
Irradiation Activates MZF1 to Inhibit miR-541-5p Expression and Promote Epithelial-Mesenchymal Transition (EMT) in Radiation-Induced Pulmonary Fibrosis (RIPF) by Upregulating Slug

 and
and
Abstract
:1. Introduction
2. Results
2.1. Ionizing Radiation Can Reduce the Expression of miR-541-5p
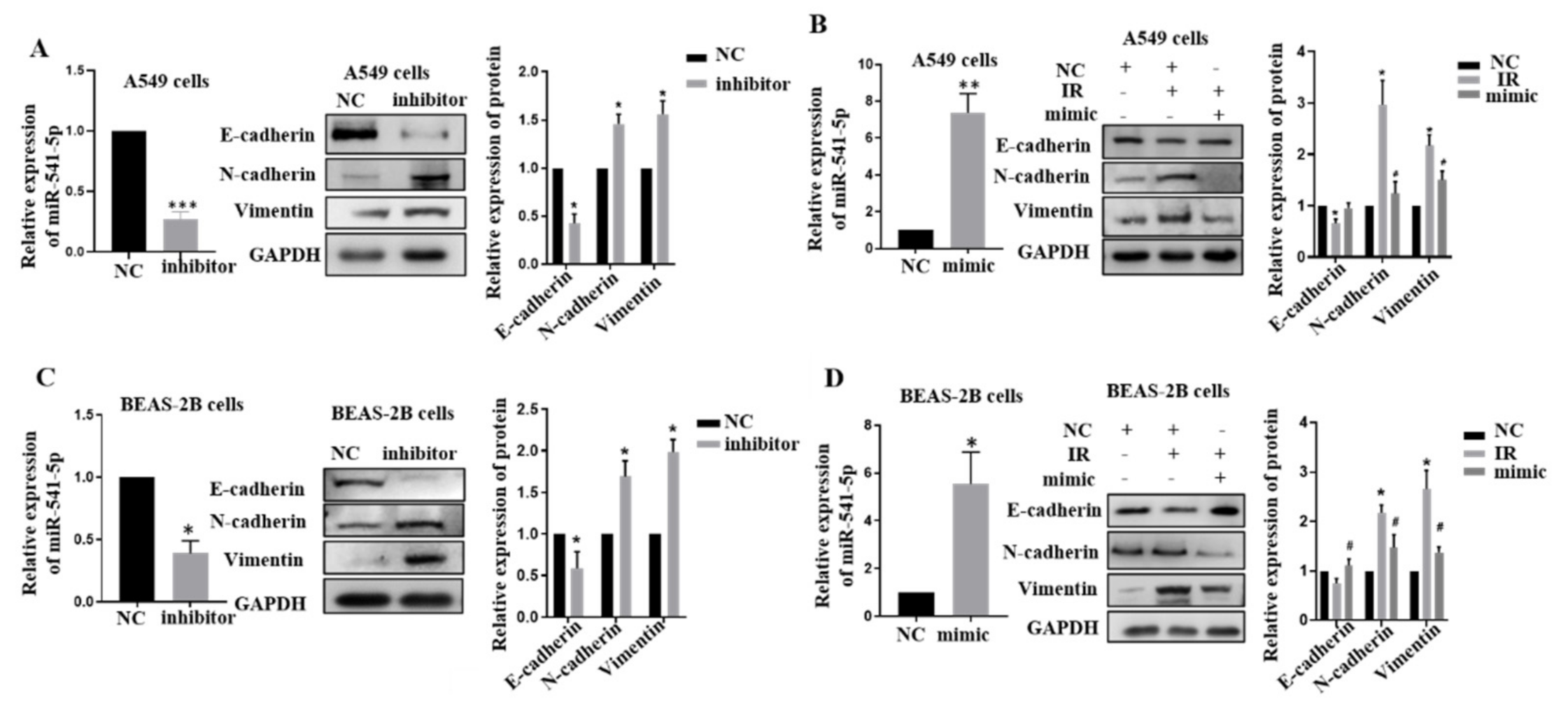
2.2. Knockdown of miR-541-5p Could Promote the Occurrence of EMT
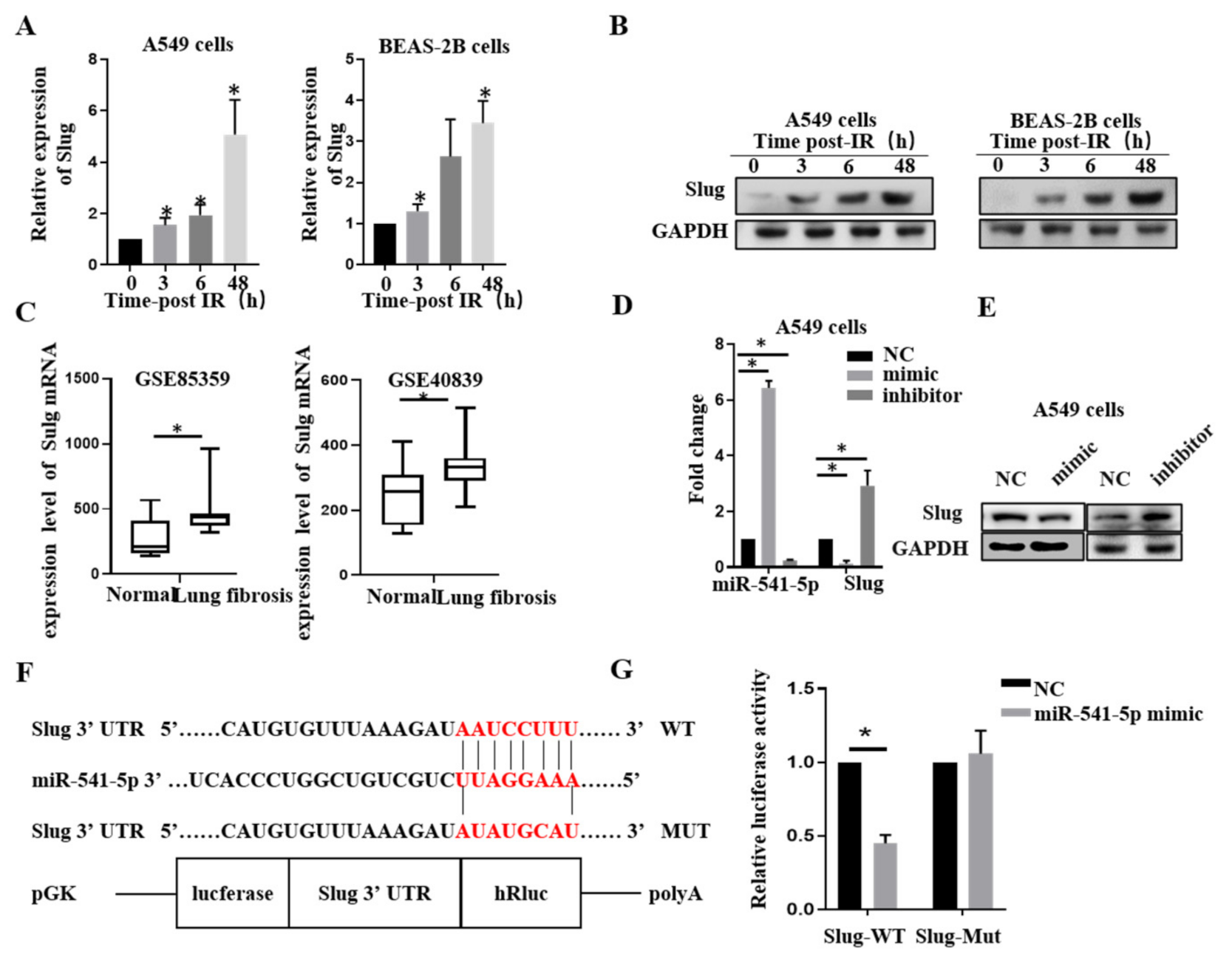
2.3. miR-541-5p Directly Suppressed Slug via Binding to the 3′-UTR Region
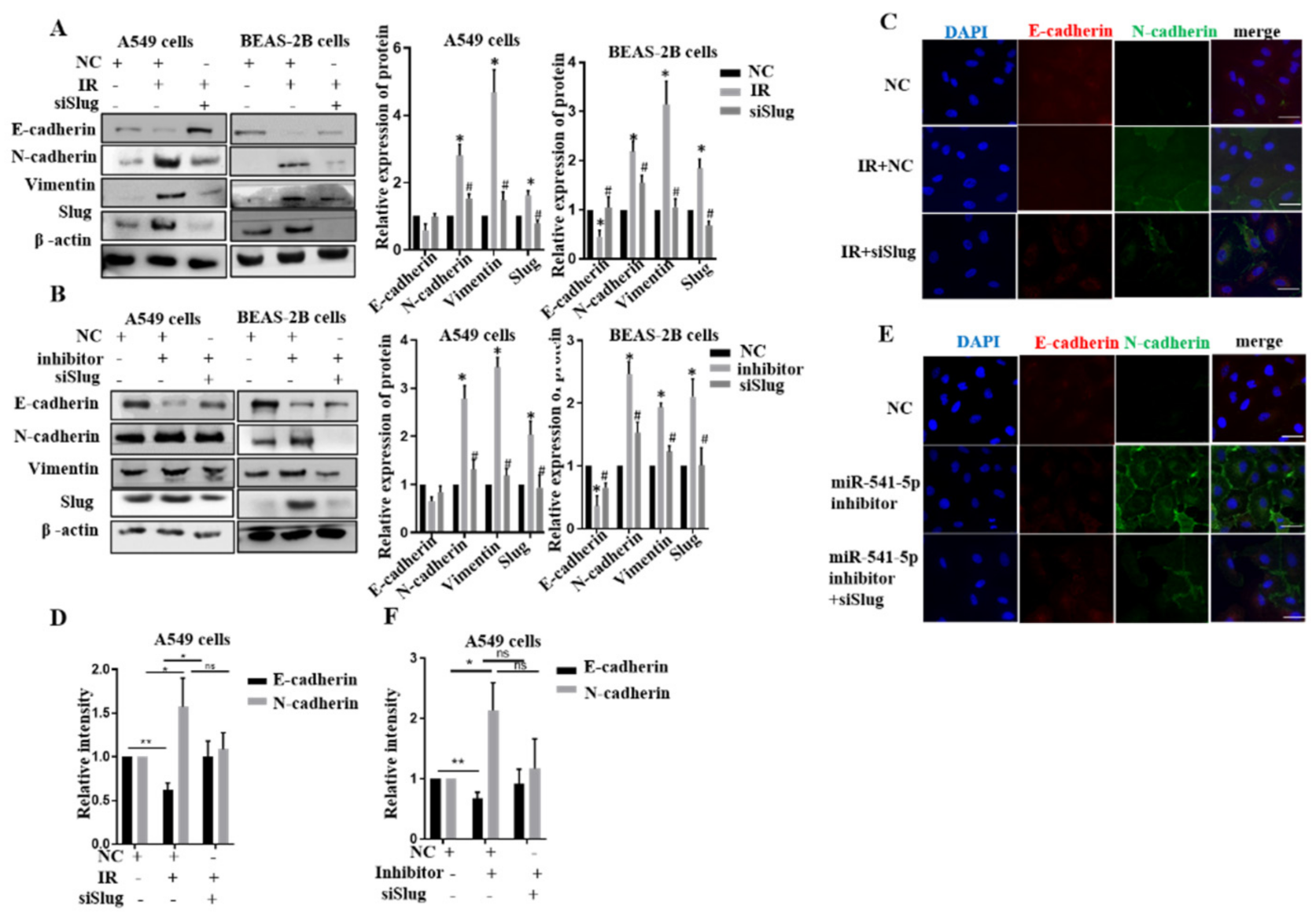
2.4. miR-541-5p Negatively Regulated EMT by Inhibiting Slug
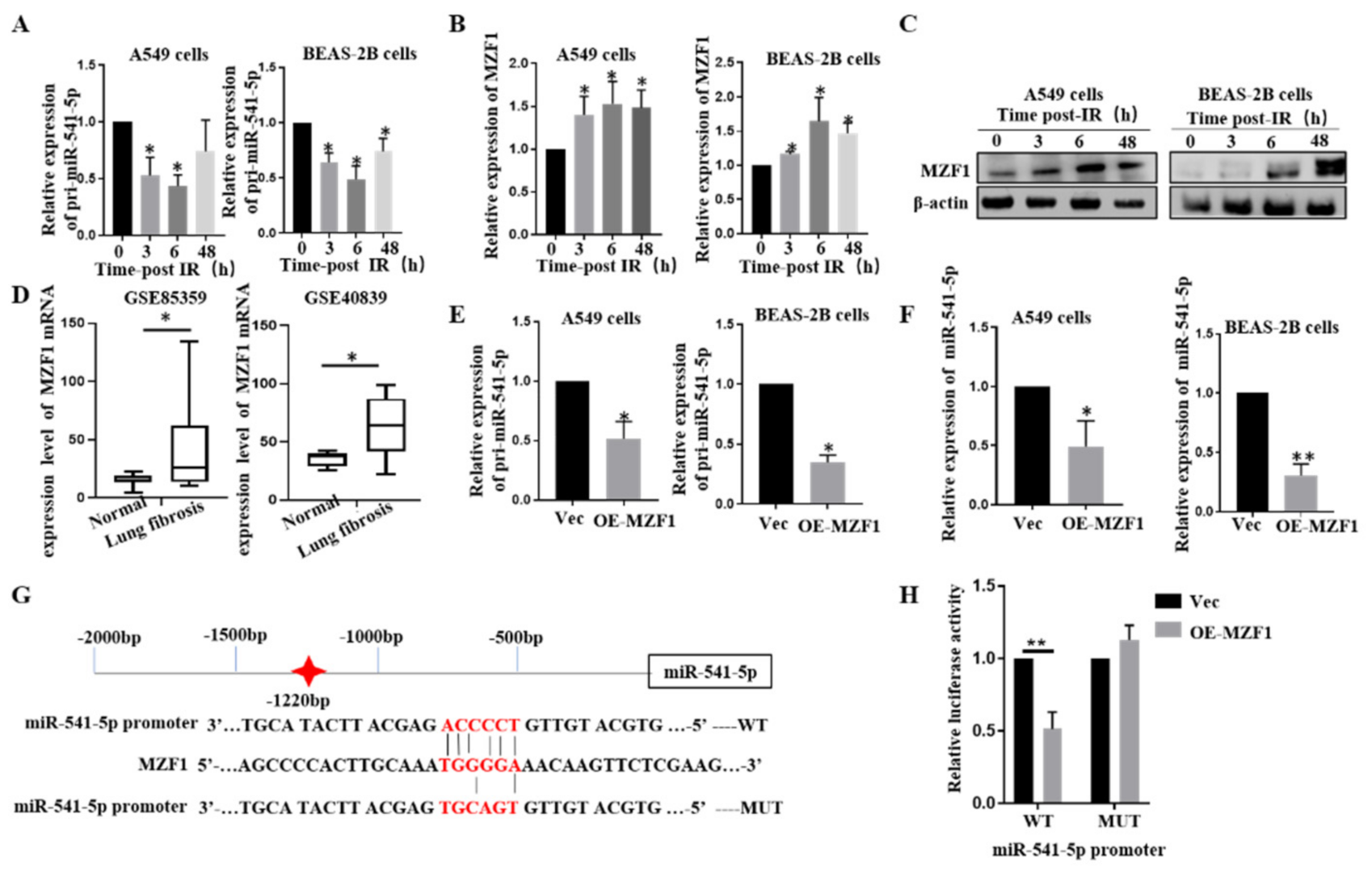
2.5. IR Downgrades miR-541-5p via MZF1
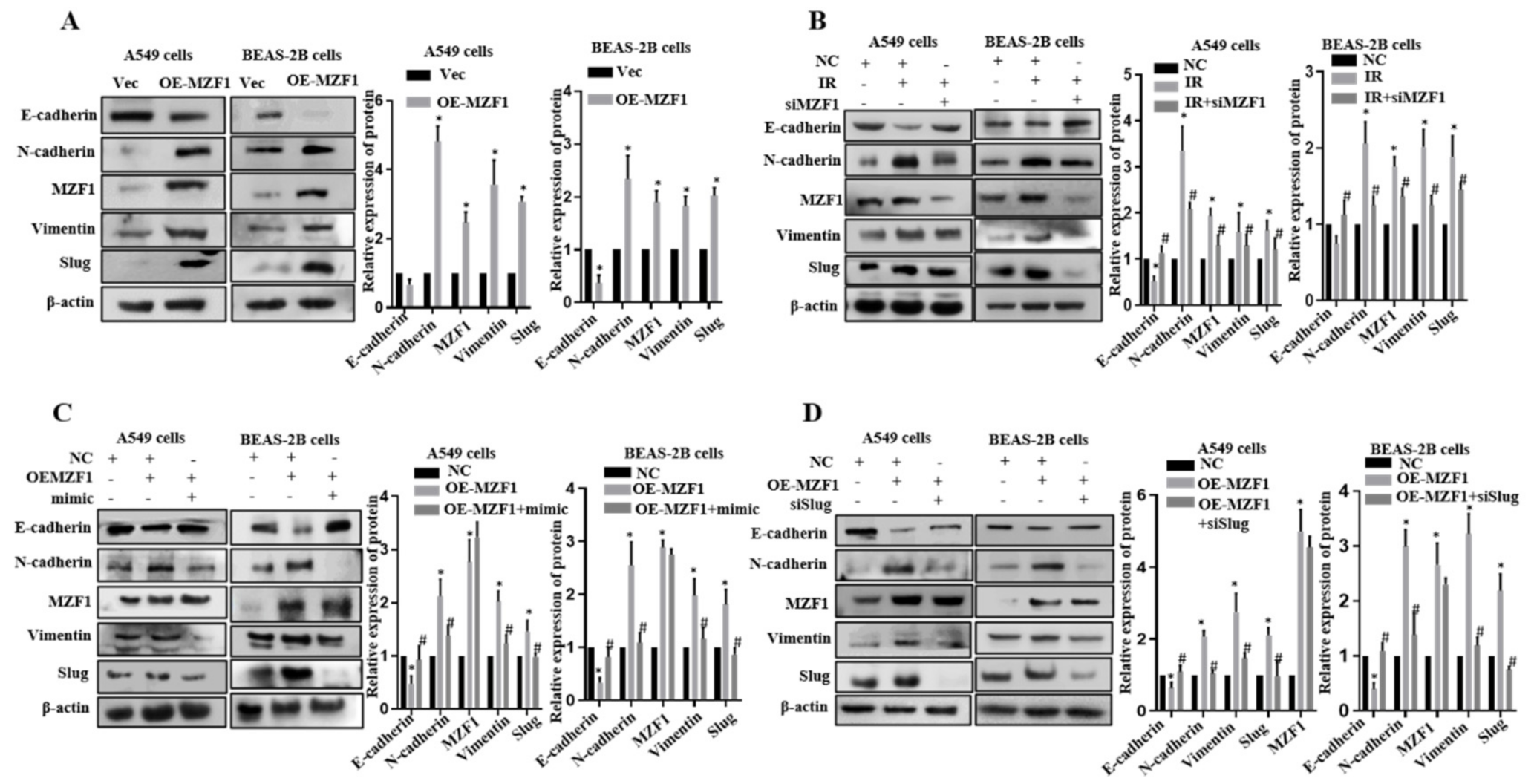
2.6. MZF1 Promotes EMT via Repression of miR-541-5p Following IR
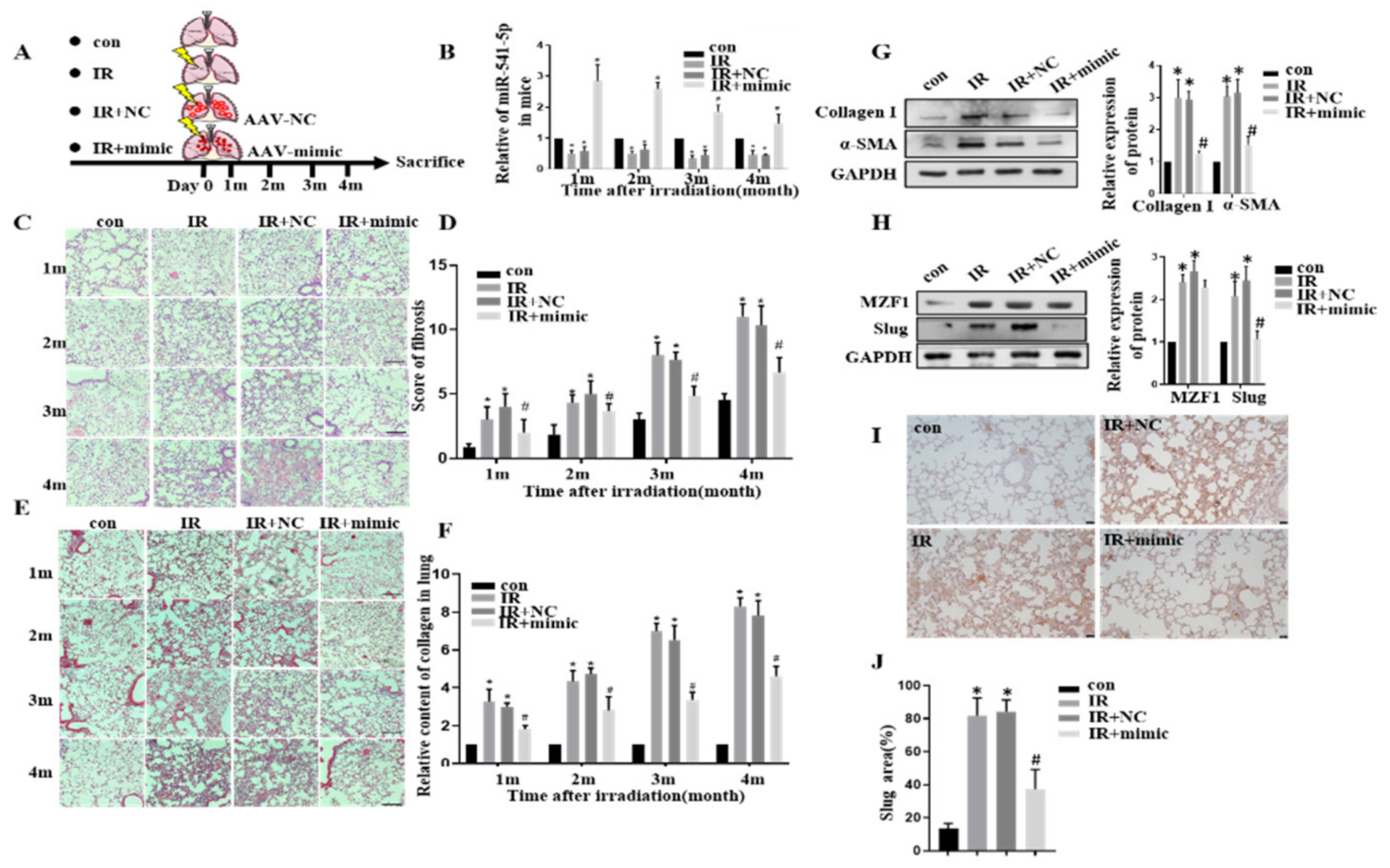
2.7. MZF1 Mediates RIPF via miR-541-5p/Slug Axis
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. RNA Isolation, Reverse Transcription, and qRT-PCR
4.3. Irradiation and Transfection
4.4. Mice and Mice Treatment
4.5. Western Blot Analysis and Antibodies
4.6. Immunofluorescence Analysis
4.7. Dual-Luciferase Reporter Gene Assay
4.8. Hematoxylin and Eosin (H&E) and Masson’s Triple Stain
4.9. Immunohistochemistry (IHC) Assay
4.10. Statistical Analysis
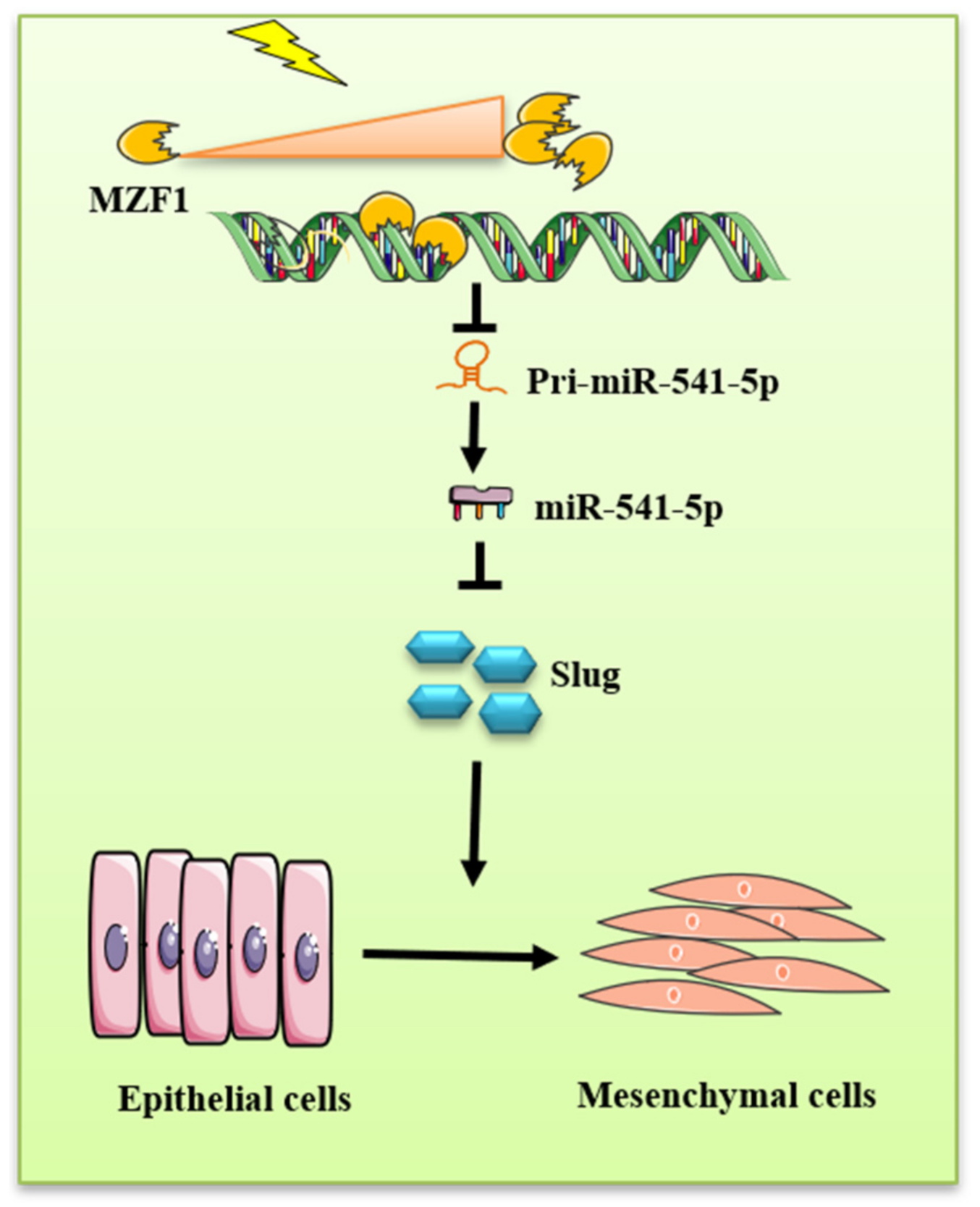
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hanania, A.; Mainwaring, W.; Ghebre, Y.; Hanania, N.; Ludwig, M.J.C. Radiation-Induced Lung Injury: Assessment and Management. Chest 2019, 156, 150–162. [Google Scholar] [CrossRef] [PubMed]
- Delanian, S.; Balla-Mekias, S.; Lefaix, J. Striking regression of chronic radiotherapy damage in a clinical trial of combined pentoxifylline and tocopherol. J. Clin. Oncol. 1999, 17, 3283–3290. [Google Scholar] [CrossRef] [PubMed]
- López-Novoa, J.; Nieto, M. Inflammation and EMT: An alliance towards organ fibrosis and cancer progression. EMBO Mol. Med. 2009, 1, 303–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, S.; Williams, C.; Akahori, Y.; Tanaka, T.; Aikawa, H.; Tong, Y.; Childs-Disney, J.; Disney, M. Small molecule recognition of disease-relevant RNA structures. Chem. Soc. Rev. 2020, 49, 7167–7199. [Google Scholar] [CrossRef]
- Hausser, J.; Zavolan, M. Identification and consequences of miRNA-target interactions–beyond repression of gene expression. Nat. Rev. Genet. 2014, 15, 599–612. [Google Scholar] [CrossRef]
- Rupaimoole, R.; Calin, G.; Lopez-Berestein, G.; Sood, A. miRNA Deregulation in Cancer Cells and the Tumor Microenvironment. Cancer Discov. 2016, 6, 235–246. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Rana, T. Therapeutic targeting of microRNAs: Current status and future challenges. Nat. Rev. Drug Discov. 2014, 13, 622–638. [Google Scholar] [CrossRef]
- Xu, W.; Liu, J.; Feng, J.; Zhu, C.; Yang, Y.; Zhou, W.; Ding, J.; Huang, C.; Cui, Y.; Ding, C.; et al. miR-541 potentiates the response of human hepatocellular carcinoma to sorafenib treatment by inhibiting autophagy. Gut 2020, 69, 1309–1321. [Google Scholar] [CrossRef]
- Liu, C.; Yi, X. miR-541 serves as a prognostic biomarker of osteosarcoma and its regulatory effect on tumor cell proliferation, migration and invasion by targeting TGIF2. Diagn. Pathol. 2020, 15, 96. [Google Scholar] [CrossRef]
- He, Z.; Shen, F.; Qi, P.; Zhai, Z.; Wang, Z. miR-541-3p enhances the radiosensitivity of prostate cancer cells by inhibiting HSP27 expression and downregulating β-catenin. Cell Death Discov. 2021, 7, 18. [Google Scholar] [CrossRef]
- Zhou, W.; Gross, K.; Kuperwasser, C. Molecular regulation of Snai2 in development and disease. J. Cell Sci. 2019, 132, jcs235127. [Google Scholar] [CrossRef]
- Daniel, Y.; Lelou, E.; Aninat, C.; Corlu, A.; Cabillic, F. Interplay between Metabolism Reprogramming and Epithelial-to-Mesenchymal Transition in Cancer Stem Cells. Cancers 2021, 13, 1973. [Google Scholar] [CrossRef]
- Bucay, N.; Bhagirath, D.; Sekhon, K.; Yang, T.; Fukuhara, S.; Majid, S.; Shahryari, V.; Tabatabai, Z.; Greene, K.; Hashimoto, Y.; et al. A novel microRNA regulator of prostate cancer epithelial-mesenchymal transition. Cell Death Differ. 2017, 24, 1263–1274. [Google Scholar] [CrossRef]
- Zirkel, A.; Lederer, M.; Stöhr, N.; Pazaitis, N.; Hüttelmaier, S. IGF2BP1 promotes mesenchymal cell properties and migration of tumor-derived cells by enhancing the expression of LEF1 and SNAI2 (SLUG). Nucleic Acids Res. 2013, 41, 6618–6636. [Google Scholar] [CrossRef] [Green Version]
- Morris, J.; Rauscher, F.; Davis, B.; Klemsz, M.; Xu, D.; Tenen, D.; Hromas, R. The myeloid zinc finger gene, MZF-1, regulates the CD34 promoter in vitro. Blood 1995, 86, 3640–3647. [Google Scholar]
- Vishwamitra, D.; Curry, C.; Alkan, S.; Song, Y.; Gallick, G.; Kaseb, A.; Shi, P.; Amin, H. The transcription factors Ik-1 and MZF1 downregulate IGF-IR expression in NPM-ALK+ T-cell lymphoma. Mol. Cancer 2015, 14, 53. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Liu, Z.; Yan, Z.; Liang, X.; Liu, X.; Liu, Y.; Wang, P.; Bai, C.; Gu, Y.; Zhou, P. MiRNA-155-5p inhibits epithelium-to-mesenchymal transition (EMT) by targeting GSK-3β during radiation-induced pulmonary fibrosis. Arch. Biochem. Biophys. 2021, 697, 108699. [Google Scholar] [CrossRef]
- Liu, Z.; Liang, X.; Li, X.; Liu, X.; Zhu, M.; Gu, Y.; Zhou, P. MiRNA-21 functions in ionizing radiation-induced epithelium-to-mesenchymal transition (EMT) by downregulating PTEN. Toxicol. Res. 2019, 8, 328–340. [Google Scholar] [CrossRef]
- Lei, X.; He, N.; Zhu, L.; Zhou, M.; Zhang, K.; Wang, C.; Huang, H.; Chen, S.; Li, Y.; Liu, Q.; et al. Mesenchymal Stem Cell-Derived Extracellular Vesicles Attenuate Radiation-Induced Lung Injury miRNA-214-3p. Antioxid. Redox Signal. 2021, 35, 849–862. [Google Scholar] [CrossRef]
- Xie, L.; Zhou, J.; Zhang, S.; Chen, Q.; Lai, R.; Ding, W.; Song, C.; Meng, X.; Wu, J. Integrating microRNA and mRNA expression profiles in response to radiation-induced injury in rat lung. Radiat. Oncol. 2014, 9, 111. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z.; Tie, Y.; Lv, G.; Zhu, J.; Fu, H.; Zheng, X. Transcriptional activation of miR-320a by ATF2, ELK1 and YY1 induces cancer cell apoptosis under ionizing radiation conditions. Int. J. Oncol. 2018, 53, 1691–1702. [Google Scholar] [CrossRef] [Green Version]
- Ren, L.; Yang, C.; Dou, Y.; Zhan, R.; Sun, Y.; Yu, Y. MiR-541-5p regulates lung fibrosis by targeting cyclic nucleotide phosphodiesterase 1A. Exp. Lung Res. 2017, 43, 249–258. [Google Scholar] [CrossRef]
- Kroboth, S.; Layton, K.; Jia, F.; Littin, S.; Yu, H.; Hennig, J.; Zaitsev, M. Switching Circuit Optimization for Matrix Gradient Coils. Tomography 2019, 5, 248–259. [Google Scholar] [CrossRef]
- Kota, J.; Chivukula, R.; O’Donnell, K.; Wentzel, E.; Montgomery, C.; Hwang, H.; Chang, T.; Vivekanandan, P.; Torbenson, M.; Clark, K.; et al. Therapeutic microRNA delivery suppresses tumorigenesis in a murine liver cancer model. Cell 2009, 137, 1005–1017. [Google Scholar] [CrossRef] [Green Version]
- Mori, M.; Ludwig, R.; Garcia-Martin, R.; Brandão, B.; Kahn, C. Extracellular miRNAs: From Biomarkers to Mediators of Physiology and Disease. Cell Metab. 2019, 30, 656–673. [Google Scholar] [CrossRef]
- Shirley, S.; Hudson, L.; He, J.; Kusewitt, D. The skinny on Slug. Mol. Carcinog. 2010, 49, 851–861. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.; Kuperwasser, C. SLUG: Critical regulator of epithelial cell identity in breast development and cancer. Cell Adh. Migr. 2014, 8, 578–587. [Google Scholar] [CrossRef] [Green Version]
- Shih, J.; Yang, P. The EMT regulator slug and lung carcinogenesis. Carcinogenesis 2011, 32, 1299–1304. [Google Scholar] [CrossRef] [Green Version]
- Alves, C.; Carneiro, F.; Hoefler, H.; Becker, K. Role of the epithelial-mesenchymal transition regulator Slug in primary human cancers. Front. Biosci. 2009, 14, 3035–3050. [Google Scholar] [CrossRef] [Green Version]
- Recouvreux, M.; Moldenhauer, M.; Galenkamp, K.; Jung, M.; James, B.; Zhang, Y.; Lowy, A.; Bagchi, A.; Commisso, C. Glutamine depletion regulates Slug to promote EMT and metastasis in pancreatic cancer. J Exp Med. 2020, 217, e20200388. [Google Scholar] [CrossRef]
- Krstic, M.; Kolendowski, B.; Cecchini, M.; Postenka, C.; Hassan, H.; Andrews, J.; MacMillan, C.; Williams, K.; Leong, H.; Brackstone, M.; et al. TBX3 promotes progression of pre-invasive breast cancer cells by inducing EMT and directly upregulating SLUG. J. Pathol. 2019, 248, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Leong, K.; Niessen, K.; Kulic, I.; Raouf, A.; Eaves, C.; Pollet, I.; Karsan, A. Jagged1-mediated Notch activation induces epithelial-to-mesenchymal transition through Slug-induced repression of E-cadherin. J. Exp. Med. 2007, 204, 2935–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchikado, Y.; Natsugoe, S.; Okumura, H.; Setoyama, T.; Matsumoto, M.; Ishigami, S.; Aikou, T. Slug Expression in the E-cadherin preserved tumors is related to prognosis in patients with esophageal squamous cell carcinoma. Clin. Cancer Res. 2005, 11, 1174–1180. [Google Scholar] [PubMed]
- Dai, X.; Xin, Y.; Xu, W.; Tian, X.; Wei, X.; Zhang, H. CBP-mediated Slug acetylation stabilizes Slug and promotes EMT and migration of breast cancer cells. Sci. China Life Sci. 2021, 64, 563–574. [Google Scholar] [CrossRef]
- Peng, Y.; Li, Y.; Li, Y.; Wu, A.; Fan, L.; Huang, W.; Fu, C.; Deng, Z.; Wang, K.; Zhang, Y.; et al. HOXC10 promotes tumour metastasis by regulating the EMT-related gene Slug in ovarian cancer. Aging 2020, 12, 19375–19398. [Google Scholar] [CrossRef]
- Zabaleta, N.; Dai, W.; Bhatt, U.; Chichester, J.; Estelien, R.; Sanmiguel, J.; Michalson, K.; Diop, C.; Maciorowski, D.; Qi, W.; et al. Immunogenicity of an AAV-based, room-temperature stable, single dose COVID-19 vaccine in mice and non-human primates. bioRxiv 2021. [Google Scholar] [CrossRef]
- Pan, J.; Huang, G.; Yin, Z.; Cai, X.; Gong, E.; Li, Y.; Xu, C.; Ye, Z.; Cao, Z. Circular RNA FLNA acts as a sponge of miR-486-3p in promoting lung cancer progression via regulating XRCC1 and CYP1A1. Cancer Gene Ther. 2021, 8, 1–21. [Google Scholar] [CrossRef]
- Liu, H.; Han, D.; Hsu, C.; Wang, J. Circulating MicroRNA-486 and MicroRNA-146a serve as potential biomarkers of sarcopenia in the older adults. BMC Geriatr. 2021, 21, 86. [Google Scholar] [CrossRef]
- Beermann, J.; Piccoli, M.; Viereck, J.; Thum, T. Non-coding RNAs in Development and Disease: Background, Mechanisms, and Therapeutic Approaches. Physiol. Rev. 2016, 96, 1297–1325. [Google Scholar] [CrossRef] [Green Version]
- Brix, D.; Bundgaard Clemmensen, K.; Kallunki, T. Zinc Finger Transcription Factor MZF1-A Specific Regulator of Cancer Invasion. Cells 2020, 9, 223. [Google Scholar] [CrossRef] [Green Version]
- Eguchi, T.; Prince, T.; Wegiel, B.; Calderwood, S. Role and Regulation of Myeloid Zinc Finger Protein 1 in Cancer. J. Cell. Biochem. 2015, 116, 2146–2154. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Wang, F.; Xu, F.; Fang, K.; Fang, Z.; Shuai, X.; Cai, K.; Chen, J.; Hu, P.; Chen, D.; et al. A reciprocal feedback of Myc and lncRNA MTSS1-AS contributes to extracellular acidity-promoted metastasis of pancreatic cancer. Theranostics 2020, 10, 10120–10140. [Google Scholar] [CrossRef]
- Zhang, S.; Shi, W.; Ramsay, E.; Bliskovsky, V.; Eiden, A.; Connors, D.; Steinsaltz, M.; DuBois, W.; Mock, B. The transcription factor MZF1 differentially regulates murine promoter variants linked to tumor susceptibility. J. Biol. Chem. 2019, 294, 16756–16764. [Google Scholar] [CrossRef]
- Lin, S.; Wang, X.; Pan, Y.; Tian, R.; Lin, B.; Jiang, G.; Chen, K.; He, Y.; Zhang, L.; Zhai, W.; et al. Transcription Factor Myeloid Zinc-Finger 1 Suppresses Human Gastric Carcinogenesis by Interacting with Metallothionein 2A. Clin. Cancer Res. 2019, 25, 1050–1062. [Google Scholar] [CrossRef] [Green Version]
- Yue, C.; Liu, J.; Chi, C.; Hu, C.; Tan, K.; Huang, F.; Pan, Y.; Lin, K.; Lee, C. Myeloid Zinc Finger 1 (MZF1) Maintains the Mesenchymal Phenotype by Down-regulating IGF1R/p38 MAPK/ERα Signaling Pathway in High-level MZF1-expressing TNBC cells. Anticancer Res. 2019, 39, 4149–4164. [Google Scholar] [CrossRef]
- Ko, H.; Kim, S.; Yang, K.; Kim, K. Phosphorylation-dependent stabilization of MZF1 upregulates N-cadherin expression during protein kinase CK2-mediated epithelial-mesenchymal transition. Oncogenesis 2018, 7, 27. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer ID | Sequence (5′-3′) |
|---|---|
| Has-miR-541-5p-F | AGGATTCTGCTGTCGGT |
| Has-miR-541-5p-R | GGTCCAGTTTTTTTTTTTTTTTAGTG |
| Slug-F | GACTGACCCGTCGTGACG |
| Slug-R | GCAGACGACGGGTCAGAT |
| Pri-miR-541-5p-F | ACGGTGCATGTCATCTGTTC |
| Pri-miR-541-5p-R | AAGATGTCACAGACGACTTC |
| MZF1-F | GGGCCTGCAGGTGAAAGAG |
| MZF1-R | GGCAGCTAGAGGCCCAGACT |
| Has-U6-F | ATTGGAACGATACAGAGAAGAAT |
| Has-U6-R | GGAACGCTTCACGAATTTG |
| β-actin-F | GAATCAATGCAAGTTCGGTTCC |
| β-actin-R | TCATCTCCGCTATTAGCTCCG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, X.; Yan, Z.; Wang, P.; Liu, Y.; Ao, X.; Liu, Z.; Wang, D.; Liu, X.; Zhu, M.; Gao, S.; et al. Irradiation Activates MZF1 to Inhibit miR-541-5p Expression and Promote Epithelial-Mesenchymal Transition (EMT) in Radiation-Induced Pulmonary Fibrosis (RIPF) by Upregulating Slug. Int. J. Mol. Sci. 2021, 22, 11309. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111309
Liang X, Yan Z, Wang P, Liu Y, Ao X, Liu Z, Wang D, Liu X, Zhu M, Gao S, et al. Irradiation Activates MZF1 to Inhibit miR-541-5p Expression and Promote Epithelial-Mesenchymal Transition (EMT) in Radiation-Induced Pulmonary Fibrosis (RIPF) by Upregulating Slug. International Journal of Molecular Sciences. 2021; 22(21):11309. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111309
Chicago/Turabian StyleLiang, Xinxin, Ziyan Yan, Ping Wang, Yuhao Liu, Xingkun Ao, Zheng Liu, Duo Wang, Xiaochang Liu, Maoxiang Zhu, Shanshan Gao, and et al. 2021. "Irradiation Activates MZF1 to Inhibit miR-541-5p Expression and Promote Epithelial-Mesenchymal Transition (EMT) in Radiation-Induced Pulmonary Fibrosis (RIPF) by Upregulating Slug" International Journal of Molecular Sciences 22, no. 21: 11309. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111309