
Mechanisms of MEHP Inhibitory Action and Analysis of Potential Replacement Plasticizers on Leydig Cell Steroidogenesis



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
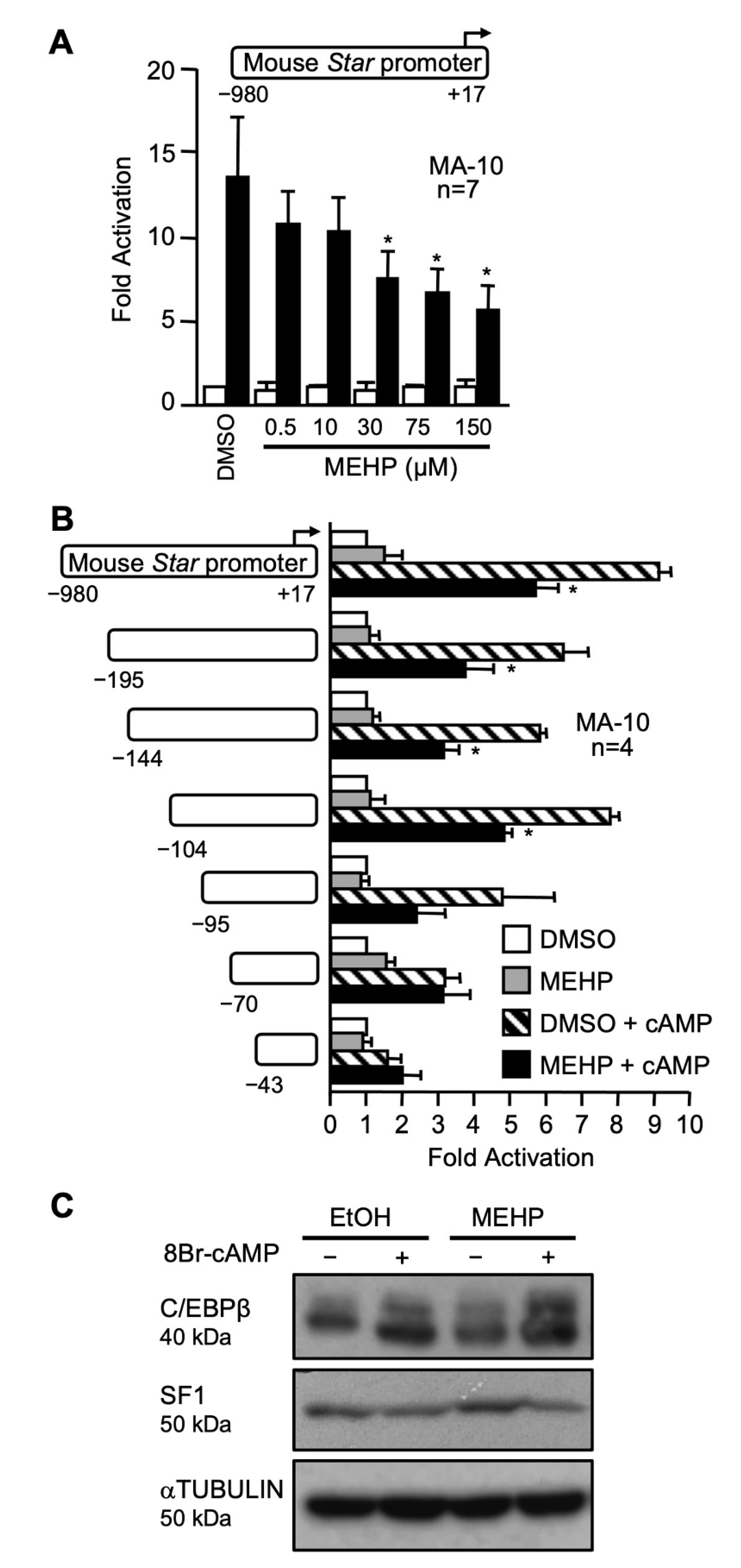
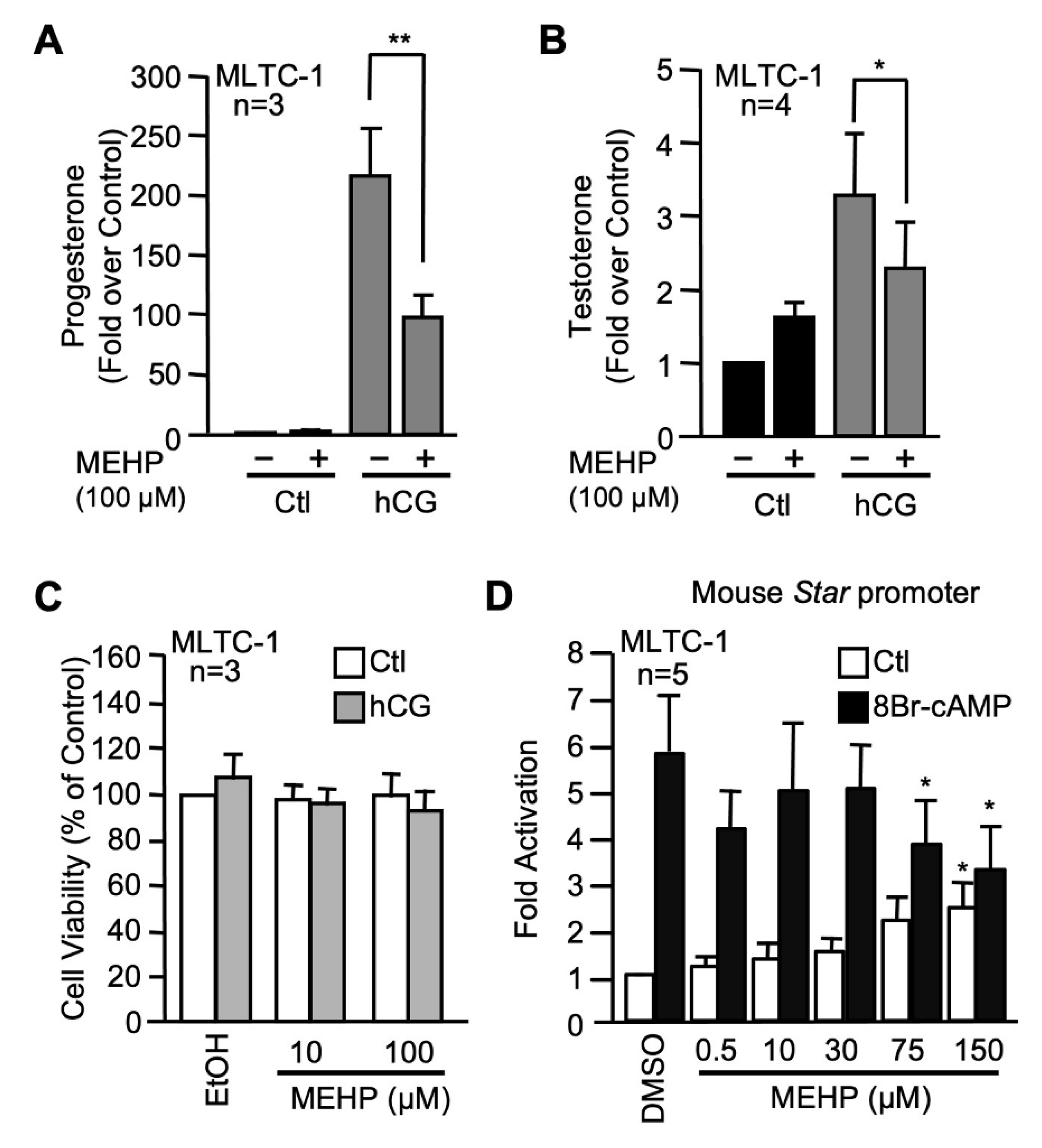
2.1. MEHP Represses Steroidogenesis by Affecting Star Gene Transcription
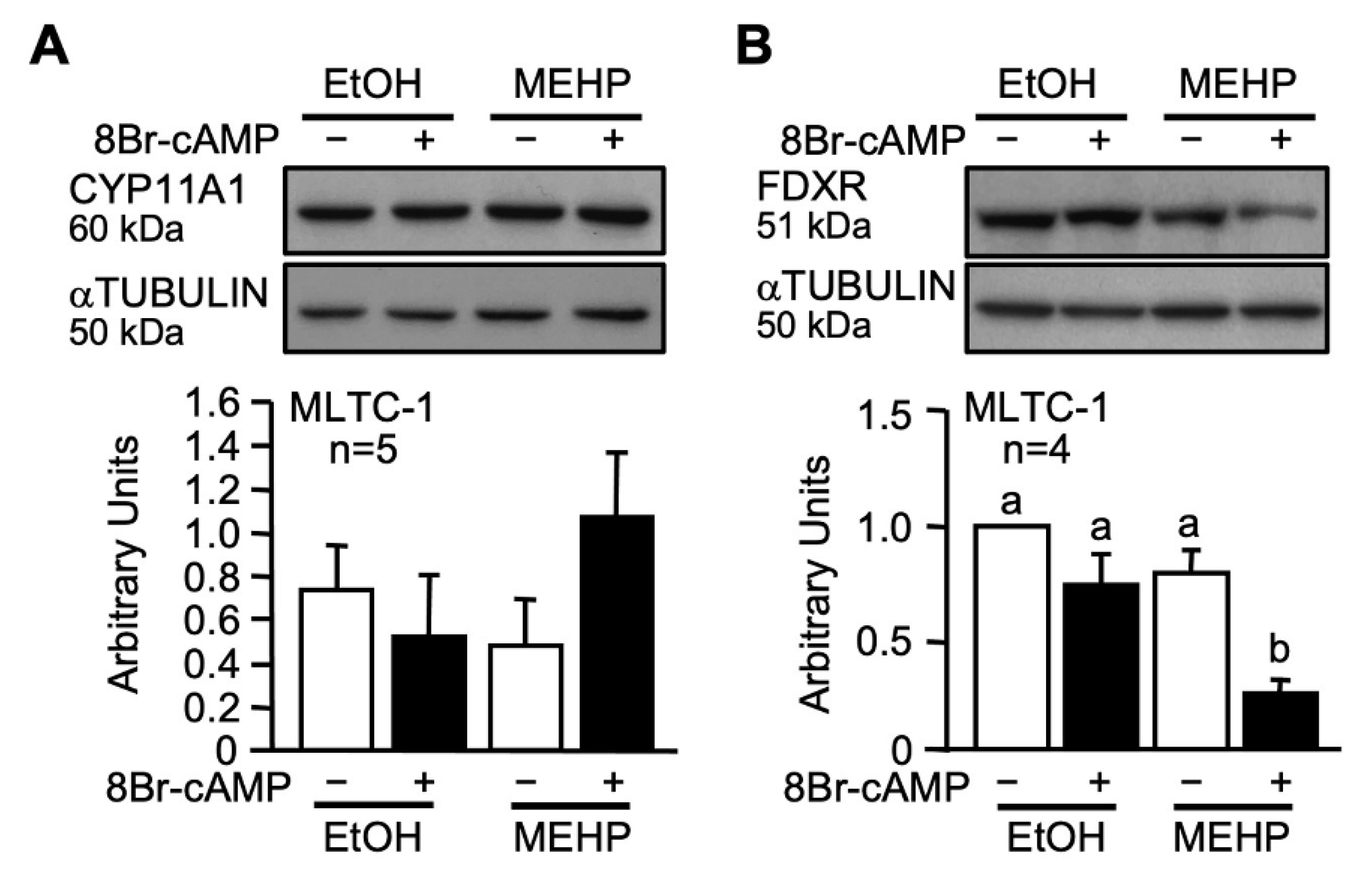
2.2. MEHP Affects FDXR Expression
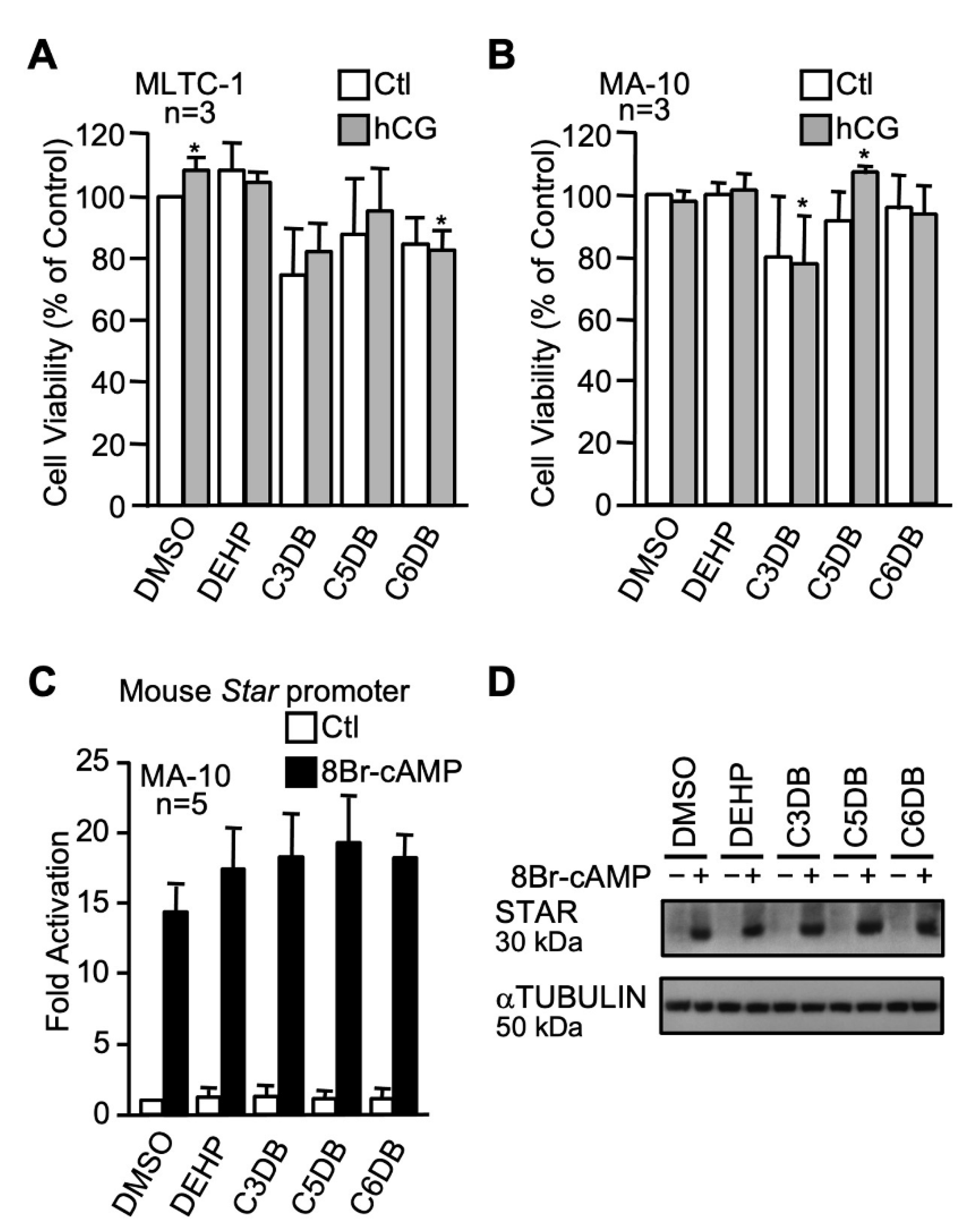
2.3. Effects of Novel Green Plasticizers on Leydig Cell Function
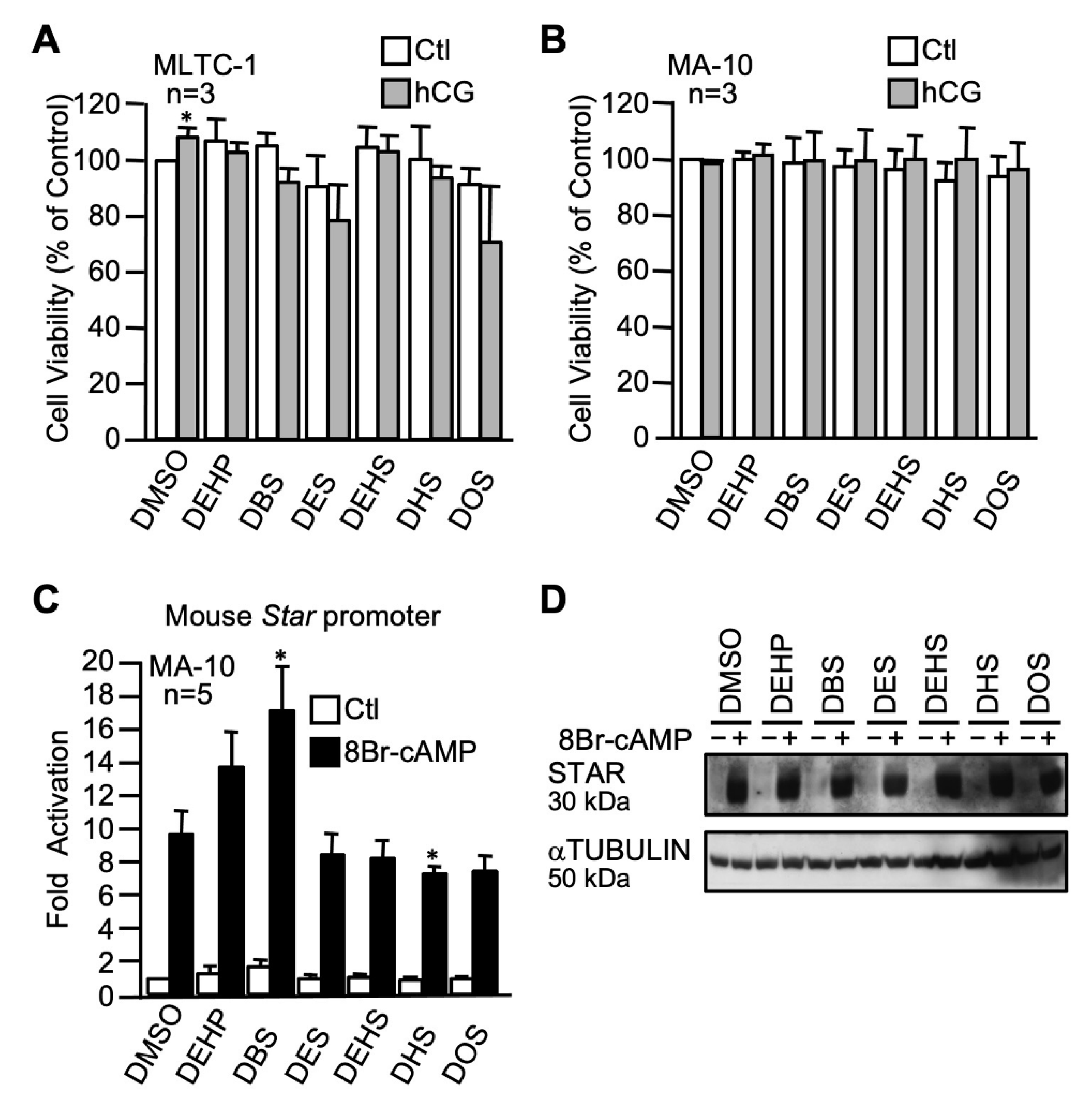
2.3.1. The n-alkyl Dibenzoate Plasticizer Series
2.3.2. The n-alkyl Succinate Plasticizer Series
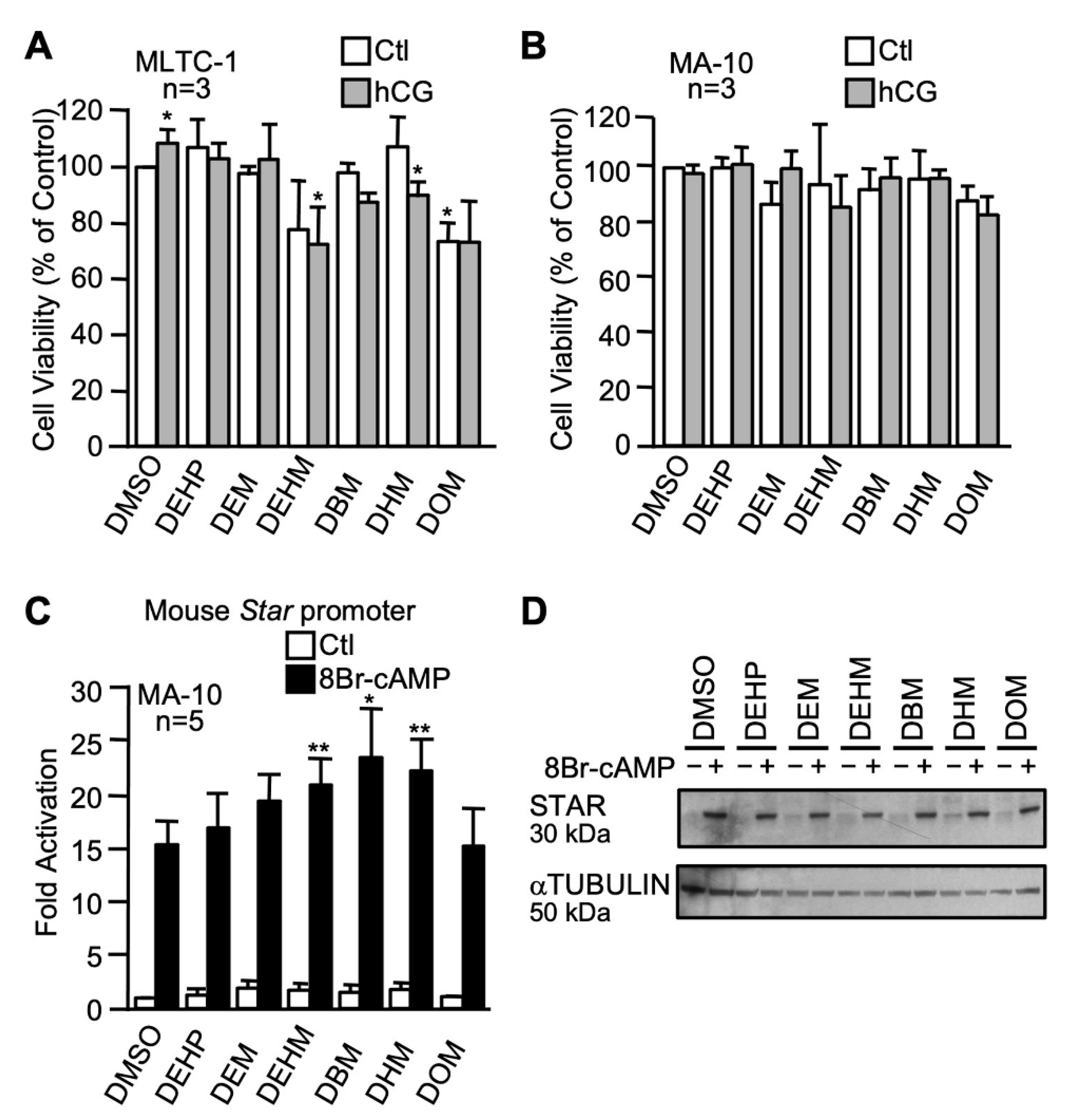
2.3.3. The n-alkyl Maleate Plasticizer Series
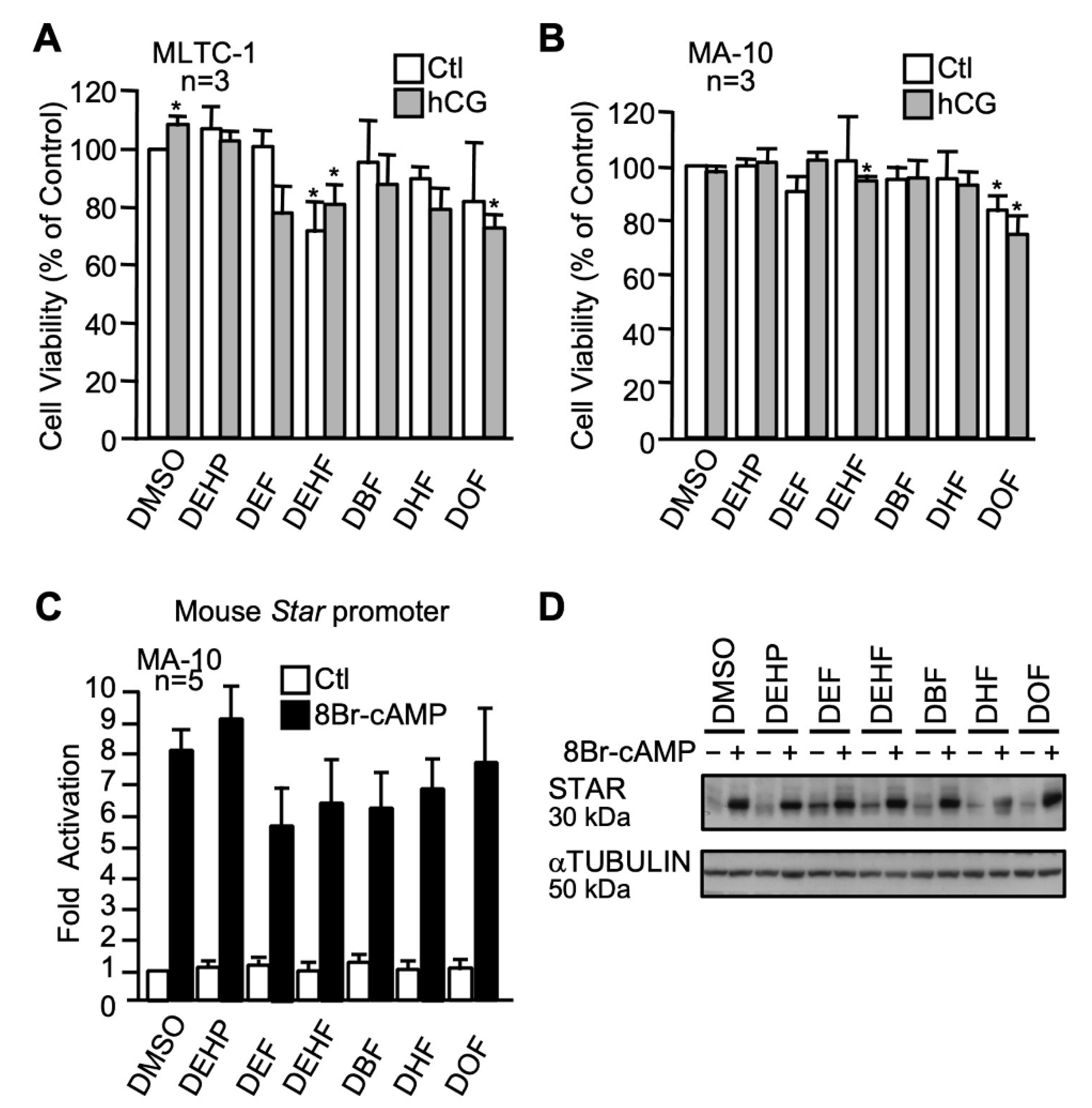
2.3.4. The n-alkyl Fumarate Plasticizer Series
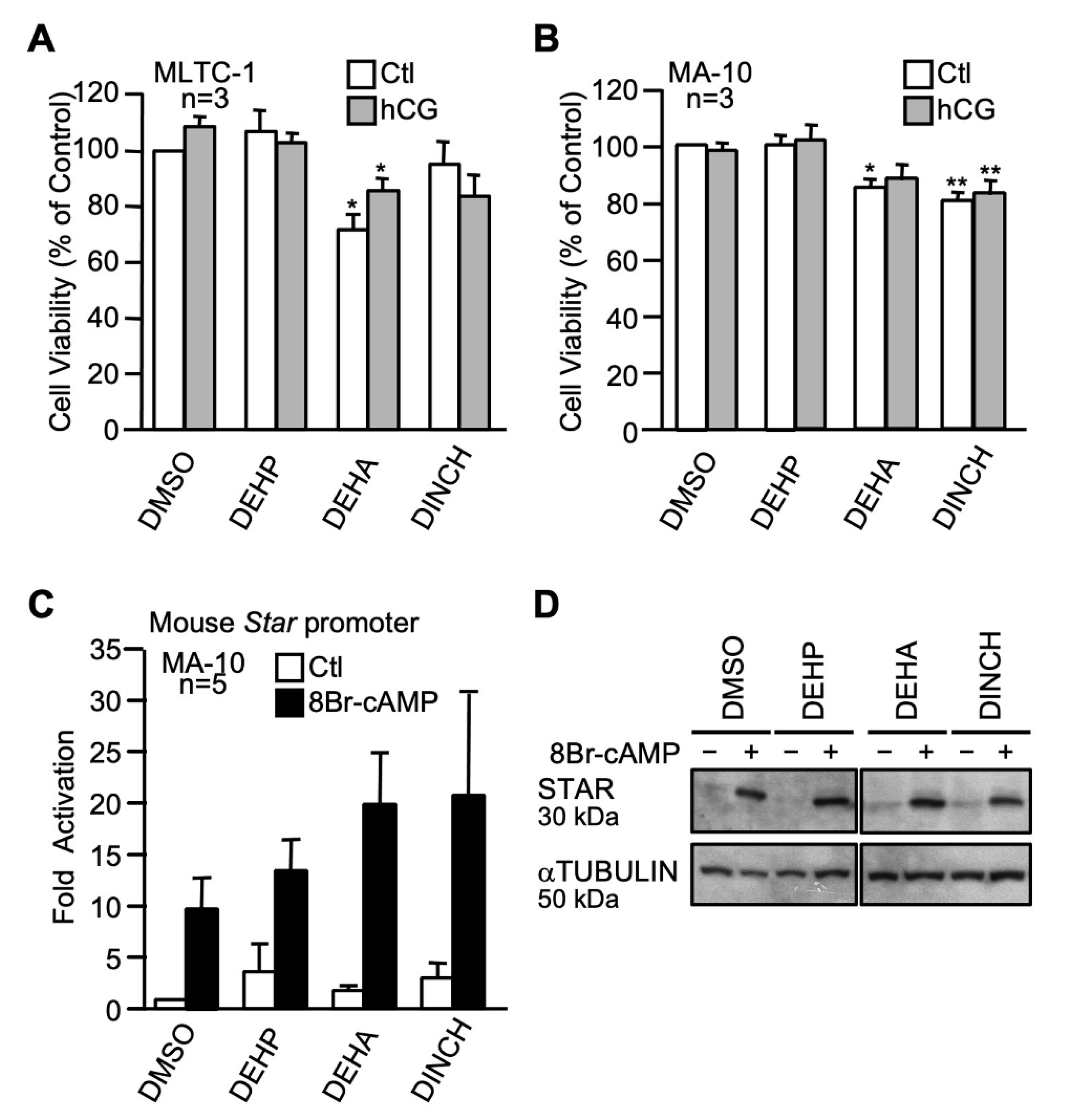
2.3.5. The Commercial Plasticizers Hexamoll-DINCH and DEHA
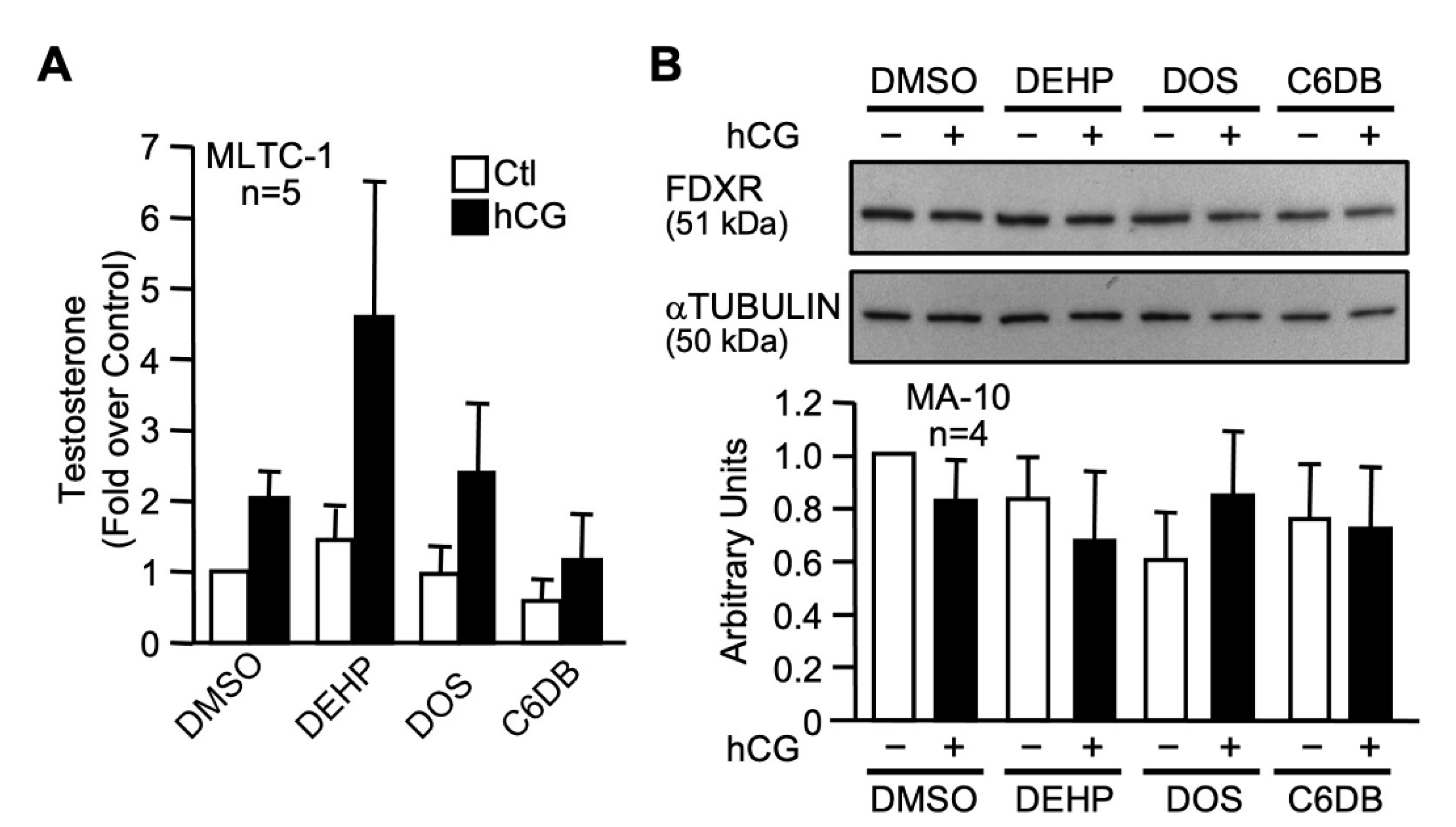
2.4. Effect of Two Novel Candidate Plasticizers on Leydig Cell Line Function and FDXR Levels
3. Discussion
3.1. MEHP Represses Leydig Cells Steroidogenesis by Decreasing Star Gene Transcription
3.2. MEHP Reduces FDXR Protein Levels in Leydig Cells
3.3. Effects of Novel Candidate Plasticizers on Leydig Cell Steroidogenesis
4. Materials and Methods
4.1. Chemicals
4.2. Cell Culture
4.3. Plasmids, Transfections and Luciferase Reporter Assays
4.4. Protein Purification and Western Blot
4.5. Progesterone and Testosterone Quantification
4.6. MTT Cell Assay
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schettler, T. Human exposure to phthalates via consumer products. Int. J. Androl. 2006, 29, 134–139. [Google Scholar] [CrossRef]
- Wormuth, M.; Scheringer, M.; Vollenweider, M.; Hungerbuhler, K. What are the sources of exposure to eight frequently used phthalic acid esters in Europeans? Risk Anal. 2006, 26, 803–824. [Google Scholar] [CrossRef]
- Fierens, T.; Servaes, K.; Van Holderbeke, M.; Geerts, L.; De Henauw, S.; Sioen, I.; Vanermen, G. Analysis of phthalates in food products and packaging materials sold on the Belgian market. Food Chem. Toxicol. 2012, 50, 2575–2583. [Google Scholar] [CrossRef]
- Guart, A.; Bono-Blay, F.; Borrell, A.; Lacorte, S. Migration of plasticizers phthalates, bisphenol A and alkylphenols from plastic containers and evaluation of risk. Food Addit. Contam. Part. Chem. Anal. Control. Expo. Risk Assess. 2011, 28, 676–685. [Google Scholar] [CrossRef]
- Koniecki, D.; Wang, R.; Moody, R.P.; Zhu, J. Phthalates in cosmetic and personal care products: Concentrations and possible dermal exposure. Environ. Res. 2011, 111, 329–336. [Google Scholar] [CrossRef]
- Guo, Y.; Kannan, K. A survey of phthalates and parabens in personal care products from the United States and its implications for human exposure. Environ. Sci. Technol. 2013, 47, 14442–14449. [Google Scholar] [CrossRef]
- Babich, M.A.; Chen, S.B.; Greene, M.A.; Kiss, C.T.; Porter, W.K.; Smith, T.P.; Wind, M.L.; Zamula, W.W. Risk assessment of oral exposure to diisononyl phthalate from children’s products. Regul. Toxicol. Pharmacol. 2004, 40, 151–167. [Google Scholar] [CrossRef]
- Wilkinson, C.F.; Lamb, J.C.T. The potential health effects of phthalate esters in children’s toys: A review and risk assessment. Regul. Toxicol. Pharmacol. 1999, 30, 140–155. [Google Scholar] [CrossRef]
- Witorsch, R.J.; Thomas, J.A. Personal care products and endocrine disruption: A critical review of the literature. Crit. Rev. Toxicol. 2010, 40, 1–30. [Google Scholar] [CrossRef]
- Net, S.; Sempere, R.; Delmont, A.; Paluselli, A.; Ouddane, B. Occurrence, fate, behavior and ecotoxicological state of phthalates in different environmental matrices. Environ. Sci. Technol. 2015, 49, 4019–4035. [Google Scholar] [CrossRef]
- Luis, C.; Algarra, M.; Camara, J.S.; Perestrelo, R. Comprehensive Insight from Phthalates Occurrence: From Health Outcomes to Emerging Analytical Approaches. Toxics 2021, 9, 157. [Google Scholar] [CrossRef]
- Wang, Y.; Qian, H. Phthalates and Their Impacts on Human Health. Healthcare 2021, 9, 603. [Google Scholar] [CrossRef]
- Strauss, A.A. Di(2-ethylhexyl)phthalate (DEHP). J. Pediatr. Pharmacol. Ther. 2004, 9, 89–95. [Google Scholar] [CrossRef] [Green Version]
- Erythropel, H.C.; Maric, M.; Nicell, J.A.; Leask, R.L.; Yargeau, V. Leaching of the plasticizer di(2-ethylhexyl)phthalate (DEHP) from plastic containers and the question of human exposure. Appl. Microbiol. Biotechnol. 2014, 98, 9967–9981. [Google Scholar] [CrossRef]
- Wang, Y.; Zhu, H.; Kannan, K. A Review of Biomonitoring of Phthalate Exposures. Toxics 2019, 7, 21. [Google Scholar] [CrossRef] [Green Version]
- Toxicology Excellence for Risk Assessment (TERA). Exposure Assessment: Composition, Production, and Use of Phthalates; University of Cincinnati: Cincinnati, OH, USA, 2015; pp. 1–180. [Google Scholar]
- Huang, S.; Qi, Z.; Ma, S.; Li, G.; Long, C.; Yu, Y. A critical review on human internal exposure of phthalate metabolites and the associated health risks. Environ. Pollut. 2021, 279, 116941. [Google Scholar] [CrossRef]
- Institute for Health and Consumer Protection. European Union Risk Assessment Report on Bis(2-ethylhexyl) Phthalate (DEHP); Office for Official Publications of the European Communities: Luxembourg, 2008; pp. 1–588. [Google Scholar]
- Wittassek, M.; Koch, H.M.; Angerer, J.; Bruning, T. Assessing exposure to phthalates—The human biomonitoring approach. Mol. Nutr. Food Res. 2011, 55, 7–31. [Google Scholar] [CrossRef]
- Heudorf, U.; Mersch-Sundermann, V.; Angerer, J. Phthalates: Toxicology and exposure. Int. J. Hyg. Environ. Health 2007, 210, 623–634. [Google Scholar] [CrossRef]
- Koch, H.M.; Drexler, H.; Angerer, J. An estimation of the daily intake of di(2-ethylhexyl)phthalate (DEHP) and other phthalates in the general population. Int. J. Hyg. Environ. Health 2003, 206, 77–83. [Google Scholar] [CrossRef]
- Koch, H.M.; Preuss, R.; Angerer, J. Di(2-ethylhexyl)phthalate (DEHP): Human metabolism and internal exposure—An update and latest results. Int. J. Androl. 2006, 29, 155–165. [Google Scholar] [CrossRef]
- Frederiksen, H.; Skakkebaek, N.E.; Andersson, A.M. Metabolism of phthalates in humans. Mol. Nutr. Food Res. 2007, 51, 899–911. [Google Scholar] [CrossRef]
- Silva, M.J.; Barr, D.B.; Reidy, J.A.; Kato, K.; Malek, N.A.; Hodge, C.C.; Hurtz, D., 3rd; Calafat, A.M.; Needham, L.L.; Brock, J.W. Glucuronidation patterns of common urinary and serum monoester phthalate metabolites. Arch. Toxicol. 2003, 77, 561–567. [Google Scholar] [CrossRef]
- Vo, T.T.; Jung, E.M.; Dang, V.H.; Jung, K.; Baek, J.; Choi, K.C.; Jeung, E.B. Differential effects of flutamide and di-(2-ethylhexyl) phthalate on male reproductive organs in a rat model. J. Reprod. Dev. 2009, 55, 400–411. [Google Scholar] [CrossRef] [Green Version]
- Vo, T.T.; Jung, E.M.; Dang, V.H.; Yoo, Y.M.; Choi, K.C.; Yu, F.H.; Jeung, E.B. Di-(2 ethylhexyl) phthalate and flutamide alter gene expression in the testis of immature male rats. Reprod. Biol. Endocrinol. 2009, 7, 104. [Google Scholar] [CrossRef] [Green Version]
- Andrade, A.J.; Grande, S.W.; Talsness, C.E.; Gericke, C.; Grote, K.; Golombiewski, A.; Sterner-Kock, A.; Chahoud, I. A dose response study following in utero and lactational exposure to di-(2-ethylhexyl) phthalate (DEHP): Reproductive effects on adult male offspring rats. Toxicology 2006, 228, 85–97. [Google Scholar] [CrossRef]
- Venturelli, A.C.; Meyer, K.B.; Fischer, S.V.; Kita, D.H.; Philipsen, R.A.; Morais, R.N.; Martino Andrade, A.J. Effects of in utero and lactational exposure to phthalates on reproductive development and glycemic homeostasis in rats. Toxicology 2019, 421, 30–40. [Google Scholar] [CrossRef]
- Li, H.; Spade, D.J. Environmental exposures, fetal testis development and function: Phthalates and beyond. Reproduction 2021, 162, F147–F167. [Google Scholar] [CrossRef]
- Foster, P.M. Disruption of reproductive development in male rat offspring following in utero exposure to phthalate esters. Int. J. Androl. 2006, 29, 140–147. [Google Scholar] [CrossRef]
- Gray, L.E., Jr.; Ostby, J.; Furr, J.; Price, M.; Veeramachaneni, D.N.; Parks, L. Perinatal exposure to the phthalates DEHP, BBP, and DINP, but not DEP, DMP, or DOTP, alters sexual differentiation of the male rat. Toxicol. Sci. 2000, 58, 350–365. [Google Scholar] [CrossRef] [Green Version]
- Parks, L.G.; Ostby, J.S.; Lambright, C.R.; Abbott, B.D.; Klinefelter, G.R.; Barlow, N.J.; Gray, L.E., Jr. The plasticizer diethylhexyl phthalate induces malformations by decreasing fetal testosterone synthesis during sexual differentiation in the male rat. Toxicol. Sci. 2000, 58, 339–349. [Google Scholar] [CrossRef]
- Howdeshell, K.L.; Rider, C.V.; Wilson, V.S.; Gray, L.E., Jr. Mechanisms of action of phthalate esters, individually and in combination, to induce abnormal reproductive development in male laboratory rats. Environ. Res. 2008, 108, 168–176. [Google Scholar] [CrossRef] [Green Version]
- Christiansen, S.; Boberg, J.; Axelstad, M.; Dalgaard, M.; Vinggaard, A.M.; Metzdorff, S.B.; Hass, U. Low-dose perinatal exposure to di(2-ethylhexyl) phthalate induces anti-androgenic effects in male rats. Reprod. Toxicol. 2010, 30, 313–321. [Google Scholar] [CrossRef]
- Culty, M.; Thuillier, R.; Li, W.; Wang, Y.; Martinez-Arguelles, D.B.; Benjamin, C.G.; Triantafilou, K.M.; Zirkin, B.R.; Papadopoulos, V. In utero exposure to di-(2-ethylhexyl) phthalate exerts both short-term and long-lasting suppressive effects on testosterone production in the rat. Biol. Reprod. 2008, 78, 1018–1028. [Google Scholar] [CrossRef] [Green Version]
- Desdoits-Lethimonier, C.; Albert, O.; Le Bizec, B.; Perdu, E.; Zalko, D.; Courant, F.; Lesne, L.; Guille, F.; Dejucq-Rainsford, N.; Jegou, B. Human testis steroidogenesis is inhibited by phthalates. Hum. Reprod. 2012, 27, 1451–1459. [Google Scholar] [CrossRef] [Green Version]
- Chang, W.H.; Li, S.S.; Wu, M.H.; Pan, H.A.; Lee, C.C. Phthalates might interfere with testicular function by reducing testosterone and insulin-like factor 3 levels. Hum. Reprod. 2015, 30, 2658–2670. [Google Scholar] [CrossRef] [Green Version]
- Araki, A.; Mitsui, T.; Miyashita, C.; Nakajima, T.; Naito, H.; Ito, S.; Sasaki, S.; Cho, K.; Ikeno, T.; Nonomura, K.; et al. Association between maternal exposure to di(2-ethylhexyl) phthalate and reproductive hormone levels in fetal blood: The Hokkaido study on environment and children’s health. PLoS ONE 2014, 9, e109039. [Google Scholar] [CrossRef] [Green Version]
- Chauvigne, F.; Plummer, S.; Lesne, L.; Cravedi, J.P.; Dejucq-Rainsford, N.; Fostier, A.; Jegou, B. Mono-(2-ethylhexyl) phthalate directly alters the expression of Leydig cell genes and CYP17 lyase activity in cultured rat fetal testis. PLoS ONE 2011, 6, e27172. [Google Scholar] [CrossRef] [Green Version]
- Chauvigne, F.; Menuet, A.; Lesne, L.; Chagnon, M.C.; Chevrier, C.; Regnier, J.F.; Angerer, J.; Jegou, B. Time- and dose-related effects of di-(2-ethylhexyl) phthalate and its main metabolites on the function of the rat fetal testis in vitro. Environ. Health Perspect. 2009, 117, 515–521. [Google Scholar] [CrossRef] [Green Version]
- Svechnikov, K.; Svechnikova, I.; Soder, O. Inhibitory effects of mono-ethylhexyl phthalate on steroidogenesis in immature and adult rat Leydig cells in vitro. Reprod. Toxicol. 2008, 25, 485–490. [Google Scholar] [CrossRef]
- Sekaran, S.; Jagadeesan, A. In utero exposure to phthalate downregulates critical genes in Leydig cells of F1 male progeny. J. Cell Biochem. 2015, 116, 1466–1477. [Google Scholar] [CrossRef]
- Miller, W.L.; Auchus, R.J. The molecular biology, biochemistry, and physiology of human steroidogenesis and its disorders. Endocr. Rev. 2011, 32, 81–151. [Google Scholar] [CrossRef] [Green Version]
- Martin, L.J.; Boucher, N.; Brousseau, C.; Tremblay, J.J. The orphan nuclear receptor NUR77 regulates hormone-induced StAR transcription in Leydig cells through a cooperation with CaMKI. Mol. Endocrinol. 2008, 22, 2021–2037. [Google Scholar] [CrossRef] [Green Version]
- Stocco, D.M.; Wang, X.; Jo, Y.; Manna, P.R. Multiple signaling pathways regulating steroidogenesis and steroidogenic acute regulatory protein expression: More complicated than we thought. Mol. Endocrinol. 2005, 19, 2647–2659. [Google Scholar] [CrossRef] [Green Version]
- Tremblay, J.J. Molecular regulation of steroidogenesis in endocrine Leydig cells. Steroids 2015, 103, 3–10. [Google Scholar] [CrossRef]
- Stocco, D.M. The role of the StAR protein in steroidogenesis: Challenges for the future. J. Endocrinol. 2000, 164, 247–253. [Google Scholar] [CrossRef]
- Issop, L.; Rone, M.B.; Papadopoulos, V. Organelle plasticity and interactions in cholesterol transport and steroid biosynthesis. Mol. Cell Endocrinol. 2013, 371, 34–46. [Google Scholar] [CrossRef]
- Manna, P.R.; Stetson, C.L.; Slominski, A.T.; Pruitt, K. Role of the steroidogenic acute regulatory protein in health and disease. Endocrine 2016, 51, 7–21. [Google Scholar] [CrossRef] [Green Version]
- Aghazadeh, Y.; Zirkin, B.R.; Papadopoulos, V. Pharmacological regulation of the cholesterol transport machinery in steroidogenic cells of the testis. Vitam. Horm. 2015, 98, 189–227. [Google Scholar]
- Piché, C.D.; Sauvageau, D.; Vanlian, M.; Erythropel, H.C.; Robaire, B.; Leask, R.L. Effects of di-(2-ethylhexyl) phthalate and four of its metabolites on steroidogenesis in MA-10 cells. Ecotoxicol. Environ. Saf. 2012, 79, 108–115. [Google Scholar] [CrossRef]
- Fan, J.; Traore, K.; Li, W.; Amri, H.; Huang, H.; Wu, C.; Chen, H.; Zirkin, B.; Papadopoulos, V. Molecular mechanisms mediating the effect of mono-(2-ethylhexyl) phthalate on hormone-stimulated steroidogenesis in MA-10 mouse tumor Leydig cells. Endocrinology 2010, 151, 3348–3362. [Google Scholar] [CrossRef] [Green Version]
- Bui, T.T.; Giovanoulis, G.; Cousins, A.P.; Magner, J.; Cousins, I.T.; de Wit, C.A. Human exposure, hazard and risk of alternative plasticizers to phthalate esters. Sci. Total. Environ. 2016, 541, 451–467. [Google Scholar] [CrossRef]
- Svechnikova, K.; Svechnikova, I.; Soder, O. Gender-Specific Adverse Effects of Mono-Ethylhexyl Phthalate on Steroidogenesis in Immature Granulosa Cells and Rat Leydig cell Progenitors in vitro. Front. Endocrinol. 2011, 2, 9. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Beattie, M.C.; Lin, C.Y.; Liu, J.; Traore, K.; Papadopoulos, V.; Zirkin, B.R.; Chen, H. Oxidative stress and phthalate-induced down-regulation of steroidogenesis in MA-10 Leydig cells. Reprod. Toxicol. 2013, 42, 95–101. [Google Scholar] [CrossRef] [Green Version]
- Clark, B.J.; Cochrum, R.K. The steroidogenic acute regulatory protein as a target of endocrine disruption in male reproduction. Drug. Metab. Rev. 2007, 39, 353–370. [Google Scholar] [CrossRef]
- Erythropel, H.C.; Dodd, P.; Leask, R.L.; Maric, M.; Cooper, D.G. Designing green plasticizers: Influence of alkyl chain length on biodegradation and plasticization properties of succinate based plasticizers. Chemosphere 2013, 91, 358–365. [Google Scholar] [CrossRef]
- Erythropel, H.C.; Maric, M.; Cooper, D.G. Designing green plasticizers: Influence of molecular geometry on biodegradation and plasticization properties. Chemosphere 2012, 86, 759–766. [Google Scholar] [CrossRef]
- Erythropel, H.C.; Brown, T.; Maric, M.; Nicell, J.A.; Cooper, D.G.; Leask, R.L. Designing greener plasticizers: Effects of alkyl chain length and branching on the biodegradation of maleate based plasticizers. Chemosphere 2015, 134, 106–112. [Google Scholar] [CrossRef]
- Erythropel, H.C.; Shipley, S.; Börmann, A.; Nicell, J.A.; Maric, L.; Leask, R.L. Designing green plasticizers: Influence of molecule geometry and alkyl chain length on the plasticizing effectiveness of diester plasticizers in PVC blends. Polymer 2016, 89, 18–27. [Google Scholar] [CrossRef]
- Ascoli, M. Immortalized Leydig Cell Lines as Models for Studying Leydig Cell Physiology. In The Leydig Cell in Health and Disease, 2nd ed.; Payne, A., Hardy, M.P., Eds.; Humana Press: Totowa, NJ, USA, 2007; pp. 373–381. [Google Scholar]
- Gunnarsson, D.; Leffler, P.; Ekwurtzel, E.; Martinsson, G.; Liu, K.; Selstam, G. Mono-(2-ethylhexyl) phthalate stimulates basal steroidogenesis by a cAMP-independent mechanism in mouse gonadal cells of both sexes. Reproduction 2008, 135, 693–703. [Google Scholar] [CrossRef] [Green Version]
- Savchuk, I.; Soder, O.; Svechnikov, K. Mono-2-ethylhexyl phthalate stimulates androgen production but suppresses mitochondrial function in mouse leydig cells with different steroidogenic potential. Toxicol. Sci. 2015, 145, 149–156. [Google Scholar] [CrossRef] [Green Version]
- Dees, J.H.; Gazouli, M.; Papadopoulos, V. Effect of mono-ethylhexyl phthalate on MA-10 Leydig tumor cells. Reprod. Toxicol. 2001, 15, 171–187. [Google Scholar] [CrossRef]
- Mendoza-Villarroel, R.E.; Robert, N.M.; Martin, L.J.; Brousseau, C.; Tremblay, J.J. The nuclear receptor NR2F2 activates Star expression and steroidogenesis in mouse MA-10 and MLTC-1 Leydig cells. Biol. Reprod. 2014, 91, 26. [Google Scholar] [CrossRef] [Green Version]
- Martin, L.J.; Bergeron, F.; Viger, R.S.; Tremblay, J.J. Functional cooperation between GATA factors and cJUN on the star promoter in MA-10 Leydig cells. J. Androl. 2012, 33, 81–87. [Google Scholar] [CrossRef]
- Martin, L.J.; Tremblay, J.J. The nuclear receptors NUR77 and SF1 play additive roles with c-JUN through distinct elements on the mouse Star promoter. J. Mol. Endocrinol. 2009, 42, 119–129. [Google Scholar] [CrossRef] [Green Version]
- Manna, P.R.; Huhtaniemi, I.T.; Stocco, D.M. Mechanisms of protein kinase C signaling in the modulation of 3’,5’-cyclic adenosine monophosphate-mediated steroidogenesis in mouse gonadal cells. Endocrinology 2009, 150, 3308–3317. [Google Scholar] [CrossRef] [Green Version]
- Manna, P.R.; Stocco, D.M. Crosstalk of CREB and Fos/Jun on a single cis-element: Transcriptional repression of the steroidogenic acute regulatory protein gene. J. Mol. Endocrinol. 2007, 39, 261–277. [Google Scholar] [CrossRef]
- Manna, P.R.; Eubank, D.W.; Stocco, D.M. Assessment of the role of activator protein-1 on transcription of the mouse steroidogenic acute regulatory protein gene. Mol. Endocrinol. 2004, 18, 558–573. [Google Scholar] [CrossRef]
- Sugawara, T.; Saito, M.; Fujimoto, S. Sp1 and SF-1 interact and cooperate in the regulation of human steroidogenic acute regulatory protein gene expression. Endocrinology 2000, 141, 2895–2903. [Google Scholar] [CrossRef]
- Silverman, E.; Eimerl, S.; Orly, J. CCAAT enhancer-binding protein beta and GATA-4 binding regions within the promoter of the steroidogenic acute regulatory protein (StAR) gene are required for transcription in rat ovarian cells. J. Biol. Chem. 1999, 274, 17987–17996. [Google Scholar] [CrossRef] [Green Version]
- Christenson, L.K.; Johnson, P.F.; McAllister, J.M.; Strauss, J.F., III. CCAAT/enhancer-binding proteins regulate expression of the human steroidogenic acute regulatory protein (StAR) gene. J. Biol. Chem. 1999, 274, 26591–26598. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Dong, C.; Chen, M.; Chen, Y.; Gu, A.; Xia, Y.; Sun, H.; Li, Z.; Wang, Y. Effects of monobutyl phthalate on steroidogenesis through steroidogenic acute regulatory protein regulated by transcription factors in mouse Leydig tumor cells. J. Endocrinol. Invest. 2015, 38, 875–884. [Google Scholar] [CrossRef]
- Scott, H.M.; Mason, J.I.; Sharpe, R.M. Steroidogenesis in the fetal testis and its susceptibility to disruption by exogenous compounds. Endocr. Rev. 2009, 30, 883–925. [Google Scholar] [CrossRef] [Green Version]
- Payne, A.H. Steroidogenic Enzymes in Leydig Cells. In The Leydig Cell in Health and Disease, 2nd ed.; Payne, A., Hardy, M.P., Eds.; Humana Press: Totowa, NJ, USA, 2007; pp. 157–171. [Google Scholar]
- Slone, J.D.; Yang, L.; Peng, Y.; Queme, L.F.; Harris, B.; Rizzo, S.J.S.; Green, T.; Ryan, J.L.; Jankowski, M.P.; Reinholdt, L.G.; et al. Integrated analysis of the molecular pathogenesis of FDXR-associated disease. Cell Death Dis. 2020, 11, 423. [Google Scholar] [CrossRef]
- Slone, J.; Peng, Y.; Chamberlin, A.; Harris, B.; Kaylor, J.; McDonald, M.T.; Lemmon, M.; El-Dairi, M.A.; Tchapyjnikov, D.; Gonzalez-Krellwitz, L.A.; et al. Biallelic mutations in FDXR cause neurodegeneration associated with inflammation. J. Hum. Genet. 2018, 63, 1211–1222. [Google Scholar] [CrossRef]
- Peng, Y.; Shinde, D.N.; Valencia, C.A.; Mo, J.S.; Rosenfeld, J.; Truitt Cho, M.; Chamberlin, A.; Li, Z.; Liu, J.; Gui, B.; et al. Biallelic mutations in the ferredoxin reductase gene cause novel mitochondriopathy with optic atrophy. Hum. Mol. Genet. 2017, 26, 4937–4950. [Google Scholar] [CrossRef] [Green Version]
- Boisvert, A.; Jones, S.; Issop, L.; Erythropel, H.C.; Papadopoulos, V.; Culty, M. In vitro functional screening as a means to identify new plasticizers devoid of reproductive toxicity. Environ. Res. 2016, 150, 496–512. [Google Scholar] [CrossRef]
- Kermanshahi Pour, A.; Cooper, D.G.; Mamer, O.A.; Maric, M.; Nicell, J.A. Mechanisms of biodegradation of dibenzoate plasticizers. Chemosphere 2009, 77, 258–263. [Google Scholar] [CrossRef]
- Kermanshahi Pour, A.; Mamer, O.A.; Cooper, D.G.; Maric, M.; Nicell, J.A. Metabolites from the biodegradation of 1,6-hexanediol dibenzoate, a potential green plasticizer, by Rhodococcus rhodochrous. J. Mass Spectrom 2009, 44, 662–671. [Google Scholar] [CrossRef]
- Kastner, J.; Cooper, D.G.; Maric, M.; Dodd, P.; Yargeau, V. Aqueous leaching of di-2-ethylhexyl phthalate and “green” plasticizers from poly(vinyl chloride). Sci. Total Environ. 2012, 432, 357–364. [Google Scholar] [CrossRef] [Green Version]
- Albert, O.; Nardelli, T.C.; Lalancette, C.; Hales, B.F.; Robaire, B. Effects of In Utero and Lactational Exposure to New Generation Green Plasticizers on Adult Male Rats: A Comparative Study With Di(2-Ethylhexyl) Phthalate. Toxicol. Sci. 2018, 164, 129–141. [Google Scholar] [CrossRef]
- Nardelli, T.C.; Albert, O.; Lalancette, C.; Culty, M.; Hales, B.F.; Robaire, B. In Utero and Lactational Exposure Study in Rats to Identify Replacements for Di(2-ethylhexyl) Phthalate. Sci. Rep. 2017, 7, 3862. [Google Scholar] [CrossRef] [Green Version]
- Ascoli, M. Characterization of several clonal lines of cultured Leydig tumor cells: Gonadotropin receptors and steroidogenic responses. Endocrinology 1981, 108, 88–95. [Google Scholar] [CrossRef]
- Martin, L.J.; Boucher, N.; El-Asmar, B.; Tremblay, J.J. cAMP-induced expression of the orphan nuclear receptor Nur77 in testicular Leydig cells involves a CaMKI pathway. J. Androl. 2009, 30, 134–145. [Google Scholar] [CrossRef]
- Abdou, H.S.; Villeneuve, G.; Tremblay, J.J. The calcium signaling pathway regulates leydig cell steroidogenesis through a transcriptional cascade involving the nuclear receptor NR4A1 and the steroidogenic acute regulatory protein. Endocrinology 2013, 154, 511–520. [Google Scholar] [CrossRef] [Green Version]
- Abdou, H.S.; Bergeron, F.; Tremblay, J.J. A cell-autonomous molecular cascade initiated by AMP-activated protein kinase represses steroidogenesis. Mol. Cell Biol. 2014, 34, 4257–4271. [Google Scholar] [CrossRef] [Green Version]
- Enangue Njembele, A.N.; Bailey, J.L.; Tremblay, J.J. In vitro exposure of Leydig cells to an environmentally relevant mixture of organochlorines represses early steps of steroidogenesis. Biol. Reprod. 2014, 90, 118. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Enangue Njembele, A.N.; Tremblay, J.J. Mechanisms of MEHP Inhibitory Action and Analysis of Potential Replacement Plasticizers on Leydig Cell Steroidogenesis. Int. J. Mol. Sci. 2021, 22, 11456. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111456
Enangue Njembele AN, Tremblay JJ. Mechanisms of MEHP Inhibitory Action and Analysis of Potential Replacement Plasticizers on Leydig Cell Steroidogenesis. International Journal of Molecular Sciences. 2021; 22(21):11456. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111456
Chicago/Turabian StyleEnangue Njembele, Annick N., and Jacques J. Tremblay. 2021. "Mechanisms of MEHP Inhibitory Action and Analysis of Potential Replacement Plasticizers on Leydig Cell Steroidogenesis" International Journal of Molecular Sciences 22, no. 21: 11456. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222111456