
The Beneficial Effect of Rosmarinic Acid on Benzophenone-3-Induced Alterations in Human Skin Fibroblasts


Abstract
:1. Introduction
2. Results
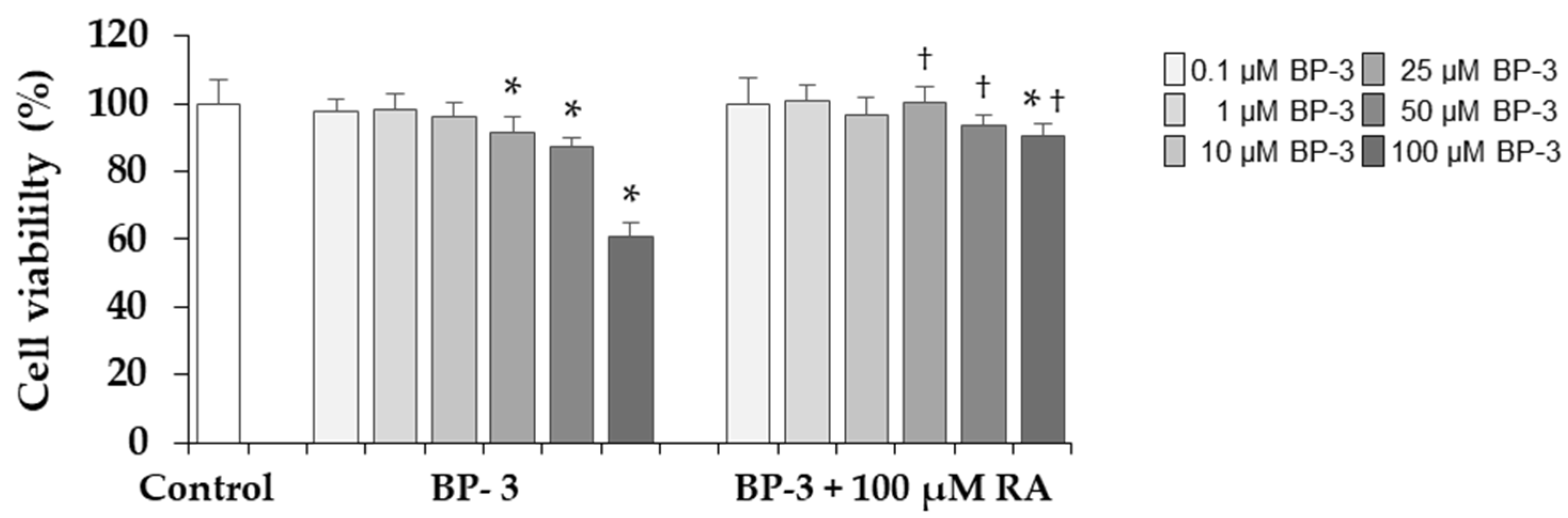
2.1. The Influence of BP-3 and BP-3 in Combination with RA on the Viability of Fibroblasts
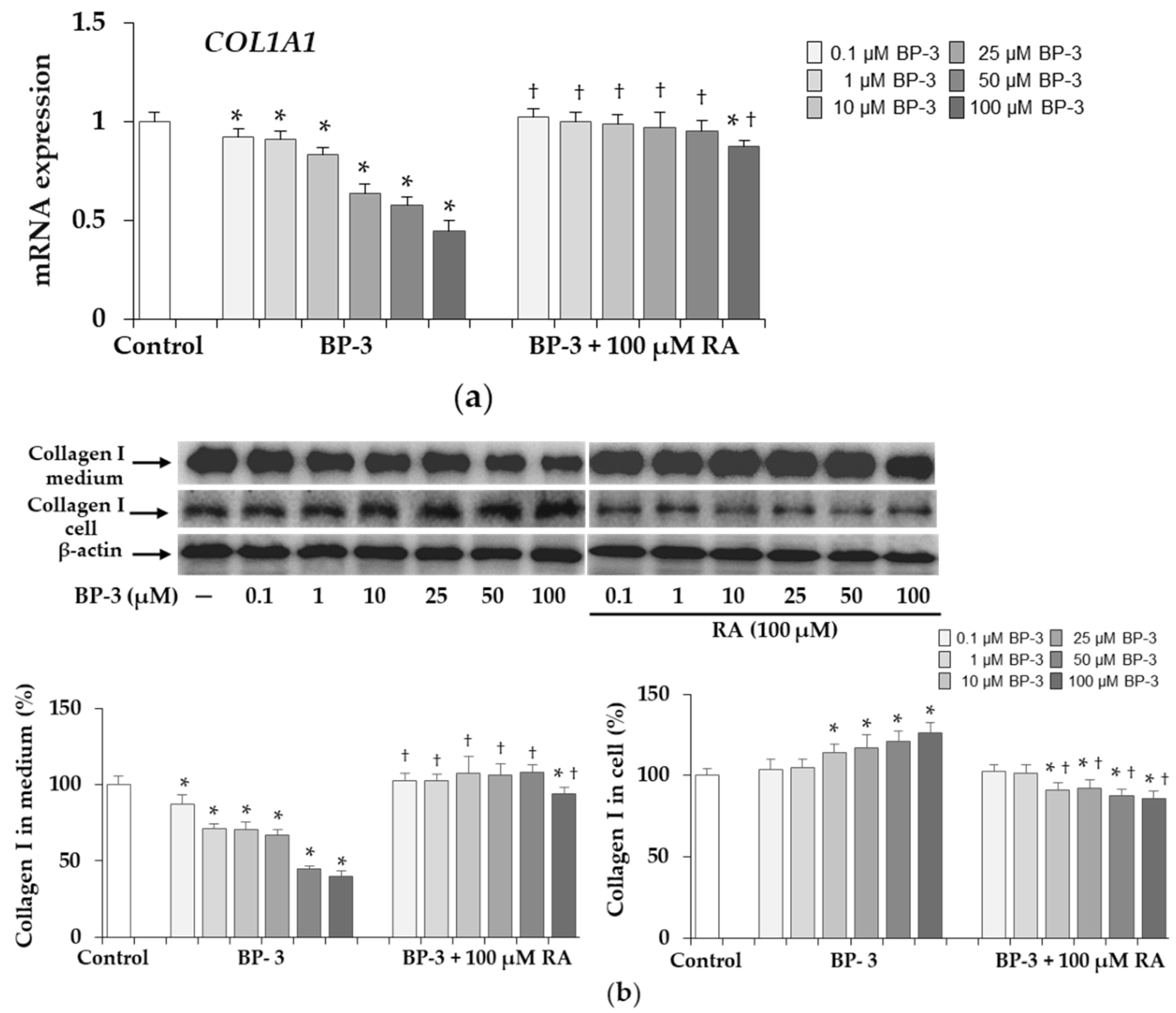
2.2. Effect of BP-3 and BP-3 in Combination with RA on the Expression of Collagen Type I in Fibroblasts
2.3. Expression of HSP47, Protein Disulfide Isomerase, and Glucosyltransferase in Fibroblasts under Influence of in BP-3 and BP-3 with RA
2.4. The Expression of Xbp-1 in Fibroblasts Exposed to BP-3 Alone and in Combination with RA
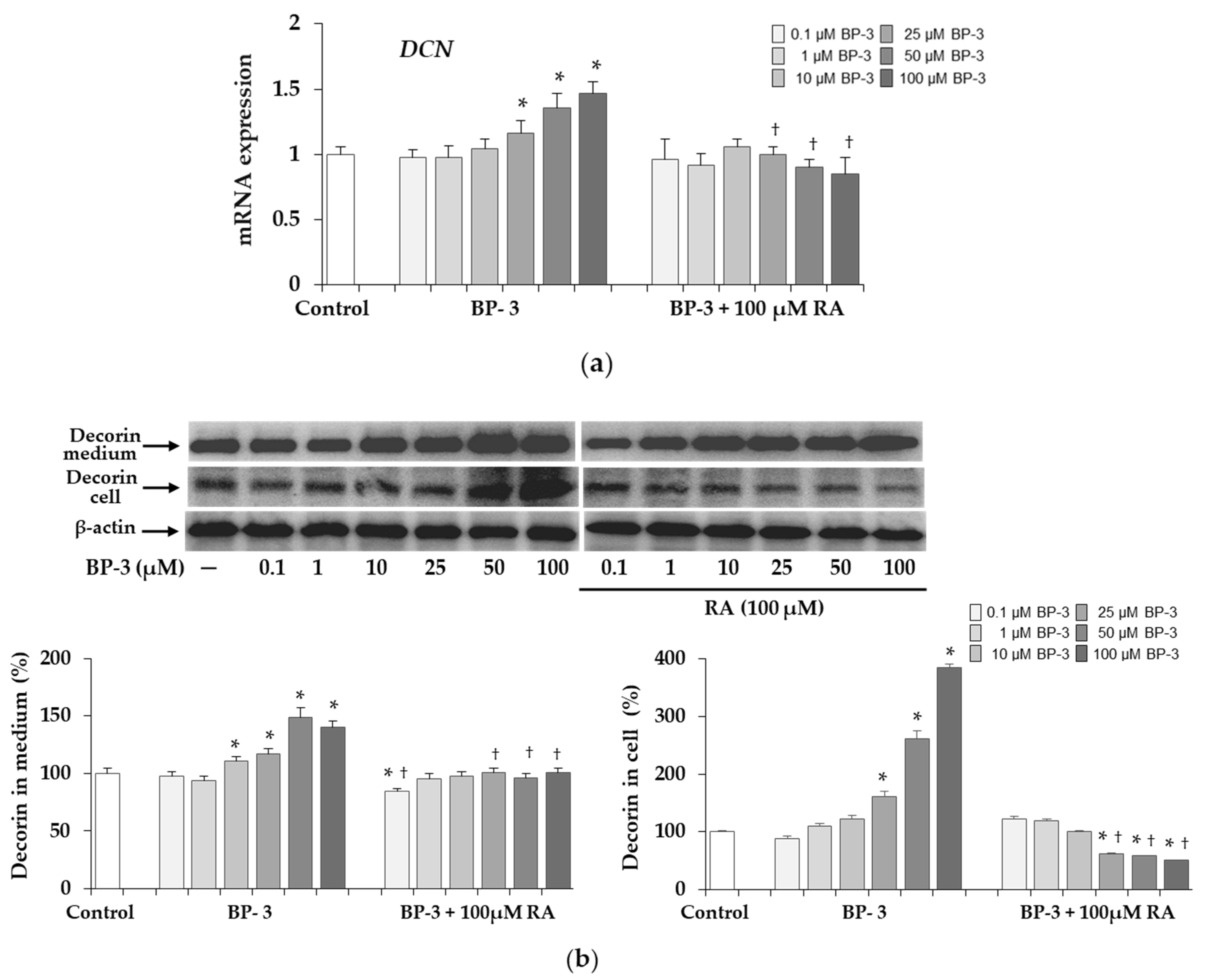
2.5. Expression of Decorin in Fibroblasts Exposed to BP-3 and BP-3 in Combination with RA
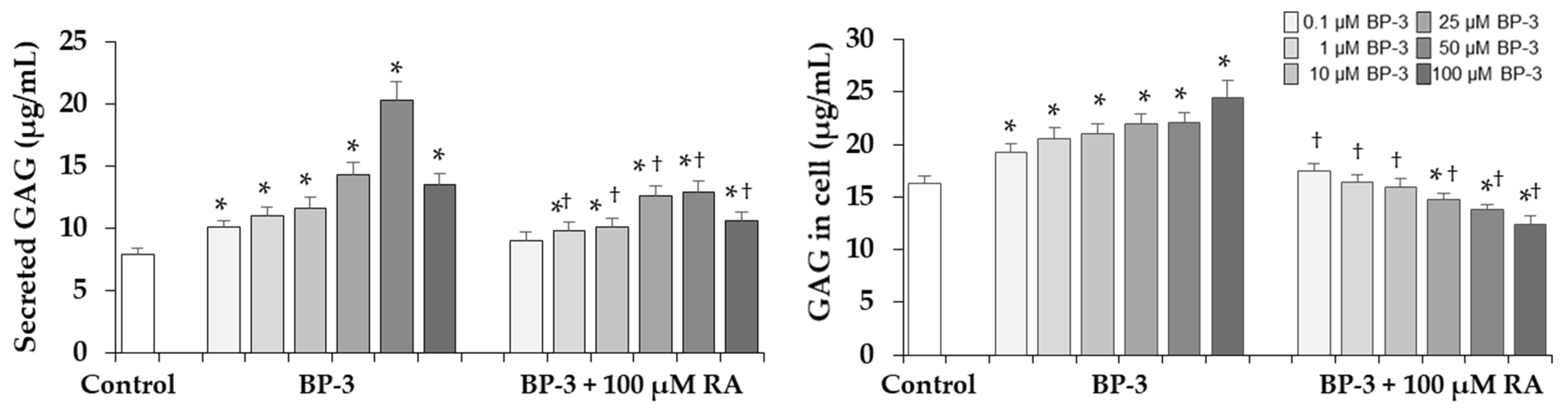
2.6. Effect of BP-3 Alone and in Combination with RA on Sulfated GAGs Content in Fibroblasts
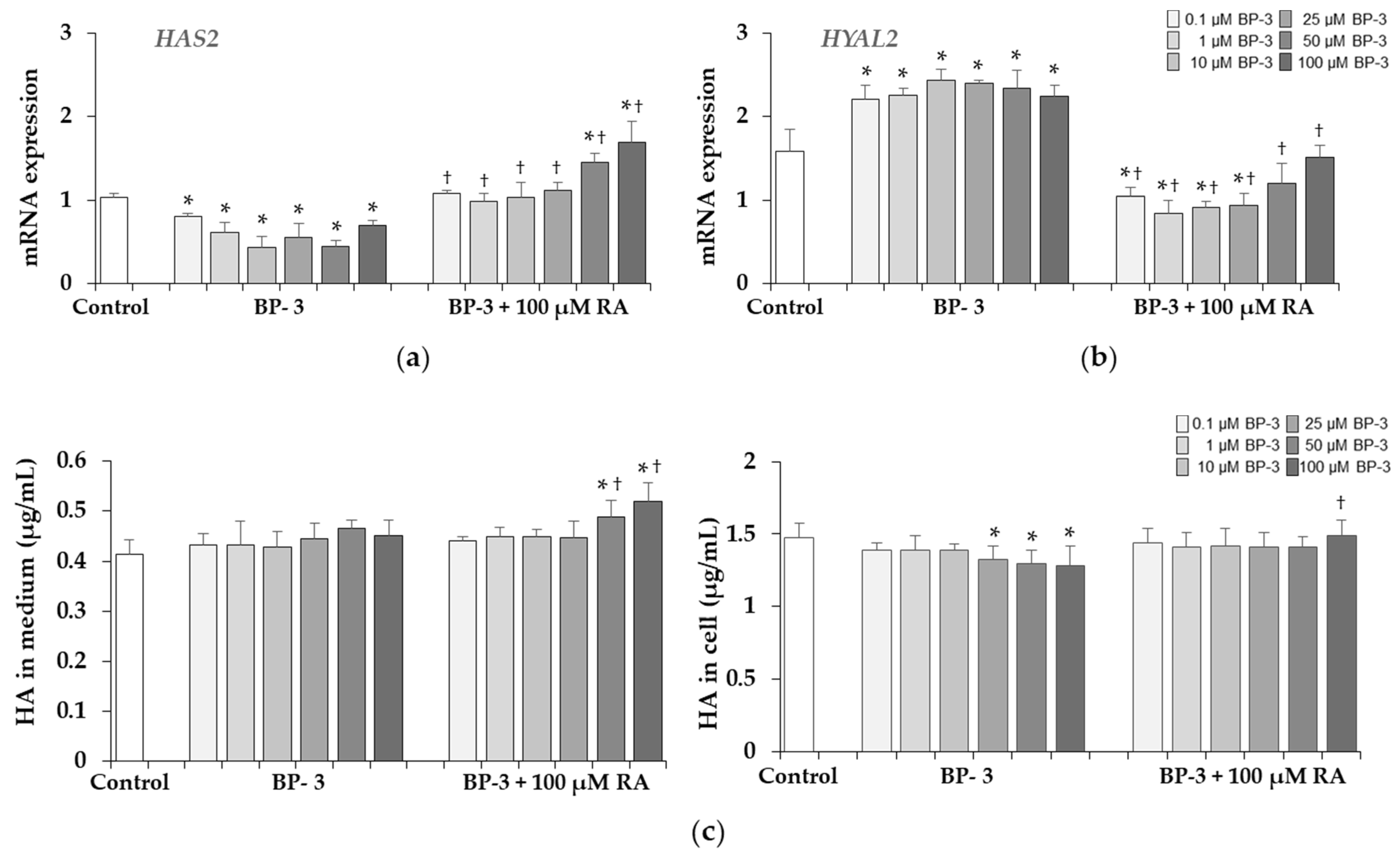
2.7. Effect of BP-3 Alone and in Combination with RA on the Expression of HAS2, HYAL2 and HA Content in Fibroblasts
2.8. The Elastin Content and Activity of Elastase in Fibroblasts Exposed to BP-3 Alone and in Combination with RA
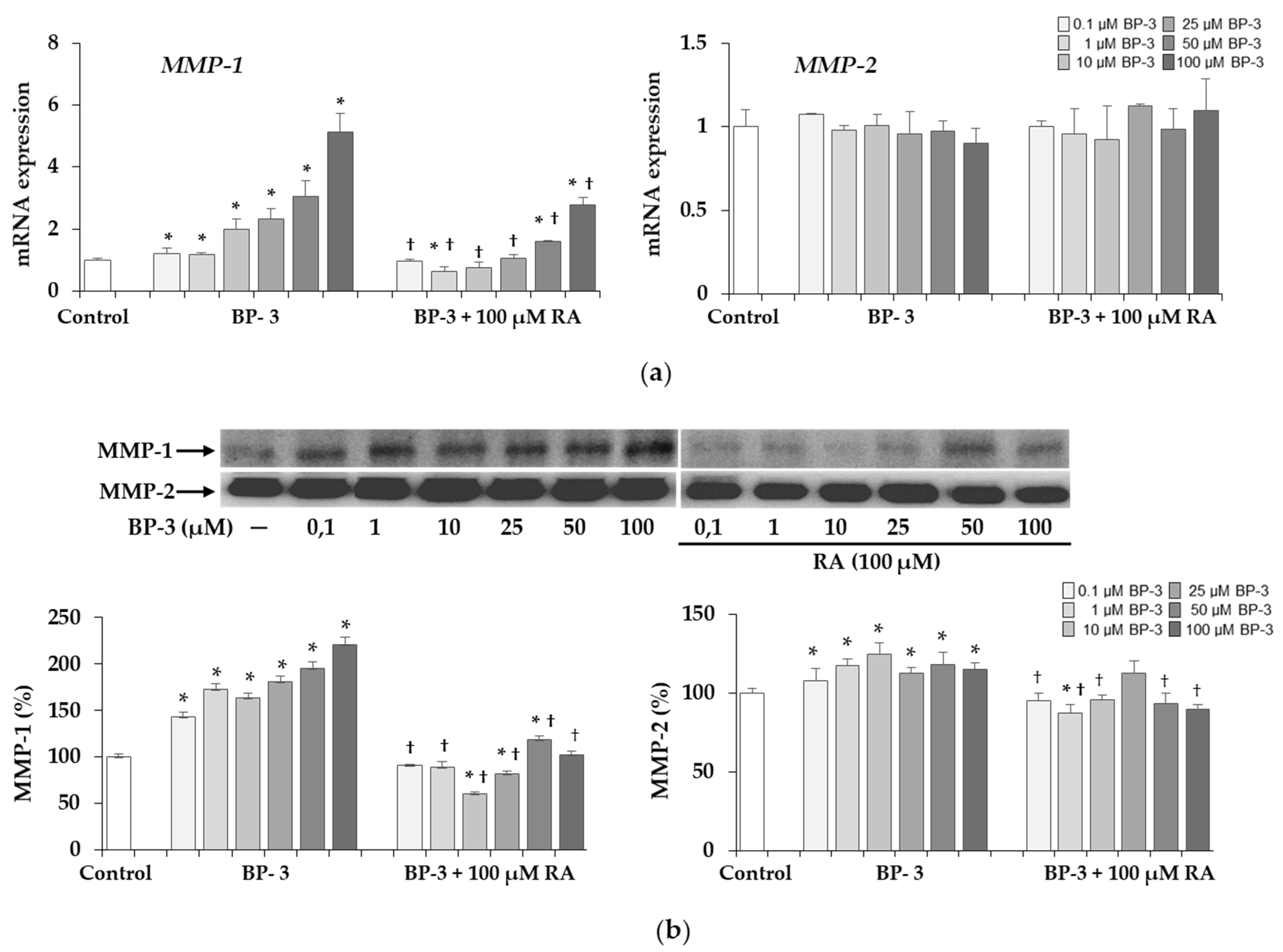
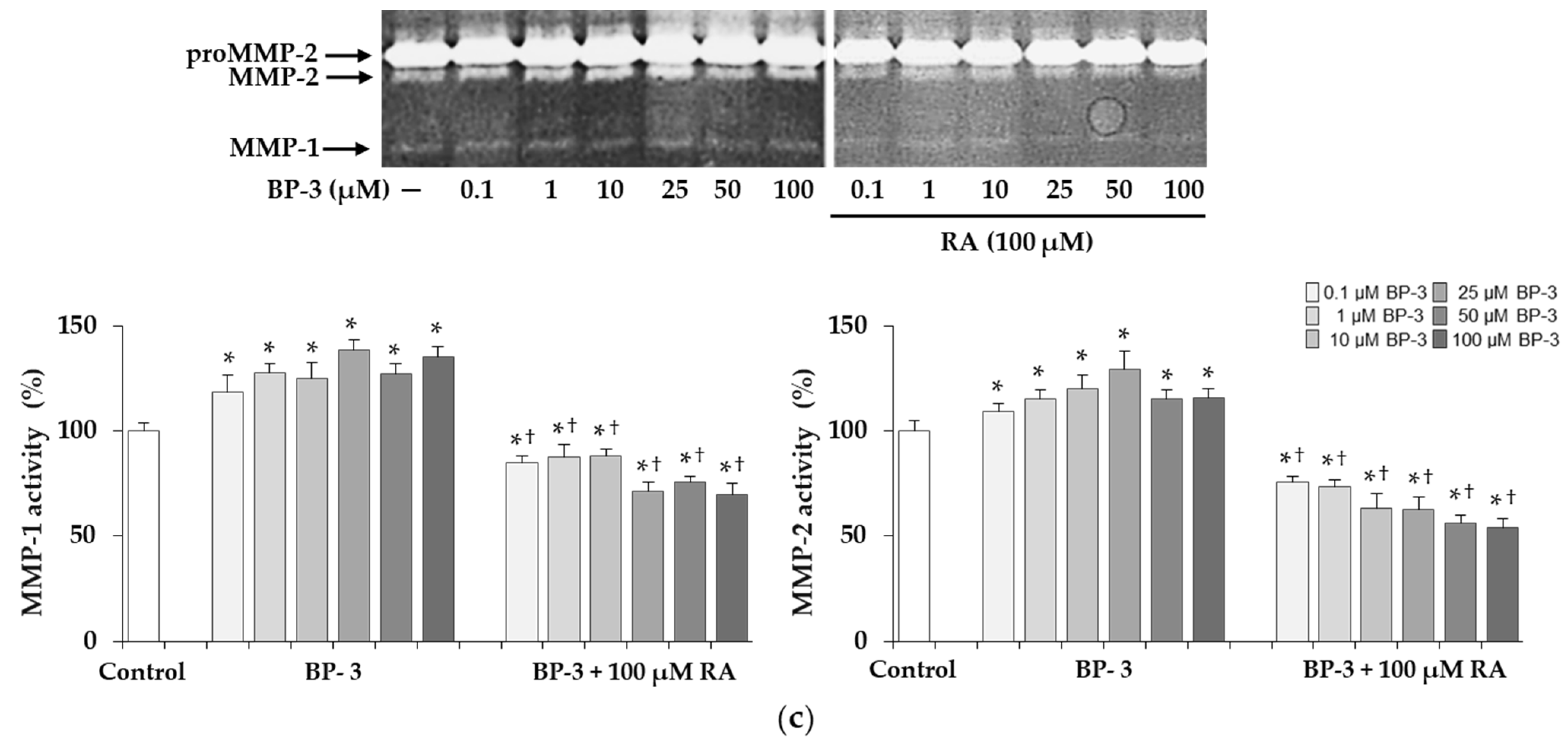
2.9. The Expression and Activity of MMP-1 and MMP-2 under Influence of BP-3 Alone and in Combination with RA in Fibroblasts
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Fibroblast Culture and Treatment
4.3. Determination of Cell Viability with MTT Test
4.4. Quantitative Real-Time PCR Analysis
4.5. XBP1 Splicing Analysis
4.6. Western Blot
4.7. The Quantitative Measurement of Total Sulfated Glycosaminoglycans, Hyaluronic Acid, and Elastin
4.8. The Measurement of Elastase Activity
4.9. Zymography
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| BP-3 | Benzophenone-3 |
| BiP | Binding immunoglobulin protein |
| DCN | Gene coding decorin |
| ECM | Extracellular matrix |
| EHM | Ethylhexyl methoxycinnamate |
| ER | Endoplasmic reticulum |
| GAG | Glycosaminoglycan |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| GLT25D1 | Collagen beta(1-O)galactosyltransferase 1 |
| HA | Hyaluronic acid |
| HAS-2 | Hyaluronan Synthase 2 |
| HS | Homosalate |
| HSP47 | Heat Shock Protein 47 |
| HYAL-2 | Hyaluronidase 2 |
| IRE1α | Inositol requiring enzyme 1 |
| 4-MBC | 4-methylbenzilidenecamphor |
| MMP | Matrix metalloproteinase |
| OI | Osteogenesis imperfecta |
| OMC | Octylmethoxycinnamate |
| P4HB | Prolyl 4-hydroxylase subunit beta |
| PDI | Protein disulfide isomerase |
| PG | Proteoglycan |
| RA | Rosmarinic acid |
| ROS | Reactive oxygen species |
| STANA | N-succinyl-tri-alanyl-p-nitroanilide |
| UPR | Unfolded protein response |
| UV | Ultraviolet |
| Xbp-1s | X-box 1 binding protein |
References
- Watson, M.; Holman, D.M.; Maguire-Eisen, M. Ultraviolet radiation exposure and its impact on skin cancer risk. Semin. Oncol. Nurs. 2016, 32, 241–254. [Google Scholar] [CrossRef] [Green Version]
- Lee, L.Y.; Liu, S.X. Pathogenesis of photoaging in human dermal fibroblasts. Int. J. Dermatol. Venereol. 2020, 3, 37–42. [Google Scholar] [CrossRef]
- Mancuso, J.B.; Maruthi, R.; Wang, S.Q.; Lim, H.W. Sunscreens: An update. Am. J. Clin. Dermatol. 2017, 18, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Ngoc, L.T.N.; Tran, V.V.; Moon, J.-Y.; Chae, M.; Park, D.; Lee, Y.-C. Recent trends of sunscreen cosmetic: An update review. Cosmetics 2019, 6, 64. [Google Scholar] [CrossRef] [Green Version]
- Fivenson, D.; Sabzevari, N.; Qiblawi, S.; Blitz, J.; Norton, B.B.; Norton, S.A. Sunscreens: UV filters to protect us: Part 2—Increasing awareness of UV filters and their potential toxicities to us and our environment. Int. J. Womens Dermatol. 2020, 7, 45–69. [Google Scholar] [CrossRef]
- Sabzevari, N.; Qiblawi, S.; Norton, S.A.; Fivenson, D. Sunscreens: UV filters to protect us: Part 1: Changing regulations and choices for optimal sun protection. Int. J. Womens Dermatol. 2021, 7, 28–44. [Google Scholar] [CrossRef]
- Siller, A.; Blaszak, S.C.; Lazar, M.; Olasz Harken, E. Update about the effects of the sunscreen ingredients oxybenzone and octinoxate on humans and the environment. Plast. Surg. Nurs. 2019, 39, 157–160. [Google Scholar] [CrossRef] [PubMed]
- DiNardo, J.C.; Downs, C.A. Dermatological and environmental toxicological impact of the sunscreen ingredient oxybenzone/benzophenone-3. J. Cosmet. Dermatol. 2018, 17, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Schlumpf, M.; Cotton, B.; Conscience, M.; Haller, V.; Steinmann, B.; Lichtensteiger, W. In vitro and in vivo estrogenicity of UV screens. Environ. Health Perspect. 2001, 109, 239–244. [Google Scholar] [CrossRef]
- Kim, S.; Choi, K. Occurrences, toxicities, and ecological risks of benzophenone-3, a common component of organic sunscreen products: A mini-review. Environ. Int. 2014, 70, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Alamer, M.; Darbre, P.D. Effects of exposure to six chemical ultraviolet filters commonly used in personal care products on motility of MCF-7 and MDA-MB-231 human breast cancer cells in vitro. J. Appl. Toxicol. 2018, 38, 148–159. [Google Scholar] [CrossRef]
- Barr, L.; Alamer, M.; Darbre, P.D. Measurement of concentrations of four chemical ultraviolet filters in human breast tissue at serial locations across the breast. J. Appl. Toxicol. 2018, 38, 1112–1120. [Google Scholar] [CrossRef]
- Kunisue, T.; Chen, Z.; Buck Louis, G.M.; Sundaram, R.; Hediger, M.L.; Sun, L.; Kannan, K. Urinary concentrations of benzophenone-type UV filters in U.S. women and their association with endometriosis. Environ. Sci. Technol. 2012, 46, 4624–4632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huo, W.; Cai, P.; Chen, M.; Li, H.; Tang, J.; Xu, C.; Zhu, D.; Tang, W.; Xia, Y. The relationship between prenatal exposure to BP-3 and Hirschsprung’s disease. Chemosphere 2016, 144, 1091–1097. [Google Scholar] [CrossRef] [PubMed]
- Scinicariello, F.; Buser, M.C. Serum testosterone concentrations and urinary bisphenol A, benzophenone-3, triclosan, and paraben levels in male and female children and adolescents: NHANES 2011–2012. Environ. Health Perspect. 2016, 124, 1898–1904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wnuk, A.; Rzemieniec, J.; Lasoń, W.; Krzeptowski, W.; Kajta, M. Benzophenone-3 impairs autophagy, alters epigenetic status, and disrupts retinoid X receptor signaling in apoptotic neuronal cells. Mol. Neurobiol. 2018, 55, 5059–5074. [Google Scholar] [CrossRef] [Green Version]
- Wnuk, A.; Rzemieniec, J.; Staroń, J.; Litwa, E.; Lasoń, W.; Bojarski, A.; Kajta, M. Prenatal exposure to benzophenone-3 impairs autophagy, disrupts RXRs/PPARγ signaling, and alters epigenetic and post-translational statuses in brain neurons. Mol. Neurobiol. 2019, 56, 4820–4837. [Google Scholar] [CrossRef] [Green Version]
- Heurung, A.R.; Raju, S.I.; Warshaw, E.M. Benzophenones. Dermatitis 2014, 25, 3–10. [Google Scholar] [CrossRef]
- Schneider, S.L.; Lim, H.W. Review of environmental effects of oxybenzone and other sunscreen active ingredients. J. Am. Acad. Dermatol. 2019, 80, 266–271. [Google Scholar] [CrossRef]
- Commission Regulation (EU) 2017/238. Annex VI to Regulation (EC) No. 1223/2009 of the European Parliament and of the Council on Cosmetic Products. 2017. Available online: http://data.europa.eu/eli/reg/2017/238/oj (accessed on 11 February 2017).
- Hayden, C.G.; Roberts, M.S.; Benson, H.A. Systemic absorption of sunscreen after topical application. Lancet 1997, 350, 863–864. [Google Scholar] [CrossRef]
- Gonzalez, H.; Farbrot, A.; Larkö, O.; Wennberg, A.M. Percutaneous absorption of the sunscreen benzophenone-3 after repeated whole-body applications, with and without ultraviolet irradiation. Br. J. Dermatol. 2006, 154, 337–340. [Google Scholar] [CrossRef]
- Jiang, R.; Roberts, M.S.; Collins, D.M.; Benson, H.A. Absorption of sunscreens across human skin: An evaluation of commercial products for children and adults. Br. J. Clin. Pharmacol. 1999, 48, 635–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janjua, N.R.; Mogensen, B.; Andersson, A.M.; Petersen, J.H.; Henriksen, M.; Skakkebaek, N.E.; Wulf, H.C. Systemic absorption of the sunscreens benzophenone-3, octyl-methoxycinnamate, and 3-(4-methylbenzylidene) camphor after whole-body topical application and reproductive hormone levels in humans. J. Investig. Dermatol. 2004, 123, 57–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janjua, N.R.; Kongshoj, B.; Andersson, A.M.; Wulf, H.C. Sunscreens in human plasma and urine after repeated whole-body topical application. J. Eur. Acad. Dermatol. Venereol. 2008, 22, 456–461. [Google Scholar] [CrossRef] [PubMed]
- Matta, M.K.; Florian, J.; Zusterzeel, R.; Pilli, N.R.; Patel, V.; Volpe, D.A.; Yang, Y.; Oh, L.; Bashaw, E.; Zineh, I.; et al. Effect of sunscreen application on plasma concentration of sunscreen active ingredients: A ramdomized clinical trial. JAMA 2020, 323, 256–267. [Google Scholar] [CrossRef] [PubMed]
- Chatelain, E.; Gabard, B.; Surber, C. Skin penetration and sun protection factor of five UV filters: Effect of the vehicle. Skin Pharmacol. Appl. Skin Physiol. 2003, 16, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Calafat, A.M.; Wong, L.Y.; Ye, X.; Reidy, J.A.; Needham, L.L. Concentrations of the sunscreen agent benzophenone-3 in residents of the United States: National Health and Nutrition Examination Survey 2003–2004. Environ. Health Perspect. 2008, 116, 893–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okereke, C.S.; Abdel-Rhaman, M.S.; Friedman, M.A. Disposition of benzophenone-3 after dermal administration in male rats. Toxicol. Lett. 1994, 73, 113–122. [Google Scholar] [CrossRef]
- Fediuk, D.J.; Wang, T.; Raizman, J.E.; Parkinson, F.E.; Gu, X. Tissue deposition of the insect repellent DEET and the sunscreen oxybenzone from repeated topical skin applications in rats. Int. J. Toxicol. 2010, 29, 594–603. [Google Scholar] [CrossRef]
- Wang, L.; Asimakopoulos, A.G.; Kannan, K. Accumulation of 19 environmental phenolic and xenobiotic heterocyclic aromatic compounds in human adipose tissue. Environ. Int. 2015, 78, 45–50. [Google Scholar] [CrossRef]
- Valle-Sistac, J.; Molins-Delgado, D.; Díaz, M.; Ibáñez, L.; Barceló, D.; Silvia Díaz-Cruz, M. Determination of parabens and benzophenone-type UV filters in human placenta. First description of the existence of benzyl paraben and benzophenone-4. Environ. Int. 2016, 88, 243–249. [Google Scholar] [CrossRef]
- Hany, J.; Nagel, R. Detection of sunscreen agents in human breast milk. Dtsch. Lebensmittel-Rundsch. 1995, 91, 341–345. [Google Scholar]
- Xu, C.; Parsons, P.G. Cell cycle delay, mitochondrial stress and uptake of hydrophobic cations induced by sunscreens in cultured human cells. Photochem. Photobiol. 1999, 69, 611–616. [Google Scholar] [CrossRef]
- Majewska, N.; Zaręba, I.; Surażyński, A.; Galicka, A. Methylparaben-induced decrease in collagen production and viability of cultured human dermal fibroblasts. J. Appl. Toxicol. 2017, 37, 1117–1124. [Google Scholar] [CrossRef] [PubMed]
- Matwiejczuk, N.; Galicka, A.; Zaręba, I.; Brzóska, M.M. The protective effect of rosmarinic acid against unfavorable influence of methylparaben and propylparaben on collagen in human skin fibroblasts. Nutrients 2020, 12, 1282. [Google Scholar] [CrossRef] [PubMed]
- Fernando, P.M.; Piao, M.J.; Kang, K.A.; Ryu, Y.S.; Hewage, S.R.; Chae, S.W.; Hyun, J.W. Rosmarinic acid attenuates cell damage against UVB radiation-induced oxidative stress via enhancing antioxidant effects in human HaCaT cells. Biomol. Ther. 2016, 24, 75–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Macedo, L.M.; Santos, É.M.D.; Militão, L.; Tundisi, L.L.; Ataide, J.A.; Souto, E.B.; Mazzola, P.G. Rosemary (Rosmarinus officinalis L., syn Sylvia rosmarinus Spenn.) and its topical applications: A review. Plants 2020, 9, 651. [Google Scholar] [CrossRef]
- Mao, F.; Wu, Z.; Shen, Y.; Ren, Y.; Yang, M.; Li, X.; Zhang, L. Regulatory mechanism of rosemary extract (Rosmarinus officinalis) on human skin fibroblasts during ultraviolet A photoaging. Iran. J. Sci. Technol. Trans Sci. 2020, 45, 427–436. [Google Scholar] [CrossRef]
- González-Minero, F.J.; Bravo-Díaz, L.; Ayala-Gómez, A. Rosmarinus officinalis L. (Rosemary): An ancient plant with uses in personal healthcare and cosmetics. Cosmetics 2020, 7, 77. [Google Scholar] [CrossRef]
- Martin, R.; Pierrard, C.; Lejeune, F.; Hilaire, P.; Breton, L.; Bernerd, F. Photoprotective effect of a water-soluble extract of Rosmarinus officinalis L. against UV-induced matrix metalloproteinase-1 in human dermal fibroblasts and reconstructed skin. Eur. J. Dermatol. 2008, 18, 128–135. [Google Scholar] [CrossRef]
- Pattananandecha, T.; Apichai, S.; Julsrigival, J.; Ungsurungsie, M.; Samuhasaneetoo, S.; Chulasiri, P.; Kwankhao, P.; Pitiporn, S.; Ogata, F.; Kawasaki, N.; et al. Antioxidant activity and anti-photoaging effects on UVA-irradiated human fibroblasts of rosmarinic acid enriched extract prepared from Thunbergia laurifolia leaves. Plants 2021, 10, 1648. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, M.; Okano, Y.; Masaki, H. An Ocimum basilicum extract containing rosmarinic acid restores the disruption of collagen fibers caused by repetitive UVA irradiation of dermal fibroblasts. J. Oleo Sci. 2020, 69, 1487–1495. [Google Scholar] [CrossRef] [PubMed]
- Theocharis, A.D.; Skandalis, S.S.; Gialeli, C.; Karamanos, N.K. Extracellular matrix structure. Adv. Drug Deliv. Rev. 2016, 97, 4–27. [Google Scholar] [CrossRef]
- Kielty, C.M.; Grant, M.E. The collagen family: Structure, assembly, and organization in the extracellular matrix. In Connective Tissue and Its Heritable Disorders; Royce, P.M., Steinmann, B., Eds.; Wiley: Hoboken, NJ, USA, 2002; pp. 159–221. [Google Scholar] [CrossRef]
- Gajko-Galicka, A. Mutations in type I collagen genes resulting in osteogenesis imperfecta in humans. Acta Biochim. Pol. 2002, 49, 433–441. [Google Scholar] [CrossRef] [Green Version]
- Besio, R.; Garibaldi, N.; Leoni, L.; Cipolla, L.; Sabbioneda, S.; Biggiogera, M.; Mottes, M.; Aglan, M.; Otaify, G.A.; Temtamy, S.A.; et al. Cellular stress due to impairment of collagen prolyl hydroxylation complex is rescued by the chaperone 4-phenylbutyrate. Dis. Model. Mech. 2019, 12, dmm038521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garibaldi, N.; Contento, B.M.; Babini, G.; Morini, J.; Siciliani, S.; Biggiogera, M.; Raspanti, M.; Marini, J.C.; Rossi, A.; Forlino, A.; et al. Targeting cellular stress in vitro improves osteoblast homeostasis, matrix collagen content and mineralization in two murine models of osteogenesis imperfecta. Matrix Biol. 2021, 98, 1–20. [Google Scholar] [CrossRef]
- Schegg, B.; Hülsmeier, A.J.; Rutschmann, C.; Maag, C.; Hennet, T. Core glycosylation of collagen is initiated by two beta(1-O)galactosyltransferases. Mol. Cell Biol. 2009, 29, 943–952. [Google Scholar] [CrossRef] [Green Version]
- Nagai, N.; Hosokawa, M.; Itohara, S.; Adachi, E.; Matsushita, T.; Hosokawa, N.; Nagata, K. Embryonic lethality of molecular chaperone hsp47 knockout mice is associated with defects in collagen biosynthesis. J. Cell Biol. 2000, 150, 1499–1506. [Google Scholar] [CrossRef] [Green Version]
- Sepulveda, D.; Rojas-Rivera, D.; Rodríguez, D.A.; Groenendyk, J.; Köhler, A.; Lebeaupin, C.; Ito, S.; Urra, H.; Carreras-Sureda, A.; Hazari, Y.; et al. Interactome screening identifies the ER luminal chaperone Hsp47 as a regulator of the unfolded protein response transducer IRE1α. Mol. Cell. 2018, 69, 238–252. [Google Scholar] [CrossRef] [Green Version]
- Laronha, H.; Caldeira, J. Structure and function of human matrix metalloproteinases. Cells. 2020, 9, 1076. [Google Scholar] [CrossRef]
- Rühland, C.; Schönherr, E.; Robenek, H.; Hansen, U.; Iozzo, R.V.; Bruckner, P.; Seidler, D.G. The glycosaminoglycan chain of decorin plays an important role in collagen fibril formation at the early stages of fibrillogenesis. FEBS J. 2007, 274, 4246–4255. [Google Scholar] [CrossRef]
- Ishikawa, Y.; Rubin, K.; Bächinger, H.P.; Kalamajski, S. The endoplasmic reticulum-resident collagen chaperone Hsp47 interacts with and promotes the secretion of decorin, fibromodulin, and lumican. J. Biol. Chem. 2018, 293, 13707–13716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, W.J.; Ahn, H.M.; Roh, H.; Na, Y.; Choi, I.K.; Lee, J.H.; Kim, Y.O.; Lew, D.H.; Yun, C.O. Decorin expressing adenovirus decreases collagen synthesis and upregulates MMP expression in keloid fibroblasts and keloid spheroids. Exp. Dermatol. 2005, 24, 591–597. [Google Scholar] [CrossRef] [PubMed]
- Lochner, K.; Gaemlich, A.; Südel, K.M.; Venzke, K.; Moll, I.; Knott, A.; Stäb, F.; Wenck, H.; Döring, O.; Böttger, M.; et al. Expression of decorin and collagens I and III in different layers of human skin in vivo: A laser capture microdissection study. Biogerontology 2007, 8, 269–282. [Google Scholar] [CrossRef]
- Carrino, D.A.; Sorrell, J.M.; Caplan, A.I. Age-related changes in the proteoglycans of human skin. Arch. Biochem. Biophys. 2000, 373, 91–101. [Google Scholar] [CrossRef]
- Shin, J.W.; Kwon, S.H.; Choi, J.Y.; Na, J.I.; Huh, C.H.; Choi, H.R.; Park, K.C. Molecular Mechanisms of Dermal Aging and Antiaging Approaches. Int. J. Mol. Sci. 2019, 20, 2126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papakonstantinou, E.; Roth, M.; Karakiulakis, G. Hyaluronic acid: A key molecule in skin aging. Dermatoendocrinology 2012, 4, 253–258. [Google Scholar] [CrossRef] [Green Version]
- Hanson, K.M.; Gratton, E.; Bardeen, C.J. Sunscreen enhancement of UV-induced reactive oxygen species in the skin. Free Radic. Biol. Med. 2006, 41, 1205–1212. [Google Scholar] [CrossRef] [Green Version]
- Schallreuter, K.U.; Wood, J.M.; Farwell, D.W.; Moore, J.; Edwards, H.G. Oxybenzone oxidation following solar irradiation of skin: Photoprotection versus antioxidant inactivation. J. Investig. Dermatol. 1996, 106, 583–586. [Google Scholar] [CrossRef] [Green Version]
- Vigetti, D.; Viola, M.; Karousou, E.; De Luca, G.; Passi, A. Metabolic control of hyaluronan synthases. Matrix Biol. 2014, 35, 8–13. [Google Scholar] [CrossRef] [Green Version]
- Moczar, M.; Robert, L. Stimulation of cell proliferation by hyaluronidase during in vitro aging of human skin fibroblasts. Exp. Gerontol. 1993, 28, 59–68. [Google Scholar] [CrossRef]
- Buhren, B.A.; Schrumpf, H.; Gorges, K.; Reiners, O.; Bölke, E.; Fischer, J.W.; Homey, B.; Gerber, P.A. Dose- and time-dependent effects of hyaluronidase on structural cells and the extracellular matrix of the skin. Eur. J. Med. Res. 2020, 25, 60. [Google Scholar] [CrossRef] [PubMed]
- Uitto, J. The role of elastin and collagen in cutaneous aging: Intrinsic aging versus photoexposure. J. Drugs Dermatol. 2008, 7, 12–16. [Google Scholar]
- Sutkowska, J.; Hupert, N.; Gawron, K.; Strawa, J.W.; Tomczyk, M.; Forlino, A.; Galicka, A. The stimulating effect of rosmarinic acid and extracts from rosemary and lemon balm on collagen type I biosynthesis in osteogenesis imperfecta type I skin fibroblasts. Pharmaceutics 2021, 13, 938. [Google Scholar] [CrossRef] [PubMed]
- Tampucci, S.; Burgalassi, S.; Chetoni, P.; Monti, D. Cutaneous permeation and penetration of sunscreens: Formulation strategies and in vitro methods. Cosmetics 2018, 5, 1. [Google Scholar] [CrossRef] [Green Version]
- Shanbhag, S.; Nayak, A.; Narayan, R.; Nayak, U.Y. Anti-aging and sunscreens: Paradigm shift in cosmetics. Adv. Pharm. Bull. 2019, 9, 348–359. [Google Scholar] [CrossRef] [Green Version]
- Chaiyana, W.; Chansakaow, S.; Intasai, N.; Kiattisin, K.; Lee, K.H.; Lin, W.C.; Lue, S.C.; Leelapornpisid, P. Chemical constituents, antioxidant, anti-MMPs, and anti-hyaluronidase activities of Thunbergia laurifolia Lindl. leaf extracts for skin aging and skin damage prevention. Molecules 2020, 25, 1923. [Google Scholar] [CrossRef] [PubMed] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence | |
|---|---|---|
| COL1A1 | forward | 5′-GCTCGTGGAAATGATGGTGC-3′ |
| reverse | 5′-ACCCTGGGGACCTTCAGAG-3′ | |
| MMP-1 | forward | 5′-CATTGATGGCATCCAAGCC-3′ |
| reverse | 5′-GGCTGGACAGGATTTTGGG-3′ | |
| MMP-2 | forward | 5′-TGTGTCTTCCCCTTCACTTT-3′ |
| reverse | 5′-GATCTGAGCGATGCCATCAA-3′ | |
| DCN | forward | 5′-TGATGCAGCTAGCCTGAAAGG-3′ |
| reverse | 5′-AGGCGTGTTGGCCAGAGAG-3′ | |
| HSP47 | forward | 5′-AGAGGTCACCAAGGATGTGGAG-3′ |
| reverse | 5′-TGGGGCATGAGGATGATGAG-3′ | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galicka, A.; Sutkowska-Skolimowska, J. The Beneficial Effect of Rosmarinic Acid on Benzophenone-3-Induced Alterations in Human Skin Fibroblasts. Int. J. Mol. Sci. 2021, 22, 11451. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111451
Galicka A, Sutkowska-Skolimowska J. The Beneficial Effect of Rosmarinic Acid on Benzophenone-3-Induced Alterations in Human Skin Fibroblasts. International Journal of Molecular Sciences. 2021; 22(21):11451. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111451
Chicago/Turabian StyleGalicka, Anna, and Joanna Sutkowska-Skolimowska. 2021. "The Beneficial Effect of Rosmarinic Acid on Benzophenone-3-Induced Alterations in Human Skin Fibroblasts" International Journal of Molecular Sciences 22, no. 21: 11451. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111451