
Systematic Analysis of FASTK Gene Family Alterations in Cancer

 and
and
Abstract
:1. Introduction
2. Results
2.1. Genetic Alterations of FASTK Family Genes across Human Cancers
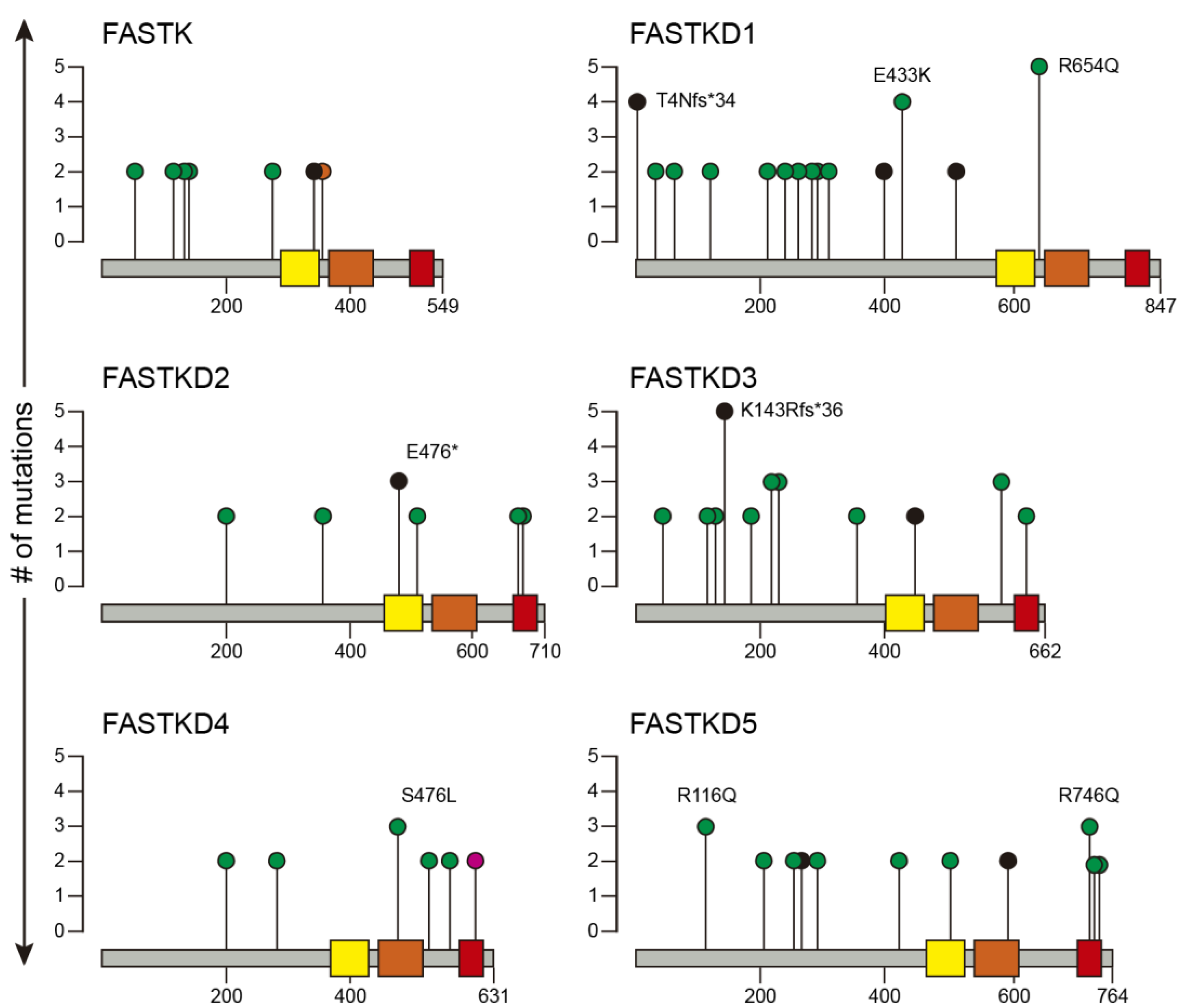
2.2. Recurrent Mutations in FASTK Genes
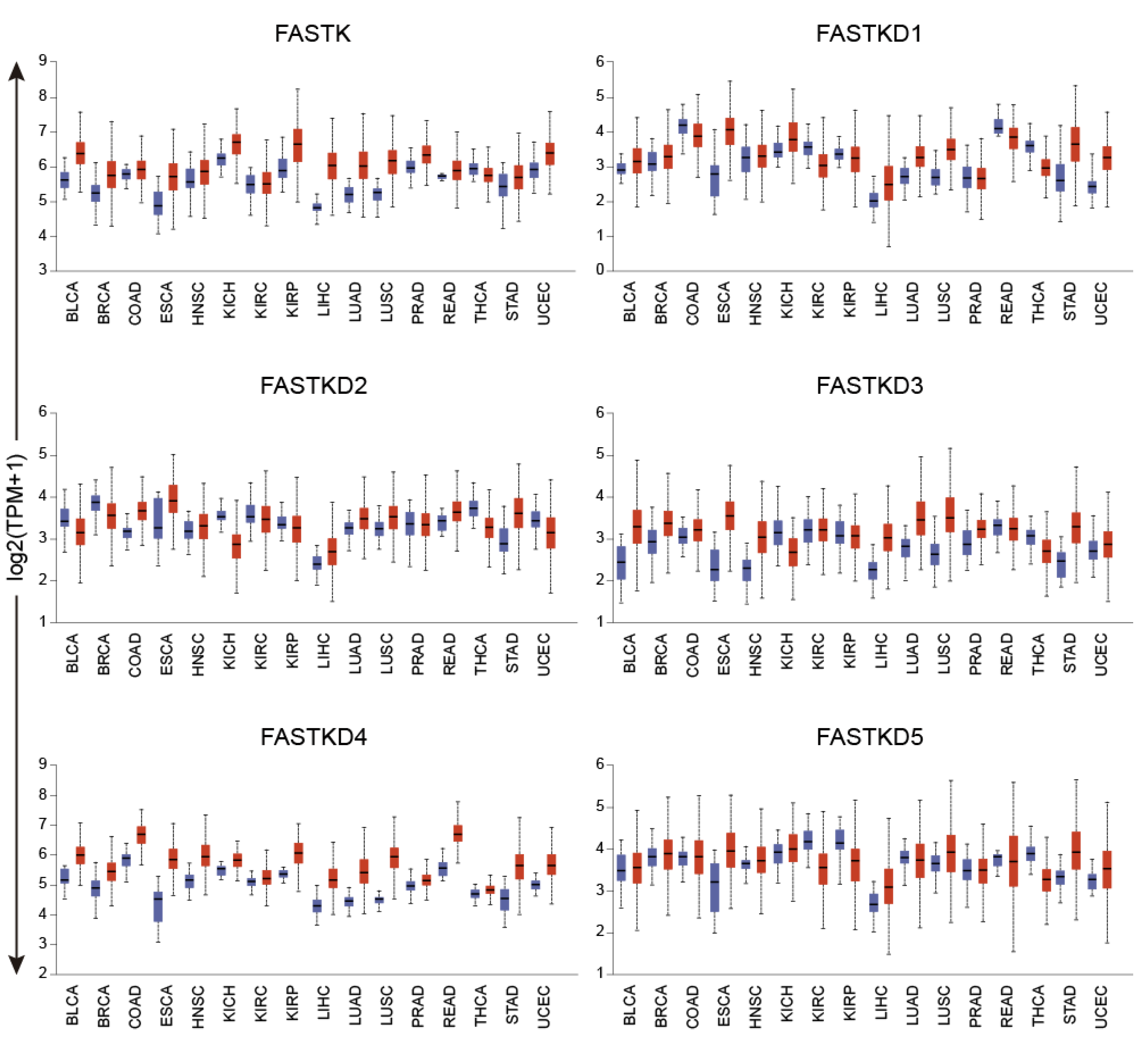
2.3. Expression Profile of FASTK Genes across Human Cancers
2.4. Protein Interaction Network of FASTK Members: Pathways and Molecular Functions Enrichment Analysis
3. Discussion
4. Materials and Methods
4.1. TCGA Data Source Selection and Processing for Genetic Alterations Analysis
4.2. TCGA Data Source Selection and Processing for Expression Analysis
4.3. Construction of the Protein-Protein Interaction Network of FASTK Proteins
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Simarro, M.; Gimenez-Cassina, A.; Kedersha, N.; Lazaro, J.B.; Adelmant, G.O.; Marto, J.A.; Rhee, K.; Tisdale, S.; Danial, N.; Benarafa, C.; et al. Fast kinase domain-containing protein 3 is a mitochondrial protein essential for cellular respiration. Biochem. Biophys. Res. Commun. 2010, 401, 440–446. [Google Scholar] [CrossRef] [Green Version]
- Lee, I.; Hong, W. RAP—A putative RNA-binding domain. Trends Biochem. Sci. 2004, 29, 567–570. [Google Scholar] [CrossRef]
- Jourdain, A.A.; Popow, J.; de la Fuente, M.A.; Martinou, J.-C.; Anderson, P.; Simarro, M. The FASTK family of proteins: Emerging regulators of mitochondrial RNA biology. Nucleic Acids Res. 2017, 45, 10941–10947. [Google Scholar] [CrossRef] [Green Version]
- Ghezzi, D.; Saada, A.; D’Adamo, P.; Fernandez-Vizarra, E.; Gasparini, P.; Tiranti, V.; Elpeleg, O.; Zeviani, M. FASTKD2 Nonsense Mutation in an Infantile Mitochondrial Encephalomyopathy Associated with Cytochrome C Oxidase Deficiency. Am. J. Hum. Genet. 2008, 83, 415–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, D.H.; Choi, Y.C.; Nam, D.E.; Choi, S.S.; Kim, J.W.; Choi, B.O.; Chung, K.W. Identification of FASTKD2 compound heterozygous mutations as the underlying cause of autosomal recessive MELAS-like syndrome. Mitochondrion 2017, 35, 54–58. [Google Scholar] [CrossRef]
- Simarro, M.; Chen, M.; De la Fuente, M.A.; Eiros, J.M.; Orduña, A.; Anderson, P. Deletion of FAST (Fas-activated serine/threonine phosphoprotein) ameliorates immune complex arthritis in mice. Mod. Rheumatol. 2016, 26, 630–632. [Google Scholar] [CrossRef]
- Simarro, M.; Giannattasio, G.; De la Fuente, M.A.; Benarafa, C.; Subramanian, K.K.; Ishizawar, R.; Balestrieri, B.; Andersson, E.M.; Luo, H.R.; Orduña, A.; et al. Fas-Activated Serine/Threonine Phosphoprotein Promotes Immune-Mediated Pulmonary Inflammation. J. Immunol. 2010, 184, 5325–5332. [Google Scholar] [CrossRef] [PubMed]
- García Del Río, A.; Delmiro, A.; Martín, M.A.; Cantalapiedra, R.; Carretero, R.; Durántez, C.; Menegotto, F.; Morán, M.; Serrano-Lorenzo, P.; De la Fuente, M.A.; et al. The Mitochondrial Isoform of FASTK Modulates Nonopsonic Phagocytosis of Bacteria by Macrophages via Regulation of Respiratory Complex I. J. Immunol. 2018, 201, 2977–2985. [Google Scholar] [CrossRef] [PubMed]
- Bauer, A.; Kleeff, J.; Bier, M.; Wirtz, M.; Kayed, H.; Esposito, I.; Korc, M.; Hafner, M.; Hoheisel, J.D.; Friess, H. Identification of malignancy factors by analyzing cystic tumors of the pancreas. Pancreatology 2009, 9, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Van Doorn, R.; Van Kester, M.S.; Dijkman, R.; Vermeer, M.H.; Mulder, A.A.; Szuhai, K.; Knijnenburg, J.; Boer, J.M.; Willemze, R.; Tensen, C.P. Oncogenomic analysis of mycosis fungoides reveals major differences with Sézary syndrome. Blood 2009, 113, 127–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhi, F.; Chen, X.; Wang, S.; Xia, X.; Shi, Y.; Guan, W.; Shao, N.; Qu, H.; Yang, C.; Zhang, Y.; et al. The use of hsa-miR-21, hsa-miR-181b and hsa-miR-106a as prognostic indicators of astrocytoma. Eur. J. Cancer 2010, 46, 1640–1649. [Google Scholar] [CrossRef]
- Zhi, F.; Zhou, G.; Shao, N.; Xia, X.; Shi, Y.; Wang, Q.; Zhang, Y.; Wang, R.; Xue, L.; Wang, S.; et al. miR-106a-5p Inhibits the Proliferation and Migration of Astrocytoma Cells and Promotes Apoptosis by Targeting FASTK. PLoS ONE 2013, 8, e72390. [Google Scholar] [CrossRef]
- Colas, E.; Perez, C.; Cabrera, S.; Pedrola, N.; Monge, M.; Castellvi, J.; Eyzaguirre, F.; Gregorio, J.; Ruiz, A.; Llaurado, M.; et al. Molecular markers of endometrial carcinoma detected in uterine aspirates. Int. J. Cancer 2011, 129, 2435–2444. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Mi, J.Q.; Debernardi, A.; Vitte, A.L.; Emadali, A.; Meyer, J.A.; Charmpi, K.; Ycart, B.; Callanan, M.B.; Carroll, W.L.; et al. A six gene expression signature defines aggressive subtypes and predicts outcome in childhood and adult acute lymphoblastic leukemia. Oncotarget 2015, 6, 16527–16542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, R.; Zhang, B.; Lu, X.; Jin, X.; Liu, T. FASTKD2 promotes cancer cell progression through upregulating Myc expression in pancreatic ductal adenocarcinoma. J. Cell. Biochem. 2020, 121, 2458–2466. [Google Scholar] [CrossRef]
- Wu, Y.; Liu, Y.; He, A.; Guan, B.; He, S.; Zhang, C.; Kang, Z.; Gong, Y.; Li, X.; Zhou, L. Identification of the Six-RNA-Binding Protein Signature for Prognosis Prediction in Bladder Cancer. Front. Genet. 2020, 11, 992. [Google Scholar] [CrossRef] [PubMed]
- Yeung, K.T.; Das, S.; Zhang, J.; Lomniczi, A.; Ojeda, S.R.; Xu, C.-F.; Neubert, T.A.; Samuels, H.H. A Novel Transcription Complex That Selectively Modulates Apoptosis of Breast Cancer Cells through Regulation of FASTKD2. Mol. Cell. Biol. 2011, 31, 2287–2298. [Google Scholar] [CrossRef] [Green Version]
- Das, S.; Yeung, K.T.; Mahajan, M.A.; Samuels, H.H. Fas Activated Serine-Threonine Kinase Domains 2 (FASTKD2) mediates apoptosis of breast and prostate cancer cells through its novel FAST2 domain. BMC Cancer 2014, 14, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.Z.; Xue, M.Z.; Shen, H.J.; Li, X.G.; Ma, D.; Gong, Y.; Liu, Y.R.; Qiao, F.; Xie, H.Y.; Lian, B.; et al. PHF5A epigenetically inhibits apoptosis to promote breast cancer progression. Cancer Res. 2018, 78, 3190–3206. [Google Scholar] [CrossRef] [Green Version]
- Campbell, P.J.; Getz, G.; Korbel, J.O.; Stuart, J.M.; Jennings, J.L.; Stein, L.D.; Perry, M.D.; Nahal-Bose, H.K.; Ouellette, B.F.F.; Li, C.H.; et al. Pan-cancer analysis of whole genomes. Nature 2020, 578, 82–93. [Google Scholar] [CrossRef] [Green Version]
- Frampton, G.M.; Fichtenholtz, A.; Otto, G.A.; Wang, K.; Downing, S.R.; He, J.; Schnall-Levin, M.; White, J.; Sanford, E.M.; An, P.; et al. Development and validation of a clinical cancer genomic profiling test based on massively parallel DNA sequencing. Nat. Biotechnol. 2013, 31, 1023–1031. [Google Scholar] [CrossRef]
- Ellrott, K.; Bailey, M.H.; Saksena, G.; Covington, K.R.; Kandoth, C.; Stewart, C.; Hess, J.; Ma, S.; Chiotti, K.E.; McLellan, M.D.; et al. Scalable Open Science Approach for Mutation Calling of Tumor Exomes Using Multiple Genomic Pipelines. Cell Syst. 2018, 6, 271–281. [Google Scholar] [CrossRef] [Green Version]
- Tan, H.; Bao, J.; Zhou, X. Genome-wide mutational spectra analysis reveals significant cancer-specific heterogeneity. Sci. Rep. 2015, 5, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Wood, L.D.; Parsons, D.W.; Jones, S.; Lin, J.; Sjöblom, T.; Leary, R.J.; Shen, D.; Boca, S.M.; Barber, T.; Ptak, J.; et al. The genomic landscapes of human breast and colorectal cancers. Science 2007, 318, 1108–1113. [Google Scholar] [CrossRef] [Green Version]
- Grasso, D.; Zampieri, L.X.; Capelôa, T.; Van De Velde, J.A.; Sonveaux, P. Mitochondria in cancer. Cell Stress 2020, 4, 114–146. [Google Scholar] [CrossRef]
- Matsui, A.; Ihara, T.; Suda, H.; Mikami, H.; Semba, K. Gene amplification: Mechanisms and involvement in cancer. Biomol. Concepts 2013, 4, 567–582. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wen, S.; Wu, Y.; Shi, L.; Xu, X.; Shen, B. Efficacy and safety of first-generation epidermal growth factor receptor (EGFR) tyrosine kinase inhibitors (TKIs) combined with chemotherapy or antiangiogenic therapy as first-line treatment in patients with EGFR-mutant non-small cell lung cancer: A systematic review and meta-analysis. Crit. Rev. Oncol. Hematol. 2021, 163, 103393. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Wang, G.; Liu, Y.; Feng, L.; Wang, M.; Liu, J.; Chen, Y.; Ouyang, L. Development of small-molecule tropomyosin receptor kinase (TRK) inhibitors for NTRK fusion cancers. Acta Pharm. Sin. B 2021, 11, 355–372. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.W.; Brognard, J.; Coughlin, C.; You, Z.; Dolled-Filhart, M.; Aslanian, A.; Manning, G.; Abraham, R.T.; Hunter, T. The F Box Protein Fbx6 Regulates Chk1 Stability and Cellular Sensitivity to Replication Stress. Mol. Cell 2009, 35, 442–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.Z.; Wang, Z.Q.; Shan, H.Z.; Zhou, L.; Yang, L.; Lei, H.; Liu, B.; Wu, Y.L. Overexpression of Fbxo6 inactivates spindle checkpoint by interacting with Mad2 and BubR1. Cell Cycle 2018, 17, 2779–2789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio Cancer Genomics Portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, J.; Arman Aksoy, B.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Onur Sumer, S.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative Analysis of Complex Cancer Genomics and Clinical Profiles Using the cBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Jiménez, F.; Muiños, F.; Sentís, I.; Deu-Pons, J.; Reyes-Salazar, I.; Arnedo-Pac, C.; Mularoni, L.; Pich, O.; Bonet, J.; Kranas, H.; et al. A compendium of mutational cancer driver genes. Nat. Rev. Cancer 2020, 20, 555–572. [Google Scholar] [CrossRef] [PubMed]
- Chandrashekar, D.S.; Bashel, B.; Balasubramanya, S.A.H.; Creighton, C.J.; Ponce-Rodriguez, I.; Chakravarthi, B.V.S.K.; Varambally, S. UALCAN: A Portal for Facilitating Tumor Subgroup Gene Expression and Survival Analyses. Neoplasia 2017, 19, 649–658. [Google Scholar] [CrossRef]
- Tang, Z.; Kang, B.; Li, C.; Chen, T.; Zhang, Z. GEPIA2: An enhanced web server for large-scale expression profiling and interactive analysis. Nucleic Acids Res. 2019, 47, W556–W560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Ruotti, V.; Stewart, R.M.; Thomson, J.A.; Dewey, C.N. RNA-Seq gene expression estimation with read mapping uncertainty. Bioinformatics 2009, 26, 493–500. [Google Scholar] [CrossRef] [Green Version]
- Magraner-Pardo, L.; Laskowski, R.A.; Pons, T.; Thornton, J.M. A computational and structural analysis of germline and somatic variants affecting the DDR mechanism, and their impact on human diseases. Sci. Rep. 2021, 11, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Oughtred, R.; Stark, C.; Breitkreutz, B.J.; Rust, J.; Boucher, L.; Chang, C.; Kolas, N.; O’Donnell, L.; Leung, G.; McAdam, R.; et al. The BioGRID interaction database: 2019 update. Nucleic Acids Res. 2019, 47, D529–D541. [Google Scholar] [CrossRef] [Green Version]
- Orchard, S.; Ammari, M.; Aranda, B.; Breuza, L.; Briganti, L.; Broackes-Carter, F.; Campbell, N.H.; Chavali, G.; Chen, C.; Del-Toro, N.; et al. The MIntAct project—IntAct as a common curation platform for 11 molecular interaction databases. Nucleic Acids Res. 2014, 42, D358–D363. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Cancer Type |
|---|---|
| ACC | Adrenocortical carcinoma |
| AML | Acute myeloid leukemia |
| BLCA | Bladder urotelial carcinoma |
| BRCA | Breast invasive carcinoma |
| CESC | Cervical squamous cell carcinoma and endocervical adenocarcinoma |
| CHOL | Cholangiocarcinoma |
| COAD | Colorectal adenocarcinoma |
| DLBC | Lymphoid Neoplasm Diffuse Large B-cell Lymphoma |
| ESCA | Esophageal carcinoma |
| GBM | Glioblastoma multiforme |
| HNSC | Head and neck squamous cell carcinoma |
| KICH | Kidney Chromophobe carcinoma |
| KIRC | Kidney renal clear cell carcinoma |
| KIRP | Kidney renal papillary cell carcinoma |
| LGG | Brain Lower Grade Glioma |
| LIHC | Liver hepatocellular carcinoma |
| LUAD | Lung adenocarcinoma |
| LUSC | Lung squamous cell carcinoma |
| MESO | Mesothelioma |
| OV | Ovarian serous cystadenocarcinoma |
| PAAD | Pancreas adenocarcinoma |
| PCPG | Pheochromocytoma and paraganglioma |
| PRAD | Prostate adenocarcinoma |
| READ | Rectal adenocarcinoma |
| SARC | Sarcoma |
| SKCM | Skin cutaneous melanoma |
| STAD | Stomach adenocarcinoma |
| TGCT | Testicular Germ Cell Tumors |
| THCA | Thyroid carcinoma |
| THYM | Thymoma |
| UCEC | Uterine Corpus Endometrial Carcinoma |
| UCS | Uterine Carcinosarcoma |
| UVM | Uveal Melanoma |
| Cluster 1 | Log10 (p) |
| (R-HSA-5205685) Pink/Parkin Mediated Mitophagy | −39.5 |
| (R-HSA-5205647) Mitophagy | −37.1 |
| (R-HSA-9663891) Selective autophagy | −29.7 |
| Cluster 2 | Log10 (p) |
| (hsa05200) Pathways in cancer | −7.2 |
| (hsa04144) Endocytosis | −6.1 |
| (M153) PID P75 NTR Pathway | −5.9 |
| Cluster 3 | Log10 (p) |
| (GO:0032543) Mitochondrial translation | −7.2 |
| (GO:0140053) Mitochondrial gene expression | −6.9 |
| (R-HSA-5693607) Processing of DNA double-strand break ends | −5.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magraner-Pardo, L.; Gobelli, D.; de la Fuente, M.A.; Pons, T.; Simarro, M. Systematic Analysis of FASTK Gene Family Alterations in Cancer. Int. J. Mol. Sci. 2021, 22, 11337. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111337
Magraner-Pardo L, Gobelli D, de la Fuente MA, Pons T, Simarro M. Systematic Analysis of FASTK Gene Family Alterations in Cancer. International Journal of Molecular Sciences. 2021; 22(21):11337. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111337
Chicago/Turabian StyleMagraner-Pardo, Lorena, Dino Gobelli, Miguel A. de la Fuente, Tirso Pons, and María Simarro. 2021. "Systematic Analysis of FASTK Gene Family Alterations in Cancer" International Journal of Molecular Sciences 22, no. 21: 11337. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111337