, Z.S.; Burney, R.O.; Stallings, J.D. Expression Patterns of Progesterone Receptor Membrane Components 1 and 2 in Endometria From Women With and Without Endometriosis. Reprod. Sci. 2013, 21, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Branchini, G.; Schneider, L.; Cericatto, R.; Capp, E.; Brum, I. Progesterone receptors A and B and estrogen receptor alpha expression in normal breast tissue and fibroadenomas. Endocrine 2009, 35, 459–466. [Google Scholar] [CrossRef]
- Kim, J.; Kurita, T.; Bulun, S. Progesterone action in endometrial cancer, endometriosis, uterine fibroids, and breast cancer. Endocr. Rev. 2013, 34, 130–162. [Google Scholar] [CrossRef] [Green Version]
- Hirota, Y. Progesterone governs endometrial proliferation-differentiation switching and blastocyst implantation. Endocr. J. 2019, 66, 199–206. [Google Scholar] [CrossRef] [Green Version]
- Liao, S.; Mi, H.; Chai, L.; Wang, H. Effects of progesterone receptor on proliferation of uterine leiomyoma cells. J. Biol. Regul. Homeost. Agents 2019, 33, 1685–1693. [Google Scholar] [CrossRef]
- Wu, Y.; Strawn, E.; Basir, Z.; Halverson, G.; Guo, S. Promoter hypermethylation of progesterone receptor isoform B (PR-B) in endometriosis. Epigenetics 2006, 1, 106–111. [Google Scholar] [CrossRef] [Green Version]
- Burney, R.; Talbi, S.; Hamilton, A.; Vo, K.; Nyegaard, M.; Nezhat, C.; Lessey, B.; Giudice, L. Gene expression analysis of endometrium reveals progesterone resistance and candidate susceptibility genes in women with endometriosis. Endocrinology 2007, 148, 3814–3826. [Google Scholar] [CrossRef] [Green Version]
- Ehn, N.L.; Cooper, M.E.; Orr, K.; Shi, M.; Johnson, M.K.; Caprau, D.; Dagle, J.; Steffen, K.; Johnson, K.; Marazita, M.L.; et al. Evaluation of Fetal and Maternal Genetic Variation in the Progesterone Receptor Gene for Contributions to Preterm Birth. Pediatr. Res. 2007, 62, 630–635. [Google Scholar] [CrossRef] [PubMed]
- Ito, K.; Utsunomiya, H.; Yaegashi, N.; Sasano, H. Biological roles of estrogen and progesterone in human endometrial carcinoma--new developments in potential endocrine therapy for endometrial cancer. Endocr. J. 2007, 54, 667–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kowalik, M.; Rekawiecki, R.; Kotwica, J. The putative roles of nuclear and membrane-bound progesterone receptors in the female reproductive tract. Reprod. Biol. 2013, 13, 279–289. [Google Scholar] [CrossRef]
- Horie, K.; Takakura, K.; Fujiwara, H.; Suginami, H.; Liao, S.; Mori, T. Immunohistochemical localization of androgen receptor in the human ovary throughout the menstrual cycle in relation to oestrogen and progesterone receptor expression. Hum. Reprod. 1992, 7, 184–190. [Google Scholar] [CrossRef]
- Iwai, T.; Nanbu, Y.; Iwai, M.; Taii, S.; Fujii, S.; Mori, T. Immunohistochemical localization of oestrogen receptors and progesterone receptors in the human ovary throughout the menstrual cycle. Virchows Arch. A Pathol. Anat. Histopathol. 1990, 417, 369–375. [Google Scholar] [CrossRef]
- Thijssen, J. Progesterone receptors in the human uterus and their possible role in parturition. J. Steroid Biochem. Mol. Biol. 2005, 97, 397–400. [Google Scholar] [CrossRef]
- Amso, N.; Crow, J.; Shaw, W. Comparative immunohistochemical study of oestrogen and progesterone receptors in the Fallopian tube and uterus at different stages of the menstrual cycle and the menopause. Hum. Reprod. 1994, 9, 1027–1037. [Google Scholar] [CrossRef] [PubMed]
- Barton, B.; Herrera, G.; Anamthathmakula, P.; Rock, J.; Willie, A.; Harris, E.; Takemaru, K.; Winuthayanon, W. Roles of steroid hormones in oviductal function. Reproduction 2020, 159, R125–R137. [Google Scholar] [CrossRef] [PubMed]
- Shanker, Y.; Rao, A. Regulation of progesterone biosynthesis in the human placenta by estradiol 17 beta and progesterone. Biochem. Mol. Biol. Int. 1997, 43, 591–599. [Google Scholar] [CrossRef]
- Abid, S.; Gokral, J.; Maitra, A.; Meherji, P.; Kadam, S.; Pires, E.; Modi, D. Altered expression of progesterone receptors in testis of infertile men. Reprod. Biomed. Online 2008, 17, 175–184. [Google Scholar] [CrossRef]
- Plant, T.M. The hypothalamo-pituitary-gonadal axis. J. Endocrinol. 2015, 226, T41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Orozco, J.; Camacho-Arroyo, I. Progesterone Actions During Central Nervous System Development. Front. Neurosci. 2019, 13, 503. [Google Scholar] [CrossRef] [Green Version]
- Brinton, R.D.; Thompson, R.F.; Foy, M.R.; Baudry, M.; Wang, J.M.; Finch, C.E.; Morgan, T.E.; Pike, C.J.; Mack, W.J.; Stanczyk, F.Z.; et al. Progesterone receptors: Form and function in brain. Front. Neuroendocrinol. 2008, 29, 313–339. [Google Scholar] [CrossRef] [Green Version]
- Doglioni, C.; Gambacorta, M.; Zamboni, G.; Coggi, G.; Vialet, G.; Roma, B.; Ca, O. Immunocytochemical Localization of Progesterone Receptors in Endocrine Cells of the Human Pancreas. Am. J. Pathol. 1990, 137, 999–1005. [Google Scholar]
- Bland, R. Steroid hormone receptor expression and action in bone. Clin. Sci. 2000, 98, 217–240. [Google Scholar] [CrossRef]
- Batra, S.; Iosif, C. Progesterone receptors in the female lower urinary tract. J. Urol. 1987, 138, 1301–1304. [Google Scholar] [CrossRef]
- Dressing, G.; Goldberg, J.; Charles, N.; Schwertfeger, K.; Lange, C. Membrane progesterone receptor expression in mammalian tissues: A review of regulation and physiological implications. Steroids 2011, 76, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Rice, C.; Pang, Y.; Pace, M.; Thomas, P. Cloning, expression, and characterization of a membrane progestin receptor and evidence it is an intermediary in meiotic maturation of fish oocytes. Proc. Natl. Acad. Sci. USA 2003, 100, 2231–2236. [Google Scholar] [CrossRef] [Green Version]
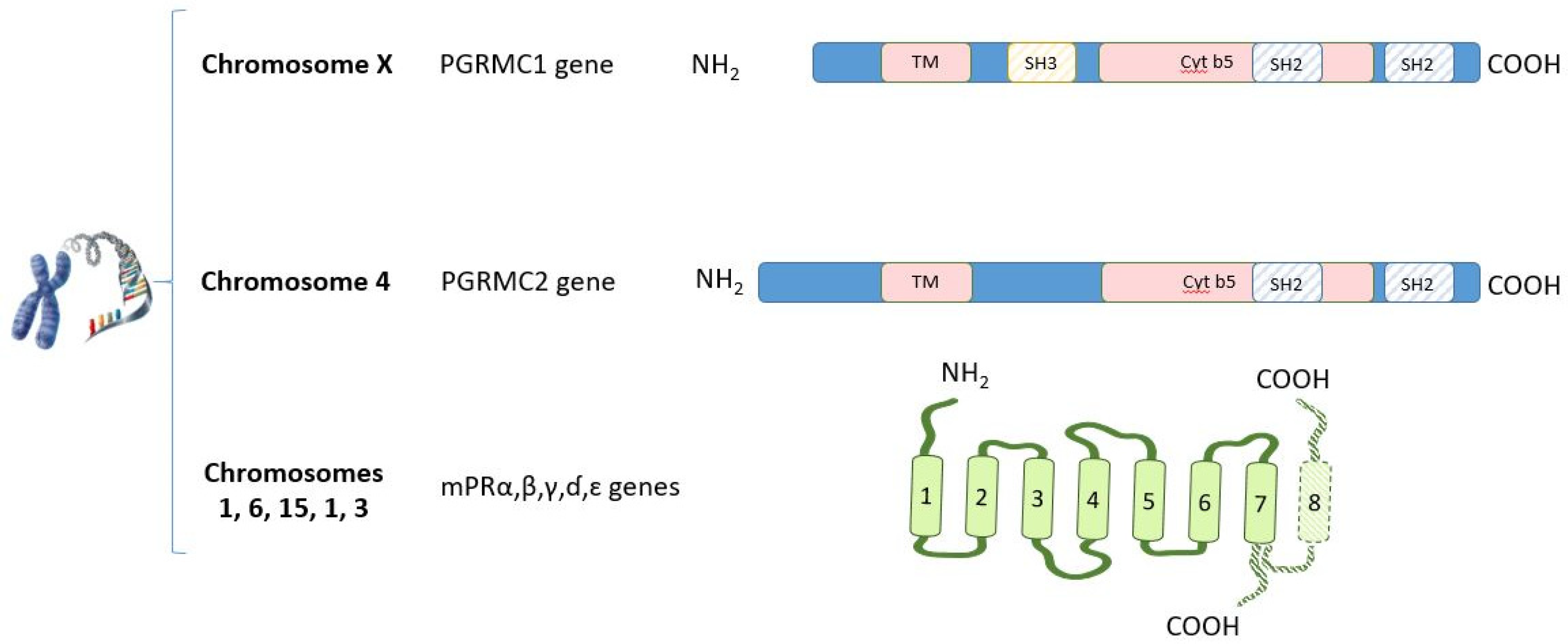
- Cahill, M. Progesterone receptor membrane component 1: An integrative review. J. Steroid Biochem. Mol. Biol. 2007, 105, 16–36. [Google Scholar] [CrossRef]
- Aparicio, I.M.; Garcia-Herreros, M.; O’Shea, L.C.; Hensey, C.; Lonergan, P.; Fair, T. Expression, Regulation, and Function of Progesterone Receptors in Bovine Cumulus Oocyte Complexes During In Vitro Maturation. Biol. Reprod. 2011, 84, 910–921. [Google Scholar] [CrossRef] [PubMed]
- Hagiwara, A.; Ogiwara, K.; Takahashi, T. Expression of Membrane Progestin Receptors (mPRs) in Granulosa Cells of Medaka Preovulatory Follicles. Zoolog. Sci. 2016, 33, 98–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peluso, J.; Liu, X.; Gawkowska, A.; Johnston-MacAnanny, E. Progesterone activates a progesterone receptor membrane component 1-dependent mechanism that promotes human granulosa/luteal cell survival but not progesterone secretion. J. Clin. Endocrinol. Metab. 2009, 94, 2644–2649. [Google Scholar] [CrossRef]
- Gellersen, B.; Fernandes, M.S.; Brosens, J.J. Non-genomic progesterone actions in female reproduction. Hum. Reprod. Update 2009, 15, 119–138. [Google Scholar] [CrossRef] [Green Version]
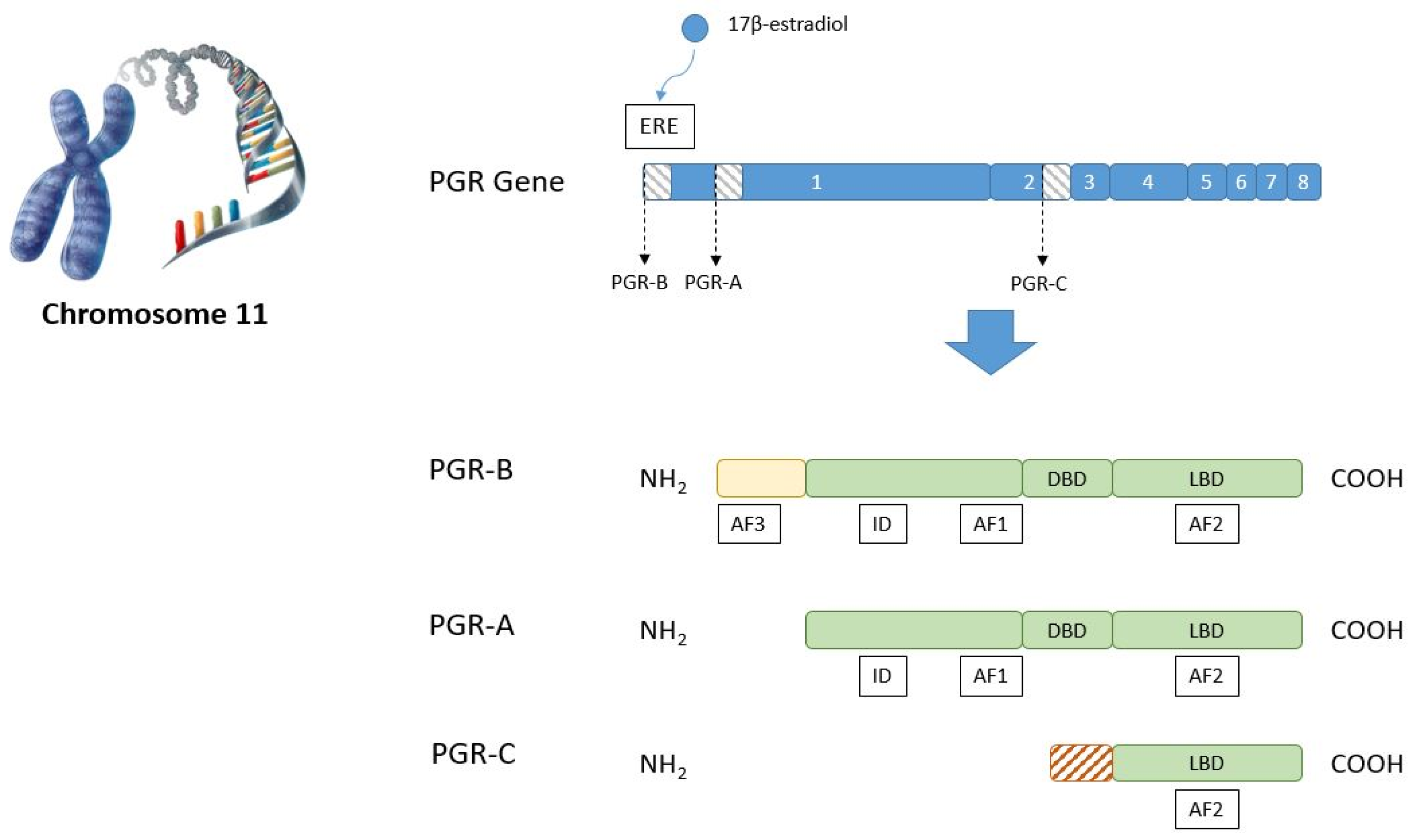
- Misrahi, M.; Venencie, P.Y.; Saugier-Veber, P.; Sar, S.; Dessen, P.; Milgrom, E. Structure of the human progesterone receptor gene. Biochim. Biophys. Acta Gene Struct. Expr. 1993, 1216, 289–292. [Google Scholar] [CrossRef]
- Jacobsen, B.M.; Horwitz, K.B. Progesterone receptors, their isoforms and progesterone regulated transcription. Mol. Cell. Endocrinol. 2012, 357, 18–29. [Google Scholar] [CrossRef] [Green Version]
- Hill, K.K.; Roemer, S.C.; Churchill, M.E.A.; Edwards, D.P. Structural and Functional Analysis of Domains of the Progesterone Receptor. Mol. Cell. Endocrinol. 2012, 348, 418. [Google Scholar] [CrossRef] [Green Version]
- Wei, L.L.; Hawkins, P.; Baker, C.; Norris, B.; Sheridan, P.L.; Quinn, P.G. An amino-terminal truncated progesterone receptor isoform, PRc, enhances progestin-induced transcriptional activity. Mol. Endocrinol. 1996, 10, 1379–1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scarpin, K.M.; Graham, J.D.; Mote, P.A.; Clarke, C.L. Progesterone action in human tissues: Regulation by progesterone receptor (PR) isoform expression, nuclear positioning and coregulator expression. Nucl. Recept. Signal. 2009, 7, nrs-07009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pieber, D.; Allport, V.C.; Bennett, P.R. Progesterone receptor isoform A inhibits isoform B-mediated transactivation in human amnion. Eur. J. Pharmacol. 2001, 427, 7–11. [Google Scholar] [CrossRef]
- Untitled. Available online: https://app.biorender.com/illustrations/6140d7d4a43e4400a5a10e7d (accessed on 15 September 2021).
- Gerdes, D.; Wehling, M.; Leube, B.; Falkenstein, E. Cloning and tissue expression of two putative steroid membrane receptors. Biol. Chem. 1998, 379, 907–911. [Google Scholar] [CrossRef]
- Saner, K.J.; Welter, B.H.; Zhang, F.; Hansen, E.; Dupont, B.; Wei, Y.; Price, T.M. Cloning and expression of a novel, truncated, progesterone receptor. Mol. Cell. Endocrinol. 2003, 200, 155–163. [Google Scholar] [CrossRef]
- Migliaccio, A.; Piccolo, D.; Castoria, G.; Di Domenico, M.; Bilancio, A.; Lombardi, M.; Gong, W.; Beato, M.; Auricchio, F. Activation of the Src/p21ras/Erk pathway by progesterone receptor via cross-talk with estrogen receptor. EMBO J. 1998, 17, 2008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierson-Mullany, L.K.; Lange, C.A. Phosphorylation of Progesterone Receptor Serine 400 Mediates Ligand-Independent Transcriptional Activity in Response to Activation of Cyclin-Dependent Protein Kinase 2. Mol. Cell. Biol. 2004, 24, 10542–10557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Y.; Hu, T.; Arterburn, M.; Boyle, B.; Bright, J.; Emtage, P.; Funk, W. PAQR proteins: A novel membrane receptor family defined by an ancient 7-transmembrane pass motif. J. Mol. Evol. 2005, 61, 372–380. [Google Scholar] [CrossRef]
- Peluso, J.J. Non-genomic actions of progesterone in the normal and neoplastic mammalian ovary. Semin. Reprod. Med. 2007, 25, 198–207. [Google Scholar] [CrossRef]
- Thomas, P. Characteristics of membrane progestin receptor alpha (mPRα) and progesterone membrane receptor component one (PGMRC1) and their roles in mediating rapid progestin actions. Front. Neuroendocrinol. 2008, 29, 292. [Google Scholar] [CrossRef] [Green Version]
- Thomas, P.; Pang, Y. Membrane progesterone receptors: Evidence for neuroprotective, neurosteroid signaling and neuroendocrine functions in neuronal cells. Neuroendocrinology 2012, 96, 162–171. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Bond, J.; Thomas, P. Identification, classification, and partial characterization of genes in humans and other vertebrates homologous to a fish membrane progestin receptor. Proc. Natl. Acad. Sci. USA 2003, 100, 2237–2242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karteris, E.; Zervou, S.; Pang, Y.; Dong, J.; Hillhouse, E.; Randeva, H.; Thomas, P. Progesterone signaling in human myometrium through two novel membrane G protein-coupled receptors: Potential role in functional progesterone withdrawal at term. Mol. Endocrinol. 2006, 20, 1519–1534. [Google Scholar] [CrossRef] [Green Version]
- Falkenstein, E.; Meyer, C.; Eisen, C.; Scriba, P.; Wehling, M. Full-length cDNA sequence of a progesterone membrane-binding protein from porcine vascular smooth muscle cells. Biochem. Biophys. Res. Commun. 1996, 229, 86–89. [Google Scholar] [CrossRef]
- Meyer, C.; Schmid, R.; Scriba, P.C.; Wehling, M. Purification and Partial Sequencing of High-Affinity Progesterone-Binding Site(s) from Porcine Liver Membranes. Eur. J. Biochem. 1996, 239, 726–731. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.; Pang, Y.; Dong, J. Enhancement of cell surface expression and receptor functions of membrane progestin receptor α (mPRα) by progesterone receptor membrane component 1 (PGRMC1): Evidence for a role of PGRMC1 as an adaptor protein for steroid receptors. Endocrinology 2014, 155, 1107–1119. [Google Scholar] [CrossRef] [PubMed]
- Sueldo, C.; Liu, X.; Peluso, J. Progestin and AdipoQ Receptor 7, Progesterone Membrane Receptor Component 1 (PGRMC1), and PGRMC2 and Their Role in Regulating Progesterone’s Ability to Suppress Human Granulosa/Luteal Cells from Entering into the Cell Cycle. Biol. Reprod. 2015, 93, 63. [Google Scholar] [CrossRef] [Green Version]
- Krietsch, T.; Fernandes, M.S.; Kero, J.; Lösel, R.; Heyens, M.; Lam, E.W.F.; Huhtaniemi, I.; Brosens, J.J.; Gellersen, B. Human homologs of the putative G protein-coupled membrane progestin receptors (mPRα, β, and γ) localize to the endoplasmic reticulum and are not activated by progesterone. Mol. Endocrinol. 2006, 20, 3146–3164. [Google Scholar] [CrossRef] [Green Version]
- Frye, C.; Sumida, K.; Dudek, B.; Harney, J.; Lydon, J.; O’Malley, B.; Pfaff, D.; Rhodes, M. Progesterone’s effects to reduce anxiety behavior of aged mice do not require actions via intracellular progestin receptors. Psychopharmacology 2006, 186, 312–322. [Google Scholar] [CrossRef]
- Finidori-Lepicard, J.; Schorderet-Slatkine, S.; Hanoune, J.; Baulieu, E.-E. Progesterone inhibits membrane-bound adenylate cyclase in Xenopus laevis oocytes. Nature 1981, 292, 255–257. [Google Scholar] [CrossRef]
- Foresta, C.; Rossato, M.; Di Virgilio, F. Ion fluxes through the progesterone-activated channel of the sperm plasma membrane. Biochem. J. 1993, 294, 279–283. [Google Scholar] [CrossRef] [Green Version]
- Viéro, C.; Méchaly, I.; Aptel, H.; Puech, S.; Valmier, J.; Bancel, F.; Dayanithi, G. Rapid inhibition of Ca2+ influx by neurosteroids in murine embryonic sensory neurones. Cell Calcium 2006, 40, 383–391. [Google Scholar] [CrossRef]
- Barbagallo, M.; Dominguez, L.; Licata, G.; Shan, J.; Bing, L.; Karpinski, E.; Pang, P.; Resnick, L. Vascular Effects of Progesterone: Role of Cellular Calcium Regulation. Hypertension 2001, 37, 142–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Head, G.; Downing, J.; Brucker, C.; Mentlein, R.; Kendall, M. Rapid progesterone actions on thymulin-secreting epithelial cells cultured from rat thymus. Neuroimmunomodulation 1999, 6, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Scott, R.E.M.; Wu-Peng, X.-S.; Pfaff, D.W. Regulation and Expression of Progesterone Receptor mRNA Isoforms A and B in the Male and Female Rat Hypothalamus and Pituitary Following Oestrogen Treatment. J. Neuroendocrinol. 2002, 14, 175–183. [Google Scholar] [CrossRef]
- Bashour, N.M.; Wray, S. Progesterone Directly and Rapidly Inhibits GnRH Neuronal Activity via Progesterone Receptor Membrane Component 1. Endocrinology 2012, 153, 4457–4469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuloaga, D.G.; Yahn, S.L.; Pang, Y.; Quihuis, A.M.; Oyola, M.G.; Reyna, A.; Thomas, P.; Handa, R.J.; Mani, S.K. Distribution and estrogen regulation of membrane progesterone receptor-β in the female rat brain. Endocrinology 2012, 153, 4432–4443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Revelli, A.; Paahioni, D.; Cassoni, P.; Bussolati, G.; Massobrio, M. In situ hybridization study of messenger RNA for estrogen receptor and immunohistochemical detection of estrogen and progesterone receptors in the human ovary. Gynecol. Endocrinol. 1996, 10, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Griffin, D.; Liu, X.; Pru, C.; Pru, J.; Peluso, J. Expression of progesterone receptor membrane component-2 within the immature rat ovary and its role in regulating mitosis and apoptosis of spontaneously immortalized granulosa cells. Biol. Reprod. 2014, 91, 36. [Google Scholar] [CrossRef]
- Will, E.; Liu, X.; Peluso, J. AG 205, a progesterone receptor membrane component 1 antagonist, ablates progesterone’s ability to block oxidative stress-induced apoptosis of human granulosa/luteal cells†. Biol. Reprod. 2017, 96, 843–854. [Google Scholar] [CrossRef] [Green Version]
- Dressing, G.; Pang, Y.; Dong, J.; Thomas, P. Progestin signaling through mPRα in Atlantic croaker granulosa/theca cell cocultures and its involvement in progestin inhibition of apoptosis. Endocrinology 2010, 151, 5916–5926. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.; Zhang, C.; Wang, Y.; Feng, L.; Wang, Z.; Niu, W.; Du, X.; Tang, W.; Li, Y.; Wang, C.; et al. Progesterone Receptor Membrane Component 1 Mediates Progesterone-Induced Suppression of Oocyte Meiotic Prophase I and Primordial Folliculogenesis. Sci. Rep. 2016, 6, 36869. [Google Scholar] [CrossRef]
- Peluso, J.J.; Griffin, D.; Liu, X.; Horne, M. Progesterone Receptor Membrane Component-1 (PGRMC1) and PGRMC-2 Interact to Suppress Entry into the Cell Cycle in Spontaneously Immortalized Rat Granulosa Cells1. Biol. Reprod. 2014, 91, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peluso, J.J.; Pru, C.; Liu, X.; Kelp, N.; Pru, J.K. Progesterone Receptor Membrane Component 1 and 2 regulate granulosa cell mitosis and survival through a NF?B-dependent mechanism John. Biol. Reprod. 2019, 100, 1571–1580. [Google Scholar] [CrossRef]
- Skiadas, C.; Duan, S.; Correll, M.; Rubio, R.; Karaca, N.; Ginsburg, E.; Quackenbush, J.; Racowsky, C. Ovarian reserve status in young women is associated with altered gene expression in membrana granulosa cells. Mol. Hum. Reprod. 2012, 18, 362–371. [Google Scholar] [CrossRef] [Green Version]
- Schuster, J.; Karlsson, T.; Karlström, P.; Poromaa, I.; Dahl, N. Down-regulation of progesterone receptor membrane component 1 (PGRMC1) in peripheral nucleated blood cells associated with premature ovarian failure (POF) and polycystic ovary syndrome (PCOS). Reprod. Biol. Endocrinol. 2010, 8, 58. [Google Scholar] [CrossRef] [Green Version]
- Mansouri, M.; Schuster, J.; Badhai, J.; Stattin, E.; Lösel, R.; Wehling, M.; Carlsson, B.; Hovatta, O.; Karlström, P.; Golovleva, I.; et al. Alterations in the expression, structure and function of progesterone receptor membrane component-1 (PGRMC1) in premature ovarian failure. Hum. Mol. Genet. 2008, 17, 3776–3783. [Google Scholar] [CrossRef] [PubMed]
- Conneely, I.; Mulac-Jericevic, B.; DeMayo, F.; Lydon, J.; O’Malley, B. Reproductive functions of progesterone receptors. Recent Prog. Horm. Res. 2002, 57, 339–355. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.-J.; Zhu, Y. Downregulation of nuclear progestin receptor (Pgr) and subfertility in double knockouts of progestin receptor membrane component 1 (pgrmc1) and pgrmc2 in zebrafish. Gen. Comp. Endocrinol. 2020, 285, 113275. [Google Scholar] [CrossRef]
- Hasegawa, J.; Yanaihara, A.; Iwasaki, S.; Otsuka, Y.; Negishi, M.; Akahane, T.; Okai, T. Reduction of progesterone receptor expression in human cumulus cells at the time of oocyte collection during IVF is associated with good embryo quality. Hum. Reprod. 2005, 20, 2194–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lodde, V.; Peluso, J. A novel role for progesterone and progesterone receptor membrane component 1 in regulating spindle microtubule stability during rat and human ovarian cell mitosis. Biol. Reprod. 2011, 84, 715–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terzaghi, L.; Tessaro, I.; Raucci, F.; Merico, V.; Mazzini, G.; Garagna, S.; Zuccotti, M.; Franciosi, F.; Lodde, V. PGRMC1 participates in late events of bovine granulosa cells mitosis and oocyte meiosis. Cell Cycle 2016, 15, 2019–2032. [Google Scholar] [CrossRef] [Green Version]
- Jühlen, R.; Landgraf, D.; Huebner, A.; Koehler, K. Identification of a novel putative interaction partner of the nucleoporin ALADIN. Biol. Open 2016, 5, 1697–1705. [Google Scholar] [CrossRef] [Green Version]
- Hazano, K.; Haneda, S.; Kayano, M.; Matsui, M. Local sex steroid hormone milieu in the bovine oviduct ipsilateral and contralateral to preovulatory follicle or corpus luteum during the periovulatory phase. Domest. Anim. Endocrinol. 2021, 74, 106515. [Google Scholar] [CrossRef] [PubMed]
- Saint-Dizier, M.; Sandra, O.; Ployart, S.; Chebrout, M.; Constant, F. Expression of nuclear progesterone receptor and progesterone receptor membrane components 1 and 2 in the oviduct of cyclic and pregnant cows during the post-ovulation period. Reprod. Biol. Endocrinol. 2012, 10, 76. [Google Scholar] [CrossRef] [PubMed]
- Kowalik, M.K.; Martyniak, M.; Rekawiecki, R.; Kotwica, J. Expression and immunolocalization of membrane progesterone receptors in the bovine oviduct. Domest. Anim. Endocrinol. 2016, 55, 83–96. [Google Scholar] [CrossRef]
- Tahir, M.; Reynaud, K.; Grimard, B.; Thoumire, S.; Chastant-Maillard, S.; Saint-Dizier, M. Expression of nuclear and membrane progesterone receptors in the canine oviduct during the periovulatory period. Reprod. Fertil. Dev. 2013, 25, 1065–1076. [Google Scholar] [CrossRef]
- Petousis, S.; Prapas, Y.; Margioula-Siarkou, C.; Ravanos, K.; Milias, S.; Mavromatidis, G.; Kalogiannidis, I.; Haitoglou, C.; Athanasiadis, A.; Prapas, N.; et al. Unexplained infertility patients present the mostly impaired levels of progesterone receptors: Prospective observational study. Am. J. Reprod. Immunol. 2018, 79, e12828. [Google Scholar] [CrossRef]
- Hosseinirad, H.; Novin, M.; Hosseini, S.; Nazarian, H.; Safaei, Z.; Hashemi, T.; Paktinat, S.; Mofarahe, Z. Evaluation of Expression and Phosphorylation of Progesterone Receptor in Endometrial Stromal Cells of Patients with Recurrent Implantation Failure Compared to Healthy Fertile Women. Reprod. Sci. 2021, 28, 1457–1465. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, M.S.; Pierron, V.; Michalovich, D.; Astle, S.; Thornton, S.; Peltoketo, H.; Lam, E.W.F.; Gellersen, B.; Huhtaniemi, I.; Allen, J.; et al. Regulated expression of putative membrane progestin receptor homologues in human endometrium and gestational tissues. J. Endocrinol. 2005, 187, 89–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salsano, S.; Quiñonero, A.; Pérez, S.; Garrido Gómez, T.; Simón, C.; Dominguez, F. Dynamic expression of PGRMC1 and SERBP1 in human endometrium: An implication in the human decidualization process. Fertil. Steril. 2017, 108, 832–842.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mccallum, M.L.; Pru, C.A.; Niikura, Y.; Yee, S.P.; Lydon, J.P.; Peluso, J.J.; Pru, J.K. Conditional ablation of progesterone receptor membrane component 1 results in subfertility in the female and development of endometrial cysts. Endocrinology 2016, 157, 3309–3319. [Google Scholar] [CrossRef] [Green Version]
- Garrido-Gómez, T.; Quiñonero, A.; Antúnez, O.; Díaz-Gimeno, P.; Bellver, J.; Simón, C.; Domínguez, F. Deciphering the proteomic signature of human endometrial receptivity. Hum. Reprod. 2014, 29, 1957–1967. [Google Scholar] [CrossRef] [Green Version]
- Gemzell-Danielsson, K.; Swahn, M.-L.; Svalander, P.; Bygdeman, M. Early luteal phase treatment with mifepristone (RU 486) for fertility regulation. Hum. Reprod. 1993, 8, 870–873. [Google Scholar] [CrossRef]
- Avrech, O.M.; Golan, A.; Weinraub, Z.; Bukovsky, I.; Caspi, E. Mifepristone (RU486) alone or in combination with a prostaglandin analogue for termination of early pregnancy: A review. Fertil. Steril. 1991, 56, 385–393. [Google Scholar] [CrossRef]
- Wu, W.; Shi, S.Q.; Huang, H.J.; Balducci, J.; Garfield, R.E. Changes in PGRMC1, a potential progesterone receptor, in human myometrium during pregnancy and labour at term and preterm. Mol. Hum. Reprod. 2011, 17, 233–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ackerman, W.E., IV; Summerfield, T.L.; Mesiano, S.; Schatz, F.; Lockwood, C.J.; Kniss, D.A. Agonist-Dependent Downregulation of Progesterone Receptors in Human Cervical Stromal Fibroblasts. Reprod. Sci. 2016, 23, 112. [Google Scholar] [CrossRef] [Green Version]
- Akison, L.; Robker, R. The critical roles of progesterone receptor (PGR) in ovulation, oocyte developmental competence and oviductal transport in mammalian reproduction. Reprod. Domest. Anim. 2012, 47, 288–296. [Google Scholar] [CrossRef]
- Pelosi, E.; Forabosco, A.; Schlessinger, D. Genetics of the ovarian reserve. Front. Genet. 2015, 6, 308. [Google Scholar] [CrossRef]
- Peluso, J.; Liu, X.; Uliasz, T.; Pru, C.; Kelp, N.; Pru, J. PGRMC1/2 promotes luteal vascularization and maintains the primordial follicles of mice. Reproduction 2018, 156, 365–373. [Google Scholar] [CrossRef] [Green Version]
- Robker, R.L.; Richards, J.S. Progesterone: Lessons from the Progesterone Receptor Knockout. Ovulation 2000, 121–129. [Google Scholar] [CrossRef]
- Peluso, J.; Pru, J. Non-canonical progesterone signaling in granulosa cell function. Reproduction 2014, 147, 169–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lösel, R.; Besong, D.; Peluso, J.; Wehling, M. Progesterone receptor membrane component 1—Many tasks for a versatile protein. Steroids 2008, 73, 929–934. [Google Scholar] [CrossRef]
- Friel, A.; Zhang, L.; Pru, C.; Clark, N.; McCallum, M.; Blok, L.; Shioda, T.; Peluso, J.; Rueda, B.; Pru, K.; et al. Progesterone receptor membrane component 1 deficiency attenuates growth while promoting chemosensitivity of human endometrial xenograft tumors. Cancer Lett. 2015, 356, 434–442. [Google Scholar] [CrossRef] [Green Version]
- Luciano, A.; Lodde, V.; Franciosi, F.; Ceciliani, F.; Peluso, J. Progesterone receptor membrane component 1 expression and putative function in bovine oocyte maturation, fertilization, and early embryonic development. Reproduction 2010, 140, 663–672. [Google Scholar] [CrossRef] [Green Version]
- Elassar, A.; Liu, X.; Scranton, V.; Wu, C.; Peluso, J. The relationship between follicle development and progesterone receptor membrane component-1 expression in women undergoing in vitro fertilization. Fertil. Steril. 2012, 97, 572–578. [Google Scholar] [CrossRef] [Green Version]
- Vaitsopoulou, C.I.; Kolibianakis, E.M.; Bosdou, J.K.; Neofytou, E.; Lymperi, S.; Makedos, A.; Savvaidou, D.; Chatzimeletiou, K.; Grimbizis, G.F.; Lambropoulos, A.; et al. Expression of genes that regulate follicle development and maturation during ovarian stimulation in poor responders. Reprod. Biomed. Online 2021, 42, 248–259. [Google Scholar] [CrossRef]
- Fujiwara, H.; Maeda, M.; Honda, T.; Yamada, S.; Ueda, M.; Kanzaki, H.; Suginami, H.; Mori, T. Granulosa cells express integrin alpha 6: Possible involvement of integrin alpha 6 in folliculogenesis. Horm. Res. 1996, 46 (Suppl. S1), 24–30. [Google Scholar] [CrossRef] [PubMed]
- Bridges, P.J.; Cho, J.; Ko, C. Endothelins in regulating ovarian and oviductal function. Front. Biosci. Schol. Ed. 2011, 3, 145. [Google Scholar] [PubMed] [Green Version]
- Ashkenazi, H.; Cao, X.; Motola, S.; Popliker, M.; Conti, M.; Tsafriri, A. Epidermal Growth Factor Family Members: Endogenous Mediators of the Ovulatory Response. Endocrinology 2005, 146, 77–84. [Google Scholar] [CrossRef] [Green Version]
- Azziz, R. Polycystic Ovary Syndrome. Obstet. Gynecol. 2018, 132, 321–336. [Google Scholar] [CrossRef]
- Artimani, T.; Saidijam, M.; Aflatoonian, R.; Amiri, I.; Ashrafi, M.; Shabab, N.; Mohammadpour, N.; Mehdizadeh, M. Estrogen and progesterone receptor subtype expression in granulosa cells from women with polycystic ovary syndrome. Gynecol. Endocrinol. 2015, 31, 379–383. [Google Scholar] [CrossRef] [PubMed]
- Mehlmann, L.M. Stops and starts in mammalian oocytes: Recent advances in understanding the regulation of meiotic arrest and oocyte maturation. Reproduction 2005, 130, 791–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teilmann, S.C.; Clement, C.A.; Thorup, J.; Byskov, A.G.; Christensen, S.T. Expression and localization of the progesterone receptor in mouse and human reproductive organs. J. Endocrinol. 2006, 191, 525–535. [Google Scholar] [CrossRef]
- Deng, J.; Carbajal, L.; Evaul, K.; Rasar, M.; Jamnongjit, M.; Hammes, S.R. Nongenomic Steroid-Triggered Oocyte Maturation: Of Mice and Frogs. Steroids 2009, 74, 595. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, A.; Gentilello, A.; Balboula, A.; Shrivastava, V.; Ohring, J.; Schindler, K. Phosphorylation of threonine 3 on histone H3 by haspin kinase is required for meiosis I in mouse oocytes. J. Cell Sci. 2014, 127, 5066–5078. [Google Scholar] [CrossRef] [Green Version]
- Bonner, M.K.; Poole, D.S.; Xu, T.; Sarkeshik, A.; Yates, J.R., III; Skop, A.R. Mitotic Spindle Proteomics in Chinese Hamster Ovary Cells. PLoS ONE 2011, 6, e20489. [Google Scholar] [CrossRef] [Green Version]
- Nousiainen, M.; Silljé, H.H.W.; Sauer, G.; Nigg, E.A.; Körner, R. Phosphoproteome analysis of the human mitotic spindle. Proc. Natl. Acad. Sci. USA 2006, 103, 5391–5396. [Google Scholar] [CrossRef] [Green Version]
- Carvalhal, S.; Ribeiro, S.A.; Arocena, M.; Kasciukovic, T.; Temme, A.; Koehler, K.; Huebner, A.; Griffis, E.R. The nucleoporin ALADIN regulates Aurora A localization to ensure robust mitotic spindle formation. Mol. Biol. Cell 2015, 26, 3424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalhal, S.; Stevense, M.; Koehler, K.; Naumann, R.; Huebner, A.; Jessberger, R.; Griffis, E. ALADIN is required for the production of fertile mouse oocytes. Mol. Biol. Cell 2017, 28, 2470–2478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, B.; Liu, X.; Thomas, P.; Pang, Y.; Xu, Y.; Li, X.; Li, X. Identification and characterization of a progestin and adipoQ receptor (PAQR) structurally related to Paqr7 in the ovary of Cynoglossus semilaevis and its potential role in regulating oocyte maturation. Gen. Comp. Endocrinol. 2016, 237, 109–120. [Google Scholar] [CrossRef]
- Thomas, P.; Pang, Y.; Zhu, Y.; Detweiler, C.; Doughty, K. Multiple rapid progestin actions and progestin membrane receptor subtypes in fish. Steroids 2004, 69, 567–573. [Google Scholar] [CrossRef]
- Roy, S.; Wang, J.; Rana, M.; Nakashima, M.; Tokumoto, T. Characterization of membrane progestin receptor α (mPRα) of the medaka and role in the induction of oocyte maturation. Biomed. Res. 2017, 38, 79–87. [Google Scholar] [CrossRef] [Green Version]
- Mahmood, T.; Saridogan, E.; Smutna, S.; Habib, A.M.; Djahanbakhch, O. The effect of ovarian steroids on epithelial ciliary beat frequency in the human Fallopian tube. Hum. Reprod. 1998, 13, 2991–2994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wånggren, K.; Stavreus-Evers, A.; Olsson, C.; Andersson, E.; Gemzell-Danielsson, K. Regulation of muscular contractions in the human Fallopian tube through prostaglandins and progestagens. Hum. Reprod. 2008, 23, 2359–2368. [Google Scholar] [CrossRef] [Green Version]
- McDonald, M.; Bellve, A. Influence of oestrogen and progesterone on flow of fluid from the Fallopian tube in the ovariectomized ewe. J. Reprod. Fertil. 1969, 20, 51–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, B.; Elguero, S.; Thakore, S.; Dahoud, W.; Bedaiwy, M.; Mesiano, S. Role of nuclear progesterone receptor isoforms in uterine pathophysiology. Hum. Reprod. Update 2015, 21, 155–173. [Google Scholar] [CrossRef] [Green Version]
- Borini, A.; Bafaro, G.; Violini, F.; Bianchi, L.; Casadio, V.; Flamigni, C. Pregnancies in postmenopausal women over 50 years old in an oocyte donation program. Fertil. Steril. 1995, 63, 258–261. [Google Scholar] [CrossRef]
- Salsano, S.; González-Martín, R.; Quiñonero, A.; Pérez-Debén, S.; Domínguez, F. Deciphering the Role of PGRMC1 During Human Decidualization Using an In Vitro Approach. J. Clin. Endocrinol. Metab. 2021, 106, 2313–2327. [Google Scholar] [CrossRef]
- Salsano, S.; González-Martín, R.; Quiñonero, A.; López-Martín, S.; Gómez-Escribano, A.; Pérez-Debén, S.; Yañez-Mo, M.; Domínguez, F. Novel nonclassic progesterone receptor PGRMC1 pulldown-precipitated proteins reveal a key role during human decidualization. Fertil. Steril. 2020, 113, 1050–1066.e7. [Google Scholar] [CrossRef]
- Zhang, L.; Kanda, Y.; Roberts, D.J.; Ecker, J.L.; Losel, R.; Wehling, M.; Peluso, J.J.; Pru, J.K. Expression of progesterone receptor membrane component 1 and its partner serpine 1 mRNA binding protein in uterine and placental tissues of the mouse and human. Mol. Cell. Endocrinol. 2008, 287, 81–89. [Google Scholar] [CrossRef]
- Kowalik, M.; Dobrzyn, K.; Rekawiecki, R.; Kotwica, J. Expression of membrane progestin receptors (mPRs) α, β and γ in the bovine uterus during the oestrous cycle and pregnancy. Theriogenology 2019, 140, 171–179. [Google Scholar] [CrossRef]
- Pru, J.K.; Clark, N.C.; Keator, C.S. PGRMC1 and PGRMC2 in uterine physiology and disease. Front. Neurosci. 2013, 7, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Williams, M.; Patel, P.; Kew, K.; Zhu, Y. Subfertility and reduced progestin synthesis in Pgrmc2 knockout zebrafish. Gen. Comp. Endocrinol. 2019, 282, 113218. [Google Scholar] [CrossRef] [PubMed]
- Keator, C.S.; Mah, K.; Slayden, O.D. Alterations in progesterone receptor membrane component 2 (PGRMC2) in the endometrium of macaques afflicted with advanced endometriosis. Mol. Hum. Reprod. 2012, 18, 308–319. [Google Scholar] [CrossRef]
- Mote, P.A.; Balleine, R.L.; Mcgowan, E.M.; Clarke, C.L. Heterogeneity of progesterone receptors A and B expression in human endometrial glands and stroma. Hum. Reprod. 2000, 15, 48–56. [Google Scholar] [CrossRef] [Green Version]
- Mesiano, S. Myometrial progesterone responsiveness. Semin. Reprod. Med. 2007, 25, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Cowan, S.; Calder, A.; Kelly, R. Decidualisation of cervical stromal cells. Eur. J. Obstet. Gynecol. Reprod. Biol. 2004, 114, 189–196. [Google Scholar] [CrossRef]
- Zhu, H.; Hou, C.; Luo, L.; Hu, Y.; Yang, W. Endometrial stromal cells and decidualized stromal cells: Origins, transformation and functions. Gene 2014, 551, 1–14. [Google Scholar] [CrossRef]
- Wetendorf, M.; DeMayo, F. The progesterone receptor regulates implantation, decidualization, and glandular development via a complex paracrine signaling network. Mol. Cell. Endocrinol. 2012, 357, 108–118. [Google Scholar] [CrossRef] [Green Version]
- Jabbour, H.; Kelly, R.; Fraser, H.; Critchley, H. Endocrine regulation of menstruation. Endocr. Rev. 2006, 27, 17–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.; Li, R.; DeMayo, F. Progesterone Receptor Regulation of Uterine Adaptation for Pregnancy. Trends Endocrinol. Metab. 2018, 29, 481–491. [Google Scholar] [CrossRef] [PubMed]
- Mulac-Jericevic, B.; Mullinax, R.; DeMayo, F.; Lydon, J.; Conneely, O. Subgroup of reproductive functions of progesterone mediated by progesterone receptor-B isoform. Science 2000, 289, 1751–1754. [Google Scholar] [CrossRef]
- Fleisch, M.; Chou, Y.; Cardiff, R.; Asaithambi, A.; Shyamala, G. Overexpression of progesterone receptor A isoform in mice leads to endometrial hyperproliferation, hyperplasia and atypia. Mol. Hum. Reprod. 2009, 15, 241–249. [Google Scholar] [CrossRef] [Green Version]
- Su, M.-T.; Lee, I.-W.; Chen, Y.-C.; Kuo, P.-L. Association of progesterone receptor polymorphism with idiopathic recurrent pregnancy loss in Taiwanese Han population. J. Assist. Reprod. Genet. 2011, 28, 239. [Google Scholar] [CrossRef] [Green Version]
- Pisarska, M.D.; Carson, S.A.; Casson, P.R.; Tong, X.; Buster, J.E.; Kieback, D.G. A mutated progesterone receptor allele is more prevalent in unexplained infertility. Fertil. Steril. 2003, 80, 651–653. [Google Scholar] [CrossRef]
- Rahnama, R.; Rafiee, M.; Fouladi, S.; Akbari-Fakhrabadi, M.; Mehrabian, F.; Rezaei, A. Gene expression analysis of membrane progesterone receptors in women with recurrent spontaneous abortion: A case control study. BMC Res. Notes 2019, 12, 790. [Google Scholar] [CrossRef]
- Lyzikova, Y.A.; Zinovkin, D.A.; Pranjol, M.Z.I. Increase in FoxP3, CD56 immune cells and decrease in glands PGRMC1 expression in the endometrium are associated with recurrent miscarriages. Eur. J. Obstet. Gynecol. Reprod. Biol. 2020, 245, 121–126. [Google Scholar] [CrossRef]
- Wang, H.; Critchley, H.O.D.; Kelly, R.W.; Shen, D.; Baird, D.T. Progesterone receptor subtype B is differentially regulated in human endometrial stroma. Mol. Hum. Reprod. 1998, 4, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Deryabin, P.; Griukova, A.; Nikolsky, N.; Borodkina, A. The link between endometrial stromal cell senescence and decidualization in female fertility: The art of balance. Cell. Mol. Life Sci. 2020, 77, 1357–1370. [Google Scholar] [CrossRef] [PubMed]
- Lucas, E.; Vrljicak, P.; Muter, J.; Diniz-da-Costa, M.; Brighton, P.; Kong, C.; Lipecki, J.; Fishwick, K.; Odendaal, J.; Ewington, L.; et al. Recurrent pregnancy loss is associated with a pro-senescent decidual response during the peri-implantation window. Commun. Biol. 2020, 3, 37. [Google Scholar] [CrossRef] [Green Version]
- Clark, N.; Pru, C.; Yee, S.; Lydon, J.; Peluso, J.; Pru, J. Conditional Ablation of Progesterone Receptor Membrane Component 2 Causes Female Premature Reproductive Senescence. Endocrinology 2017, 158, 640–651. [Google Scholar] [CrossRef]
- Jiang, J.; Wu, R.F.; Wang, Z.H.; Sun, H.C.; Xu, Z.; Xiu, H.M. Effect of mifepristone on estrogen and progesterone receptors in human endometrial and endometriotic cells in vitro. Fertil. Steril. 2002, 77, 995–1000. [Google Scholar] [CrossRef]
- Bulun, S.; Cheng, Y.; Yin, P.; Imir, G.; Utsunomiya, H.; Attar, E.; Innes, J.; Julie Kim, J. Progesterone resistance in endometriosis: Link to failure to metabolize estradiol. Mol. Cell. Endocrinol. 2006, 248, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Lyndrup, J.; Thorpe, S.; Glenthej, A.; Obel, E.; Sele, V. Altered progesterone/estrogen receptor ratios in endometriosis: A comparative study of steroid receptors and morphology in endometriosis and endometrium. Acta Obstet. Gynecol. Scand. 1987, 66, 625–629. [Google Scholar] [CrossRef] [PubMed]
- Nisolle, M.; Casanas-Roux, F.; Wyns, C.; de Menten, Y.; Mathieu, P.; Donnez, J. Immunohistochemical analysis of estrogen and progesterone receptors in endometrium and peritoneal endometriosis: A new quantitative method. Fertil. Steril. 1994, 62, 751–759. [Google Scholar] [CrossRef]
- Rocha-Junior, C.; Da Broi, M.; Miranda-Furtado, C.; Navarro, P.; Ferriani, R.; Meola, J. Progesterone Receptor B (PGR-B) Is Partially Methylated in Eutopic Endometrium From Infertile Women With Endometriosis. Reprod. Sci. 2019, 26, 1568–1574. [Google Scholar] [CrossRef]
- Sletten, E.T.; Smaglyukova, N.; Ørbo, A.; Sager, G. Expression of nuclear progesterone receptors (nPRs), membrane progesterone receptors (mPRs) and progesterone receptor membrane components (PGRMCs) in the human endometrium after 6 months levonorgestrel low dose intrauterine therapy. J. Steroid Biochem. Mol. Biol. 2020, 202, 105701. [Google Scholar] [CrossRef]
- Vázquez-Martínez, E.; Bello-Alvarez, C.; Hermenegildo-Molina, A.; Solís-Paredes, M.; Parra-Hernández, S.; Cruz-Orozco, O.; Silvestri-Tomassoni, J.; Escobar-Ponce, L.; Hernández-López, L.; Reyes-Mayoral, C.; et al. Expression of Membrane Progesterone Receptors in Eutopic and Ectopic Endometrium of Women with Endometriosis. Biomed Res. Int. 2020, 2020, 2196024. [Google Scholar] [CrossRef] [PubMed]
- Kunz, G.; Beil, D.; Deininger, H.; Wudt, L.; Leyendecker, G. The dynamics of rapid sperm transport through the female genital tract: Evidence from vaginal sonography of uterine peristalsis and hysterosalpingoscintigraphy. Hum. Reprod. 1996, 11, 627–632. [Google Scholar] [CrossRef]
- Bygdeman, M.; Swahn, M.; Gemzell-Danielsson, K.; Svalander, P. Mode of action of RU 486. Ann. Med. 1993, 25, 61–64. [Google Scholar] [CrossRef]
- Astle, S.; Slater, D.M.; Thornton, S. The involvement of progesterone in the onset of human labour. Eur. J. Obstet. Gynecol. Reprod. Biol. 2003, 108, 177–181. [Google Scholar] [CrossRef]
- Brown, A.; Leite, R.; Strauss, J. Mechanisms underlying “functional” progesterone withdrawal at parturition. Ann. N. Y. Acad. Sci. 2004, 1034, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Mesiano, S.; Chan, E.; Fitter, J.; Kwek, K.; Yeo, G.; Smith, R. Progesterone withdrawal and estrogen activation in human parturition are coordinated by progesterone receptor A expression in the myometrium. J. Clin. Endocrinol. Metab. 2002, 87, 2924–2930. [Google Scholar] [CrossRef] [PubMed]
- Ruddock, N.K.; Shi, S.Q.; Jain, S.; Moore, G.; Hankins, G.D.V.; Romero, R.; Garfield, R.E. Progesterone, but not 17-alpha-hydroxyprogesterone caproate, inhibits human myometrial contractions. Am. J. Obstet. Gynecol. 2008, 199, 391.e1–391.e7. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Sheehan, P.; Brennecke, S. Changes in myometrial expression of progesterone receptor membrane components 1 and 2 are associated with human parturition at term. Reprod. Fertil. Dev. 2016, 28, 618–627. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, M.S.; Brosens, J.J.; Gellersen, B. Honey, we need to talk about the membrane progestin receptors. Steroids 2008, 73, 942–952. [Google Scholar] [CrossRef]
- Merlino, A.A.; Welsh, T.N.; Tan, H.; Yi, L.J.; Cannon, V.; Mercer, B.M.; Mesiano, S. Nuclear Progesterone Receptors in the Human Pregnancy Myometrium: Evidence that Parturition Involves Functional Progesterone Withdrawal Mediated by Increased Expression of Progesterone Receptor-A. J. Clin. Endocrinol. Metab. 2007, 92, 1927–1933. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Gonzalez, J.; Ofori, E.; Elovitz, M. Preventing cervical ripening: The primary mechanism by which progestational agents prevent preterm birth? Am. J. Obstet. Gynecol. 2008, 198, 314.e1–314.e8. [Google Scholar] [CrossRef] [PubMed]
- Hirai, Y.; Utsugi, K.; Takeshima, N.; Kawamata, Y.; Furuta, R.; Kitagawa, T.; Kawaguchi, T.; Hasumi, K.; Noda, S.T. Putative gene loci associated with carcinogenesis and metastasis of endocervical adenocarcinomas of uterus determined by conventional and array-based CGH. Am. J. Obstet. Gynecol. 2004, 191, 1173–1182. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Reproductive Tissue | Role | Study | Model | Type of PGR | Main Conclusions |
|---|---|---|---|---|---|
| OVARY and OVIDUCT | Gonadal neurological regulation | [69] | Rat | C | Increased PGR-A and PGR-B levels in hypothalamic and pituitary tissue after exogenous estradiol treatment |
| [70] | Mouse | NC | PGRMC1 is implicated in the P4 inhibition of GnRH neuronal activity | ||
| [71] | Rat | NC | mPRs expression has been detected in the hypothalamus | ||
| Follicular development and recruitment | [22] | Human | C | Increased expression in granulosa cells from preovulatory follicles during the periovulatory period, which suggests their role in ovulation, but not in follicular development and recruitment | |
| [72] | Human | C | Detected in a small proportion of human primordial and preantral follicles | ||
| [73,74] | Mouse | NC | PGRMC1 and PGRMC2 interfere with antral follicle development | ||
| [75] | A. croaker | NC | mPRα mediates the antiapoptotic actions of progestins in ovarian follicle cells | ||
| [76] | Mouse | NC | PGRMC1 mediates P4-induced suppression of oocyte development and primordial folliculogenesis | ||
| [77] | Mouse | NC | PGRMC1 and PGRMC2 interact to suppress entry into the cell cycle in spontaneously immortalized granulosa cells | ||
| [78] | Rat | NC | PGRMC1 and PGRMC2 regulate granulosa cell mitosis and survival through an NFkB-dependent mechanism | ||
| [79,80,81] | Human | NC | Lower PGRMC1 and PGRMC2 levels in women with premature ovarian insufficiency and decreased ovarian reserve | ||
| Ovulation | [82] | Mouse | C | Ovulation was completely absent in classical PGRKO mice, but it was only impaired in PGR-AKO mice and unaffected in PGR-BKO mice | |
| [83] | Zebrafish | NC | Lower metalloproteinase expression and ovulation rate in double PGRMC1/2KO zebrafish | ||
| Oocyte acquisition of competence | [84] | Human | C | No relation between the classical PGR expression in cumulus cells of IVF patients and oocyte fertilization or cleavage rate. Association between reduced classical PGR expression and good embryo quality | |
| [85] | Rat, Human | NC | PGRMC1 regulates spindle microtubule stability during rat and human ovarian cell mitosis | ||
| [86] | Bovine | NC | PGRMC1 participates in late mitosis and oocyte meiosis events and interacts with AURKB | ||
| [87] | Mouse | NC | PGRMC2 participates in meiotic spindle assembly with ALADIN | ||
| [37] | Bovine | NC | Role of mPRα in oocyte maturation and embryo development regulation | ||
| Oviduct transport | [88,89,90,91] | Bovine Canine | C and NC | Classical PGR, PGRMC1, and PGRMC2 expression in different parts along the oviduct | |
| UTERUS | Endometrium | [82] | Mouse | C | PGR-A expression, but not PGR-B, is sufficient for successful implantation and pregnancy |
| [92] | Human | C | Infertile patients with unexplained infertility with the lowest levels of endometrial epithelial expression of both PGR-A and PGR-B | ||
| [93] | Human | C | Association of recurrent implantation failure with decreased classical PGR expression in the endometrium | ||
| [11,94,95] | Human | C and NC | PGRMC1, mPRγ, and mPRε transcripts are up-regulated in the proliferative phase and progressively decrease in the secretory phase, whereas mPRα and PGRMC2 mRNA are significantly overexpressed in the secretory phase. mPRα do not significantly change during the menstrual cycle. | ||
| [96] | Mouse | NC | Conditional ablation of PGRMC1 results in female subfertility | ||
| [97] | Human | NC | PGRMC1 behaves differently in a receptive vs. a non-receptive endometrium | ||
| Myometrium | [98,99] | Human | C | P4 action via these receptors regulates myometrium contractile activity | |
| [57,94,100] | Human | NC | Presence of mPRα, mPRβ, PGRMC1, and PGRMC2 in myometrial cells | ||
| Cervix | [101] | Human | C | Expressed in stromal fibroblasts and basal squamous epithelial cells of the cervix |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Medina-Laver, Y.; Rodríguez-Varela, C.; Salsano, S.; Labarta, E.; Domínguez, F. What Do We Know about Classical and Non-Classical Progesterone Receptors in the Human Female Reproductive Tract? A Review. Int. J. Mol. Sci. 2021, 22, 11278. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011278
Medina-Laver Y, Rodríguez-Varela C, Salsano S, Labarta E, Domínguez F. What Do We Know about Classical and Non-Classical Progesterone Receptors in the Human Female Reproductive Tract? A Review. International Journal of Molecular Sciences. 2021; 22(20):11278. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011278
Chicago/Turabian StyleMedina-Laver, Yassmin, Cristina Rodríguez-Varela, Stefania Salsano, Elena Labarta, and Francisco Domínguez. 2021. "What Do We Know about Classical and Non-Classical Progesterone Receptors in the Human Female Reproductive Tract? A Review" International Journal of Molecular Sciences 22, no. 20: 11278. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222011278