
New Investigations with Lupane Type A-Ring Azepane Triterpenoids for Antimycobacterial Drug Candidate Design

 ,
, .jpg) , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results and Discussion
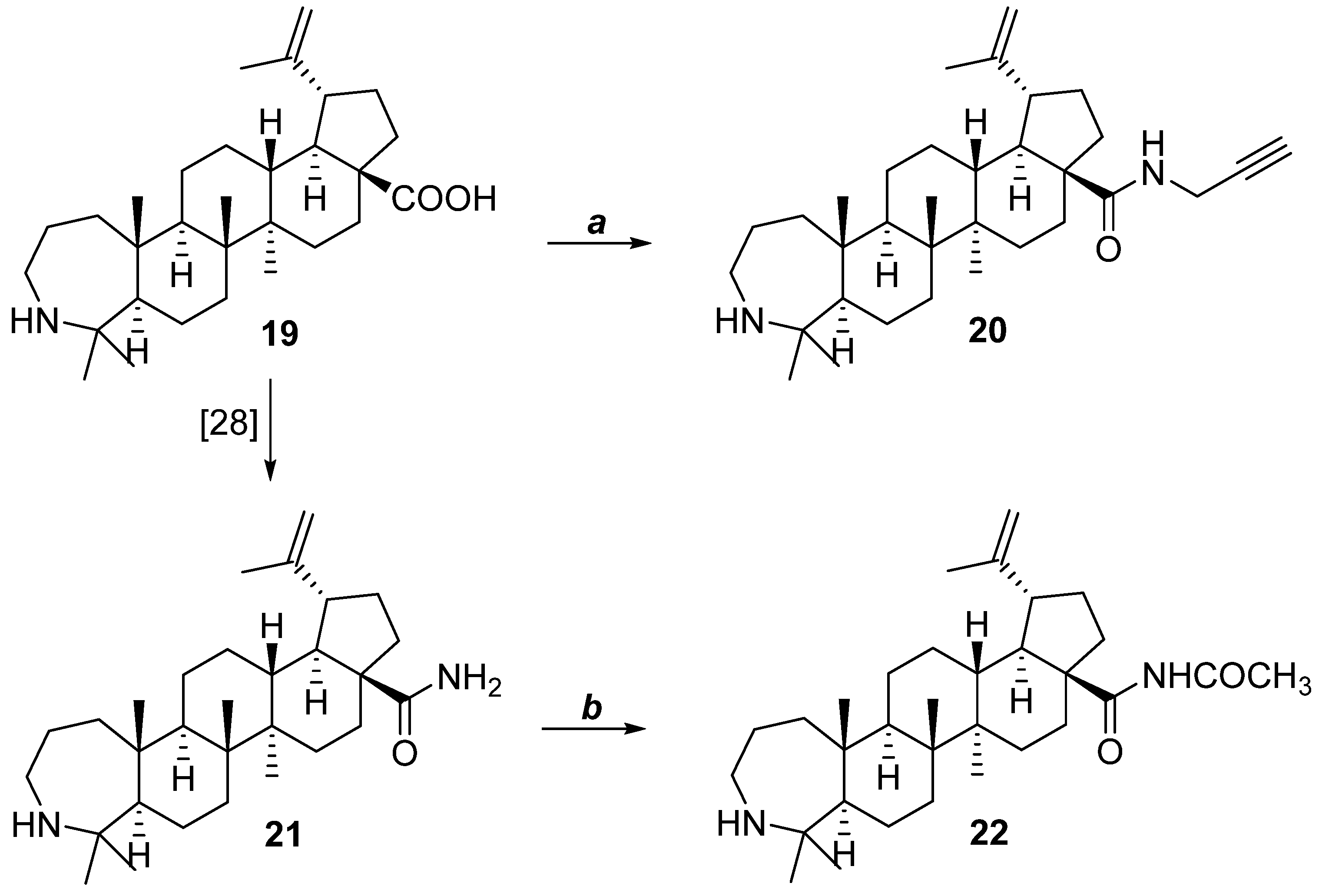
2.1. Chemistry
2.2. Machine Learning Antitubercular Activity and Compound-Drug Similarity Prediction
2.3. MIC Determination against M. Tuberculosis and NTM Strains
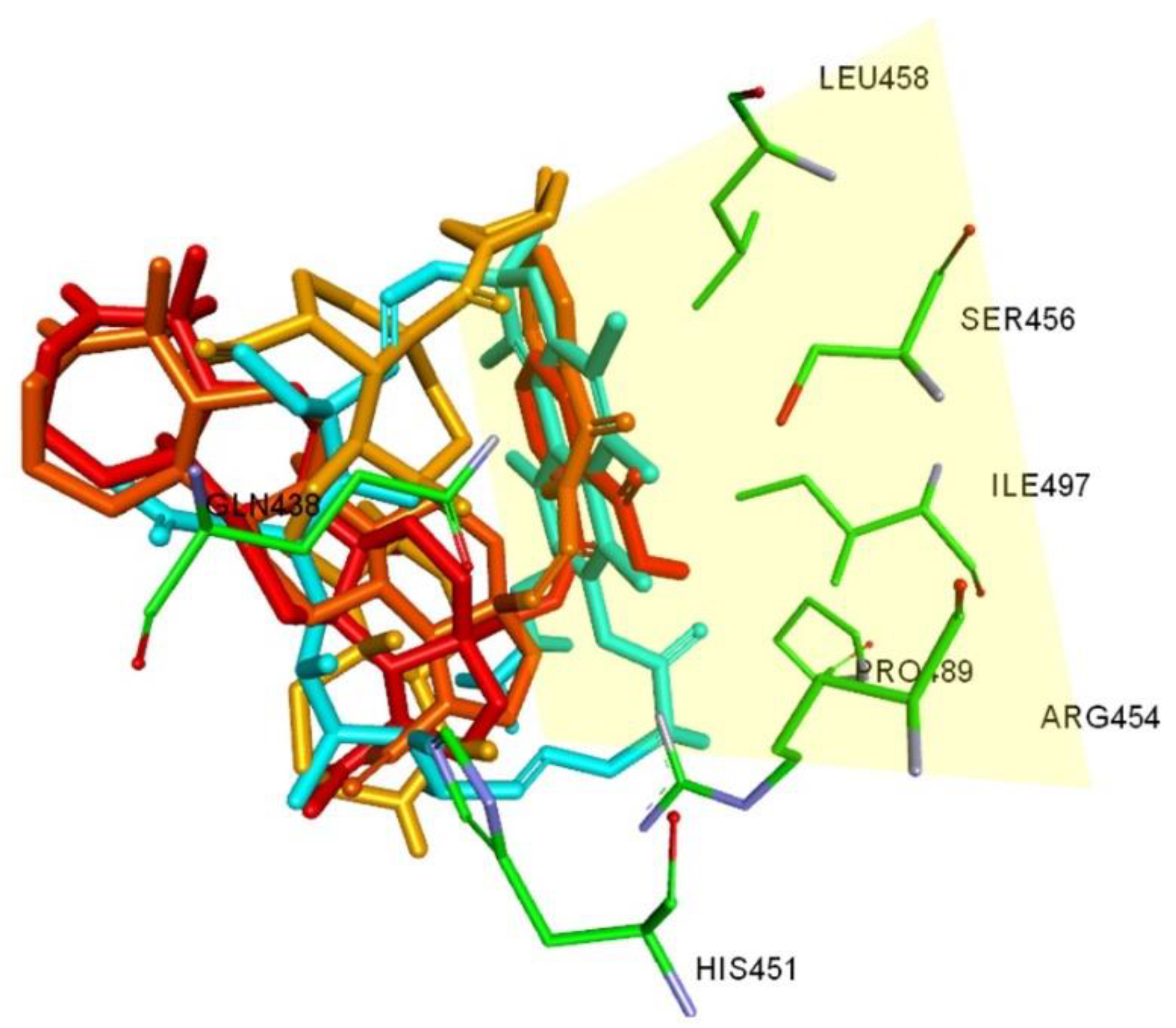
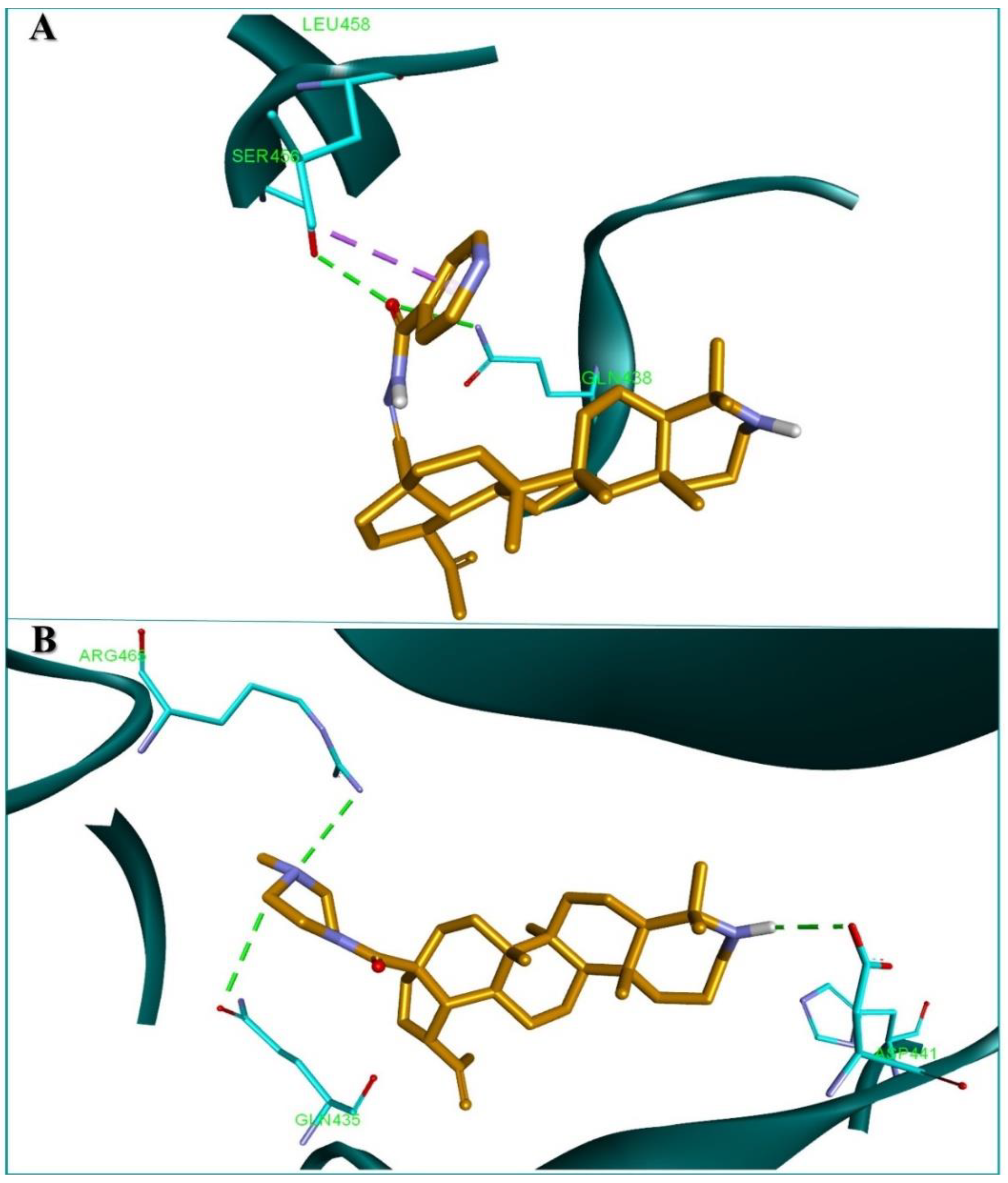
2.4. Molecular Docking
2.5. Ex Vivo Efficacy and Cytotoxicity
2.6. Combinatorial Testing with Standard of Care Drugs against M. tuberculosis H37Rv
2.7. Frequency of Resistance
3. Conclusions
4. Experimental Part
4.1. Chemistry
4.2. Computational Prediction of Tuberculostatic Compound Activity and Mechanism of Action
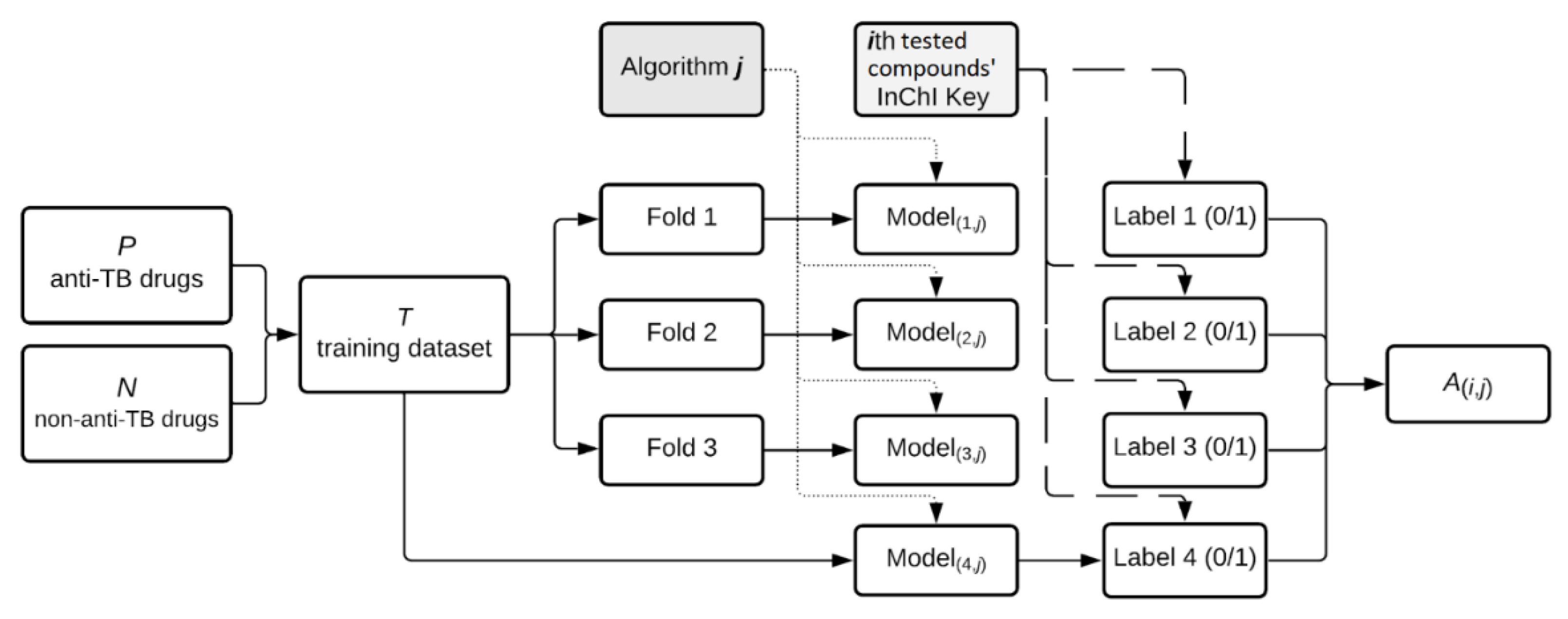
4.2.1. Machine Learning Based Antitubercular Activity Prediction
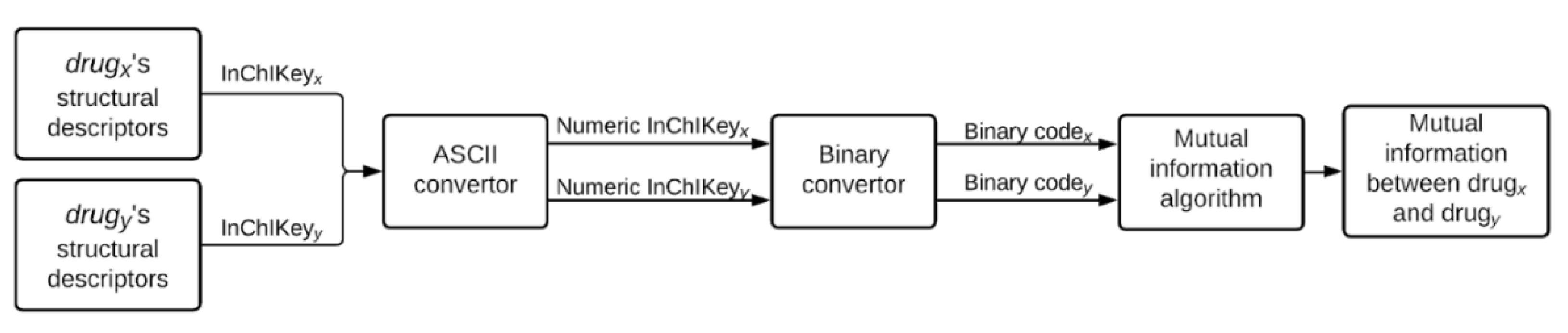
4.2.2. Mutual Information-Based Compound-Drug Similarity Prediction
4.2.3. Molecular Docking Based Compound Mechanism of Action Assessment
4.3. Compound Sample Preparation for In Vitro Biological Assessment
4.4. MIC Determination against M. tuberculosis Strains
4.5. MIC Determination against NTM Strains
4.6. Ex Vivo Testing
4.6.1. In Vitro Infection of Differentiated THP-1 Human Macrophages
4.6.2. Cytotoxicity Evaluation
4.7. Antimicrobial Synergy Study against M. tuberculosis H37Rv
4.8. Assessment of Frequency of Resistance
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- WHO. The End TB Strategy, (n.d.). Available online: https://www.who.int/teams/global-tuberculosis-programme/the-end-tb-strategy (accessed on 4 June 2021).
- WHO. World Tuberculosis Day 2021-The Clock Is Ticking. (n.d.). 2021. Available online: https://www.who.int/news-room/events/detail/2021/03/24/default-calendar/world-tuberculosis-day-2021---the-clock-is-ticking (accessed on 4 June 2021).
- Ghante, M.H.; Jamkhande, P.G. Role of pentacyclic triterpenoids in chemoprevention and anticancer treatment: An overview on targets and underling mechanisms. J. Pharmacopunct. 2019, 22, 55–67. [Google Scholar] [CrossRef]
- Dehelean, C.A.; Soica, C.; Peev, C.; Gruia, A.T.; Seclaman, E. Physico-chemical and Molecular Analysis of Antitumoral Pentacyclic Triterpenes in Complexation with Gamma-cyclodextrin. Rev. Chim. 2008, 59, 887–890. [Google Scholar]
- Kamble, S.; Goyal, S.; Patil, C. Multifunctional pentacyclic triterpenoids as adjuvants in cancer chemotherapy: A review. RSC Adv. 2014, 4, 33370–33382. [Google Scholar] [CrossRef]
- Cháirez-Ramírez, M.H.; Moreno-Jiménez, M.R.; González-Laredo, R.F.; Gallegos-Infante, J.A.; Rocha-Guzmán, N.E. Lupane-type triterpenes and their anti-cancer activities against most common malignant tumors: A review. EXCLI J. 2016, 15, 758–771. [Google Scholar] [CrossRef]
- Laszczyk, M.N. Pentacyclic triterpenes of the lupane, oleanane and ursane group as tools in cancer therapy. Planta Med. 2009, 75, 1549–1560. [Google Scholar] [CrossRef] [Green Version]
- Ulici, U.A.; Milan, M.A.; Mioc, M.M.; Ghiulai, G.R.; Racoviceanu, R.R.; Șoica, Ș.C. Ring-Modified Triterpene Derivatives as Potential Pharmacological Active Compounds. Timis. Med. J. 2021, 2020, 7. [Google Scholar] [CrossRef]
- Demikhova, O.V.; Balakshin, V.V.; Presnova, G.A.; Bocharova, I.V.; Lepekha, L.N.; Chernousova, L.N.; Smirnova, T.G.; Pospelov, L.E.; Chistiakov, A.N. Antimycobacterial activity of a dry birch bark extract on a model of experimental pulmonary tuberculosis. Probl. Tuberk. I Bolezn. Legk. 2006, 1, 55–57. [Google Scholar]
- Fomogne-Fodjo, M.C.Y.; Ndinteh, D.T.; Olivier, D.K.; Kempgens, P.; van Vuuren, S.; Krause, R.W.M. Secondary metabolites from Tetracera potatoria stem bark with anti-mycobacterial activity. J. Ethnopharmacol. 2017, 195, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Lall, N.; Meyer, J.J.M.; Wang, Y.; Bapela, N.B.; van Rensburg, C.E.J.; Fourie, B.; Franzblau, S.G. Characterization of Intracellular Activity of Antitubercular Constituents the Roots of Euclea natalensis. Pharm. Biol. 2005, 43, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Bocharova, I.V.; Demikhova, O.V.; Erokhin, V.V.; Pospelov, L.E.; Balakshin, V.V.; Chistyakov, A.N.; Mishin, V.J.; Deriabin, P.G. Tuberculosis Treating And Preventing Agent. WIPO Patent WO/2006/001733, 1 May 2006. [Google Scholar]
- Oladosu, I.A.; Lawson, L.; Aiyelaagbe, O.O.; Emenyonu, N.; Afieroho, O.E. Anti-tuberculosis lupane-type isoprenoids from Syzygium guineense Wild DC. (Myrtaceae) stem bark. Future J. Pharm. Sci. 2017, 3, 148–152. [Google Scholar] [CrossRef]
- Akihisa, T.; Franzblau, S.G.; Ukiya, M.; Okuda, H.; Zhang, F.; Yasukawa, K.; Suzuki, T.; Kimura, Y. Antitubercular activity of triterpenoids from Asteraceae flowers. Biol. Pharm. Bull. 2005, 28, 158–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suksamrarn, S.; Panseeta, P.; Kunchanawatta, S.; Distaporn, T.; Ruktasing, S.; Suksamrarn, A. Ceanothane- and lupane-type triterpenes with antiplasmodial and antimycobacterial activities from Ziziphus cambodiana. Chem. Pharm. Bull. 2006, 54, 535–537. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Webster, D.; Johnson, J.A.; Gray, C.A. Anti-mycobacterial triterpenes from the Canadian medicinal plant Alnus incana. J. Ethnopharmacol. 2015, 165, 148–151. [Google Scholar] [CrossRef] [PubMed]
- Thi, T.A.D.; Tuyet, N.T.K.; Pham, C.; Nguyen, H.T.; Thi, C.B.; Duy, T.D.; D’hooghe, M.; van Nguyen, T. Synthesis and cytotoxic evaluation of novel ester-triazole-linked triterpenoid-AZT conjugates. Bioorg. Med. Chem. Lett. 2014, 24, 5190–5194. [Google Scholar] [CrossRef]
- Ledeti, I.; Avram, S.; Bercean, V.; Vlase, G.; Vlase, T.; Ledeti, A.; Zupko, I.; Mioc, M.; Suta, L.-M.; Soica, C.; et al. Solid-State Characterization and Biological Activity of Betulonic Acid Derivatives. Molecules 2015, 20, 22691–22702. [Google Scholar] [CrossRef] [PubMed]
- Pinzaru, I.; Trandafirescu, C.; Szabadai, Z.; Mioc, M.; Ledeti, I.; Coricovac, D.; Ciurlea, S.; Ghiulai, R.M.; Crainiceanu, Z.; Simu, G. Synthesis and Biological Evaluation of Some Pentacyclic Lupane Triterpenoid Esters. Rev. Chim. 2014, 65, 848–851. [Google Scholar]
- Mioc, M.; Avram, S.; Bercean, V.; Kurunczi, L.; Ghiulai, R.M.; Oprean, C.; Coricovac, D.E.; Dehelean, C.; Mioc, A.; Balan-Porcarasu, M.; et al. Design, Synthesis and Biological Activity Evaluation of S-Substituted 1H-5-Mercapto-1,2,4-Triazole Derivatives as Antiproliferative Agents in Colorectal Cancer. Front. Chem. 2018, 6, 373. [Google Scholar] [CrossRef] [Green Version]
- Dehelean, C.A.; Soica, C.; Peev, C.; Ciurlea, S.; Feflea, S.; Kasa, P. A pharmaco-toxicological evaluation of betulinic acid mixed with hydroxipropilgamma cyclodextrin on in vitro and in vivo models. Farmacia 2011, 59, 51–59. [Google Scholar]
- Patra, N.; Rath, J.P. 3-Epi-Betulinic Acid Acetate as A Drug Candidate for Tuberculosis. Biosci. Biotechnol. Res. Commun. 2020, 13, 175–177. [Google Scholar]
- Tanachatchairatana, T.; Bremner, J.B.; Chokchaisiri, R.; Suksamrarn, A. Antimycobacterial activity of cinnamate-based esters of the triterpenes betulinic, oleanolic and ursolic acids. Chem. Pharm. Bull. 2008, 56, 194–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fadipe, V.O.; Mongalo, N.I.; Opoku, A.R.; Dikhoba, P.M.; Makhafola, T.J. Isolation of anti-mycobacterial compounds from Curtisia dentata (Burm.f.) C.A.Sm (Curtisiaceae), BMC Complement. Altern. Med. 2017, 17, 306. [Google Scholar] [CrossRef] [Green Version]
- Kazakova, O.; Medvedeva, N.; Samoilova, I.; Baikova, I.; Tolstikov, G.; Kataev, V.; Mironov, V. Conjugates of several lupane, oleanane, and ursane triterpenoids with the antituberculosis drug isoniazid and pyridinecarboxaldehydes. Chem. Nat. Compd. 2011, 47, 752–758. [Google Scholar] [CrossRef]
- Medvedeva, N.I.; Kazakova, O.B.; Lopatina, T.V.; Smirnova, I.E.; Giniyatullina, G.V.; Baikova, I.P.; Kataev, V.E. Synthesis and antimycobacterial activity of triterpenic A-ring azepanes. Eur. J. Med. Chem. 2018, 143, 464–472. [Google Scholar] [CrossRef]
- Kazakova, O.B.; Medvedeva, N.I.; Smirnova, I.E.; Lopatina, T.V.; Veselovsky, A.V. The Introduction of Hydrazone, Hydrazide, or Azepane Moieties to the Triterpenoid Core Enhances an Activity Against M. tuberculosis. Med. Chem. 2020, 17, 134–145. [Google Scholar] [CrossRef] [PubMed]
- Kazakova, O.; Lopatina, T.; Giniyatullina, G.; Mioc, M.; Soica, C. Antimycobacterial activity of azepanobetulin and its derivative: In vitro, in vivo, ADMET and docking studies. Bioorg. Chem. 2020, 104, 104209. [Google Scholar] [CrossRef] [PubMed]
- Kazakova, O.B.; Lopatina, T.V.; Baikova, I.P.; Zileeva, Z.R.; Vakhitova, Y.V.; Suponitsky, K.Y. Synthesis, evaluation of cytotoxicity, and antimicrobial activity of A-azepano- and A-seco-3-amino-C28-aminolupanes. Med. Chem. Res. 2020, 29, 1507–1519. [Google Scholar] [CrossRef]
- Lopatina, T.V.; Medvedeva, N.I.; Baikova, I.P.; Iskhakov, A.S.; Kazakova, O.B. Synthesis and Cytotoxicity of О- and N-Acyl Derivatives of Azepanobetulin. Russ. J. Bioorganic Chem. 2019, 45, 292–301. [Google Scholar] [CrossRef]
- Petrova, A.V.; Lopatina, T.V.; Mustafin, A.G.; Kazakova, O.B. Modification of Azepanobetulin at the Isopropenyl Group. Russ. J. Org. Chem. 2020, 56, 1582–1587. [Google Scholar] [CrossRef]
- Kazakova, O.; Tret’yakova, E.; Baev, D. Evaluation of A-azepano-triterpenoids and related derivatives as antimicrobial and antiviral agents. J. Antibiot. 2021, 74, 559–573. [Google Scholar] [CrossRef]
- Kazakova, O.; Smirnova, I.; Lopatina, T.; Giniyatullina, G.; Petrova, A.; Khusnutdinova, E.; Csuk, R.; Serbian, I.; Loesche, A. Synthesis and cholinesterase inhibiting potential of A-ring azepano- and 3-amino-3,4-seco-triterpenoids. Bioorg. Chem. 2020, 101, 104001. [Google Scholar] [CrossRef]
- Giniyatullina, G.V.; Kazakova, O.B.; Baikova, I.P.; Yamansarov, E.Y.; Osterman, I.A.; Komarova, E.S.; Skvortsov, D.A.; Saltikova, I.V.; Majouga, A.G.; Ivanenkov, Y.A. Synthesis and Сytotoxicity of A-Azepanobetulinic Acid N-Methyl-Piperazinylamide. Nat. Prod. Commun. 2019, 14, 1934578X19860670. [Google Scholar] [CrossRef] [Green Version]
- Kazakova, O.B.; Brunel, J.M.; Khusnutdinova, E.F.; Negrel, S.; Giniyatullina, G.V.; Lopatina, T.V.; Petrova, A.V. A-Ring-Modified Triterpenoids and Their Spermidine–Aldimines with Strong Antibacterial Activity. Molbank 2019, 2019, M1078. [Google Scholar] [CrossRef] [Green Version]
- Udrescu, L.; Bogdan, P.; Chiş, A.; Sîrbu, I.O.; Topîrceanu, A.; Văruţ, R.-M.; Udrescu, M. Uncovering New Drug Properties in Target-Based Drug–Drug Similarity Networks. Pharmaceutics 2020, 12, 879. [Google Scholar] [CrossRef]
- Čmelo, I.; Voršilák, M.; Svozil, D. Profiling and analysis of chemical compounds using pointwise mutual information. J. Cheminform. 2021, 13, 3. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, L.; Cheng, K.; Jin, B.; Lu, X.; Che, C.; Liu, Y. Exploring Multi-level Mutual Information for Drug-target Interaction Prediction. In Proceedings of the 2020 IEEE International Conference on Bioinformatics and Biomedicine (BIBM), Seoul, Korea, 16–19 December 2020; pp. 251–256. [Google Scholar] [CrossRef]
- Papadopoulou, M.V.; Bloomer, W.D.; Rosenzweig, H.S. The antitubercular activity of various nitro(triazole/imidazole)-based compounds. Bioorganic Med. Chem. 2017, 25, 6039–6048. [Google Scholar] [CrossRef] [PubMed]
- Huitric, E.; Verhasselt, P.; Koul, A.; Andries, K.; Hoffner, S.; Andersson, D.I. Rates and mechanisms of resistance development in Mycobacterium tuberculosis to a novel diarylquinoline ATP synthase inhibitor. Antimicrob. Agents Chemother. 2010, 54, 1022–1028. [Google Scholar] [CrossRef] [Green Version]
- Suresh, A.; Srinivasarao, S.; Khetmalis, Y.M.; Nizalapur, S.; Sankaranarayanan, M.; Sekhar, K.V.G.C. Inhibitors of pantothenate synthetase of Mycobacterium tuberculosis–a medicinal chemist perspective. RSC Adv. 2020, 10, 37098–37115. [Google Scholar] [CrossRef]
- Kumar, S.; Jena, L. Understanding Rifampicin Resistance in Tuberculosis through a Computational Approach. Genom. Inform. 2014, 12, 276–282. [Google Scholar] [CrossRef]
- Chien, J.-Y.; Chiu, W.-Y.; Chien, S.-T.; Chiang, C.-J.; Yu, C.-J.; Hsueh, P.-R. Mutations in gyrA and gyrB among Fluoroquinolone- and Multidrug-Resistant Mycobacterium tuberculosis Isolates. Antimicrob. Agents Chemother. 2016, 60, 2090–2096. [Google Scholar] [CrossRef] [Green Version]
- Hsu, L.-Y.; Lai, L.-Y.; Hsieh, P.-F.; Lin, T.-L.; Lin, W.-H.; Tasi, H.-Y.; Lee, W.-T.; Jou, R.; Wang, J.-T. Two Novel katG Mutations Conferring Isoniazid Resistance in Mycobacterium tuberculosis. Front. Microbiol. 2020, 11, 1644. [Google Scholar] [CrossRef]
- Ducati, R.G.; Ruffino-Netto, A.; Basso, L.A.; Santos, D.S. The resumption of consumption -- a review on tuberculosis. Mem. Inst. Oswaldo Cruz. 2006, 101, 697–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, M.M.; Odell, J.A. Nontuberculous mycobacterial pulmonary infections. J. Thorac. Dis. 2014, 6, 210–220. [Google Scholar] [CrossRef]
- Bento, C.M.; Gomes, M.S.; Silva, T. Looking beyond Typical Treatments for Atypical Mycobacteria. Antibiotics 2020, 9, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, W.; Mandal, S.; Degen, D.; Liu, Y.; Ebright, Y.W.; Li, S.; Feng, Y.; Zhang, Y.; Mandal, S.; Jiang, Y.; et al. Ebright, Structural Basis of Mycobacterium tuberculosis Transcription and Transcription Inhibition. Mol. Cell. 2017, 66, 169–179.e8. [Google Scholar] [CrossRef] [Green Version]
- Rothstein, D.M. Rifamycins, Alone and in Combination. Cold Spring Harb. Perspect. Med. 2016, 6, a027011. [Google Scholar] [CrossRef] [PubMed]
- Mahmud, H.A.; Seo, H.; Kim, S.; Islam, M.I.; Sultana, O.F.; Nam, K.W.; Lee, B.E.; Sadu, V.S.; Lee, K.I.; Song, H.Y. Synthesis and activity of BNF15 against drug-resistant Mycobacterium tuberculosis. Future Med. Chem. 2021, 13, 251–267. [Google Scholar] [CrossRef]
- Zheng, X.; Av-Gay, Y. System for Efficacy and Cytotoxicity Screening of Inhibitors Targeting Intracellular Mycobacterium tuberculosis. J. Vis. Exp. 2017, 122, 55273. [Google Scholar] [CrossRef]
- Zhou, J.; Xu, R.; Du, X.-Z.; Zhou, X.-D.; Li, Q. Saxifragifolin D attenuates phagosome maturation arrest in Mycobacterium tuberculosis-infected macrophages via an AMPK and VPS34-dependent pathway. AMB Express 2017, 7, 11. [Google Scholar] [CrossRef] [Green Version]
- Gopalaswamy, R.; Shanmugam, S.; Mondal, R.; Subbian, S. Of tuberculosis and non-tuberculous mycobacterial infections-a comparative analysis of epidemiology, diagnosis and treatment. J. Biomed. Sci. 2020, 27, 74. [Google Scholar] [CrossRef]
- Wu, M.-L.; Aziz, D.B.; Dartois, V.; Dick, T. NTM drug discovery: Status, gaps and the way forward. Drug Discov. Today 2018, 23, 1502–1519. [Google Scholar] [CrossRef]
- Saxena, S.; Spaink, H.P.; Forn-Cuní, G. Drug Resistance in Nontuberculous Mycobacteria: Mechanisms and Models. Biology 2021, 10, 96. [Google Scholar] [CrossRef] [PubMed]
- Pires, C.T.A.; Brenzan, M.A.; Scodro, R.B.D.; Cortez, D.A.G.; Lopes, L.D.G.; Siqueira, V.L.D.; Cardoso, R.F. Anti-Mycobacterium tuberculosis activity and cytotoxicity of Calophyllum brasiliense Cambess (Clusiaceae). Mem. Inst. Oswaldo Cruz. 2014, 109, 324–329. [Google Scholar] [CrossRef]
- Scandorieiro, S.; de Camargo, L.C.; Lancheros, C.A.C.; Yamada-Ogatta, S.F.; Nakamura, C.V.; de Oliveira, A.G.; Andrade, C.G.T.J.; Duran, N.; Nakazato, G.; Kobayashi, R.K.T. Synergistic and Additive Effect of Oregano Essential Oil and Biological Silver Nanoparticles against Multidrug-Resistant Bacterial Strains. Front. Microbiol. 2016, 7, 760. [Google Scholar] [CrossRef] [PubMed]
- Shang, J.-H.; Xu, G.-W.; Zhu, H.-T.; Wang, D.; Yang, C.-R.; Zhang, Y.-J. Anti-inflammatory and Cytotoxic Triterpenes from the Rot Roots of Panax notoginseng. Nat. Products Bioprospect. 2019, 9, 287–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirzayev, F.; Viney, K.; Linh, N.N.; Gonzalez-Angulo, L.; Gegia, M.; Jaramillo, E.; Zignol, M.; Kasaeva, T. World Health Organization recommendations on the treatment of drug-resistant tuberculosis, 2020 update. Eur. Respir. J. 2021, 57, 2003300. [Google Scholar] [CrossRef]
- O’Neill, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations; Government of UK: London, UK, 2016. [Google Scholar]
- Keam, S.J. Pretomanid: First Approval. Drugs 2019, 79, 1797–1803. [Google Scholar] [CrossRef]
- Khoshnood, S.; Goudarzi, M.; Taki, E.; Darbandi, A.; Kouhsari, E.; Heidary, M.; Motahar, M.; Moradi, M.; Bazyar, H. Bedaquiline: Current status and future perspectives. J. Glob. Antimicrob. Resist. 2021, 25, 48–59. [Google Scholar] [CrossRef]
- Lee, J.J.; Lee, S.-K.; Song, N.; Nathan, T.O.; Swarts, B.M.; Eum, S.-Y.; Ehrt, S.; Cho, S.-N.; Eoh, H. Transient drug-tolerance and permanent drug-resistance rely on the trehalose-catalytic shift in Mycobacterium tuberculosis. Nat. Commun. 2019, 10, 2928. [Google Scholar] [CrossRef]
- Freedman, D. Statistical Models: Theory and Practice; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Hidalgo, B.; Goodman, M. Multivariate or multivariable regression? Am. J. Public Health 2013, 103, 39–40. [Google Scholar] [CrossRef]
- Fix, E.; Hodges, J.L. Discriminatory Analysis. Nonparametric Discrimination: Consistency Properties. Int. Stat. Rev./Rev. Int. Stat. 1989, 57, 238–247. [Google Scholar] [CrossRef]
- Ho, T.K. Random decision forests. In Proceedings of the 3rd International Conference on Document Analysis and Recognition, Montreal, QC, Canada, 14–16 August 1995; Volume 1, pp. 278–282. [Google Scholar] [CrossRef]
- Géron, A. Hands-on Machine Learning with Scikit-Learn, Keras, and TensorFlow: Concepts, Tools, and Techniques to Build Intelligent Systems; O’Reilly Media: Newton, MA, USA, 2017. [Google Scholar]
- Pedregosa, F.; Gaël, V.; Alexandre, G.; Vincent, M.; Bertrand, T.; Olivier, G.; Mathieu, B. Scikit-learn: Machine learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- Buitinck, L.; Louppe, G.; Blondel, M.; Pedregosa, F.; Mueller, A.; Grisel, O.; Niculae, V.; Prettenhofer, P.; Gramfort, A.; Grobler, J.; et al. API Design for Machine Learning Software: Experiences from the Scikit-Learn Project. 2013. Available online: https://arxiv.org/abs/1309.0238 (accessed on 17 July 2021).
- Wishart, D.S.; Feunang, Y.D.; Guo, A.C.; Lo, E.J.; Marcu, A.; Grant, J.R.; Sajed, T.; Johnson, D.; Li, C.; Sayeeda, Z.; et al. Wilson, DrugBank 5.0: A major update to the DrugBank database for 2018. Nucleic Acids Res. 2018, 46, D1074–D1082. [Google Scholar] [CrossRef] [PubMed]
- Cover, T.M.; Thomas, J.A. Elements of Information Theory Second Edition Solutions to Problems; WILEY: Hoboken, NJ, USA, 2006. [Google Scholar]
- Jianu, C.; Stoin, D.; Cocan, I.; David, I.; Pop, G.; Lukinich-Gruia, A.T.; Mioc, M.; Mioc, A.; Șoica, C.; Muntean, D.; et al. In Silico and In Vitro Evaluation of the Antimicrobial and Antioxidant Potential of Mentha × smithiana R. GRAHAM Essential Oil from Western Romania. Foods 2021, 10, 815. [Google Scholar] [CrossRef]
- Mioc, M.; Soica, C.; Bercean, V.; Avram, S.; Balan-Porcarasu, M.; Coricovac, D.; Ghiulai, R.; Muntean, D.; Andrica, F.; Dehelean, C.; et al. Design, synthesis and pharmaco-toxicological assessment of 5-mercapto-1,2,4-triazole derivatives with antibacterial and antiproliferative activity. Int. J. Oncol. 2017, 50, 1175–1183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oprean, C.; Mioc, M.; Csányi, E.; Ambrus, R.; Bojin, F.; Tatu, C.; Cristea, M.; Ivan, A.; Danciu, C.; Dehelean, C.; et al. Improvement of ursolic and oleanolic acids’ antitumor activity by complexation with hydrophilic cyclodextrins. Biomed. Pharmacother. 2016, 83, 1095–1104. [Google Scholar] [CrossRef]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The protein data bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- EUCAST, ESCMID. Terminology relating to methods for the determination of susceptibility of bacteria to antimicrobial agents. Clin. Microbiol. Infect. 2000, 6, 503–508. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | KMeans | Linear Regression | KNeigh | MLP | SVC | Linear SVC | Random Forest | Aggregated Score |
|---|---|---|---|---|---|---|---|---|
| 1 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0.57 |
| 2 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0.57 |
| 3 | 1 | 0 | 1 | 0 | 1 | 1 | 0 | 0.57 |
| 4 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0.14 |
| 5 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0.14 |
| 6 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 0.71 |
| 7 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 0.86 |
| 8 | 0 | 0 | 1 | 1 | 1 | 0 | 1 | 0.57 |
| 9 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 0.71 |
| 10 | 0 | 1 | 1 | 1 | 1 | 1 | 0 | 0.71 |
| 11 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 0.86 |
| 12 | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 0.71 |
| 13 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0.14 |
| 14 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0.29 |
| 15 | 1 | 0 | 0 | 1 | 1 | 1 | 0 | 0.57 |
| 16 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0.14 |
| 17 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0.29 |
| 18 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0.29 |
| 20 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0.14 |
| 22 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0.14 |
| Compound | Most Similar Drug | Second Most Similar Drug | Third Most Similar Drug |
|---|---|---|---|
| 1 | Pyrazinamide | Rifampicin | Protionamide |
| 2 | Cycloserine | Protionamide | Rifapentine |
| 3 | Rifampicin | Protionamide | Rifapentine |
| 4 | Rifapentine | Protionamide | Protionamide |
| 5 | Rifampicin | Protionamide | Ethambutol |
| 6 | Rifampicin | Pyrazinamide | Rifapentine |
| 7 | Protionamide | Terizidone | Enviomycin |
| 8 | Terizidone | Rifampicin | Protionamide |
| 9 | Rifampicin | Protionamide | Rifapentine |
| 10 | Rifampicin | Pyrazinamide | Enviomycin |
| 11 | Rifapentine | Protionamide | Protionamide |
| 12 | Rifampicin | Protionamide | Rifapentine |
| 13 | Protionamide | Pyrazinamide | Enviomycin |
| 14 | Rifampicin | Protionamide | Rifapentine |
| 15 | Rifampicin | Terizidone | Streptomycin |
| 16 | Rifampicin | Protionamide | Pyrazinamide |
| 17 | Rifampicin | Protionamide | Rifapentine |
| 18 | Rifampicin | Rifapentine | Streptomycin |
| 20 | Protionamide | Terizidone | Rifampicin |
| 22 | Protionamide | Rifamycin | Rifampicin |
| Compound | MIC (µM) | |||||
|---|---|---|---|---|---|---|
| H37Rv | H37Rv (rpoBS450L) | H37Rv (katGdel) | H37Rv (gyrAD94K) | M. abscessus 19977 | M. avium 25291 | |
| 1 | 4 * | 4 * | 2 * | 4 * | 8 * | 0.125 * |
| 2 | 4 | 4 | 4 | 4 | 4 | 1 |
| 3 | 8 | 4 | 4 | 8 | 16 | 2 |
| 4 | 16 | 8 | 8 | 16 | 16 | 4 |
| 5 | 8 | 8 | 4 | 8 | 16 | 1 |
| 6 | 8 | 4 | 4 | 4 | 8 | 1 |
| 7 | 0.5 | 0.5 | 8 | 0.5 | 8 | 4 |
| 8 | 8 | 4 | 4 | 4 | 32 | 4 |
| 9 | 2 | 2 | 1 | 2 | 2 | 4 |
| 10 | 4 | 4 | 4 | 4 | 8 | 2 |
| 11 | 8 | 4 | 8 | 8 | 16 | 4 |
| 12 | 32 | 32 | 32 | >32 | 8 | 4 |
| 13 | 32 | 8 | 8 | 16 | 16 | 2 |
| 14 | >32 | >32 | 16 | >32 | >64 | 0.0625 |
| 15 | >32 | >32 | 32 | >32 | >64 | 64 |
| 16 | 4 | 32 | 2 | 2 | >64 | 4 |
| 17 | 16 | 8 | 8 | 16 | 16 | 0.5 |
| 18 | 16 | 16 | 16 | 8 | 32 | 4 |
| 22 | >32 | 4 | 16 | >32 | >64 | 64 |
| 20 | 4 | 8 | 4 | 4 | 16 | 4 |
| Rifampin | ≤0.004 | >1 | ≤0.004 | ≤0.004 | ND | 0.0625 |
| Isoniazid | 0.03 | 0.06 | >2 | 0.06 | - | - |
| Moxifloxacin | 0.06 | 0.03 | 0.03 | >2 | - | - |
| Pretomanid | 0.125 | 0.06 | 0.06 | 0.125 | - | - |
| Clarithromycin | - | - | - | - | 4 | 0.125 |
| Amikacin | - | - | - | - | 8 | ND |
| Protein PBD ID | 5UHB | 5UHE | |
|---|---|---|---|
| Ligand | Binding Free Energy ∆G (kcal/mol) | ||
| Native ligand | −10.3 | −10.9 | |
| Compound 1 | −7 | −8 | |
| Compound 2 | −7.1 | −6.5 | |
| Compound 3 | −6.3 | −5.6 | |
| Compound 4 | −7.9 | −6.0 | |
| Compound 5 | −7.4 | −5.6 | |
| Compound 6 | −7.6 | −6.1 | |
| Compound 7 | −8.3 | −2.5 | |
| Compound 8 | −7.2 | −7.6 | |
| Compound 9 | −8.2 | −4.5 | |
| Compound 10 | −7.4 | −5.6 | |
| Compound 11 | −8 | −4.7 | |
| Compound 12 | −7.2 | −4.3 | |
| Compound 13 | −7.8 | −9.0 | |
| Compound 14 | −6.9 | −4.5 | |
| Compound 15 | −7.2 | −4.8 | |
| Compound 16 | −6.9 | −4.8 | |
| Compound 17 | −6.7 | −4,8 | |
| Compound 18 | −6.9 | −5.1 | |
| Compound 20 | −7.6 | −5.9 | |
| Compound 22 | −7.8 | −3.5 | |
| Compound | M. avium Ex Vivo Efficacy (>90% Inhibition µM) | M. abscessus Ex Vivo Efficacy (>90% Inhibition µM) | M. tuberculosis Ex Vivo Efficacy (% Inhibition at MIC) |
|---|---|---|---|
| 1 | 8 | >128 | ND |
| 2 | >128 | >128 | 87% |
| 3 | >128 | >128 | 68% |
| 5 | 32 | >128 | 84% |
| 7 | 128 | >128 | 86% |
| 8 | 16 | >128 | 60% |
| 9 | >128 | >128 | 92% |
| 10 | 64 | >128 | 86% |
| 17 | >128 | >128 | 87% |
| 18 | 128 | >128 | 80% |
| 22 | 128 | >128 | ND |
| Clarithromycin | 8 | 128 | ND |
| Isoniazid | ND | ND | 91% |
| Rifampin | ND | ND | 93% |
| Compound | THP-1 Cytotoxicity (CC50 µM) | THP-1 SI | HepG2 Cytotoxicity (CC50 µM) | HepG2 SI | HeLa Cytotoxicity (CC50 µM) | HeLa SI |
|---|---|---|---|---|---|---|
| 2 | 2.006 | 0.5 | 0.3305 | 0.08 | 2.064 | 0.52 |
| 3 | 8.046 | 1 | 0.5936 | 0.07 | 2.767 | 0.34 |
| 5 | 15.78 | 1.97 | 5.914 | 0.74 | 17.73 | 2.22 |
| 7 | 10.25 | 20.5 | 1.917 | 3.83 | 11.26 | 22.52 |
| 8 | 4.39 | 0.55 | 0.7154 | 0.09 | 0.9302 | 0.46 |
| 9 | 3.334 | 1.66 | 0.2422 | 0.12 | 2.419 | 1.21 |
| 10 | 11.54 | 2.88 | 1.522 | 0.38 | 9.542 | 2.38 |
| 17 | 7.534 | 0.47 | 0.7124 | 0.04 | 3.771 | 0.24 |
| 18 | 13.03 | 0.81 | 1.098 | 0.07 | 8.893 | 0.55 |
| 22 | 4.972 | 0.15 | 1.432 | 0.09 | 2.21 | 0.07 |
| Mitomycin C | 3.094 | 16.6 | 8.89 |
| Compound | Rifampin | Isoniazid | Ethambutol |
|---|---|---|---|
| 3 | 1.0 Additive | 1.0 Additive | 1.5 Indifferent |
| 7 | 1.0 Additive | 0.8 Additive | 1.0 Additive |
| 9 | 1.5 Indifferent | N/A | 2.5 Indifferent |
| 22 | 1.0 Additive | N/A | 1.5 Indifferent |
| Compound | ~105 CFU Plated | ~106 CFU Plated | ~107 CFU Plated |
|---|---|---|---|
| 2× the MIC = 4.00 µg mL−1 Bedaquiline (BDQ) | 0 | 3.13 ×10−7 | 1.43 ×10−7 |
| 8× the MIC = 16.00 µg mL−1 Bedaquiline (BDQ) | 0 | 3.13 ×10−7 | 3.57 ×10−8 |
| 16× the MIC = 32.00 µg mL−1 Bedaquiline (BDQ) | 0 | 0 | 3.57 ×10−8 |
| 2× the MIC = 2.00 µg mL−1 Ethambutol (EMB) | 9.44 ×10−5 | 2.78 ×10−5 | 3.61 ×10−6 |
| 8× the MIC = 8.00 µg mL−1 Ethambutol (EMB) | 1.11 ×10−5 | 3.75 ×10−6 | 8.57 ×10−7 |
| 16× the MIC = 16.00 µg mL−1 Ethambutol (EMB) | 0 | 0 | 1.79 ×10−7 |
| 2× the MIC = 0.25 µg mL−1 Isoniazid (INH) | 1.39 ×10−4 | 3.75 ×10−5 | 5.00 ×10−6 |
| 8× the MIC = 1.00 µg mL−1 Isoniazid (INH) | 5.56 ×10−5 | 3.31 ×10−5 | 3.93 ×10−6 |
| 16× the MIC = 2.00 µg mL−1 Isoniazid (INH) | 1.67 ×10−5 | 2.63 ×10−5 | 2.71 ×10−6 |
| 2× the MIC = 2.00 µg mL−1 Ofloxacin (OFX) | 0 | 1.56 ×10−6 | 5.36 ×10−7 |
| 8× the MIC = 8.00 µg mL−1 Ofloxacin (OFX) | 0 | 9.38 ×10−7 | 7.14 ×10−8 |
| 16× the MIC = 16.00 µg mL−1 Ofloxacin (OFX) | 0 | 0 | 3.57 ×10−8 |
| 2× the MIC = 1.00 µg mL−1 Pretomanid (PRT) | 6.94 ×10−5 | 1.06 ×10−5 | 1.79 ×10−6 |
| 8× the MIC = 4.00 µg mL−1 Pretomanid (PRT) | 3.89 ×10−5 | 6.88 ×10−6 | 8.57 ×10−7 |
| 16× the MIC = 8.00 µg mL−1 Pretomanid (PRT) | 8.33 ×10−6 | 3.13 ×10−7 | 1.43 ×10−7 |
| 2× the MIC = 1.00 µg mL−1 Rifampicin (RIF) | 2.78 ×10−6 | 3.13 ×10−6 | 5.36 ×10−7 |
| 8× the MIC = 4.00 µg mL−1 Rifampicin (RIF) | 0 | 3.13 ×10−7 | 7.14 ×10−8 |
| 16× the MIC = 8.00 µg mL−1 Rifampicin (RIF) | 0 | 0 | 3.57 ×10−8 |
| 2× the MIC = 8.00 µg mL−1 compound 1 | 2.78 ×10−6 | 3.13 ×10−7 | 1.07 ×10−7 |
| 8× the MIC = 32.00 µg mL−1 compound 1 | 0 | 9.38 ×10−7 | 7.14 ×10−8 |
| 16× the MIC = 64.00 µg mL−1 compound 1 | 0 | 0 | 0 |
| 2× the MIC = 32.00 µg mL−1 compound 18 | 5.56 ×10−6 | 7.19 ×10−6 | 4.64 ×10−7 |
| 8× the MIC = 128.00 µg mL−1 compound 18 | 0 | 3.13 ×10−7 | 3.57 ×10−8 |
| 16× the MIC = 256.00 µg mL−1 compound 18 | 0 | 0 | 0 |
| 2× the MIC = 64.00 µg mL−1 compound 22 | 2.78 ×10−6 | 1.25 ×10−6 | 5.36 ×10−7 |
| 8× the MIC = 256.00 µg mL−1 compound 22 | 0 | 1.56 ×10−6 | 1.07 ×10−7 |
| 16× the MIC = 512.00 µg mL−1 compound 22 | 0 | 0 | 0 |
| Protein | PDB ID | Grid Box Center Coordinates | Grid Box Size | Conformers Generated Per Ligand |
|---|---|---|---|---|
| Mycobacterium tuberculosis RNA polymerase (Mtb RNAP)-rifampin complex | 5UHB | Center_x = 164.393813216 center_y = 162.642568841 center_z = 18.7402222979 | size_x = 17.2315566064 size_y = 25.0 size_z = 19.8647468819 | 8 |
| Mycobacterium tuberculosisRNA polymerase- N-aroyl-N-aryl-phenylalaninamides (AAPs) complex | 5UHE | Center_x = −33.0902019309 center_y = −21.9465676304 center_z = 13.5807318 | size_x = 14.3058881805 size_y = 25.0 size_z = 13.9275973862 | 8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kazakova, O.; Racoviceanu, R.; Petrova, A.; Mioc, M.; Militaru, A.; Udrescu, L.; Udrescu, M.; Voicu, A.; Cummings, J.; Robertson, G.; et al. New Investigations with Lupane Type A-Ring Azepane Triterpenoids for Antimycobacterial Drug Candidate Design. Int. J. Mol. Sci. 2021, 22, 12542. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212542
Kazakova O, Racoviceanu R, Petrova A, Mioc M, Militaru A, Udrescu L, Udrescu M, Voicu A, Cummings J, Robertson G, et al. New Investigations with Lupane Type A-Ring Azepane Triterpenoids for Antimycobacterial Drug Candidate Design. International Journal of Molecular Sciences. 2021; 22(22):12542. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212542
Chicago/Turabian StyleKazakova, Oxana, Roxana Racoviceanu, Anastasiya Petrova, Marius Mioc, Adrian Militaru, Lucreția Udrescu, Mihai Udrescu, Adrian Voicu, Jason Cummings, Gregory Robertson, and et al. 2021. "New Investigations with Lupane Type A-Ring Azepane Triterpenoids for Antimycobacterial Drug Candidate Design" International Journal of Molecular Sciences 22, no. 22: 12542. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212542