
Inhibitory Effects of Sulfonamide Derivatives on the β-Carbonic Anhydrase (MpaCA) from Malassezia pachydermatis, a Commensal, Pathogenic Fungus Present in Domestic Animals

 , ,
, ,  , , and
, , and 
Abstract
:1. Introduction
2. Results and Discussion
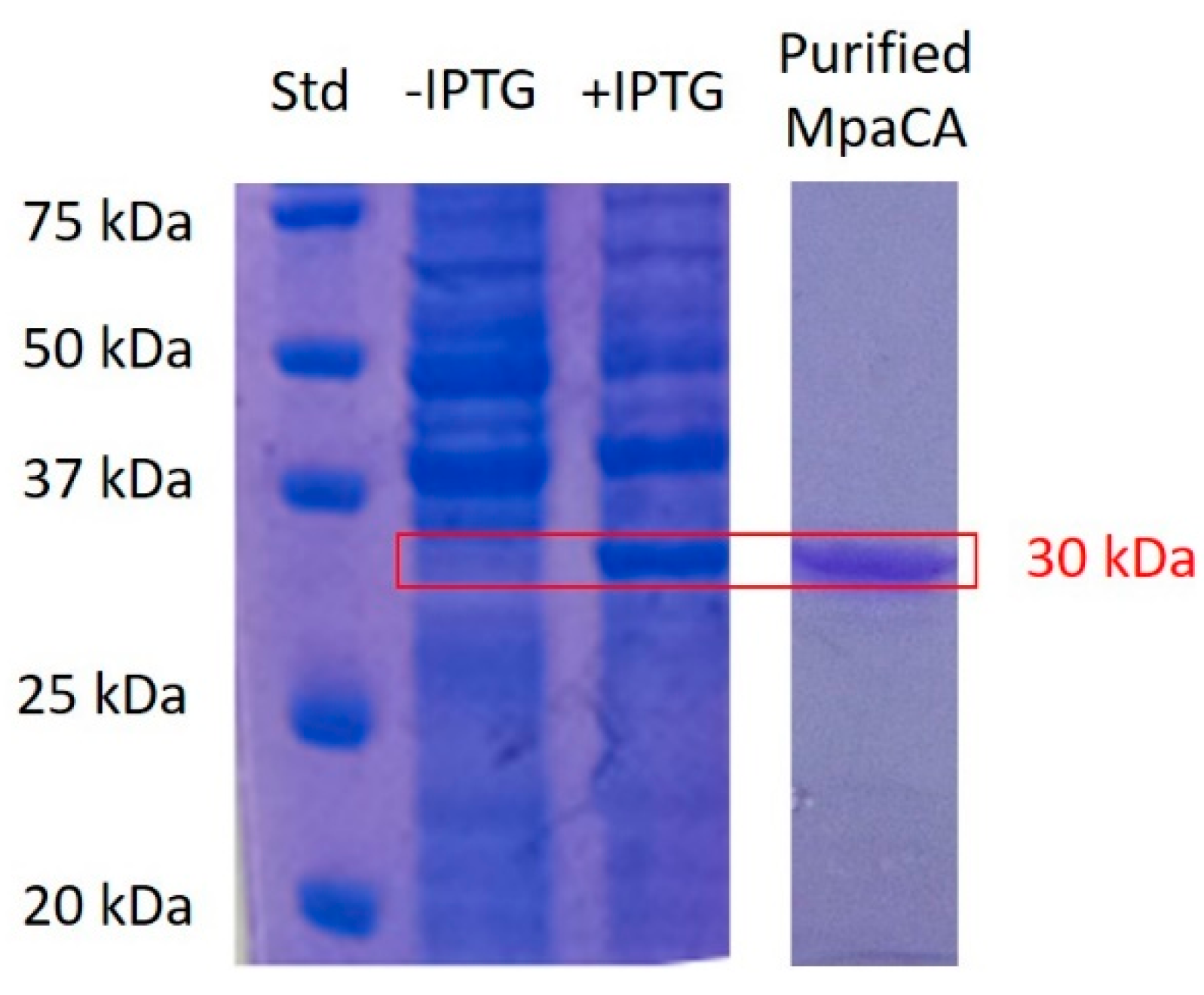
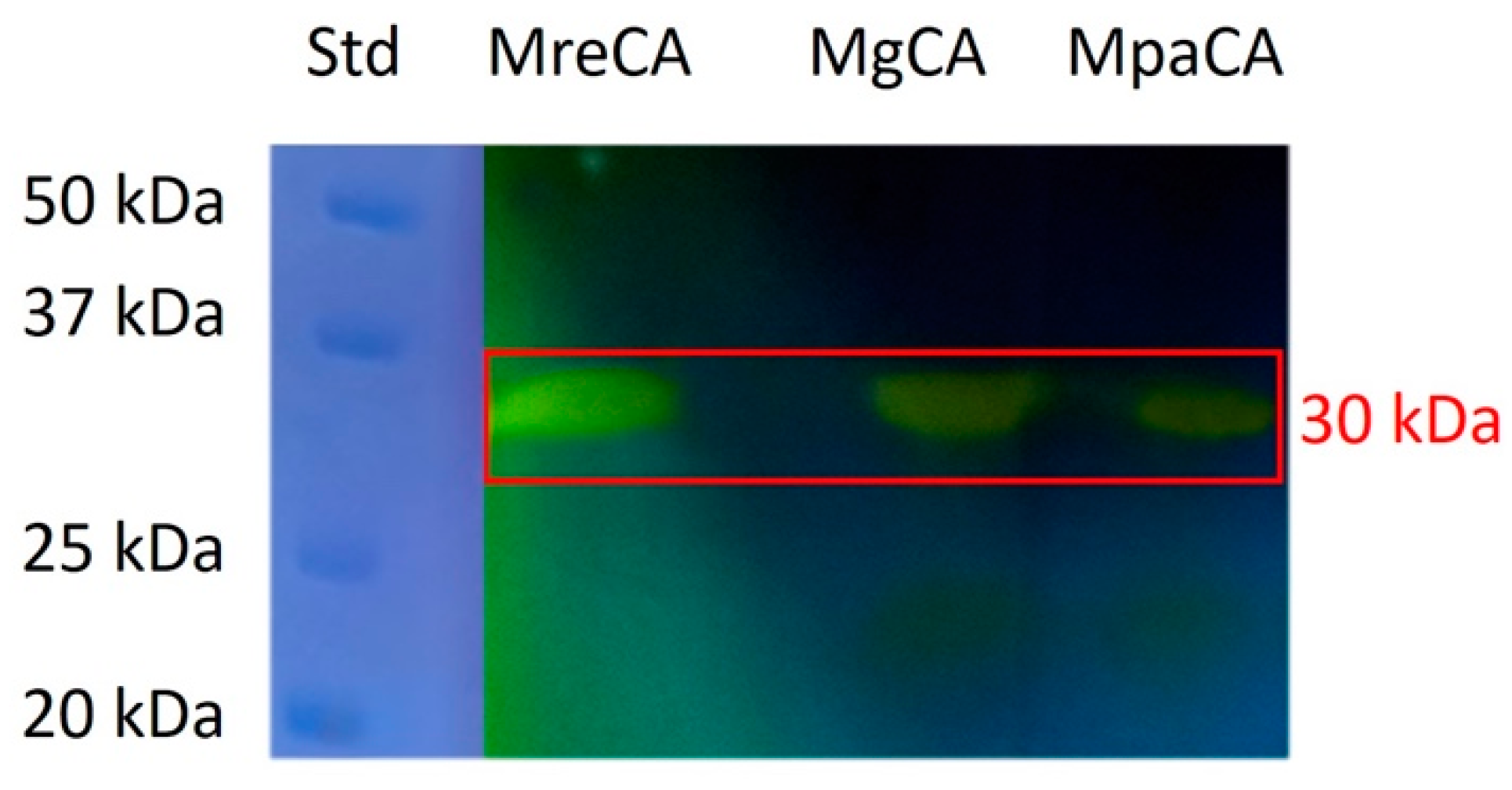
2.1. Biochemical Characterization of MpaCA
2.2. Inhibition Profile of MpaCA with Sulfonamides
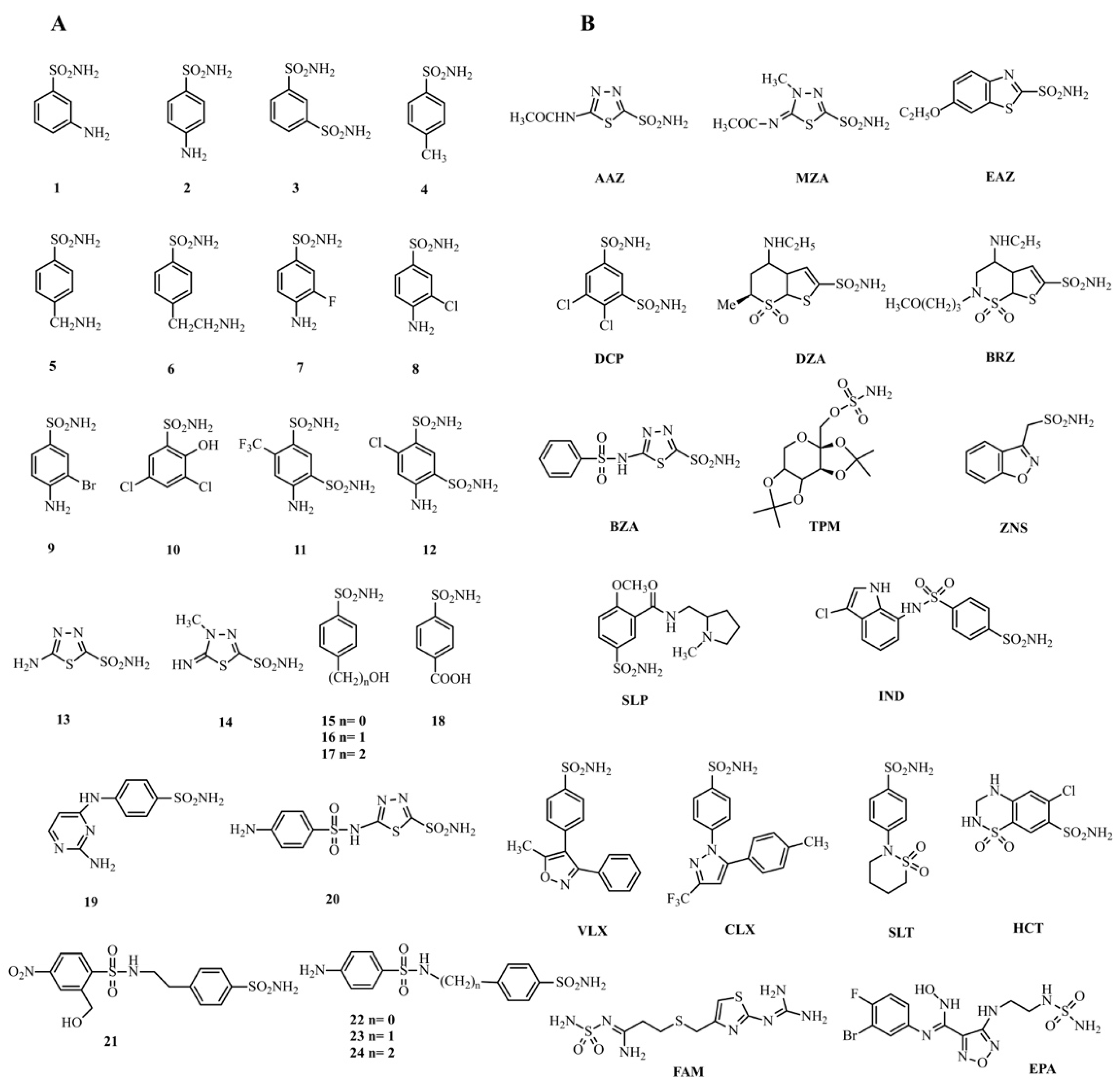
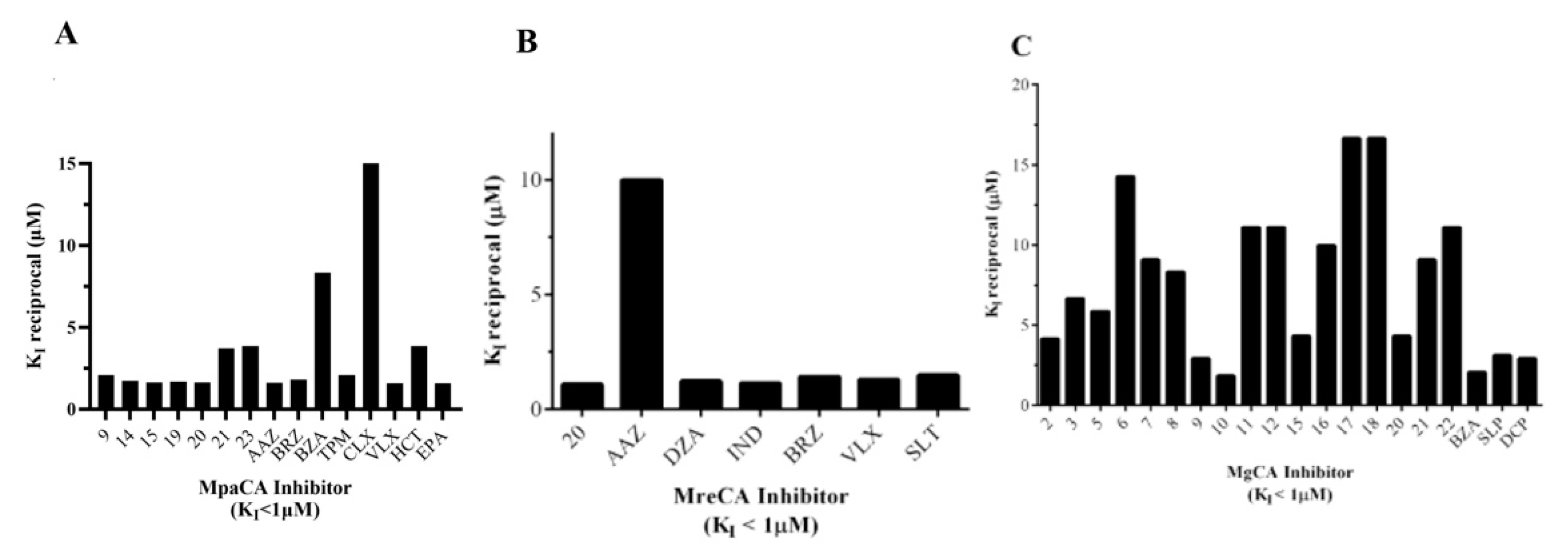
- Only fifteen drugs inhibited MpaCA with inhibition constant (KIs) values less than 1.0 μM. Sulfonamide inhibitors of the series 1–24, such as 9, 14, 15, 19, 20, 21, 23, and clinically used sulfonamide drugs of the series AAZ-EPA, such as AAZ, BRZ, BZA, TMP, CLX, VLX, HCT, and EPA, are significant examples. All these inhibitors showed KI values in the range 0.06–0.62 µM (Table 2 and Figure 5A). MreCA showed only seven “good inhibitors” (20, AAZ, DZA, BRZ, IND, VLX, and SLT) with KI values <1.0 µM (Table 2 and Figure 5B), while MgCA was well inhibited (KI values 0.06–0.54 µM) by the following twenty compounds: 2, 3, 5, 6, 7, 8, 9, 10, 11, 12, 15, 16, 17, 18, 20, 21, 22, BZA, SLP and DCP (Table 2 and Figure 5C).
- 2.
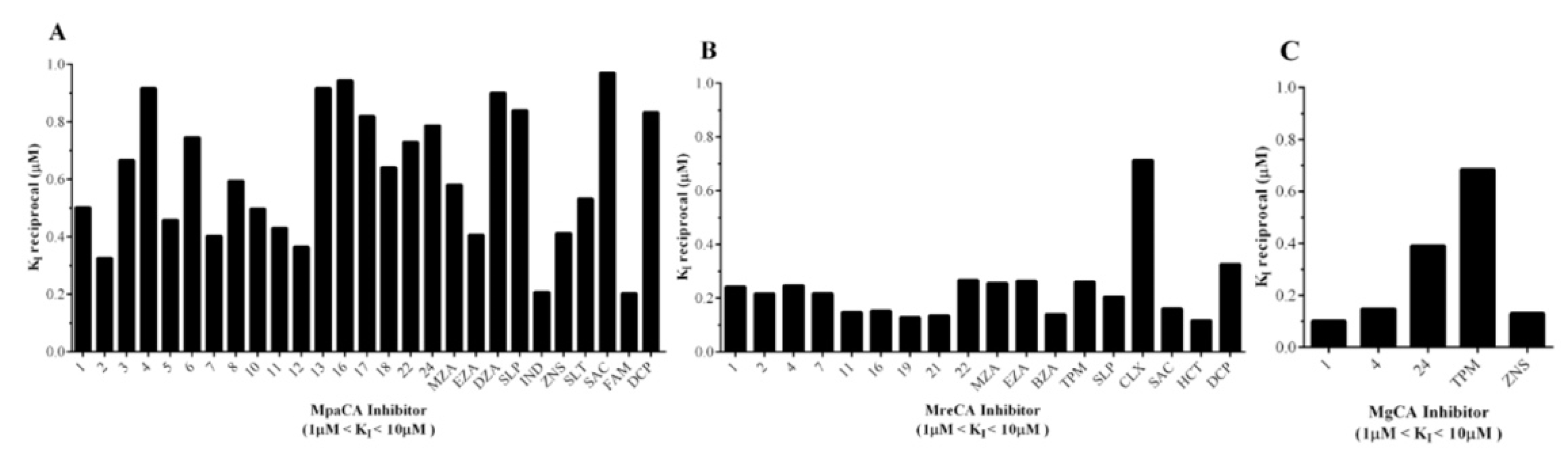
- Many compounds of the series 1–24 and AAZ-EPA (1, 2, 3, 4, 5, 6, 7, 8, 10, 11, 12, 13, 16, 17, 18, 22, 24, MZA, EZA, DZA, SLP, IND, ZNS, SLT, SAC, FAM, and DCP) examined on MpaCA demonstrated a moderate inhibitory effect on this enzyme, with KI values in the range 1.06–4.91 µM (Table 2 and Figure 6A). A number of these small molecules, namely 1, 4, 13, 24, MZA, EZA, ZNS, FAM, and SAC, were also weak inhibitors of MreCA and MgCA, showing KI values higher than 1.0 µM. Figure 6B,C provide a graphical representation of these findings, showing sulfonamide inhibitors with 1 µM < KIs < 10 µM for these fungal enzymes.
- 3.
- As mentioned above, many of the chemicals reported in Table 2 were weak inhibitors of MreCA (Ki > 10 µM) and were already demonstrated to be effective and moderate inhibitors of the human isoenzyme II (hCA II) [26], MpaCA and MgCA, respectively. As highlighted above, MreCA showed an inhibition pattern markedly different from those of the other two homologous enzymes MpaCA and MgCA (Table 2).
3. Materials and Methods
3.1. MpaCA Production: Synthetic Gene, Cloning, Heterologous Expression, and Purification
3.2. Enzyme Protonography
3.3. Enzyme Assays
3.4. Inhibition Assays
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Cummins, E.P.; Selfridge, A.C.; Sporn, P.H.; Sznajder, J.I.; Taylor, C.T. Carbon dioxide-sensing in organisms and its implications for human disease. Cell. Mol. Life Sci. 2014, 71, 831–845. [Google Scholar] [CrossRef] [Green Version]
- Endeward, V.; Musa-Aziz, R.; Cooper, G.J.; Chen, L.M.; Pelletier, M.F.; Virkki, L.V.; Supuran, C.T.; King, L.S.; Boron, W.F.; Gros, G. Evidence that aquaporin 1 is a major pathway for CO2 transport across the human erythrocyte membrane. FASEB J. 2006, 20, 1974–1981. [Google Scholar] [CrossRef] [Green Version]
- Swenson, E.R. Does Aerobic Respiration Produce Carbon Dioxide or Hydrogen Ion and Bicarbonate? Anesthesiology 2018, 128, 873–879. [Google Scholar] [CrossRef]
- Supuran, C.T.; Capasso, C. A Highlight on the Inhibition of Fungal Carbonic Anhydrases as Drug Targets for the Antifungal Armamentarium. Int. J. Mol. Sci. 2021, 22, 4324. [Google Scholar] [CrossRef]
- Angiolella, L.; Carradori, S.; Maccallini, C.; Giusiano, G.; Supuran, C.T. Targeting Malassezia species for Novel Synthetic and Natural Antidandruff Agents. Curr. Med. Chem. 2017, 24, 2392–2412. [Google Scholar] [CrossRef] [PubMed]
- Annunziato, G.; Angeli, A.; D’Alba, F.; Bruno, A.; Pieroni, M.; Vullo, D.; De Luca, V.; Capasso, C.; Supuran, C.T.; Costantino, G. Discovery of New Potential Anti-Infective Compounds Based on Carbonic Anhydrase Inhibitors by Rational Target-Focused Repurposing Approaches. ChemMedChem 2016, 11, 1904–1914. [Google Scholar] [CrossRef] [PubMed]
- Ozensoy Guler, O.; Capasso, C.; Supuran, C.T. A magnificent enzyme superfamily: Carbonic anhydrases, their purification and characterization. J. Enzyme Inhib. Med. Chem. 2016, 31, 689–694. [Google Scholar] [CrossRef]
- Del Prete, S.; Vullo, D.; De Luca, V.; Carginale, V.; Ferraroni, M.; Osman, S.M.; AlOthman, Z.; Supuran, C.T.; Capasso, C. Sulfonamide inhibition studies of the beta-carbonic anhydrase from the pathogenic bacterium Vibrio cholerae. Bioorg. Med. Chem. 2016, 24, 1115–1120. [Google Scholar] [CrossRef]
- Del Prete, S.; De Luca, V.; De Simone, G.; Supuran, C.T.; Capasso, C. Cloning, expression and purification of the complete domain of the eta-carbonic anhydrase from Plasmodium falciparum. J. Enzyme Inhib. Med. Chem. 2016, 31, 54–59. [Google Scholar] [CrossRef] [Green Version]
- Capasso, C.; Supuran, C.T. An Overview of the Carbonic Anhydrases from Two Pathogens of the Oral Cavity: Streptococcus mutans and Porphyromonas gingivalis. Curr. Top. Med. Chem. 2016, 16, 2359–2368. [Google Scholar] [CrossRef] [PubMed]
- Capasso, C.; Supuran, C.T. An overview of the alpha-, beta- and gamma-carbonic anhydrases from Bacteria: Can bacterial carbonic anhydrases shed new light on evolution of bacteria? J. Enzyme Inhib. Med. Chem. 2015, 30, 325–332. [Google Scholar] [CrossRef] [Green Version]
- Supuran, C.T.; Capasso, C. An Overview of the Bacterial Carbonic Anhydrases. Metabolites 2017, 7, 56. [Google Scholar] [CrossRef] [Green Version]
- Ohkuni, K.; Hayashi, M.; Yamashita, I. Bicarbonate-mediated social communication stimulates meiosis and sporulation of Saccharomyces cerevisiae. Yeast 1998, 14, 623–631. [Google Scholar] [CrossRef]
- D’Souza, C.A.; Heitman, J. Conserved cAMP signaling cascades regulate fungal development and virulence. FEMS Microbiol. Rev. 2001, 25, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Staniszewska, M. Virulence Factors in Candida species. Curr. Protein Pept. Sci. 2020, 21, 313–323. [Google Scholar] [CrossRef]
- Mogensen, E.G.; Janbon, G.; Chaloupka, J.; Steegborn, C.; Fu, M.S.; Moyrand, F.; Klengel, T.; Pearson, D.S.; Geeves, M.A.; Buck, J.; et al. Cryptococcus neoformans senses CO2 through the carbonic anhydrase Can2 and the adenylyl cyclase Cac1. Eukaryot. Cell 2006, 5, 103–111. [Google Scholar] [CrossRef] [Green Version]
- Schlicker, C.; Hall, R.A.; Vullo, D.; Middelhaufe, S.; Gertz, M.; Supuran, C.T.; Muhlschlegel, F.A.; Steegborn, C. Structure and inhibition of the CO2-sensing carbonic anhydrase Can2 from the pathogenic fungus Cryptococcus neoformans. J. Mol. Biol. 2009, 385, 1207–1220. [Google Scholar] [CrossRef] [PubMed]
- Klengel, T.; Liang, W.J.; Chaloupka, J.; Ruoff, C.; Schroppel, K.; Naglik, J.R.; Eckert, S.E.; Mogensen, E.G.; Haynes, K.; Tuite, M.F.; et al. Fungal adenylyl cyclase integrates CO2 sensing with cAMP signaling and virulence. Curr. Biol. 2005, 15, 2021–2026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, J.C.; Oude-Elferink, R.P. Role of the bicarbonate-responsive soluble adenylyl cyclase in pH sensing and metabolic regulation. Front. Physiol. 2014, 5, 42. [Google Scholar] [CrossRef] [Green Version]
- Hewitson, K.S.; Vullo, D.; Scozzafava, A.; Mastrolorenzo, A.; Supuran, C.T. Molecular cloning, characterization, and inhibition studies of a beta-carbonic anhydrase from Malassezia globosa, a potential antidandruff target. J. Med. Chem. 2012, 55, 3513–3520. [Google Scholar] [CrossRef]
- Rhimi, W.; Theelen, B.; Boekhout, T.; Otranto, D.; Cafarchia, C. Malassezia spp. Yeasts of Emerging Concern in Fungemia. Front. Cell. Infect. Microbiol. 2020, 10, 370. [Google Scholar] [CrossRef]
- Guillot, J.; Bond, R. Malassezia Yeasts in Veterinary Dermatology: An Updated Overview. Front. Cell. Infect. Microbiol. 2020, 10, 79. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.M.; Huang, W.M.; Li, S.F.; Wu, G.F.; Lai, K.; Chen, R.Y. Granulomatous skin infection caused by Malassezia pachydermatis in a dog owner. Arch. Dermatol. 2006, 142, 1181–1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillot, J.; Bond, R. Malassezia pachydermatis: A review. Med. Mycol. 1999, 37, 295–306. [Google Scholar] [CrossRef] [Green Version]
- Del Prete, S.; Vullo, D.; Ghobril, C.; Hitce, J.; Clavaud, C.; Marat, X.; Capasso, C.; Supuran, C.T. Cloning, Purification, and Characterization of a beta-Carbonic Anhydrase from Malassezia restricta, an Opportunistic Pathogen Involved in Dandruff and Seborrheic Dermatitis. Int. J. Mol. Sci. 2019, 20, 2447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Prete, S.; Angeli, A.; Ghobril, C.; Hitce, J.; Clavaud, C.; Marat, X.; Supuran, C.T.; Capasso, C. Sulfonamide Inhibition Profile of the beta-Carbonic Anhydrase from Malassezia restricta, An Opportunistic Pathogen Triggering Scalp Conditions. Metabolites 2020, 10, 39. [Google Scholar] [CrossRef] [Green Version]
- Del Prete, S.; Vullo, D.; Osman, S.M.; AlOthman, Z.; Capasso, C.; Supuran, C.T. Anion inhibition studies of the dandruff-producing fungus Malassezia globosa beta-carbonic anhydrase MgCA. Bioorg. Med. Chem. Lett. 2015, 25, 5194–5198. [Google Scholar] [CrossRef]
- Entezari Heravi, Y.; Bua, S.; Nocentini, A.; Del Prete, S.; Saboury, A.A.; Sereshti, H.; Capasso, C.; Gratteri, P.; Supuran, C.T. Inhibition of Malassezia globosa carbonic anhydrase with phenols. Bioorg. Med. Chem. 2017, 25, 2577–2582. [Google Scholar] [CrossRef]
- Nocentini, A.; Vullo, D.; Del Prete, S.; Osman, S.M.; Alasmary, F.A.S.; AlOthman, Z.; Capasso, C.; Carta, F.; Gratteri, P.; Supuran, C.T. Inhibition of the beta-carbonic anhydrase from the dandruff-producing fungus Malassezia globosa with monothiocarbamates. J. Enzyme Inhib. Med. Chem. 2017, 32, 1064–1070. [Google Scholar] [CrossRef] [Green Version]
- Vullo, D.; Del Prete, S.; Nocentini, A.; Osman, S.M.; AlOthman, Z.; Capasso, C.; Bozdag, M.; Carta, F.; Gratteri, P.; Supuran, C.T. Dithiocarbamates effectively inhibit the beta-carbonic anhydrase from the dandruff-producing fungus Malassezia globosa. Bioorg. Med. Chem. 2017, 25, 1260–1265. [Google Scholar] [CrossRef]
- Nocentini, A.; Bua, S.; Del Prete, S.; Heravi, Y.E.; Saboury, A.A.; Karioti, A.; Bilia, A.R.; Capasso, C.; Gratteri, P.; Supuran, C.T. Natural Polyphenols Selectively Inhibit beta-Carbonic Anhydrase from the Dandruff-Producing Fungus Malassezia globosa: Activity and Modeling Studies. ChemMedChem 2018, 13, 816–823. [Google Scholar] [CrossRef]
- Bua, S.; Osman, S.M.; AlOthman, Z.; Supuran, C.T.; Nocentini, A. Benzenesulfonamides incorporating nitrogenous bases show effective inhibition of beta-carbonic anhydrases from the pathogenic fungi Cryptococcus neoformans, Candida glabrata and Malassezia globosa. Bioorg. Chem. 2019, 86, 39–43. [Google Scholar] [CrossRef]
- Paloukopoulou, C.; Govari, S.; Soulioti, A.; Stefanis, I.; Angeli, A.; Matheeussen, A.; Capasso, C.; Cos, P.; Supuran, C.T.; Karioti, A. Phenols from Origanum dictamnus L. and Thymus vulgaris L. and their activity against Malassezia globosa carbonic anhydrase. Nat. Prod. Res. 2021, 35, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Del Prete, S.; De Luca, V.; Vullo, D.; Osman, S.M.; AlOthman, Z.; Carginale, V.; Supuran, C.T.; Capasso, C. A new procedure for the cloning, expression and purification of the beta-carbonic anhydrase from the pathogenic yeast Malassezia globosa, an anti-dandruff drug target. J. Enzyme Inhib. Med. Chem. 2016, 31, 1156–1161. [Google Scholar] [CrossRef] [PubMed]
- Kolaczek, A.; Fusiarz, I.; Lawecka, J.; Branowska, D. Biological activity and synthesis of sulfonamide derivatives: A brief review. Chemik 2014, 68, 625–628. [Google Scholar]
- Supuran, C.T.; Capasso, C. Antibacterial carbonic anhydrase inhibitors: An update on the recent literature. Expert Opin. Ther. Pat. 2020, 30, 963–982. [Google Scholar] [CrossRef]
- Supuran, C.T. Emerging role of carbonic anhydrase inhibitors. Clin. Sci. 2021, 135, 1233–1249. [Google Scholar] [CrossRef]
- Angeli, A.; Carta, F.; Nocentini, A.; Winum, J.Y.; Zalubovskis, R.; Onnis, V.; Eldehna, W.M.; Capasso, C.; Carradori, S.; Donald, W.A.; et al. Response to Perspectives on the Classical Enzyme Carbonic Anhydrase and the Search for Inhibitors. Biophys. J. 2021, 120, 178–181. [Google Scholar] [CrossRef]
- Ponka, D. Approach to managing patients with sulfa allergy—Use of antibiotic and nonantibiotic sulfonamides. Can. Fam. Physician 2006, 52, 1434–1438. [Google Scholar]
- Keilin, D.; Mann, T. Carbonic anhydrase. Purification and nature of the enzyme. Biochem. J. 1940, 34, 1163–1176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krebs, H.A. Inhibition of carbonic anhydrase by sulphonamides. Biochem. J. 1948, 43, 525–528. [Google Scholar] [CrossRef] [Green Version]
- Shaldam, M.; Eldehna, W.M.; Nocentini, A.; Elsayed, Z.M.; Ibrahim, T.M.; Salem, R.; El-Domany, R.A.; Capasso, C.; Abdel-Aziz, H.A.; Supuran, C.T. Development of novel benzofuran-based SLC-0111 analogs as selective cancer-associated carbonic anhydrase isoform IX inhibitors. Eur. J. Med. Chem. 2021, 216, 113283. [Google Scholar] [CrossRef] [PubMed]
- Capasso, C.; Supuran, C.T. An Overview of the Selectivity and Efficiency of the Bacterial Carbonic Anhydrase Inhibitors. Curr. Med. Chem. 2015, 22, 2130–2139. [Google Scholar] [CrossRef]
- Angeli, A.; Kartsev, V.; Petrou, A.; Pinteala, M.; Vydzhak, R.M.; Panchishin, S.Y.; Brovarets, V.; De Luca, V.; Capasso, C.; Geronikaki, A.; et al. New Sulfanilamide Derivatives Incorporating Heterocyclic Carboxamide Moieties as Carbonic Anhydrase Inhibitors. Pharmaceuticals 2021, 14, 828. [Google Scholar] [CrossRef] [PubMed]
- Vullo, D.; Leewattanapasuk, W.; Muhlschlegel, F.A.; Mastrolorenzo, A.; Capasso, C.; Supuran, C.T. Carbonic anhydrase inhibitors: Inhibition of the beta-class enzyme from the pathogenic yeast Candida glabrata with sulfonamides, sulfamates and sulfamides. Bioorg. Med. Chem. Lett. 2013, 23, 2647–2652. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- De Luca, V.; Del Prete, S.; Supuran, C.T.; Capasso, C. Protonography, a new technique for the analysis of carbonic anhydrase activity. J. Enzyme Inhib. Med. Chem. 2015, 30, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Khalifah, R.G. The carbon dioxide hydration activity of carbonic anhydrase. I. Stop-flow kinetic studies on the native human isoenzymes B and C. J. Biol. Chem. 1971, 246, 2561–2573. [Google Scholar] [CrossRef]
- Carta, F.; Temperini, C.; Innocenti, A.; Scozzafava, A.; Kaila, K.; Supuran, C.T. Polyamines inhibit carbonic anhydrases by anchoring to the zinc-coordinated water molecule. J. Med. Chem. 2010, 53, 5511–5522. [Google Scholar] [CrossRef] [Green Version]
- Nocentini, A.; Angeli, A.; Carta, F.; Winum, J.Y.; Zalubovskis, R.; Carradori, S.; Capasso, C.; Donald, W.A.; Supuran, C.T. Reconsidering anion inhibitors in the general context of drug design studies of modulators of activity of the classical enzyme carbonic anhydrase. J. Enzyme Inhib. Med. Chem. 2021, 36, 561–580. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CAI | Commercial Name |
|---|---|
| AAZ | Acetazolamide |
| MZA | Methazolamide |
| EZA | Ethoxzolamide |
| DZA | Dorzolamide |
| BRZ | Brinzolamide |
| BZA | Benzolamide |
| TPM | Topiramate |
| SLP | Sulpiride |
| IND | Indisulam E7070 |
| ZNS | Zonisamide |
| CLX | Celecoxib |
| VLX | Valdecoxib |
| SLT | Sulthiame |
| SAC | Saccharin |
| HCT | Hydrochlorothiazide |
| FAM | Famotidine |
| DCP | Dichlorophenamide |
| EPA | Epacadostat |
| KI (µM) * | |||
|---|---|---|---|
| Inhibitor | MpaCA | a MreCA | a MgCA |
| 1 | 1.99 | 4.12 | 9.8 |
| 2 | 3.07 | 4.62 | 0.24 |
| 3 | 1.5 | >10 | 0.15 |
| 4 | 1.09 | 4.04 | 6.74 |
| 5 | 2.18 | >10 | 0.17 |
| 6 | 1.34 | >10 | 0.07 |
| 7 | 2.48 | 4.59 | 0.11 |
| 8 | 1.68 | >10 | 0.12 |
| 9 | 0.48 | >10 | 0.34 |
| 10 | 2.01 | >10 | 0.54 |
| 11 | 2.32 | 6.76 | 0.09 |
| 12 | 2.74 | >10 | 0.09 |
| 13 | 1.09 | >10 | >10 |
| 14 | 0.58 | >10 | >10 |
| 15 | 0.61 | >10 | 0.23 |
| 16 | 1.06 | 6.51 | 0.10 |
| 17 | 1.22 | >10 | 0.06 |
| 18 | 1.56 | >10 | 0.06 |
| 19 | 0.59 | 7.79 | >10 |
| 20 | 0.61 | 0.91 | 0.23 |
| 21 | 0.27 | 7.4 | 0.11 |
| 22 | 1.37 | 3.74 | 0.09 |
| 23 | 0.26 | >10 | >10 |
| 24 | 1.27 | >10 | 2.56 |
| AAZ | 0.62 | 0.1 | >10 |
| MZA | 1.72 | 3.9 | >10 |
| EZA | 2.46 | 3.79 | >10 |
| DZA | 1.11 | 0.81 | >10 |
| BRZ | 0.55 | 0.7 | >10 |
| BZA | 0.12 | 7.15 | 0.48 |
| TPM | 0.48 | 3.83 | 1.46 |
| SLP | 1.19 | 4.85 | 0.32 |
| IND | 4.82 | 0.87. | n.d. |
| ZNS | 2.42 | >10 | 7.65 |
| CLX | 0.06 | 1.4 | >10 |
| VLX | 0.63 | 0.77 | >10 |
| SLT | 1.88 | 0.67 | n.d. |
| SAC | 1.03 | 6.2 | n.d. |
| HCT | 0.26 | 8.5 | n.d. |
| FAM | 4.91 | >10 | n.d. |
| DCP | 1.20 | 3.06 | 0.34 |
| EPA | 0.63 | n.d. | n.d. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Luca, V.; Angeli, A.; Mazzone, V.; Adelfio, C.; Carta, F.; Selleri, S.; Carginale, V.; Scaloni, A.; Supuran, C.T.; Capasso, C. Inhibitory Effects of Sulfonamide Derivatives on the β-Carbonic Anhydrase (MpaCA) from Malassezia pachydermatis, a Commensal, Pathogenic Fungus Present in Domestic Animals. Int. J. Mol. Sci. 2021, 22, 12601. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212601
De Luca V, Angeli A, Mazzone V, Adelfio C, Carta F, Selleri S, Carginale V, Scaloni A, Supuran CT, Capasso C. Inhibitory Effects of Sulfonamide Derivatives on the β-Carbonic Anhydrase (MpaCA) from Malassezia pachydermatis, a Commensal, Pathogenic Fungus Present in Domestic Animals. International Journal of Molecular Sciences. 2021; 22(22):12601. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212601
Chicago/Turabian StyleDe Luca, Viviana, Andrea Angeli, Valeria Mazzone, Claudia Adelfio, Fabrizio Carta, Silvia Selleri, Vincenzo Carginale, Andrea Scaloni, Claudiu T. Supuran, and Clemente Capasso. 2021. "Inhibitory Effects of Sulfonamide Derivatives on the β-Carbonic Anhydrase (MpaCA) from Malassezia pachydermatis, a Commensal, Pathogenic Fungus Present in Domestic Animals" International Journal of Molecular Sciences 22, no. 22: 12601. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222212601