
Lower Spermatozoal PIWI-LIKE 1 and 2 Transcript Levels Are Significantly Associated with Higher Fertilization Rates in IVF


Abstract
:1. Introduction
2. Results
2.1. Distribution of PIWI-LIKE Gene Transcripts in the Spermatozoa
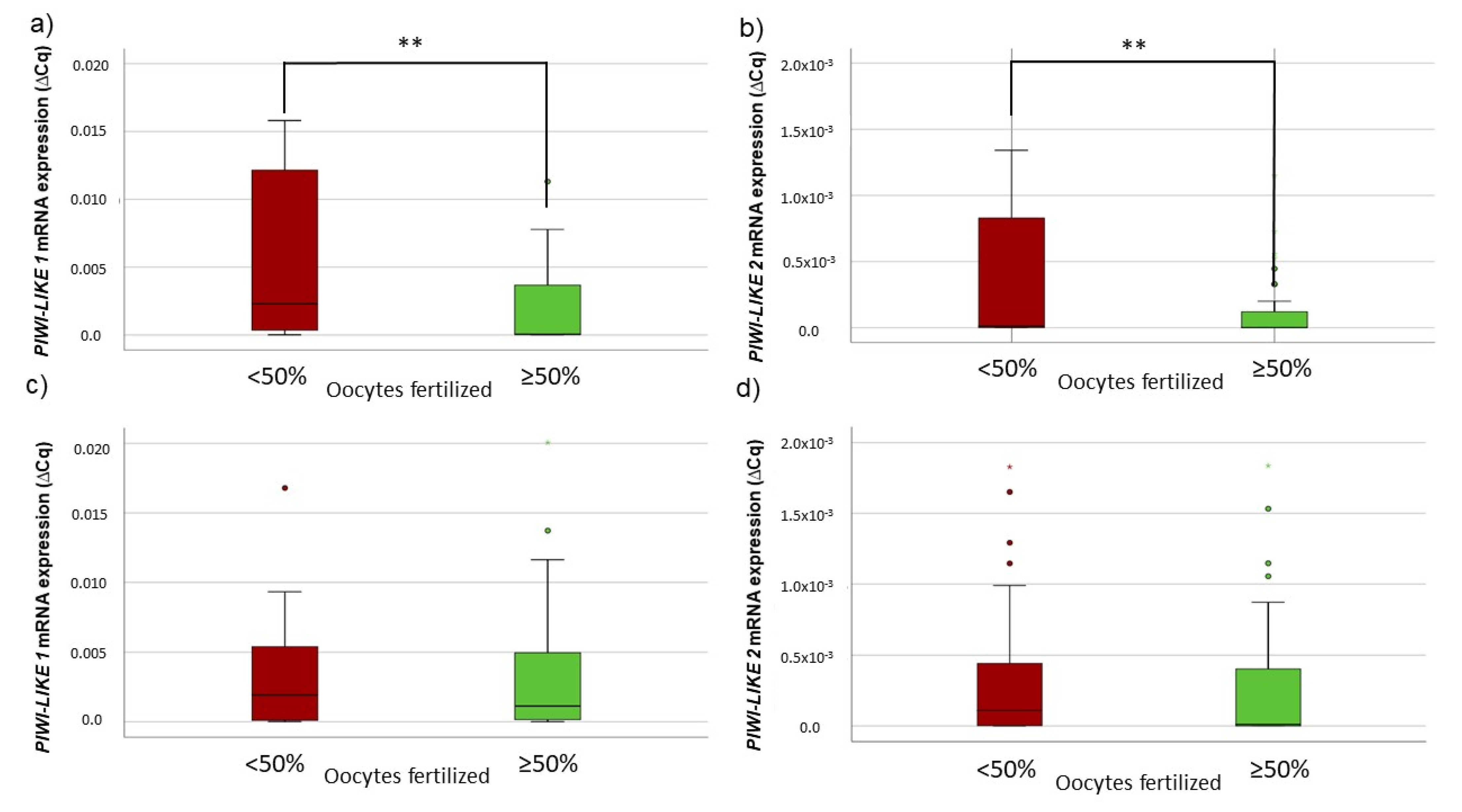
2.2. Lower PIWI-LIKE 1 and 2 mRNA Expression Is Associated with Higher Fertilization Rates in ART
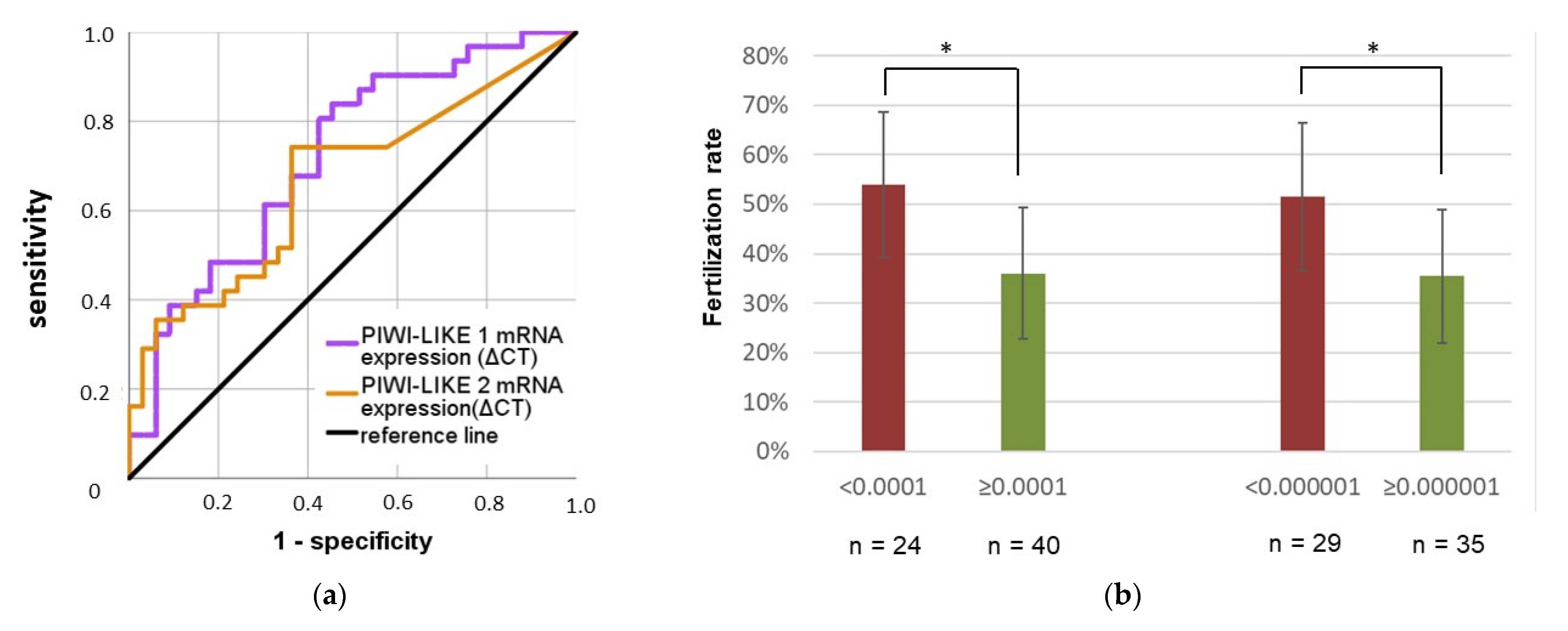
2.3. PIWI-LIKE 1 and 2 mRNA Expression Is a Predictor for Fertilization in IVF, but Not ICSI Cycles
3. Discussion
4. Materials and Methods
4.1. Ethical Approval
4.2. Study Population
4.3. Specimen Preparation and RNA Isolation
4.4. cDNA Synthesis and Quantitative Real-Time PCR
4.5. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Agarwal, A.; Mulgund, A.; Hamada, A.; Chyatte, M.R. A unique view on male infertility around the globe. Reprod. Biol. Endocrinol. 2015, 13, 37. [Google Scholar] [CrossRef] [Green Version]
- Jensen, T.K.; Lindahl-Jacobsen, R.; Christensen, K.; Nielsen, N.C.; Bostofte, E. Good Semen Quality and Life Expectancy: A Cohort Study of 43,277 Men. Am. J. Epidemiol. 2009, 170, 559–565. [Google Scholar] [CrossRef] [Green Version]
- Glazer, C.H.; Eisenberg, M.L.; Tøttenborg, S.S.; Giwercman, A.; Flachs, E.M.; Bräuner, E.; Vassard, D.; Pinborg, A.; Schmidt, L.; Bonde, J.P. Male factor infertility and risk of death: A nationwide record-linkage study. Hum. Reprod. 2019, 34, 2266–2273. [Google Scholar] [CrossRef]
- Thilagavathi, J.; Venkatesh, S.; Kumar, R.; Dada, R. Segregation of sperm subpopulations in normozoospermic infertile men. Syst. Biol. Reprod. Med. 2012, 58, 313–318. [Google Scholar] [CrossRef]
- Albani, E.; Castellano, S.; Gurrieri, B.; Arruzzolo, L.; Negri, L.; Borroni, E.M.; Levi-Setti, P.E. Male age: Negative impact on sperm DNA fragmentation. Aging 2019, 11, 2749–2761. [Google Scholar] [CrossRef]
- Poenicke, K.; Grunewald, S.; Glander, H.; Paasch, U.; Kamini, A.R.; Agarwal, A.; Srinivas, M. Sperm Selection in Assisted Reproductive Techniques. In Andrology Laboratory Manual; Jaypee Brothers Pvt Ltd.: New Delhi, India, 2010; pp. 173–187. [Google Scholar] [CrossRef]
- Dutta, S.; Henkel, R.; Agarwal, A. Comparative analysis of tests used to assess sperm chromatin integrity and DNA fragmentation. Andrologia 2020, 53, e13718. [Google Scholar] [CrossRef]
- Evenson, D.P.; Wixon, R. Clinical aspects of sperm DNA fragmentation detection and male infertility. Theriogenology 2006, 65, 979–991. [Google Scholar] [CrossRef]
- Tang, L.; Rao, M.; Yang, W.; Yao, Y.; Luo, Q.; Lu, L.; Wang, L.; Zhao, S. Predictive value of the sperm DNA fragmentation index for low or failed IVF fertilization in men with mild-to-moderate asthenozoospermia. J. Gynecol. Obstet. Hum. Reprod. 2020, 50, 101868. [Google Scholar] [CrossRef]
- Oleszczuk, K.; Giwercman, A.; Bungum, M. Sperm chromatin structure assay in prediction of in vitro fertilization outcome. Andrology 2016, 4, 290–296. [Google Scholar] [CrossRef] [Green Version]
- Lin, M.-H.; Lee, R.K.-K.; Li, S.-H.; Lu, C.-H.; Sun, F.-J.; Hwu, Y.-M. Sperm chromatin structure assay parameters are not related to fertilization rates, embryo quality, and pregnancy rates in in vitro fertilization and intracytoplasmic sperm injection, but might be related to spontaneous abortion rates. Fertil. Steril. 2008, 90, 352–359. [Google Scholar] [CrossRef]
- Green, K.A.; Patounakis, G.; Dougherty, M.P.; Werner, M.D.; Scott, R.T.; Franasiak, J.M. Sperm DNA fragmentation on the day of fertilization is not associated with embryologic or clinical outcomes after IVF/ICSI. J. Assist. Reprod. Genet. 2019, 37, 71–76. [Google Scholar] [CrossRef]
- Speyer, B.E.; Pizzey, A.R.; Ranieri, M.; Joshi, R.; Delhanty, J.D.A.; Serhal, P. Fall in implantation rates following ICSI with sperm with high DNA fragmentation. Hum. Reprod. 2010, 25, 1609–1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, C.; Li, T.; Xie, Y.; Guo, Y.; Yang, Q.; Liang, X.; Deng, C.; Liu, G. Sperm DNA fragmentation index influences assisted reproductive technology outcome: A systematic review and meta-analysis combined with a retrospective cohort study. Andrologia 2019, 51, e13263. [Google Scholar] [CrossRef] [PubMed]
- Rex, A.S.; Wu, C.; Aagaard, J.; Fedder, J. Implementation of an in-house flow cytometric analysis of DNA fragmentation in spermatozoa. Asian J. Androl. 2020, 22, 246. [Google Scholar] [CrossRef] [PubMed]
- Hamada, A.J.; Esteves, S.C.; Agarwal, A. A comprehensive review of genetics and genetic testing in azoospermia. Clinics 2013, 68, 39–60. [Google Scholar] [CrossRef]
- Sasaki, T.; Shiohama, A.; Minoshima, S.; Shimizu, N. Identification of eight members of the Argonaute family in the human genome☆. Genomics 2003, 82, 323–330. [Google Scholar] [CrossRef]
- Kuramochi-Miyagawa, S.; Kimura, T.; Yomogida, K.; Kuroiwa, A.; Tadokoro, Y.; Fujita, Y.; Sato, M.; Matsuda, Y.; Nakano, T. Two mouse piwi-related genes: Miwi and mili. Mech. Dev. 2001, 108, 121–133. [Google Scholar] [CrossRef]
- Deng, W.; Lin, H. miwi, a Murine Homolog of piwi, Encodes a Cytoplasmic Protein Essential for Spermatogenesis. Dev. Cell 2002, 2, 819–830. [Google Scholar] [CrossRef] [Green Version]
- Kuramochi-Miyagawa, S.; Kimura, T.; Ijiri, T.W.; Isobe, T.; Asada, N.; Fujita, Y.; Ikawa, M.; Iwai, N.; Okabe, M.; Deng, W.; et al. Mili, a mammalian member ofpiwifamily gene, is essential for spermatogenesis. Development 2004, 131, 839–849. [Google Scholar] [CrossRef] [Green Version]
- Carmell, M.A.; Girard, A.; van de Kant, H.J.; Bourc’His, D.; Bestor, T.H.; de Rooij, D.; Hannon, G.J. MIWI2 Is Essential for Spermatogenesis and Repression of Transposons in the Mouse Male Germline. Dev. Cell 2007, 12, 503–514. [Google Scholar] [CrossRef] [Green Version]
- Behre, H.; Giebler, M.; Greither, T.; Müller, L.; Mösinger, C. Altered PIWI-LIKE 1 and PIWI-LIKE 2 mRNA expression in ejaculated spermatozoa of men with impaired sperm characteristics. Asian J. Androl. 2018, 20, 260–264. [Google Scholar] [CrossRef] [PubMed]
- Bak, C.W.; Yoon, T.-K.; Choi, Y. Functions of PIWI proteins in spermatogenesis. Clin. Exp. Reprod. Med. 2011, 38, 61–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hempfling, A.L.; Lim, S.L.; Adelson, D.; Evans, J.; O’Connor, A.E.; Qu, Z.P.; Kliesch, S.; Weidner, W.; O’Bryan, M.K.; Bergmann, M. Expression patterns of HENMT1 and PIWIL1 in human testis: Implications for transposon expression. Reproduction 2017, 154, 363–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, L.; Fang, L.; Shi, B.; Qiu, S.; Ye, Y. Spermatozoa Expression of piR-31704, piR-39888, and piR-40349 and Their Correlation to Sperm Concentration and Fertilization Rate after ICSI. Reprod. Sci. 2017, 25, 733–739. [Google Scholar] [CrossRef] [PubMed]
- Denomme, M.M.; McCallie, B.R.; Parks, J.C.; Schoolcraft, W.B.; Katz-Jaffe, M.G. Alterations in the sperm histone-retained epigenome are associated with unexplained male factor infertility and poor blastocyst development in donor oocyte IVF cycles. Hum. Reprod. 2017, 32, 2443–2455. [Google Scholar] [CrossRef]
- Gou, L.-T.; Kang, J.-Y.; Dai, P.; Wang, X.; Li, F.; Zhao, S.; Zhang, M.; Hua, M.-M.; Lu, Y.; Zhu, Y.; et al. Ubiquitination-Deficient Mutations in Human Piwi Cause Male Infertility by Impairing Histone-to-Protamine Exchange during Spermiogenesis. Cell 2017, 169, 1090–1104.e13. [Google Scholar] [CrossRef] [Green Version]
- Oud, M.; Volozonoka, L.; Friedrich, C.; Kliesch, S.; Nagirnaja, L.; Gilissen, C.; O’Bryan, M.; McLachlan, R.; Aston, K.; Tüttelmann, F.; et al. Lack of evidence for a role of PIWIL1 variants in human male infertility. Cell 2021, 184, 1941–1942. [Google Scholar] [CrossRef]
- Ostermeier, G.C.; Goodrich, R.J.; Moldenhauer, J.S.; Diamond, M.P.; Krawetz, S.A. A suite of novel human spermatozoal RNAs. J. Androl. 2005, 26, 70–74. [Google Scholar]
- Jodar, M.; Selvaraju, S.; Sendler, E.; Diamond, M.; Krawetz, S.A.; Reproductive Medicine Network. The presence, role and clinical use of spermatozoal RNAs. Hum. Reprod. Update 2013, 19, 604–624. [Google Scholar] [CrossRef]
- Lim, A.K.; Lorthongpanich, C.; Chew, T.G.; Tan, C.W.G.; Shue, Y.T.; Balu, S.; Gounko, N.; Kuramochi-Miyagawa, S.; Matzuk, M.M.; Chuma, S.; et al. The nuage mediates retrotransposon silencing in mouse primordial ovarian follicles. Development 2013, 140, 3819–3825. [Google Scholar] [CrossRef] [Green Version]
- Kohlrausch, F.B.; Berteli, T.S.; Wang, F.; Navarro, P.A.; Keefe, D.L. Control of LINE-1 Expression Maintains Genome Integrity in Germline and Early Embryo Development. Reprod. Sci. 2021, 1–13. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Median Expression (ΔCq) | p (MWU) | n | ||
|---|---|---|---|---|
| PIWI-LIKE 1 | swim-up fraction | 0.00113 | <0.001 | 160 |
| cell pellet | 0.00399 | |||
| PIWI-LIKE 2 | swim-up fraction | 0.00001 | n.s. | 160 |
| cell pellet | 0.000012 | |||
| PIWI-LIKE 3 | swim-up fraction | 0.00 | <0.001 | 160 |
| cell pellet | 0.00 | |||
| PIWI-LIKE 4 | swim-up fraction | 0.00 | n.s. | 160 |
| cell pellet | 0.00 |
| Parameter | Male Partner | Female Partner |
|---|---|---|
| age (y) | 37 (25–52) | 34 (26–41) |
| weight (kg) | 88.4 (58–150) | 67.1 (43–123) |
| BMI (kg/m2) | 26.9 (18.3–41.6) | 24.0 (16.5–40.2) |
| days of abstinence | 4 (2–21) | |
| semen volume (mL) | 2.8 (0.1–10.4) | |
| sperm count (total sperm/ejaculate) | 70.1 (0.1–717.6) | |
| progressive motility (%) | 50 % (0–90%) | |
| % morphologically normal spermatozoa | 8% (0–24%) |
| Total (n = 160) | IVF (n = 64) | ICSI (n = 96) | |
|---|---|---|---|
| Cycles with male indication for ART | 26 | 0 | 26 |
| Cycles with female indication for ART | 61 | 56 | 5 |
| Cycles with both partner indication for ART | 73 | 8 | 65 |
| Oocytes retrieved (n) | 9 (1–36) | 10 (1–25) | 8 (1–36) |
| Inseminated (n) | 8 (1–25) | 10 (1–25) | 6 (1–19) |
| Fertilized (n) | 4 (0–14) | 4 (0–14) | 3 (0–10) |
| Fertilization rate (%) | 50 (0–100) | 50 (0–100) | 58 (0–100) |
| Transferred embryos (n) | 2 (0–3) | 2 (0–3) | 2 (0–3) |
| Clinical pregnancy rate (n) | 26.3 | 26.6 | 26.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giebler, M.; Greither, T.; Handke, D.; Seliger, G.; Behre, H.M. Lower Spermatozoal PIWI-LIKE 1 and 2 Transcript Levels Are Significantly Associated with Higher Fertilization Rates in IVF. Int. J. Mol. Sci. 2021, 22, 11320. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111320
Giebler M, Greither T, Handke D, Seliger G, Behre HM. Lower Spermatozoal PIWI-LIKE 1 and 2 Transcript Levels Are Significantly Associated with Higher Fertilization Rates in IVF. International Journal of Molecular Sciences. 2021; 22(21):11320. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111320
Chicago/Turabian StyleGiebler, Maria, Thomas Greither, Diana Handke, Gregor Seliger, and Hermann M. Behre. 2021. "Lower Spermatozoal PIWI-LIKE 1 and 2 Transcript Levels Are Significantly Associated with Higher Fertilization Rates in IVF" International Journal of Molecular Sciences 22, no. 21: 11320. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111320