
Genome-Wide Analysis of MYB10 Transcription Factor in Fragaria and Identification of QTLs Associated with Fruit Color in Octoploid Strawberry

 ,
, 
 and
and
Abstract
:1. Introduction
2. Results
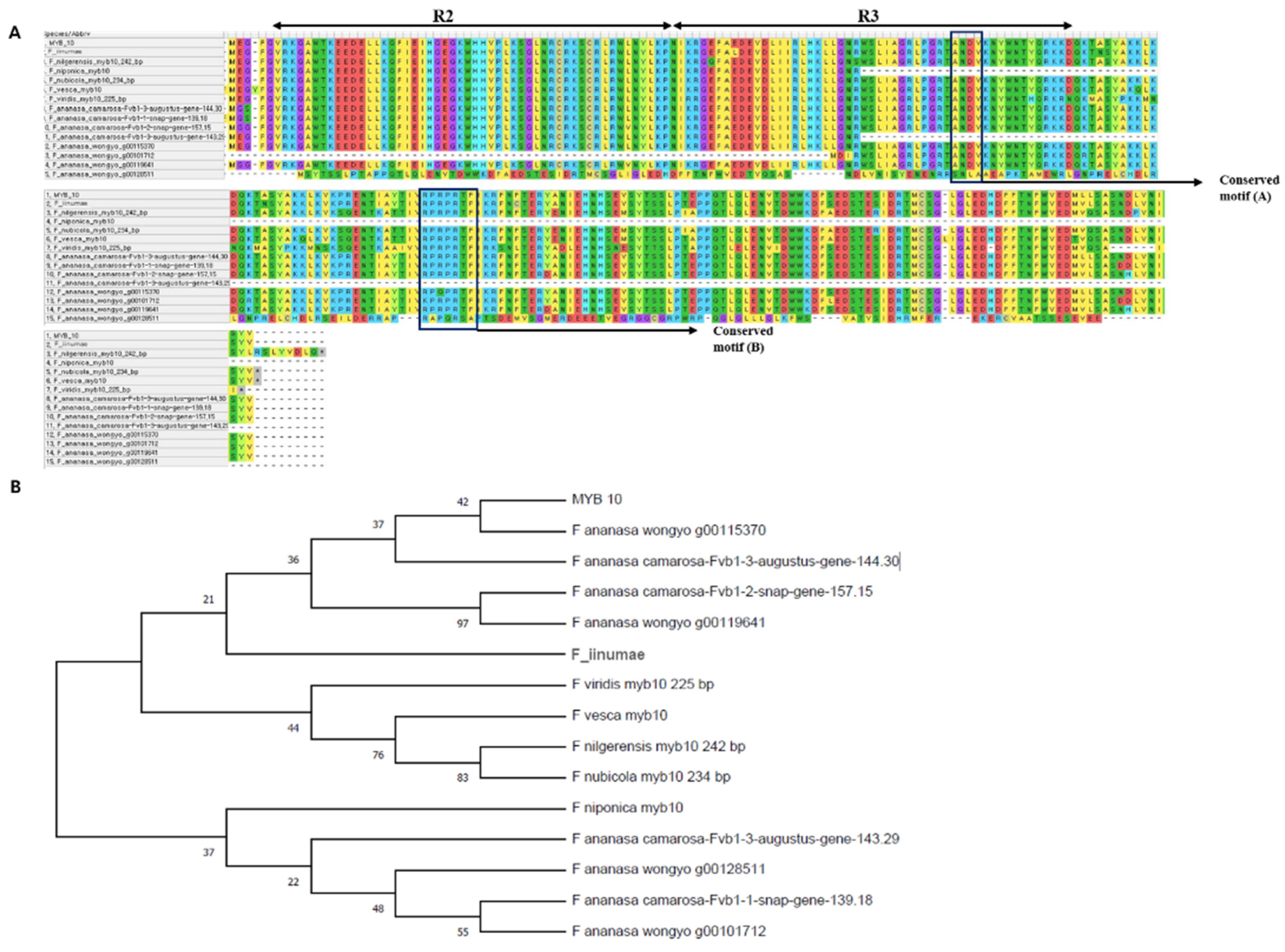
2.1. Characterization of MYB10 in Diploid and Octoploid Species of Fragaria
2.2. Structure and Phylogeny of MYB10 in Diploid and Octoploid Species of Fragaria
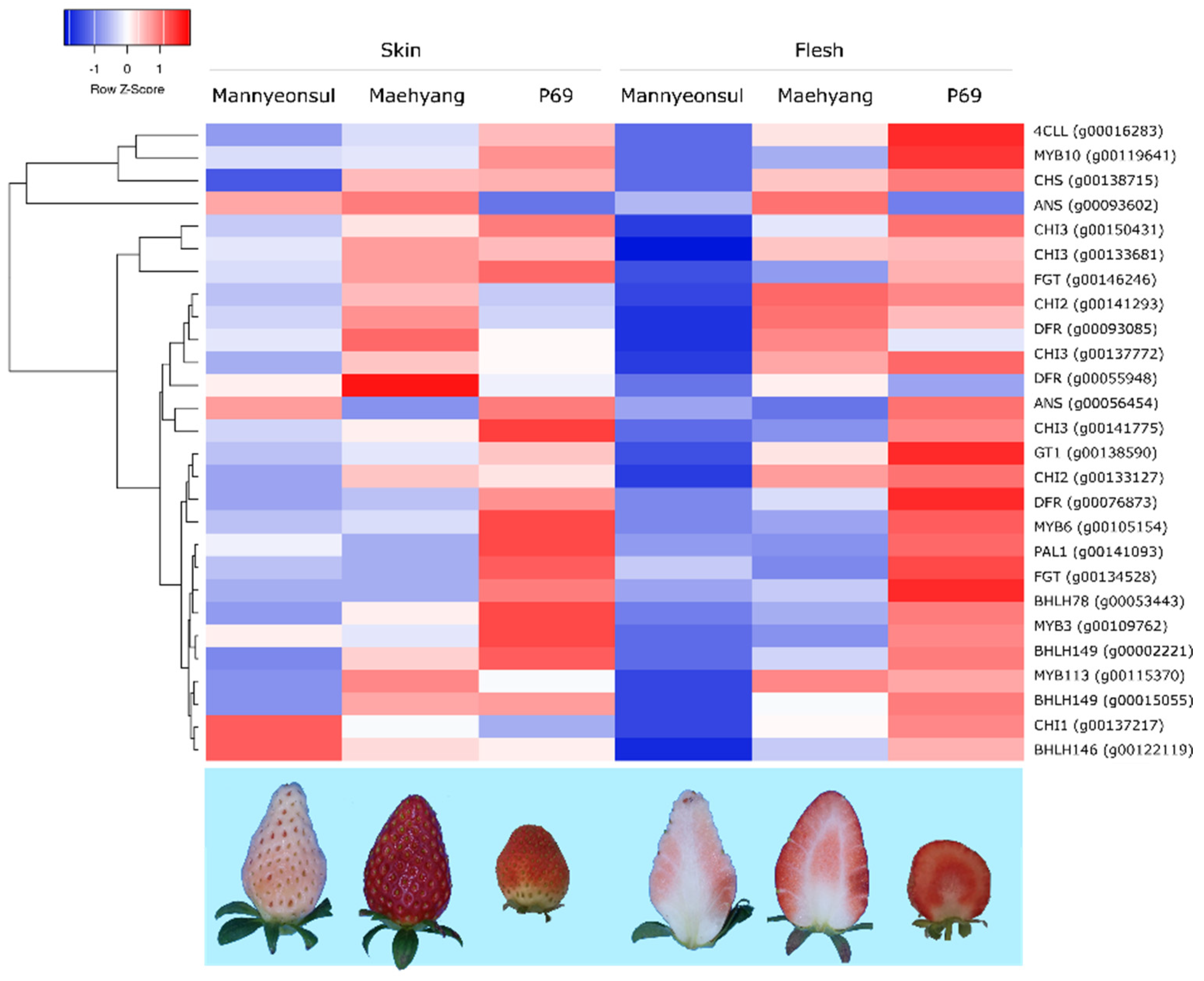
2.3. RNA-Seq Based Expression Profiling of MYB10 and Anthocyanin Biosynthesis Genes
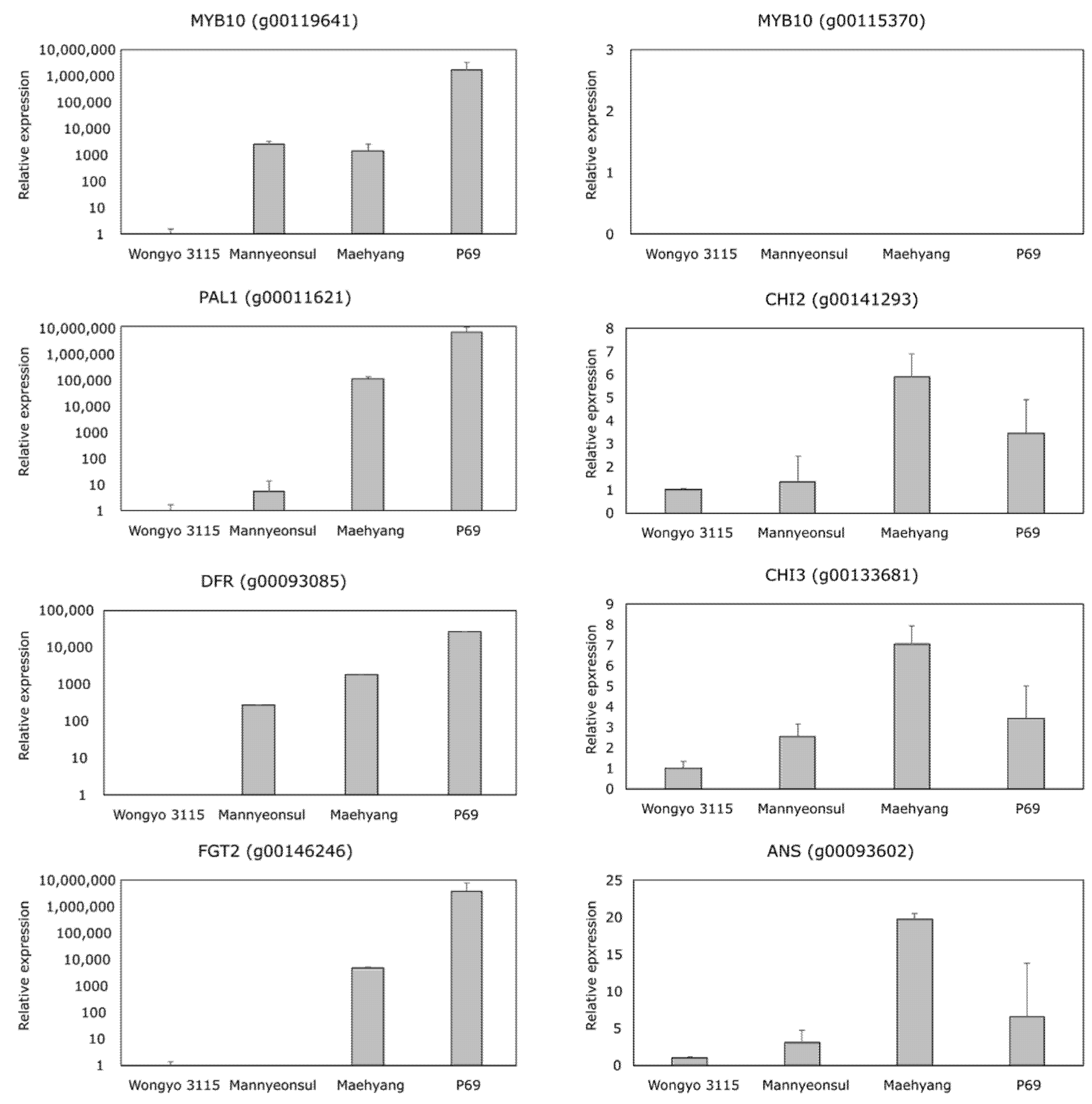
2.4. Relative Expression Analysis of MYB10 and Anthocyanin Biosynthesis Genes
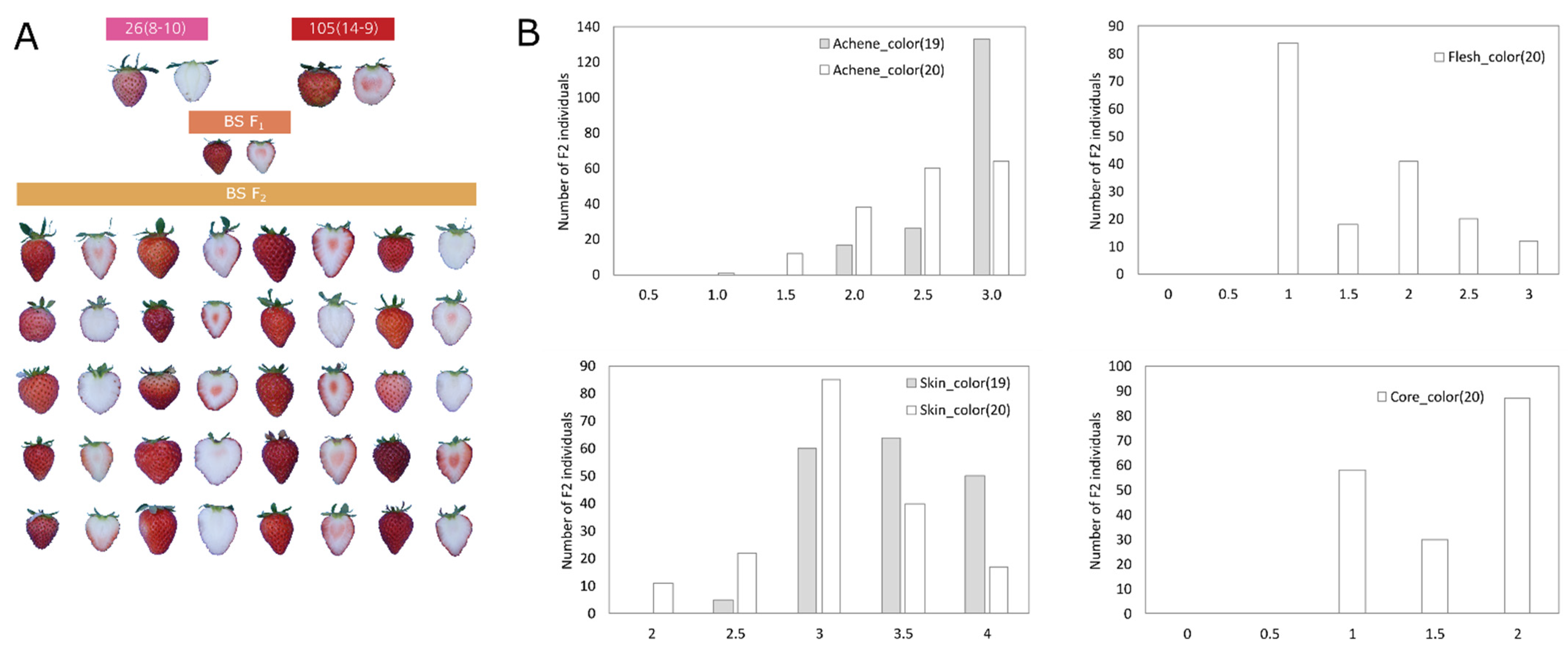
2.5. Identification of QTLs Associated with Fruit Color
2.6. Identification of Predicted Candidate Genes Associated with Fruit Color Using QTL and RNA-Seq Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Identification of MYB10 Transcription Factors in Fragaria
4.3. Analysis of Conserved Motifs, Gene Structures, and Cis-Elements
4.4. RNA-Seq Analysis of Skin and Flesh
4.5. RT-PCR Analysis
4.6. QTL Analysis and Identification of Candidate Genes
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Pabo, C.O.; Sauer, R.T. Transcription factors: Structural families and principles of DNA recognition. Annu. Rev. Biochem. 1992, 61, 1053–1095. [Google Scholar] [CrossRef] [PubMed]
- Riechmann, J.L.; Heard, J.; Martin, G.; Reuber, L.; Jiang, C.-Z.; Keddie, J.; Adam, L.; Pineda, O.; Ratcliffe, O.J.; Samaha, R.R.; et al. Arabidopsis Transcription Factors: Genome-Wide Comparative Analysis Among Eukaryotes. Science 2000, 290, 2105–2110. [Google Scholar] [CrossRef] [PubMed]
- Pillet, J.; Yu, H.-W.; Chambers, A.H.; Whitaker, V.M.; Folta, K.M. Identification of candidate flavonoid pathway genes using transcriptome correlation network analysis in ripe strawberry (Fragaria × ananassa) fruits. J. Exp. Bot. 2015, 66, 4455–4467. [Google Scholar] [CrossRef]
- Zhang, Y.; Butelli, E.; Martin, C. Engineering anthocyanin biosynthesis in plants. Curr. Opin. Plant Biol. 2014, 19, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Petroni, K.; Tonelli, C. Recent advances on the regulation of anthocyanin synthesis in reproductive organs. Plant Sci. 2011, 181, 219–229. [Google Scholar] [CrossRef]
- Wang, N.; Qu, C.; Jiang, S.; Chen, Z.; Xu, H.; Fang, H.; Su, M.; Zhang, J.; Wang, Y.; Liu, W.; et al. The proanthocyanidin specific transcription factor Md MYBPA 1 initiates anthocyanin synthesis under low temperature conditions in red-fleshed apples. Plant J. 2018, 96, 39–55. [Google Scholar] [CrossRef] [Green Version]
- Hichri, I.; Barrieu, F.; Bogs, J.; Kappel, C.; Delrot, S.; Lauvergeat, V. Recent advances in the transcriptional regulation of the flavonoid biosynthetic pathway. J. Exp. Bot. 2011, 62, 2465–2483. [Google Scholar] [CrossRef] [Green Version]
- Stracke, R.; Werber, M.; Weisshaar, B. The R2R3-MYB gene family in Arabidopsis thaliana. Curr. Opin. Plant Biol. 2001, 4, 447–456. [Google Scholar] [CrossRef]
- Zhang, J.; Lei, Y.; Wang, B.; Li, S.; Yu, S.; Wang, Y.; Li, H.; Liu, Y.; Ma, Y.; Dai, H.; et al. The high-quality genome of diploid strawberry (Fragaria nilgerrensis) provides new insights into anthocyanin accumulation. Plant Biotechnol. J. 2020, 18, 1908–1924. [Google Scholar] [CrossRef] [Green Version]
- Castillejo, C.; Waurich, V.; Wagner, H.; Ramos, R.; Oiza, N.; Muñoz, P.; Triviño, J.C.; Caruana, J.; Liu, Z.; Cobo, N.; et al. Allelic Variation of MYB10 Is the Major Force Controlling Natural Variation in Skin and Flesh Color in Strawberry (Fragaria spp.) Fruit. Plant Cell 2020, 32, 3723–3749. [Google Scholar] [CrossRef]
- Edger, P.P.; Poorten, T.J.; VanBuren, R.; Hardigan, M.A.; Colle, M.; McKain, M.R.; Smith, R.D.; Teresi, S.J.; Nelson, A.D.L.; Wai, C.M.; et al. Origin and evolution of the octoploid strawberry genome. Nat. Genet. 2019, 51, 541–547. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.-E.; Manivannan, A.; Lee, S.Y.; Han, K.; Yeum, J.-G.; Jo, J.; Kim, J.; Rho, I.R.; Lee, Y.-R.; Lee, E.S.; et al. Chromosome Level Assembly of Homozygous Inbred Line ‘Wongyo 3115’ Facilitates the Construction of a High-Density Linkage Map and Identification of QTLs Associated with Fruit Firmness in Octoploid Strawberry (Fragaria × ananassa). Front. Plant Sci. 2021, 12, 1337. [Google Scholar] [CrossRef] [PubMed]
- Lerceteau-Köhler, E.; Moing, A.; Guérin, G.; Renaud, C.; Petit, A.; Rothan, C.; Denoyes, B. Genetic dissection of fruit quality traits in the octoploid cultivated strawberry highlights the role of homoeo-QTL in their control. Theor. Appl. Genet. 2012, 124, 1059–1077. [Google Scholar] [CrossRef] [Green Version]
- Labadie, M.; Vallin, G.; Petit, A.; Ring, L.; Hoffmann, T.; Gaston, A.; Potier, A.; Schwab, W.G.; Rothan, C.; Denoyes, B. Metabolite Quantitative Trait Loci for Flavonoids Provide New Insights into the Genetic Architecture of Strawberry (Fragaria × ananassa) Fruit Quality. J. Agric. Food Chem. 2020, 68, 6927–6939. [Google Scholar] [CrossRef] [PubMed]
- Jassmine, Z.; Cabeza, A.; Domínguez, P.; Medina, J.J.; Valpuesta, V.; Denoyes, B.; Sevilla, J.F.S.; Amaya, I. Quantitative trait loci and underlying candidate genes controlling agronomical and fruit quality traits in octoploid strawberry (Fragaria × ananassa). Theor. Appl. Genet. 2011, 123, 755–778. [Google Scholar] [CrossRef]
- Pott, D.M.; Vallarino, J.G.; Cruz-Rus, E.; Willmitzer, L.; Sánchez-Sevilla, J.F.; Amaya, I.; Osorio, S. Genetic analysis of phenylpropanoids and antioxidant capacity in strawberry fruit reveals mQTL hotspots and candidate genes. Sci. Rep. 2020, 10, 20197. [Google Scholar] [CrossRef] [PubMed]
- Alarfaj, R.; El-Soda, M.; Antanaviciute, L.; Vickerstaff, R.; Hand, P.; Harrison, R.J.; Wagstaff, C. Mapping QTL under-lying fruit quality traits in an F1 strawberry population. J. Hortic. Sci. Biotechnol. 2021, 96, 634–645. [Google Scholar] [CrossRef]
- Lin-Wang, K.; Bolitho, K.; Grafton, K.; Kortstee, A.; Karunairetnam, S.; McGhie, T.K.; Espley, R.V.; Hellens, R.P.; Allan, A.C. An R2R3 MYB transcription factor associated with regulation of the anthocyanin biosynthetic pathway in Rosaceae. BMC Plant Biol. 2010, 10, 50. [Google Scholar] [CrossRef] [Green Version]
- Parra-Palma, C.; Morales-Quintana, L.; Ramos, P. Phenolic Content, Color Development, and Pigment–Related Gene Expression: A Comparative Analysis in Different Cultivars of Strawberry during the Ripening Process. Agronomy 2020, 10, 588. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Xu, P.; Pan, J.; Li, Y.; Zhou, J.; Kuang, H.; Lian, H. Inhibition of FvMYB10 transcriptional activity promotes color loss in strawberry fruit. Plant Sci. 2020, 298, 110578. [Google Scholar] [CrossRef]
- Fu, D.; Mason, A.S.; Xiao, M.; Yan, H. Effects of genome structure variation, homeologous genes and repetitive DNA on polyploid crop research in the age of genomics. Plant Sci. 2016, 242, 37–46. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Y.; Dou, Y.; Li, W.; Wang, S.; Shi, W.; Sun, Y.; Zhang, Z. Single nucleotide mutation in FvMYB10 may lead to the yellow fruit in Fragaria vesca. Mol. Breed. 2017, 37, 35. [Google Scholar] [CrossRef]
- Lin-Wang, K.; McGhie, T.K.; Wang, M.; Liu, Y.; Warren, B.; Storey, R.; Espley, R.V.; Allan, A.C. Engineering the anthocyanin regulatory complex of strawberry (Fragaria vesca). Front. Plant Sci. 2014, 5, 651. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Jiang, L.; Chen, Q.; Li, Y.; Zhang, Y.; Luo, Y.; Zhang, Y.; Sun, B.; Wang, X.; Tang, H. Comparative Transcriptome Profiling Analysis of Red- and White-Fleshed Strawberry (Fragaria × ananassa) Provides New Insight into the Regulation of the Anthocyanin Pathway. Plant Cell Physiol. 2018, 59, 1844–1859. [Google Scholar] [CrossRef] [PubMed]
- Medina-Puche, L.; Cumplido-Laso, G.; Amil-Ruiz, F.; Hoffmann, T.; Ring, L.; Rodrguez-Franco, A.; Caballero, J.L.; Schwab, W.; Muoz-Blanco, J.; Blanco-Portales, R. MYB10 plays a major role in the regulation of flavonoid/phenylpropanoid metabolism during ripening of Fragaria × ananassa fruits. J. Exp. Bot. 2014, 65, 401–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Telias, A.; Lin-Wang, K.; Stevenson, D.E.; Cooney, J.M.; Hellens, R.P.; Allan, A.C.; Hoover, E.E.; Bradeen, J.M. Apple skin patterning is associated with differential expression of MYB10. BMC Plant Biol. 2011, 11, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Zhang, X.; Zhao, Y.; Yang, J.; He, Y.; Li, G.; Ma, W.; Huang, X.; Su, J. Transcription factor PyHY5 binds to the promoters of PyWD40 and PyMYB10 and regulates its expression in red pear ‘Yunhongli No. 1’. Plant Physiol. Biochem. 2020, 154, 665–674. [Google Scholar] [CrossRef]
- Starkevič, P.; Paukstyte, J.; Kazanaviciute, V.; Denkovskiene, E.; Stanys, V.; Bendokas, V.; Siksnianas, T.; Razanskiene, A.; Ražanskas, R. Expression and Anthocyanin Biosynthesis-Modulating Potential of Sweet Cherry (Prunus avium L.) MYB10 and bHLH Genes. PLoS ONE 2015, 10, e0126991. [Google Scholar] [CrossRef]
- Zhou, H.; Liao, L.; Xu, S.; Ren, F.; Zhao, J.; Ogutu, C.; Wang, L.; Jiang, Q.; Han, Y. Two amino acid changes in the R3 repeat cause functional divergence of two clustered MYB10 genes in peach. Plant Mol. Biol. 2018, 98, 169–183. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, H.; Yang, Y.; Li, M.; Zhang, Y.; Liu, J.; Dong, J.; Li, J.; Butelli, E.; Xue, Z.; et al. The control of red colour by a family of MYB transcription factors in octoploid strawberry (Fragaria × ananassa ) fruits. Plant Biotechnol. J. 2019, 18, 1169–1184. [Google Scholar] [CrossRef] [Green Version]
- Härtl, K.; Denton, A.; Franz-Oberdorf, K.; Hoffmann, T.; Spornraft, M.; Usadel, B.; Schwab, W. Early metabolic and transcriptional variations in fruit of natural white-fruited Fragaria vesca genotypes. Sci. Rep. 2017, 7, srep45113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Symons, G.M.; Chua, Y.-J.; Ross, J.J.; Quittenden, L.; Davies, N.; Reid, J.B. Hormonal changes during non-climacteric ripening in strawberry. J. Exp. Bot. 2012, 63, 4741–4750. [Google Scholar] [CrossRef] [Green Version]
- Aharoni, A.; O’Connell, A.P. Gene expression analysis of strawberry achene and receptacle maturation using DNA microarrays. J. Exp. Bot. 2002, 53, 2073–2087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Sevilla, J.F.; Vallarino, J.G.; Osorio, S.; Bombarely, A.; Posé, D.; Merchante, C.; Botella, M.Á.; Amaya, I.; Valpuesta, V. Gene expression atlas of fruit ripening and transcriptome assembly from RNA-seq data in octoploid strawberry (Fragaria × ananassa). Sci. Rep. 2017, 7, 13737. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.P.; Suh, H.Y.; Kim, J.H.; Ahn, Y.K.; Kim, J.; Lee, E.S.; Kim, D.S.; Lee, H.E. Expression analysis of five Ara-bidopsis PDLP5 homologous in watermelon subjected to biotic and abiotic stresses. Hortic. Environ. Biotechnol. 2017, 58, 367–375. [Google Scholar] [CrossRef]
- Wang, S.; Basten, C.J.; Zeng, Z.B. Windows QTL Cartographer 2.5; Dep. of Statistics, North Carolina State University: Raleigh, NC, USA, 2012. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Chr. | Gene ID | Location | Ploidy Level |
|---|---|---|---|---|
| F. iinumae | 1 | evm.model.scaf_28.444 | 14,574,366–14,576,255 | Diploid |
| F. nilgerensis | 1 | evm.model.ctg92.252_scbg_v1.0 | 17,100,850–17,104,063 | Diploid |
| F. nubicola | 1 | evm.model.ctg173.372 | 15,072,390–15,075,138 | Diploid |
| F. viridis | 1 | evm.model.ctg108.134 | 13,877,866–13,880,523 | Diploid |
| F. vesca | 1 | FvH4_1g22020 | 13,950,290–13,952,507 | Diploid |
| F. × ananassa ‘Wongyo 3115’ | 1-1 | g00101712 | 13,223,169–13,223,636 | Octoploid |
| 1-2 | g00119641 | 12,622,806–12,626,007 | ||
| 1-3 | g00115370 | 13,537,353–13,539,011 | ||
| 1-4 | g00128511 | 13,235,156–13,238,279 | ||
| F. × ananassa ‘Camarosa’ | 1-1 | maker-Fvb1-1-snap-gene-139.18 | 13,899,904–13,911,034 | Octoploid |
| 1-2 | maker-Fvb1-2-snap-gene-157.15 | 15,722,359–15,726,641 | ||
| 1-3 | maker-Fvb1-3-augustus-gene-144.30 | 14,468,714–14,470,589 | ||
| 1-3 | maker-Fvb1-3-augustus-gene-143.29 | 14,315,382–14,315,713 |
| QTL | Chr. | Physical Position (Mbp) | Trait | Position (cM) | LOD | Additive Effect 1 | Dominant Effect 2 | R2 (%) |
|---|---|---|---|---|---|---|---|---|
| Col_1-2 | 1-2 | 12.1–24.5 | Flesh_color (2020) | 79.1–124.5 | 28.77 | 0.68 | −0.35 | 75.94 |
| Achene_color (2020) | 81.2–88.2 | 4.51 | 0.12 | −0.45 | 29.78 | |||
| Core_color (2020) | 82.2–142.9 | 32.02 | 0.41 | 0.30 | 38.48 | |||
| Achene_color (2020) | 89.2–111.5 | 4.40 | −0.17 | −0.43 | 10.00 | |||
| Skin_color (2019) | 97.5–142.9 | 7.19 | 0.23 | 0.18 | 12.91 | |||
| Skin_color (2020) | 97.5–142.9 | 11.49 | 0.45 | 0.23 | 20.19 | |||
| Col_1-4 | 1-4 | 11.8–15.9 | Achene_color (2020) | 53.4–66.6 | 5.67 | −0.22 | −0.10 | 10.04 |
| Col_2-1_1 | 2-1 | 18.8–20.8 | Flesh_color (2020) | 74.5–100.4 | 4.45 | 0.91 | −0.89 | 55.68 |
| Col_2-1_2 | 2-1 | 21.2–26.9 | Flesh_color (2020) | 30.7–47.3 | 5.52 | −0.28 | −0.07 | 6.06 |
| Col_6-4_1 | 6-4 | 3.0–6.9 | Achene_color (2019) | 7.0–17.9 | 3.97 | −0.03 | −0.25 | 5.67 |
| Col_6-4_2 | 6-4 | 15.5–29.7 | Skin_color (2019) | 127.7–143.7 | 5.00 | 0.30 | −0.25 | 0.62 |
| Achene_color (2020) | 138.7–144.2 | 4.17 | −0.26 | 0.11 | 15.09 | |||
| Col_7-4 | 7-4 | 17.8–21.8 | Achene_color (2020) | 58.5–68.1 | 4.49 | −0.18 | −0.05 | 6.50 |
| QTL | Gene ID | Chr. | Description | Function |
|---|---|---|---|---|
| Col_6-4_1 | g00015421 | 6-4 | DFR: Bifunctional dihydroflavonol 4-reductase/flavanone 4-reductase | Anthocyanin biosynthesis/Secondary metabolism |
| Col_6-4_2 | g00017208 | 6-4 | ANS1: Leucoanthocyanidin dioxygenase 1 | Anthocyanin biosynthesis/Secondary metabolism |
| Col_6-4_2 | g00018684 | 6-4 | C1: Anthocyanin regulatory C1 protein | Anthocyanin biosynthesis |
| Col_7-4 | g00137772 | 7-4 | CHI3: Probable chalcone--flavonone isomerase 3 | Anthocyanin biosynthesis/Secondary metabolism |
| Col_7-4 | g00138590 | 7-4 | GT1: Anthocyanidin 3-O-glucosyltransferase 1 | Anthocyanin biosynthesis/Secondary metabolism |
| Col_1-2 | g00119641 | 1-2 | MYB113: Transcription factor MYB113/MYB10 | DNA binding/Secondary metabolism |
| Col_1-4 | g00128511 | 1-4 | Similar to PNS1: Protein PNS1/MYB10 | DNA binding/Secondary metabolism |
| Col_7-4 | g00138251 | 7-4 | UGT88B1: UDP-glycosyltransferase 88B1 | |
| Col_6-4_2 | g00018719 | 6-4 | CYP750A1: Cytochrome P450 750A1 | ABA biosynthesis |
| Col_6-4_2 | g00019239 | 6-4 | GT6: UDP-glucose flavonoid 3-O-glucosyltransferase 6 | Anthocyanin biosynthesis/Secondary metabolism |
| Col_6-4_2 | g00018681 | 6-4 | GPI: Glucose-6-phosphate isomerase | Sugar metabolism |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manivannan, A.; Han, K.; Lee, S.Y.; Lee, H.-E.; Hong, J.P.; Kim, J.; Lee, Y.-R.; Lee, E.S.; Kim, D.-S. Genome-Wide Analysis of MYB10 Transcription Factor in Fragaria and Identification of QTLs Associated with Fruit Color in Octoploid Strawberry. Int. J. Mol. Sci. 2021, 22, 12587. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212587
Manivannan A, Han K, Lee SY, Lee H-E, Hong JP, Kim J, Lee Y-R, Lee ES, Kim D-S. Genome-Wide Analysis of MYB10 Transcription Factor in Fragaria and Identification of QTLs Associated with Fruit Color in Octoploid Strawberry. International Journal of Molecular Sciences. 2021; 22(22):12587. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212587
Chicago/Turabian StyleManivannan, Abinaya, Koeun Han, Sun Yi Lee, Hye-Eun Lee, Jong Pil Hong, Jinhee Kim, Ye-Rin Lee, Eun Su Lee, and Do-Sun Kim. 2021. "Genome-Wide Analysis of MYB10 Transcription Factor in Fragaria and Identification of QTLs Associated with Fruit Color in Octoploid Strawberry" International Journal of Molecular Sciences 22, no. 22: 12587. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212587