
Overview of Drug Transporters in Human Placenta


1
Department of Obstetrics and Gynecology, Graduate School of Medicine, Osaka University, Osaka 5650871, Japan
2
Placenta Lab, Department of Obstetrics, Jena University Hospital, Am Klinikum 1, 07747 Jena, Germany
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2021, 22(23), 13149; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222313149
Submission received: 30 October 2021
/
Revised: 25 November 2021
/
Accepted: 30 November 2021
/
Published: 5 December 2021
(This article belongs to the Special Issue Reproductive and Developmental Toxicology 2.0)
Abstract
:The transport of drugs across the placenta is a point of great importance in pharmacotherapy during pregnancy. However, the knowledge of drug transport in pregnancy is mostly based on experimental clinical data, and the underlying biological mechanisms are not fully understood. In this review, we summarize the current knowledge of drug transporters in the human placenta. We only refer to human data since the placenta demonstrates great diversity among species. In addition, we describe the experimental models that have been used in human placental transport studies and discuss their availability. A better understanding of placental drug transporters will be beneficial for the health of pregnant women who need drug treatment and their fetuses.
1. Introduction
Transporters are membrane proteins that regulate the absorption, distribution, and excretion of substrates in all cells of the body. Transporters contribute to the creation of density gradients inside of and outside of membranes by actively importing and exporting a broad variety of substances. Although transporters were originally classified based on their functions, they are now classified according to the sequence homology of the isolated genes. Transporters are categorized into two superfamilies: ATP-binding cassette (ABC) transporters, which are involved in the efflux of substances, and solute carrier (SLC) transporters, which are mainly involved in the uptake of substances [1,2]. Transporters are involved in the transport of various physiological substances such as sugars, amino acids, and lipids, but some transporters are particularly relevant to pharmacokinetics and are thus recognized as drug transporters.
Drug transporters have two main roles: one is to control the uptake of drugs during their circulation from the organs that associated with alimentation, such as the small intestine, and their transport in excretory organs, such as the liver and kidneys. Their other role is to control local drug concentrations in order to protect critical cells, tissues, and organs from xenobiotics, such as at the blood–brain barrier, blood–cerebrospinal fluid barrier, and blood–placental barrier. The body is protected biologically from xenobiotics through the coexistence of ABC transporters and SLC transporters in each organ.
The placenta connects the fetus to the mother through the umbilical cord and supplies the fetus with nutrients and oxygen. Due to a history of harmful side effects on the fetus, the placental transport of drugs has always been an important issue. Regarding detoxification from xenobiotics, metabolic enzymes are involved [3], but transporters may have similar importance. This may be due to immature detoxification functions in the developing fetus or to an insufficiency of xenobiotic detoxifying capacities in the placental metabolic enzymes.
Thus, in this review, we summarize published data on human placental drug transporters and discuss the availability of experimental models. Animal data are not mentioned in this review because the placenta is an organ with great diversity among species not only regarding its construction but also its transporter activity [4,5].
2. Literature Review Procedure
First, we identified publications that discussed human placenta transporters that were published before May 2021 using the search term “transporters in human placenta” in PubMed. From those, we selected transporters that are able to recognize drugs as their substrates and that included data from humans and animals. These selected “drug transporters existing in the human placenta” are listed in Table 1. In the last step, we collected detailed information on validation in human cells or tissues. In the table, we have noted when a transporter has no substrate information available from human cells or tissues.
In the text and the table, we use the transporter acronyms that were used in the underlying original papers. Additionally, we provide the official gene name as per human gene nomenclature (HGNC).
3. Physiological Functions of Drug Transporters
Drug transporters have other functions and transport drugs because of their similarities with physiological substrates. Therefore, we provide a brief overview of the physiological functions and main expression sites (apart from the placenta) of the drug transporters that are described in this review.
3.1. ABC Transporters
The ABC transporter superfamily is a group of transporters that discharge drugs from the inside of cells to the outside of cells using ATP hydrolysis energy. Although they were originally identified as a multidrug resistance factor from tumor cells, they are also expressed in normal tissues and are able to transport a variety of drugs with different chemical structures and mechanisms of action (e.g., anticancer drugs, immunosuppressants, and inotropic glycosides). ABC transporters can be classified into seven subgroups, ranging from ABCA to G. Among these, ABCB, C, and G are recognized as drug transporters. The ABC transporter family has been studied more in depth than the SLC family [6]. The functional proteins of eukaryotic ABC transporters typically contain two nucleotide-binding domains (NBDs) and two transmembrane domains (TMDs). Eukaryotic ABC transporters are classified into two types: full transporters (ABCB1, ABCB4, and ABCC) and half transporters, which form either homodimers or heterodimers to constitute a functional transporter (ABCG). The genes of ABC transporters are highly conserved between species, indicating that most of these genes have existed since the beginning of eukaryotic evolution.
3.1.1. ABCB (Multidrug Resistance: MDR) Family
ABCB1 (MDR1), which was initially called P-glycoprotein (P-gp), is expressed in the canalicular membrane of hepatocytes, renal proximal tubules, the luminal side membranes of the capillary bile ducts in the liver, the apical surface of intestinal enterocytes, especially in the distal ileum, and in the capillary endothelial cells in the brain and testis. MDR1 recognizes a wide range of lipophilic substrates. In the gastrointestinal mucosa, MDR1 drains substrates to the luminal side, and in cerebrovascular endothelial cells, MDR1 regulates the substrate distribution into the brain tissue [6,7,8,9]. ABCB4 (MDR3) acts as a flippase for phosphatidylcholine, a phospholipid that forms cell membranes, and changes the composition of the phospholipids that are present in the cell membranes. In the liver, MDR3 secretes the phosphatidylcholine from the hepatocytes into bile, and an MDR3 deficiency leads progressive familial intrahepatic cholestasis (PFIC) type 3 [10,11].
3.1.2. ABCC (Multidrug-Resistance-Associated Protein: MRP) Family
ABCC1 (MRP1) is widely expressed throughout the body and is suggested to be involved in antioxidant activity [12,13]. ABCC2 (MRP2) is predominantly present in the liver and gallbladder, and its loss of function causes Dubin–Johnson syndrome [14]. ABCC3 (MRP3) is widely present throughout the body. In the placenta, MRP3 may be responsible for the transport of organic cholephilic anions [15]. ABCC4 (MRP4) is mainly expressed in the prostate and in the proximal tubules in the kidney and excretes prostaglandin [16]; ABCC5 (MRP5) is expressed throughout the body and excretes cyclic nucleotides to the outside of cells [17]; and ABCC6 (MRP6) is present in the liver and kidneys. MRP6 deficiency is the cause of “pseudoxanthoma elasticum”, which promotes ectopic calcification [18]. ABCC11 (MRP8) is predominantly expressed in the liver, testis, and prostate.
3.1.3. ABCG2 (Breast Cancer Resistant Protein: BCRP)
ABCG2 (BCRP) transports uric acid and a wide variety of organic anions into the gastrointestinal tract, bile, and urine [19].
3.2. SLC Transporters
This group of transporters is responsible for secondary active transport and some passive transport. A large number of SLC transporters have been identified, with more than 400 transporters in over 60 families. Unlike ABC transporters, SLC transporters are highly diverse in structure and consist of a variety of folds. The information on the phylogenetic analysis of SLC transporters is also scarce due to a large number of subfamilies and their complex genetic structures [20,21].
3.2.1. SLCO (Organic Anion Transporting Polypeptide: OATP) Family
SLCO1A2 (OATP1A2) is mainly expressed in the brain and transports thyroid hormone. SLCO 1B1 (OATP1B1) is responsible for the uptake of bile acids into the hepatocytes. SLCO2B1 (OATP2B1) is widely expressed in organs throughout the body and carries estrone 3 sulfate and dehydroepiandrosterone sulfate. SLCO3A1 (OATP3A1) is upregulated in the liver during cholestasis and contributes to the elimination of the bile acids that accumulate in the liver [22]. SLCO4A1 (OATP4A1) is widely present throughout the body and transports bile acids and thyroid hormones [23].
3.2.2. SLC22A Family
This family includes three subfamilies: organic cation transporter (OCT), organic cation transporter novel type (OCTN), and organic anion transporter (OAT).
SLC22A1 (OCT1) is mainly expressed in the liver and is responsible for the uptake of dopamine and other substances into the liver. In the placenta, OCT1 transports acetylcholine as a substrate [24]. SLC22A2 (OCT2) is responsible for creatinine transport (efflux) in the kidney [25]. SLC22A3 (OCT3), also called extraneuronal monoamine transporter (EMT), transports monoamine neurotransmitters such as serotonin [26], norepinephrine, and dopamine and is present in more diverse tissues than OCT1 and OCT2.
SLC22A4(OCTN1) and SLC22A5 (OCTN2) are physiologically crucial transporters for L-carnitine. It has been proposed that OCTN2 plays a major role in L-carnitine transport from the mother to the fetus across the placenta [27]. SLC22A6 (OAT1) is highly expressed in the kidney and is responsible for the first step in the uptake of PGs and other substances from the blood into cells and their elimination into urine [28]. SLC22A11 (OAT4) is mainly found in kidney and placenta [28]. It may act to prevent potentially harmful organic anions from reaching the fetus. SLC22A13 (OAT10) is highly expressed in kidney and is responsible for uric acid transport. Decreased OAT10 function increases the risk of gout [29].
3.2.3. SLC29A (Equilibrative Nucleotide Transporter: ENT) Family
Both SLC29A1 (ENT1) and SLC29A2 (ENT2) are expressed in a wide range of organs, while SLC29A4 (ENT4) is highly expressed in the adipose tissue. ENT1 and ENT2 are involved in the uptake of nucleic acids into the cells [30].
3.2.4. SLC47A (Multidrug and Toxin Extrusion: MATE) Family
SLC47A1(MATE1) is present in a wide variety of organs, including the liver and kidney, while SLC47A2 (MATE2) is mainly expressed in the kidney. They excrete substrates into the urine and bile through the exchange transport of H+ and organic cations [31].
4. Morphology of the Human Placental Barrier
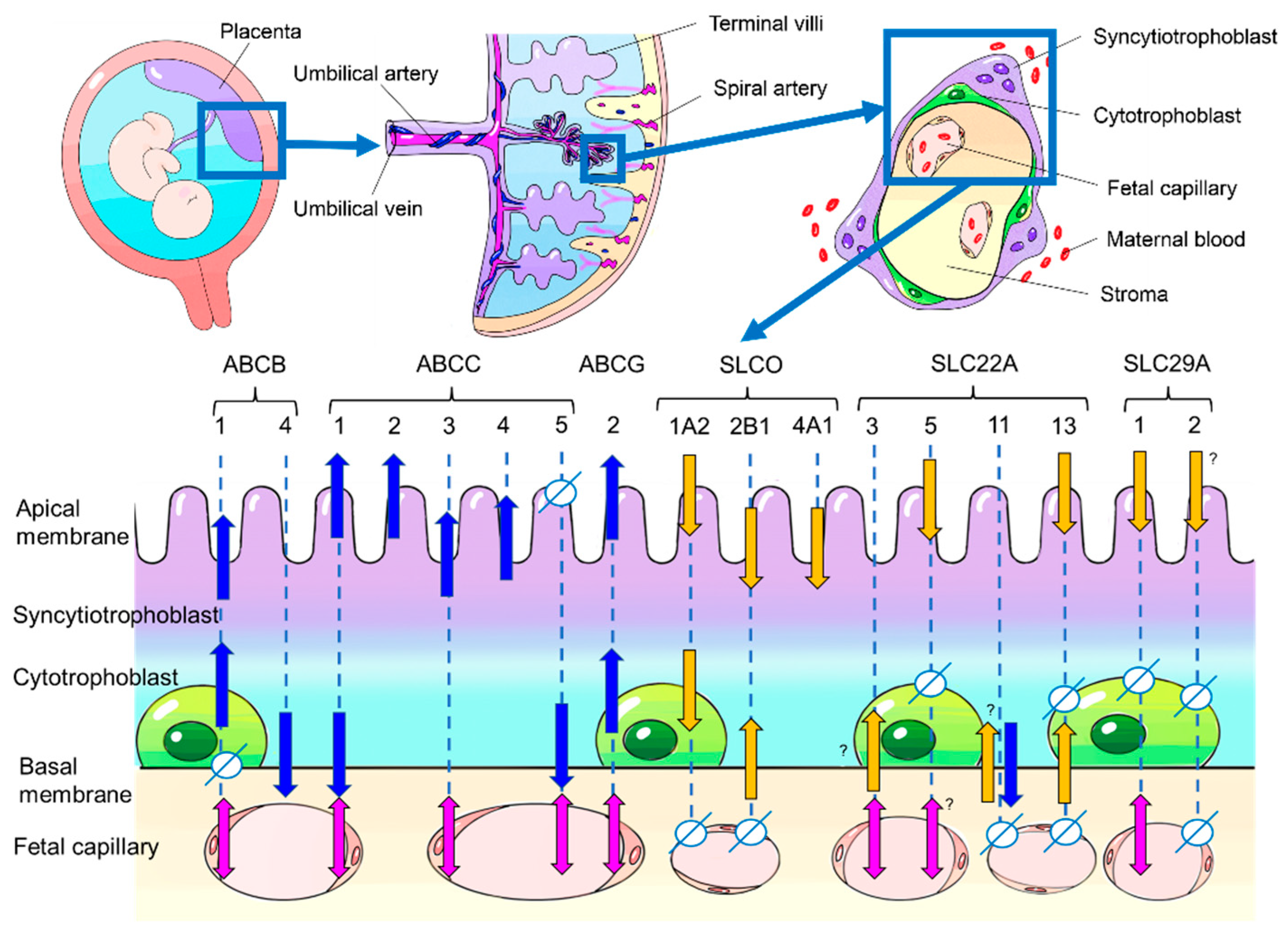
The human placenta has a hemochorial structure, where fetal-derived villous trees are surrounded by maternal blood. The cellular layers of villi consist of, from the maternal blood side, syncytiotrophoblast, cytotrophoblast cells, stroma cells, and fetal vascular endothelium, which act to separate the fetal circulation from the maternal circulation (Figure 1). Several mechanisms are involved in the transport of substances across the placenta, from simple diffusion to selective transport by membrane transporters.
5. Drug Transporters Reported in Human Placenta
In the human placenta, 28 proteins have been reported to function as drug transporters. For each transporter, we show its localization in the placenta and its expression in each trimester of pregnancy and in the cell lines that are frequently used in transporter studies (Table 1) (Figure 1).

{kind=link}
Table 1.
Detailed information on drug transporters in the human placenta. For each transporter, the following information is given: name (both original and HGNC), physiological substrates, drug substrates, localization in human placenta, change in expression level with gestational week, and expression in representative cell lines. Conflicting reports on the localization of transporters are indicated with an asterisk (*). Although the transporters are expressed in the human placenta, some functional data are only available from animals (indicated by **) or from human cell lines or organs other than the placenta (listed in parentheses after drug names). For OATP4A1, there is no substrate whose uptake has been clearly evaluated. However, it is generally believed to be involved in drug transport, as is the case with other OATPs. Abbreviations: NA: no data available, ND: not detected, R: RNA was detected, P: protein was detected, Sy: syncytiotrophoblast, Cy: cytotrophoblast, FE: fetal endothelium, LTC4: leukotriene C4, E217bG: 22C10–3 estradiol 17b D glucuronide, PG: prostaglandin, “+”: respective cells or tissues are positive.
Table 1.
Detailed information on drug transporters in the human placenta. For each transporter, the following information is given: name (both original and HGNC), physiological substrates, drug substrates, localization in human placenta, change in expression level with gestational week, and expression in representative cell lines. Conflicting reports on the localization of transporters are indicated with an asterisk (*). Although the transporters are expressed in the human placenta, some functional data are only available from animals (indicated by **) or from human cell lines or organs other than the placenta (listed in parentheses after drug names). For OATP4A1, there is no substrate whose uptake has been clearly evaluated. However, it is generally believed to be involved in drug transport, as is the case with other OATPs. Abbreviations: NA: no data available, ND: not detected, R: RNA was detected, P: protein was detected, Sy: syncytiotrophoblast, Cy: cytotrophoblast, FE: fetal endothelium, LTC4: leukotriene C4, E217bG: 22C10–3 estradiol 17b D glucuronide, PG: prostaglandin, “+”: respective cells or tissues are positive.
| Superfamily | Family | Transporter Name | HGNC Name | Physiological Substrates | Drug Substrates | Localization in Placenta | Period (Trimester) | Cell Lines | References | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sy | Cy | FE | 1st | 2nd | 3rd | Unknown | BeWo | JEG3 | JAR | HTR | |||||||
| ABC | ABCB | MDR1 | ABCB1 | hydrophobic compounds | vinblastine, vincristine, digoxin, saquinavir | + | + | + | R, P | R, P | R, P | + | + | + | + | [15,32,33,34,35,36,37,38,39,40] | |
| MDR3 | ABCB4 | bile acids | vinblastine, digoxin | + | NA | NA | R | NA | R | + | + | + | NA | [11,15,33,37] | |||
| ABCC | MRP1 | ABCC1 | bile acids, folic acid, LTC4, E217bG, | maraviroc, pravastatin | + | ND | + | R | NA | R, P | + | + | + | + | [37,41,42,43,44,45,46] | ||
| MRP2 | ABCC2 | organic anion | talinolol | + | NA | NA | R | R, P | R, P | + | +(P) | + | NA | [15,42,44,45,47,48,49,50,51] | |||
| MRP3 | ABCC3 | cholate | methotrexate (HEK293) | + | ND | + | R | NA | R, P | + | + | + | NA | [15,42,44,45,48,52] | |||
| MRP4 | ABCC4 | estradiol, cAMP, cGMP, | adefovir (kidney) | + | NA | NA | NA | NA | R, P | + | + | + | NA | [15,48,53] | |||
| MRP5 | ABCC5 | cAMP, cGMP | doxorubicin (nonsmallcell lungcancer cell-lines) | + | ND | + | R | NA | R, P | + | NA | NA | NA | [44,45,54] | |||
| MRP6 | ABCC6 | LTC4 | etoposide, doxorubicin, BQ−123 ** | NA | NA | NA | NA | NA | NA | R | NA | NA | NA | NA | [18,55,56] | ||
| MRP8 | ABCC11 | cGMP, cAMP | maraviroc | NA | NA | NA | NA | NA | R | + | + | + | NA | [15,57,58,59,60] | |||
| ABCG | BCRP | ABCG2 | organic anion | pravastatin, nitrofurantoin | + | + | + | R, P | R, P | R, P | +(P) | +(P) | + | NA | [15,35,37,38,40,50,51,61,62,63,64] | ||
| SLC | SLCO | OATP1A2 | SLCO1A2 | unconjugated bilirubin, steroids, thyroid hormones | maraviroc | + | + | ND | R, P | NA | R, P | + | + | + | NA | [11,15,41,65] | |
| OATP1B1 | SLCO1B1 | estradiol, taurocholate, leukotrienes, steroids, thyroid hormones | rifampicin (kidney), pravastatin (HEK293) | NA | NA | NA | R | NA | ND | ND | ND | ND | NA | [11,15,66,67] | |||
| OATP2B1 | SLCO2B1 | estrone 3-sulfate | fexofenadine | + | + | NA | NA | NA | R | + | + | + | NA | [11,15,68,69] | |||
| OATP3A1 | SLCO3A1 | vasopressin, PG, thyroid hormones | simvastatin (HEK293) | NA | NA | NA | R | NA | R | NA | NA | NA | NA | [11,68,70] | |||
| OATP4A1 | SLCO4A1 | taurocholate, PG | ** | + | ND | ND | R, P | NA | R, P | NA | NA | NA | NA | [11,65,71] | |||
| SLC22A | OCT1 | SLC22A1 | choline, dopamine | metformin (HEK293), pazopanib(hepatocytes), ranitidine (HEK293) | NA | NA | NA | NA | NA | R | + | + | NA | NA | [24,27,28,72,73,74,75,76] | ||
| OCT2 | SLC22A2 | histamine, dopamine, | metformin (HEK293) | NA | NA | NA | NA | NA | R | ND | ± | NA | NA | [24,27,72,76,77] | |||
| OCT3 | SLC22A3 | organic cations | metformin (HEK293) | + * | + * | + | R, P | R, P | R, P | ND | ND | NA | NA | [26,27,72,76,77,78,79] | |||
| OCTN1 | SLC22A4 | carnitine, organic cations | sulpiride ** | NA | NA | NA | NA | NA | R | + | + | NA | NA | [27,80] | |||
| OCTN2 | SLC22A5 | carnitine, organic cations | etoposide (HEK293), quinidine, verapamil, and valproate (HEK293) | + | ND | + * | NA | NA | R, P | + | + | NA | NA | [27,81,82,83,84] | |||
| OAT1 | SLC22A6 | alpha -ketoglutarate, PGE2, PGF2a, cGMP, cAMP | adefovir (kidney) | NA | NA | NA | NA | NA | NA | R | NA | NA | NA | NA | [85] | ||
| OAT4 | SLC22A11 | estrone 3-sulfate, | olmesartan | + | + | ND | R, P | R, P | R, P | + | + | NA | NA | [51,68,86,87,88,89,90] | |||
| OAT10 | SLC22A13 | urate, organic cations | cyclosporine ** | + | ND | ND | NA | R, P | R, P | + | NA | NA | NA | [88,91,92] | |||
| SLC29A | ENT1 | SLC29A1 | adenosine, inosine | entecavir, abacavir | + | ND | + | R | NA | R, P | + | NA | NA | NA | [93,94,95,96,97] | ||
| ENT2 | SLC29A2 | adenosine, inosine | entecavir | + | ND | ND | R | NA | R, P | + | NA | NA | NA | [93,94,96,97] | |||
| ENT4 | SLC29A4 | dopamine, histamine, adenosine | atenolol (HEK293) | NA | NA | NA | NA | NA | R | NA | NA | NA | NA | [73,98] | |||
| SLC47A | MATE1 | SLC47A1 | creatine, thiamine | metformin (HEK293), cimetidine (HEK293) | NA | NA | NA | R | NA | R | NA | NA | NA | NA | [76,99,100] | ||
| MATE2 | SLC47A2 | creatine, thiamine | metformin (HEK293), aciclovir (HEK293) | NA | NA | NA | R | NA | R | NA | NA | NA | NA | [76,99,101] | |||
5.1. ABC Transporters
5.1.1. ABCB (MDR) Family
ABCB1 (MDR1) is the transporter that was identified first and is the one that has been the most intensively studied. The expression of MDR1 decreases with progressing gestational age, while the expression of ABCB4 (MDR3) increases [11,34]. The expression level of MDR1 in BeWo cells, which are a frequently used model of human syncytiotrophoblast, is lower than it is in the human placenta, including in term placenta [37,40,73].
5.1.2. ABCC (MRP) Family
ABCC3 (MRP3) is highly expressed in the placenta, but it is express much less in cell lines, whereas ABCC11 (MRP8) is expressed more in cell lines [15]. It was cloned relatively recently, and data are scarce.
5.1.3. ABCG2 (BCRP)
This transporter has been comparatively well studied and is widely expressed in both the placenta and in cell lines. ABCG2(BCRP) is expressed in the apical side of syncytiotrophoblasts, cytotrophoblasts, and fetal endothelial cells. In BeWo, the expression level is higher than it is in the placenta [63]. Although there are various reports on a correlation with the expression level with gestational age, BCRP does not seem to change significantly or decrease slightly as pregnancy progress [36,38,50].
5.2. SLC Transporters
5.2.1. SLC21A (OATP) Family
This group of transporters has been isolated from various animal species, including humans. However, they have more diversity among species than other drug transporters and have many isoforms without homologues between species. SLC21A9 (OATP2B1) expression is far less abundant in cancer-derived cell lines, including in the most frequently used choriocarcinoma cell lines [15].
5.2.2. SLC22A Family
Although SLC22A1 (OCT1) and SLC22A2 (OCT2) have been reported to be expressed in very small amounts, it is commonly recognized that SLC22A3 (OCT3) is the only OCT family member that is clearly expressed in the human placenta, but there are conflicting reports on its cellular localization. One study reported OCT3 expression in the basal membranes of syncytiotrophoblast [78,79], while Kliman et al. showed its exclusive expression in cytotrophoblast cells by IHC [26]. Liu et al. classified trophoblasts into subtypes using single-cell RNA sequencing and analyzed the imprinted gene expression. In this report, OCT3 appears to be expressed in a CTB-dominant manner [102]. Single cell transcriptome research in the human placenta is summarized in a review by Li et al. [103]. For SLC22A5 (OCTN2), different localizations have been reported in fetal blood vessels [81,82]. As described above, the SLC transporters basically act in the direction of substrate uptake, while SLC22A11 (OAT4) also excretes substrates. However, the involvement of this “efflux mode” in drug transport has not been sufficiently proven [91]. OAT4 is expressed in the JEG3 choriocarcinoma cell line but only when forskolin is added [89].
5.2.3. SLC29A (ENT) Family
5.2.4. SLC47A (MATE) Family
The placental expression levels of both SLC47A1 (MATE1) and SLC47A2 (MATE2) decrease as the gestational weeks progress [99].
6. Regulation of Placental Drug Transporters
When considering the functional effects of drug transporters, the expression level of the transporters is also an important factor. As mentioned above, the expression levels of some transporters have been shown to change with the number of gestational weeks. In addition, some factors such as drug–drug interactions, some kinds of pathological conditions such as GDM or preeclampsia, and genetic variations such as single-nucleotide polymorphisms (SNPs) have been reported to cause changes in the expression levels of these transporters. The transporter regulation is summarized in the review by Staud and Ceckova [104].
7. Experimental Models for Human Placental Drug Transport
7.1. Cell Lines
The most commonly used cell line in studies on human placental transport is the choriocarcinoma cell line BeWo, which forms a confluent monolayer with the microvilli on a permeable filter support. BeWo cells are often used as a model for trophoblast maturation because morphological and biochemical changes and syncytialization occur when the intracellular cAMP concentration is increased due to stimulation with forskolin. However, it should be noted that the syncytialization of BeWo may cause changes in the permeability of the monolayer due to the widening of the intercellular spaces and increases in the transport of certain substances [105]. After BeWo, the most commonly used cell line is the choriocarcinoma-derived cell line JEG3. The addition of forskolin only alters JEG3 biochemically, but it does not form a syncytium. The choriocarcinoma cell line JAR is more similar to pre-differentiated cytotrophoblast cells and is less commonly used for transporter studies. Further, HTR-8/SVneo is a cell line that was established by immortalizing extravillous trophoblasts, which are derived from cytotrophoblasts. Their characteristics do not sufficiently reflect those of physiological trophoblast cells, and therefore, they are rarely used in transporter studies [106]. Cell lines are frequently used for functional analyses because they are easy to handle and can be used for overexpression and inhibition studies, as described below.
7.2. Placental Explants
Placental explants are less frequently used for transport studies than cell lines because syncytiotrophoblasts degrade rapidly, but they can be used for functional and toxicological analyses [71]. Placental explants have the advantage of reflecting the physiological villous structure. The localization of cells and factors of interest can be easily determined by immunohistochemistry and other methods [107,108].
7.3. Placental Membrane
The so-called isolated “membrane vesicles” from term placenta have often been used in transporter studies. Previously, they were isolated by means of the apical membrane preparation [109] and basal membrane preparation [110] methods and are mainly used to determine the localization of various transporters. However, as seen in OCT3, there are some cases where membrane preparation and IHC have demonstrated different results.
7.4. Placenta-on-a-Chip
Recently, microfluidic systems called “organs-on-a-chip” have been actively studied, including placenta-on-a-chip. To simulate the placental barrier, co-cultures of BeWo or HTR−8/SVneo and Human Umbilical Vein Endothelial Cells (HUVEC) or Human Placental Vascular Endothelial Cells (HPVEC) have been commonly used [111,112]. So far, the use of primary trophoblast cells in this model has not been published. Placenta-on-a-chip technology is still in its beginning, and only a few studies have been reported. However, it has potential for use in the evaluation of drug transport, as previously reported [113], and further development is expected.
7.5. Ex Vivo Placenta Perfusion Model
This model has been commonly used in toxicological studies. Unlike the in vitro models, which are far from the real in vivo situation, the main feature of this model is its ability to reproduce macroscopic transport in human placenta. The limitations of this model are its short maintenance time of 6 h, and situations modelling the first trimester cannot be reproduced. Nonetheless, this model is a human organ models that can mimic the in vivo situation very closely. Indeed, May et al. perfused candidate transporter inhibitors with talinolol and evaluated the transporters that were involved in the materno–fetal transfer of talinolol [47].
7.6. Functional Assays on Placental Drug Transporters
During functional transporter analyses, cell lines are frequently used. A common method is to add the inhibitors of specific transporters and to evaluate the changes that take place in terms of the substrate uptake [37,41,47,61]. This method has also been performed with placental explants [86]. The role of specific transporters during substrate uptake in cell lines can also be evaluated by overexpression or knock down using RNA mimics or siRNA [46]. Cell lines may also be used to establish hypotheses that need further confirmation in primary cells or tissues. However, there are significant differences in the expression of drug transporters between cell lines and physiological placental tissues or cells [95].
8. Discussion
In this review we have summarized the findings related drug transporters in the human placenta and in experimental models. Transporters have been intensively studied in the kidney, liver, gastrointestinal tract, and at the blood–brain barrier. Pharmacokinetic knowledge is vital in terms of drug efficacy and side effects.
Additionally, the placenta has a strong barrier function in order to ensure that maternal and fetal blood avoid contact with each other and do not mingle, but simultaneously, it has a very active transport function that is used to aliment the fetus and to eliminate its waste products. However, from the studies that have been published thus far, most do not explain the complete mechanisms of transportation of particular substances from the maternal surface to fetal circulation. Understanding the placental permeability of drugs is very important for estimating their toxicity to the fetus as well as for the pharmacological treatment of the fetus.
There are several difficulties in studying drug transporters. First, in vitro models have limitations, as shown by the different transporter localization reuslts in the human placenta when the membrane preparation method or immunostaining have been used (e.g., for OCT3 [26,78,79]). Since l transporter localization is a very important aspect for elucidating the whole picture of substrate uptake, this issue should be considered as an area of focus in the future. Second, there are many further factors to be considered in vivo, such as binding proteins and competing substrates. As it is difficult to reproduce several factors in vitro or in ex vivo models simultaneously, their results do not completely reflect the in vivo situation. The third point is the broad substrate specificity of different transporters. It is not rare that a single substrate is recognized by multiple transporters in the same organ. In other words, the recognition of a drug by a certain transporter is not equal to its importance in pharmacokinetics.
Despite the technical challenges in studying placental transporters, the continuous improvement of experimental models and the combination of different types of experimental methods will lead to better understanding. Developments in the knowledge of placental drug transporters will be beneficial for the health and optimization of drug treatment for pregnant women and their fetuses.
9. Conclusions
In this review, we described the functions and the expression and localization of drug transporters in the human placenta and in experimental models that have been previously used for their study. Detailed knowledge is fundamental for planning drug applications in pregnancy as well as for the consideration of the potential side effects of those applications.
Author Contributions
M.Y. conducted the literature research and wrote the manuscript. U.R.M. supervised the writing and revised the manuscript, and both authors jointly prepared the final version. All authors have read and agreed to the published version of the manuscript.
Funding
M.Y. is supported by a fellowship from Kanzawa Medical Research Foundation. U.R.M. is receiving research grants from the German Ministry of Education and Research (03VP08692) and the German Research Foundation (DFG, Ma1550/12-1).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Juan-Carlos, P.-D.M.; Perla-Lidia, P.-P.; Stephanie-Talia, M.-M.; Mónica-Griselda, A.-M.; Luz-María, T.-E. ABC transporter superfamily. An updated overview, relevance in cancer multidrug resistance and perspectives with personalized medicine. Mol. Biol. Rep. 2021, 48, 1883–1901. [Google Scholar] [CrossRef] [PubMed]
- Schumann, T.; König, J.; Henke, C.; Willmes, D.M.; Bornstein, S.R.; Jordan, J.; Fromm, M.F.; Birkenfeld, A.L. Solute Carrier Transporters as Potential Targets for the Treatment of Metabolic Disease. Pharmacol. Rev. 2020, 72, 343–379. [Google Scholar] [CrossRef]
- Blanco-Castañeda, R.; Galaviz-Hernández, C.; Souto, P.C.D.S.; Lima, V.V.; Giachini, F.R.; Escudero, C.; Damiano, A.E.; Barragán-Zúñiga, L.J.; Martínez-Aguilar, G.; Sosa-Macías, M. The role of xenobiotic-metabolizing enzymes in the placenta: A growing research field. Expert Rev. Clin. Pharmacol. 2020, 13, 247–263. [Google Scholar] [CrossRef] [PubMed]
- Dilworth, M.; Sibley, C. Review: Transport across the placenta of mice and women. Placenta 2012, 34, S34–S39. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, A.; Prieto, D.M.M.; Pastuschek, J.; Fröhlich, K.; Markert, U.R. Only humans have human placentas: Molecular differences between mice and humans. J. Reprod. Immunol. 2015, 108, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Dean, M.; Rzhetsky, A.; Allikmets, R. The Human ATP-Binding Cassette (ABC) Transporter Superfamily. Genome Res. 2001, 11, 1156–1166. [Google Scholar] [CrossRef] [PubMed]
- Mai, Y.; Dou, L.; Yao, Z.; Madla, C.M.; Gavins, F.K.H.; Taherali, F.; Yin, H.; Orlu, M.; Murdan, S.; Basit, A.W. Quantification of P-Glycoprotein in the Gastrointestinal Tract of Humans and Rodents: Methodology, Gut Region, Sex, and Species Matter. Mol. Pharm. 2021, 18, 1895–1904. [Google Scholar] [CrossRef]
- Fu, D. Where Is It and How Does It Get There—Intracellular Localization and Traffic of P-glycoprotein. Front. Oncol. 2013, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruckmueller, H.; Martin, P.; Kähler, M.; Haenisch, S.; Ostrowski, M.; Drozdzik, M.; Siegmund, W.; Cascorbi, I.; Oswald, S. Clinically Relevant Multidrug Transporters Are Regulated by microRNAs along the Human Intestine. Mol. Pharm. 2017, 14, 2245–2253. [Google Scholar] [CrossRef] [Green Version]
- Deleuze, J.; Jacquemin, E.; Dubuisson, C.; Cresteil, D.; Dumont, M.; Erlinger, S.; Bernard, O.; Hadchouel, M. Defect of multidrug-resistance 3 gene expression in a subtype of progressive familial intrahepatic cholestasis. Hepatology 1996, 23, 904–908. [Google Scholar] [CrossRef]
- Patel, P.; Weerasekera, N.; Hitchins, M.; Boyd, C.; Johnston, D.; Williamson, C. Semi Quantitative Expression Analysis of MDR3, FIC1, BSEP, OATP-A, OATP-C,OATP-D, OATP-E and NTCP Gene Transcripts in 1st and 3rd Trimester Human Placenta. Placenta 2003, 24, 39–44. [Google Scholar] [CrossRef]
- Sultana, R.; Butterfield, D.A. Oxidatively Modified GST and MRP1 in Alzheimer’s Disease Brain: Implications for Accumulation of Reactive Lipid Peroxidation Products. Neurochem. Res. 2004, 29, 2215–2220. [Google Scholar] [CrossRef] [PubMed]
- Mookerjee, A.; Basu, J.M.; Majumder, S.; Chatterjee, S.; Panda, G.S.; Dutta, P.; Pal, S.; Mukherjee, P.; Efferth, T.; Roy, S.; et al. A novel copper complex induces ROS generation in doxorubicin resistant Ehrlich ascitis carcinoma cells and increases activity of antioxidant enzymes in vital organs in vivo. BMC Cancer 2006, 6, 267. [Google Scholar] [CrossRef] [Green Version]
- Keitel, V.; Kartenbeck, J.; Nies, A.T.; Spring, H.; Brom, M.; Keppler, D. Impaired protein maturation of the conjugate export pump multidrug resistance protein 2 as a consequence of a deletion mutation in dubin-johnson syndrome. Hepatology 2000, 32, 1317–1328. [Google Scholar] [CrossRef] [PubMed]
- Serrano, M.A.; Macias, R.; Briz, O.; Monte, M.J.; Blazquez, A.; Williamson, C.; Kubitz, R.; Marin, J. Expression in Human Trophoblast and Choriocarcinoma Cell Lines, BeWo, Jeg-3 and JAr of Genes Involved in the Hepatobiliary-like Excretory Function of the Placenta. Placenta 2007, 28, 107–117. [Google Scholar] [CrossRef]
- Reid, G.; Wielinga, P.; Zelcer, N.; van der Heijden, I.; Kuil, A.; de Haas, M.; Wijnholds, J.; Borst, P. The human multidrug resistance protein MRP4 functions as a prostaglandin efflux transporter and is inhibited by nonsteroidal antiinflammatory drugs. Proc. Natl. Acad. Sci. USA 2003, 100, 9244–9249. [Google Scholar] [CrossRef] [Green Version]
- Jedlitschky, G.; Burchell, B.; Keppler, D. The Multidrug Resistance Protein 5 Functions as an ATP-dependent Export Pump for Cyclic Nucleotides. J. Biol. Chem. 2000, 275, 30069–30074. [Google Scholar] [CrossRef] [Green Version]
- Bergen, A.A.; Plomp, A.S.; Schuurman, E.J.; Terry, S.F.; Breuning, M.H.; Dauwerse, H.G.; Swart, J.; Kool, M.; Van Soest, S.; Baas, F.; et al. Mutations in ABCC6 cause pseudoxanthoma elasticum. Nat. Genet. 2000, 25, 228–231. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, H.; Takada, T.; Ichida, K.; Nakamura, T.; Nakayama, A.; Ikebuchi, Y.; Ito, K.; Kusanagi, Y.; Chiba, T.; Tadokoro, S.; et al. Common Defects of ABCG2, a High-Capacity Urate Exporter, Cause Gout: A Function-Based Genetic Analysis in a Japanese Population. Sci. Transl. Med. 2009, 1, 5ra11. [Google Scholar] [CrossRef]
- Fredriksson, R.; Nordström, K.J.; Stephansson, O.; Hägglund, M.G.; Schiöth, H.B. The solute carrier (SLC) complement of the human genome: Phylogenetic classification reveals four major families. FEBS Lett. 2008, 582, 3811–3816. [Google Scholar] [CrossRef] [Green Version]
- Colas, C.; Ung, P.M.-U.; Schlessinger, A. SLC transporters: Structure, function, and drug discovery. MedChemComm 2016, 7, 1069–1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Q.; Zhang, X.; Zhang, L.; Cheng, Y.; Zhao, N.; Li, F.; Zhou, X.; Chen, S.; Li, J.; Xu, S.; et al. Solute Carrier Organic Anion Transporter Family Member 3A1 Is a Bile Acid Efflux Transporter in Cholestasis. Gastroenterology 2018, 155, 1578–1592.e16. [Google Scholar] [CrossRef]
- Roth, M.; Obaidat, A.; Hagenbuch, B. OATPs, OATs and OCTs: The organic anion and cation transporters of the SLCO and SLC22A gene superfamilies. Br. J. Pharmacol. 2012, 165, 1260–1287. [Google Scholar] [CrossRef] [Green Version]
- Wessler, I.; Roth, E.; Deutsch, C.; Brockerhoff, P.; Bittinger, F.; Kirkpatrick, C.J.; Kilbinger, H. Release of non-neuronal acetylcholine from the isolated human placenta is mediated by organic cation transporters. Br. J. Pharmacol. 2001, 134, 951–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reznichenko, A.; Sinkeler, S.J.; Snieder, H.; Born, J.V.D.; De Borst, M.; Damman, J.; Van Dijk, M.C.R.F.; Van Goor, H.; Hepkema, B.G.; Hillebrands, J.-L.; et al. SLC22A2 is associated with tubular creatinine secretion and bias of estimated GFR in renal transplantation. Physiol. Genom. 2013, 45, 201–209. [Google Scholar] [CrossRef] [Green Version]
- Kliman, H.J.; Quaratella, S.B.; Setaro, A.C.; Siegman, E.C.; Subha, Z.T.; Tal, R.; Milano, K.M.; Steck, T.L. Pathway of Maternal Serotonin to the Human Embryo and Fetus. Endocrinology 2018, 159, 1609–1629. [Google Scholar] [CrossRef]
- Hasegawa, N.; Furugen, A.; Ono, K.; Koishikawa, M.; Miyazawa, Y.; Nishimura, A.; Umazume, T.; Narumi, K.; Kobayashi, M.; Iseki, K. Cellular uptake properties of lamotrigine in human placental cell lines: Investigation of involvement of organic cation transporters (SLC22A1–5). Drug Metab. Pharmacokinet. 2020, 35, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Rizwan, A.N.; Burckhardt, G. Organic Anion Transporters of the SLC22 Family: Biopharmaceutical, Physiological, and Pathological Roles. Pharm. Res. 2007, 24, 450–470. [Google Scholar] [CrossRef]
- Higashino, T.; Morimoto, K.; Nakaoka, H.; Toyoda, Y.; Kawamura, Y.; Shimizu, S.; Nakamura, T.; Hosomichi, K.; Nakayama, A.; Ooyama, K.; et al. Dysfunctional missense variant of OAT10/SLC22A13 decreases gout risk and serum uric acid levels. Ann. Rheum. Dis. 2019, 79, 164–166. [Google Scholar] [CrossRef] [Green Version]
- Boswell-Casteel, R.C.; Hays, F.A. Equilibrative nucleoside transporters—A review. Nucleosides Nucleotides Nucleic Acids 2016, 36, 7–30. [Google Scholar] [CrossRef]
- Nies, A.T.; Damme, K.; Kruck, S.; Schaeffeler, E.; Schwab, M. Structure and function of multidrug and toxin extrusion proteins (MATEs) and their relevance to drug therapy and personalized medicine. Arch. Toxicol. 2016, 90, 1555–1584. [Google Scholar] [CrossRef]
- Coles, L.D.; Lee, I.J.; Voulalas, P.J.; Eddington, N.D. Estradiol and Progesterone-Mediated Regulation of P-gp in P-gp Overexpressing Cells (NCI-ADR-RES) and Placental Cells (JAR). Mol. Pharm. 2009, 6, 1816–1825. [Google Scholar] [CrossRef]
- Ushigome, F.; Takanaga, H.; Matsuo, H.; Yanai, S.; Tsukimori, K.; Nakano, H.; Uchiumi, T.; Nakamura, T.; Kuwano, M.; Ohtani, H.; et al. Human placental transport of vinblastine, vincristine, digoxin and progesterone: Contribution of P-glycoprotein. Eur. J. Pharmacol. 2000, 408, 1–10. [Google Scholar] [CrossRef]
- Sun, M.; Kingdom, J.; Baczyk, D.; Lye, S.; Matthews, S.; Gibb, W. Expression of the Multidrug Resistance P-Glycoprotein, (ABCB1 glycoprotein) in the Human Placenta Decreases with Advancing Gestation. Placenta 2006, 27, 602–609. [Google Scholar] [CrossRef]
- Gil, S.; Saura, R.; Forestier, F.; Farinotti, R. P-glycoprotein expression of the human placenta during pregnancy. Placenta 2005, 26, 268–270. [Google Scholar] [CrossRef] [PubMed]
- Mathias, A.A.; Hitti, J.; Unadkat, J.D. P-glycoprotein and breast cancer resistance protein expression in human placentae of various gestational ages. Am. J. Physiol. Integr. Comp. Physiol. 2005, 289, R963–R969. [Google Scholar] [CrossRef]
- Evseenko, D.A.; Paxton, J.; Keelan, J. ABC drug transporter expression and functional activity in trophoblast-like cell lines and differentiating primary trophoblast. Am. J. Physiol. Integr. Comp. Physiol. 2006, 290, R1357–R1365. [Google Scholar] [CrossRef] [PubMed]
- Lye, P.; Bloise, E.; Dunk, C.; Javam, M.; Gibb, W.; Lye, S.; Matthews, S. Effect of oxygen on multidrug resistance in the first trimester human placenta. Placenta 2013, 34, 817–823. [Google Scholar] [CrossRef] [PubMed]
- Dunk, C.E.; Pappas, J.J.; Lye, P.; Kibschull, M.; Javam, M.; Bloise, E.; Lye, S.J.; Szyf, M.; Matthews, S.G. P-Glycoprotein (P-gp)/ABCB1 plays a functional role in extravillous trophoblast (EVT) invasion and is decreased in the pre-eclamptic placenta. J. Cell. Mol. Med. 2018, 22, 5378–5393. [Google Scholar] [CrossRef]
- Kallol, S.; Moser-Haessig, R.; Ontsouka, C.E.; Albrecht, C. Comparative expression patterns of selected membrane transporters in differentiated BeWo and human primary trophoblast cells. Placenta 2018, 72–73, 48–52. [Google Scholar] [CrossRef]
- Tupova, L.; Hirschmugl, B.; Sucha, S.; Pilarova, V.; Székely, V.; Bakos, É.; Novakova, L.; Özvegy-Laczka, C.; Wadsack, C.; Ceckova, M. Interplay of drug transporters P-glycoprotein (MDR1), MRP1, OATP1A2 and OATP1B3 in passage of maraviroc across human placenta. Biomed. Pharmacother. 2020, 129, 110506. [Google Scholar] [CrossRef] [PubMed]
- St-Pierre, M.V.; Serrano, M.A.; Macias, R.; Dubs, U.; Hoechli, M.; Lauper, U.; Meier, P.J.; Marin, J. Expression of members of the multidrug resistance protein family in human term placenta. Am. J. Physiol. Integr. Comp. Physiol. 2000, 279, R1495–R1503. [Google Scholar] [CrossRef] [PubMed]
- Aye, I.; Paxton, J.; Evseenko, D.; Keelan, J. Expression, Localisation and Activity of ATP Binding Cassette (ABC) Family of Drug Transporters in Human Amnion Membranes. Placenta 2007, 28, 868–877. [Google Scholar] [CrossRef]
- Pascolo, L.; Fernetti, C.; Pirulli, D.; Crovella, S.; Amoroso, A.; Tiribelli, C. Effects of maturation on RNA transcription and protein expression of four MRP genes in human placenta and in BeWo cells. Biochem. Biophys. Res. Commun. 2003, 303, 259–265. [Google Scholar] [CrossRef]
- Zu Schwabedissen, H.E.M.; Grube, M.; Heydrich, B.; Linnemann, K.; Fusch, C.; Kroemer, H.K.; Jedlitschky, G. Expression, Localization, and Function of MRP5 (ABCC5), a Transporter for Cyclic Nucleotides, in Human Placenta and Cultured Human Trophoblasts: Effects of Gestational Age and Cellular Differentiation. Am. J. Pathol. 2005, 166, 39–48. [Google Scholar] [CrossRef]
- Granitzer, S.; Ellinger, I.; Khan, R.; Gelles, K.; Widhalm, R.; Hengstschläger, M.; Zeisler, H.; Desoye, G.; Tupova, L.; Ceckova, M.; et al. In vitro function and in situ localization of Multidrug Resistance-associated Protein (MRP)1 (ABCC1) suggest a protective role against methyl mercury-induced oxidative stress in the human placenta. Arch. Toxicol. 2020, 94, 3799–3817. [Google Scholar] [CrossRef]
- May, K.; Minarikova, V.; Linnemann, K.; Zygmunt, M.; Kroemer, H.K.; Fusch, C.; Siegmund, W. Role of the Multidrug Transporter Proteins ABCB1 and ABCC2 in the Diaplacental Transport of Talinolol in the Term Human Placenta. Drug Metab. Dispos. 2008, 36, 740–744. [Google Scholar] [CrossRef] [Green Version]
- Azzaroli, F.; Mennone, A.; Feletti, V.; Simoni, P.; Baglivo, E.; Montagnani, M.; Rizzo, N.; Pelusi, G.; DE Aloysio, D.; Lodato, F.; et al. Clinical trial: Modulation of human placental multidrug resistance proteins in cholestasis of pregnancy by ursodeoxycholic acid. Aliment. Pharmacol. Ther. 2007, 26, 1139–1146. [Google Scholar] [CrossRef]
- Zu Schwabedissen, H.E.M.; Jedlitschky, G.; Gratz, M.; Haenisch, S.; Linnemann, K.; Fusch, C.; Cascorbi, I.; Kroemer, H.K. Variable expression of MRP2 (ABCC2) in human placenta: Influence of gestational age and cellular differentiation. Drug Metab. Dispos. 2005, 33, 896–904. [Google Scholar] [CrossRef] [PubMed]
- Mason, C.W.; Buhimschi, I.; Buhimschi, C.S.; Dong, Y.; Weiner, C.P.; Swaan, P.W. ATP-Binding Cassette Transporter Expression in Human Placenta as a Function of Pregnancy Condition. Drug Metab. Dispos. 2011, 39, 1000–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huuskonen, P.; Myllynen, P.; Storvik, M.; Pasanen, M. The effects of aflatoxin B1 on transporters and steroid metabolizing enzymes in JEG-3 cells. Toxicol. Lett. 2013, 218, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.S.; Belinsky, M.G.; Kruh, G.D.; Zeng, H.; Rea, P.A. Transport of methotrexate (MTX) and folates by multidrug resistance protein (MRP) 3 and MRP1: Effect of polyglutamylation on MTX transport. Cancer Res. 2001, 61, 7225–7232. [Google Scholar]
- Imaoka, T.; Kusuhara, H.; Adachi, M.; Schuetz, J.D.; Takeuchi, K.; Sugiyama, Y. Functional Involvement of Multidrug Resistance-Associated Protein 4 (MRP4/ABCC4) in the Renal Elimination of the Antiviral Drugs Adefovir and Tenofovir. Mol. Pharmacol. 2006, 71, 619–627. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, M.; Suzuki, T.; Komiya, T.; Hatashita, E.; Nishio, K.; Kazuhiko, N.; Fukuoka, M. Induction of MRP5 and SMRP mRNA by adriamycin exposure and its overexpression in human lung cancer cells resistant to adriamycin. Int. J. Cancer 2001, 94, 432–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belinsky, M.G.; Chen, Z.-S.; Shchaveleva, I.; Zeng, H.; Kruh, G.D. Characterization of the drug resistance and transport properties of multidrug resistance protein 6 (MRP6, ABCC6). Cancer Res. 2002, 62, 6172–6177. [Google Scholar] [PubMed]
- Madon, J.; Hagenbuch, B.; Landmann, L.; Meier, P.J.; Stieger, B. Transport Function and Hepatocellular Localization of mrp6 in Rat Liver. Mol. Pharmacol. 2000, 57, 634–641. [Google Scholar] [CrossRef] [PubMed]
- Yabuuchi, H.; Shimizu, H.; Takayanagi, S.-I.; Ishikawa, T. Multiple Splicing Variants of Two New Human ATP-Binding Cassette Transporters, ABCC11 and ABCC12. Biochem. Biophys. Res. Commun. 2001, 288, 933–939. [Google Scholar] [CrossRef] [PubMed]
- Tammur, J.; Prades, C.; Arnould, I.; Rzhetsky, A.; Hutchinson, A.; Adachi, M.; Schuetz, J.D.; Swoboda, K.; Ptácek, L.J.; Rosier, M.; et al. Two new genes from the human ATP-binding cassette transporter superfamily, ABCC11 and ABCC12, tandemly duplicated on chromosome 16q12. Gene 2001, 273, 89–96. [Google Scholar] [CrossRef]
- Joshi, A.A.; Vaidya, S.S.; St-Pierre, M.V.; Mikheev, A.M.; Desino, K.E.; Nyandege, A.N.; Audus, K.L.; Unadkat, J.D.; Gerk, P.M. Placental ABC Transporters: Biological Impact and Pharmaceutical Significance. Pharm. Res. 2016, 33, 2847–2878. [Google Scholar] [CrossRef] [PubMed]
- Vinot, C.; Gavard, L.; Tréluyer, J.M.; Manceau, S.; Courbon, E.; Scherrmann, J.M.; Decleves, X.; Duro, D.; Peytavin, G.; Mandelbrot, L.; et al. Placental Transfer of Maraviroc in anEx VivoHuman Cotyledon Perfusion Model and Influence of ABC Transporter Expression. Antimicrob. Agents Chemother. 2013, 57, 1415–1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feinshtein, V.; Holcberg, G.; Amash, A.; Erez, N.; Rubin, M.; Sheiner, E.; Polachek, H.; Ben-Zvi, Z. Nitrofurantoin transport by placental choriocarcinoma JAr cells: Involvement of BCRP, OATP2B1 and other MDR transporters. Arch. Gynecol. Obstet. 2009, 281, 1037–1044. [Google Scholar] [CrossRef] [PubMed]
- Afrouzian, M.; Al-Lahham, R.; Patrikeeva, S.; Xu, M.; Fokina, V.; Fischer, W.G.; Abdel-Rahman, S.Z.; Costantine, M.; Ahmed, M.S.; Nanovskaya, T. Role of the efflux transporters BCRP and MRP1 in human placental bio-disposition of pravastatin. Biochem. Pharmacol. 2018, 156, 467–478. [Google Scholar] [CrossRef] [PubMed]
- Ceckova, M.; Libra, A.; Pavek, P.; Nachtigal, P.; Brabec, M.; Fuchs, R.; Staud, F. Expression and functional activity of breast cancer resistance protein (BCRP, ABCG2) transporter in the human choriocarcinoma cell line BEWO. Clin. Exp. Pharmacol. Physiol. 2006, 33, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Lye, P.; Bloise, E.; Nadeem, L.; Gibb, W.; Lye, S.J.; Matthews, S.G. Glucocorticoids modulate multidrug resistance transporters in the first trimester human placenta. J. Cell. Mol. Med. 2018, 22, 3652–3660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loubière, L.; Vasilopoulou, E.; Bulmer, J.; Taylor, P.; Stieger, B.; Verrey, F.; McCabe, C.; Franklyn, J.; Kilby, M.; Chan, S.-Y. Expression of thyroid hormone transporters in the human placenta and changes associated with intrauterine growth restriction. Placenta 2010, 31, 295–304. [Google Scholar] [CrossRef] [Green Version]
- Litjens, C.H.C.; Heuvel, J.J.M.W.v.D.; Russel, F.G.M.; Aarnoutse, R.E.; Brake, L.H.M.T.; Koenderink, J.B. Rifampicin Transport by OATP1B1 Variants. Antimicrob. Agents Chemother. 2020, 64. [Google Scholar] [CrossRef]
- Kameyama, Y.; Yamashita, K.; Kobayashi, K.; Hosokawa, M.; Chiba, K. Functional characterization of SLCO1B1 (OATP-C) variants, SLCO1B1*5, SLCO1B1*15 and SLCO1B1*15+C1007G, by using transient expression systems of HeLa and HEK293 cells. Pharm. Genom. 2005, 15, 513–522. [Google Scholar] [CrossRef]
- Ugele, B.; St-Pierre, M.V.; Pihusch, M.; Bahn, A.; Hantschmann, P. Characterization and identification of steroid sulfate transporters of human placenta. Am. J. Physiol. Metab. 2003, 284, E390–E398. [Google Scholar] [CrossRef]
- Pinto, L.; Bapat, P.; Moreira, F.D.L.; Lubetsky, A.; Cavalli, R.D.C.; Berger, H.; Lanchote, V.L.; Koren, G. Chiral Transplacental Pharmacokinetics of Fexofenadine: Impact of P-Glycoprotein Inhibitor Fluoxetine Using the Human Placental Perfusion Model. Pharm. Res. 2021, 38, 647–655. [Google Scholar] [CrossRef] [PubMed]
- Atilano-Roque, A.; Joy, M.S. Characterization of simvastatin acid uptake by organic anion transporting polypeptide 3A1 (OATP3A1) and influence of drug-drug interaction. Toxicol. Vitr. 2017, 45, 158–165. [Google Scholar] [CrossRef]
- Lofthouse, E.M.; Torrens, C.; Manousopoulou, A.; Nahar, M.; Cleal, J.K.; O’Kelly, M.I.; Sengers, B.G.; Garbis, S.D.; Lewis, R.M. Ursodeoxycholic acid inhibits uptake and vasoconstrictor effects of taurocholate in human placenta. FASEB J. 2019, 33, 8211–8220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bottalico, B.; Larsson, I.; Brodszki, J.; Hernandez-Andrade, E.; Casslén, B.; Marsál, K.; Hansson, S. Norepinephrine Transporter (NET), Serotonin Transporter (SERT), Vesicular Monoamine Transporter (VMAT2) and Organic Cation Transporters (OCT1, 2 and EMT) in Human Placenta from Pre-eclamptic and Normotensive Pregnancies. Placenta 2004, 25, 518–529. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.; Hebert, M.F.; Prasad, B.; Easterling, T.R.; Kelly, E.J.; Unadkat, J.D.; Wang, J. Effect of Gestational Age on mRNA and Protein Expression of Polyspecific Organic Cation Transporters during Pregnancy. Drug Metab. Dispos. 2013, 41, 2225–2232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellawatty, W.E.A.; Masuo, Y.; Fujita, K.-I.; Yamazaki, E.; Ishida, H.; Arakawa, H.; Nakamichi, N.; Abdelwahed, R.; Sasaki, Y.; Kato, Y. Organic Cation Transporter 1 Is Responsible for Hepatocellular Uptake of the Tyrosine Kinase Inhibitor Pazopanib. Drug Metab. Dispos. 2017, 46, 33–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, M.J.; Seitz, T.; Brockmöller, J.; Tzvetkov, M.V. Effects of genetic polymorphisms on the OCT1 and OCT2-mediated uptake of ranitidine. PLoS ONE 2017, 12, e0189521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brake, L.H.M.T.; Heuvel, J.J.M.W.V.D.; Buaben, A.O.; van Crevel, R.; Bilos, A.; Russel, F.G.; Aarnoutse, R.E.; Koenderink, J.B. Moxifloxacin Is a Potent In Vitro Inhibitor of OCT- and MATE-Mediated Transport of Metformin and Ethambutol. Antimicrob. Agents Chemother. 2016, 60, 7105–7114. [Google Scholar] [CrossRef] [Green Version]
- Kimura, N.; Okuda, M.; Inui, K.-I. Metformin Transport by Renal Basolateral Organic Cation Transporter hOCT2. Pharm. Res. 2005, 22, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Sata, R.; Ohtani, H.; Tsujimoto, M.; Murakami, H.; Koyabu, N.; Nakamura, T.; Uchiumi, T.; Kuwano, M.; Nagata, H.; Tsukimori, K.; et al. Functional Analysis of Organic Cation Transporter 3 Expressed in Human Placenta. J. Pharmacol. Exp. Ther. 2005, 315, 888–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karahoda, R.; Horackova, H.; Kastner, P.; Matthios, A.; Cerveny, L.; Kucera, R.; Kacerovsky, M.; Tebbens, J.D.; Bonnin, A.; Abad, C.; et al. Serotonin homeostasis in the materno-foetal interface at term: Role of transporters (SERT/SLC6A4 and OCT3/SLC22A3) and monoamine oxidase A (MAO-A) in uptake and degradation of serotonin by human and rat term placenta. Acta Physiol. 2020, 229, e13478. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Sawano, T.; Jinriki, T.; Sato, J. Studies on Intestinal Absorption of Sulpiride (3): Intestinal Absorption of Sulpiride in Rats. Biol. Pharm. Bull. 2004, 27, 77–81. [Google Scholar] [CrossRef] [Green Version]
- Grube, M.; zu Schwabedissen, H.M.; Draber, K.; Präger, D.; Möritz, K.-U.; Linnemann, K.; Fusch, C.; Jedlitschky, G.; Kroemer, H.K. Expression, localization, and function of the carnitine transporter OCTN2 (SLC22A5) in human placenta. Drug Metab. Dispos. 2004, 33, 31–37. [Google Scholar] [CrossRef] [Green Version]
- Chang, T.-T.; Shyu, M.-K.; Huang, M.-C.; Hsu, C.-C.; Yeh, S.-Y.; Chen, M.-R.; Lin, C.-J.; Chun-Jung, L. Hypoxia-Mediated Down-Regulation of OCTN2 and PPARα Expression in Human Placentas and in BeWo Cells. Mol. Pharm. 2010, 8, 117–125. [Google Scholar] [CrossRef]
- Hu, C.; Lancaster, C.S.; Zuo, Z.; Hu, S.; Chen, Z.; Rubnitz, J.E.; Baker, S.; Sparreboom, A. Inhibition of OCTN2-Mediated Transport of Carnitine by Etoposide. Mol. Cancer Ther. 2012, 11, 921–929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohashi, R.; Tamai, I.; Yabuuchi, H.; Nezu, J.I.; Oku, A.; Sai, Y.; Shimane, M.; Tsuji, A. Na(+)-dependent carnitine transport by organic cation transporter (OCTN2): Its pharmacological and toxicological relevance. J. Pharmacol. Exp. Ther. 1999, 291, 778–784. [Google Scholar]
- Hosoyamada, M.; Sekine, T.; Kanai, Y.; Endou, H. Molecular cloning and functional expression of a multispecific organic anion transporter from human kidney. Am. J. Physiol. Physiol. 1999, 276, F122–F128. [Google Scholar] [CrossRef]
- Lofthouse, E.; Brooks, S.E.; Cleal, J.; Hanson, M.; Poore, K.; O’Kelly, I.M.; Lewis, R.M. Glutamate cycling may drive organic anion transport on the basal membrane of human placental syncytiotrophoblast. J. Physiol. 2015, 593, 4549–4559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cha, S.H.; Sekine, T.; Kusuhara, H.; Yu, E.; Kim, J.Y.; Kim, D.K.; Sugiyama, Y.; Kanai, Y.; Endou, H. Molecular Cloning and Characterization of Multispecific Organic Anion Transporter 4 Expressed in the Placenta. J. Biol. Chem. 2000, 275, 4507–4512. [Google Scholar] [CrossRef] [Green Version]
- Uehara, I.; Kimura, T.; Tanigaki, S.; Fukutomi, T.; Sakai, K.; Shinohara, Y.; Ichida, K.; Iwashita, M.; Sakurai, H. Paracellular route is the major urate transport pathway across the blood-placental barrier. Physiol. Rep. 2014, 2, e12013. [Google Scholar] [CrossRef] [PubMed]
- Tomi, M.; Miyata, Y.; Noguchi, S.; Nishimura, S.; Nakashima, E. Role of protein kinase A in regulating steroid sulfate uptake for estrogen production in human placental choriocarcinoma cells. Placenta 2014, 35, 658–660. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, S.; Nishimura, T.; Fujibayashi, A.; Maruyama, T.; Tomi, M.; Nakashima, E. Organic Anion Transporter 4-Mediated Transport of Olmesartan at Basal Plasma Membrane of Human Placental Barrier. J. Pharm. Sci. 2015, 104, 3128–3135. [Google Scholar] [CrossRef] [Green Version]
- Burckhardt, G. Drug transport by Organic Anion Transporters (OATs). Pharmacol. Ther. 2012, 136, 106–130. [Google Scholar] [CrossRef] [PubMed]
- Bahn, A.; Hagos, Y.; Reuter, S.; Balen, D.; Brzica, H.; Krick, W.; Burckhardt, B.C.; Sabolić, I.; Burckhardt, G. Identification of a New Urate and High Affinity Nicotinate Transporter, hOAT10 (SLC22A13). J. Biol. Chem. 2008, 283, 16332–16341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimura, T.; Sano, Y.; Takahashi, Y.; Noguchi, S.; Uchida, Y.; Takagi, A.; Tanaka, T.; Katakura, S.; Nakashima, E.; Tachikawa, M.; et al. Quantification of ENT1 and ENT2 Proteins at the Placental Barrier and Contribution of These Transporters to Ribavirin Uptake. J. Pharm. Sci. 2019, 108, 3917–3922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerveny, L.; Ptackova, Z.; Ceckova, M.; Karahoda, R.; Karbanova, S.; Jiraskova, L.; Greenwood, S.L.; Glazier, J.D.; Staud, F. Equilibrative Nucleoside Transporter 1 (ENT1, SLC29A1) Facilitates Transfer of the Antiretroviral Drug Abacavir across the Placenta. Drug Metab. Dispos. 2018, 46, 1817–1826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfeifer, E.; Parrott, J.; Lee, G.T.; Domalakes, E.; Zhou, H.; He, L.; Mason, C.W. Regulation of human placental drug transporters in HCV infection and their influence on direct acting antiviral medications. Placenta 2018, 69, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Govindarajan, R.; Bakken, A.H.; Hudkins, K.L.; Lai, Y.; Casado, F.J.; Anglada, M.P.; Tse, C.-M.; Hayashi, J.; Unadkat, J.D. In situ hybridization and immunolocalization of concentrative and equilibrative nucleoside transporters in the human intestine, liver, kidneys, and placenta. Am. J. Physiol. Integr. Comp. Physiol. 2007, 293, R1809–R1822. [Google Scholar] [CrossRef] [Green Version]
- Errasti-Murugarren, E.; Díaz, P.; Godoy, V.; Riquelme, G.; Pastor-Anglada, M. Expression and Distribution of Nucleoside Transporter Proteins in the Human Syncytiotrophoblast. Mol. Pharmacol. 2011, 80, 809–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mimura, Y.; Yasujima, T.; Ohta, K.; Inoue, K.; Yuasa, H. Functional Identification of Plasma Membrane Monoamine Transporter (PMAT/SLC29A4) as an Atenolol Transporter Sensitive to Flavonoids Contained in Apple Juice. J. Pharm. Sci. 2017, 106, 2592–2598. [Google Scholar] [CrossRef] [Green Version]
- Ahmadimoghaddam, D.; Zemankova, L.; Nachtigal, P.; Dolezelova, E.; Neumanova, Z.; Červený, L.; Ceckova, M.; Kacerovsky, M.; Micuda, S.; Staud, F. Organic Cation Transporter 3 (OCT3/SLC22A3) and Multidrug and Toxin Extrusion 1 (MATE1/SLC47A1) Transporter in the Placenta and Fetal Tissues: Expression Profile and Fetus Protective Role at Different Stages of Gestation1. Biol. Reprod. 2013, 88, 55. [Google Scholar] [CrossRef]
- Ohta, K.-Y.; Inoue, K.; Yasujima, T.; Ishimaru, M.; Yuasa, H. Functional Characteristics of Two Human MATE Transporters: Kinetics of Cimetidine Transport and Profiles of Iinhibition by Various Compounds. J. Pharm. Pharm. Sci. 2009, 12, 388–396. [Google Scholar] [CrossRef] [Green Version]
- Tanihara, Y.; Masuda, S.; Sato, T.; Katsura, T.; Ogawa, O.; Inui, K.-I. Substrate specificity of MATE1 and MATE2-K, human multidrug and toxin extrusions/H+-organic cation antiporters. Biochem. Pharmacol. 2007, 74, 359–371. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Fan, X.; Wang, R.; Lu, X.; Dang, Y.-L.; Wang, H.; Lin, H.-Y.; Zhu, C.; Ge, H.; Cross, J.C.; et al. Single-cell RNA-seq reveals the diversity of trophoblast subtypes and patterns of differentiation in the human placenta. Cell Res. 2018, 28, 819–832. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Huang, Q.; Liu, Y.; Garmire, L.X. Single cell transcriptome research in human placenta. Reproduction 2020, 160, R155–R167. [Google Scholar] [CrossRef]
- Staud, F.; Ceckova, M. Regulation of drug transporter expression and function in the placenta. Expert Opin. Drug Metab. Toxicol. 2015, 11, 533–555. [Google Scholar] [CrossRef]
- Liu, F.; Soares, M.J.; Audus, K.L. Permeability properties of monolayers of the human trophoblast cell line BeWo. Am. J. Physiol. Physiol. 1997, 273, C1596–C1604. [Google Scholar] [CrossRef] [PubMed]
- Pastuschek, J.; Nonn, O.; Gutiérrez-Samudio, R.N.; Murrieta-Coxca, J.M.; Müller, J.; Sanft, J.; Huppertz, B.; Markert, U.R.; Groten, T.; Morales-Prieto, D.M. Molecular characteristics of established trophoblast-derived cell lines. Placenta 2021, 108, 122–133. [Google Scholar] [CrossRef] [PubMed]
- Baumann, M.U.; Schneider, H.; Malek, A.; Palta, V.; Surbek, D.V.; Sager, R.; Zamudio, S.; Illsley, N.P. Regulation of Human Trophoblast GLUT1 Glucose Transporter by Insulin-Like Growth Factor I (IGF-I). PLoS ONE 2014, 9, e106037. [Google Scholar] [CrossRef] [PubMed]
- Göhner, C.; Svensson-Arvelund, J.; Pfarrer, C.; Häger, J.-D.; Faas, M.; Ernerudh, J.; Cline, J.M.; Dixon, D.; Buse, E.; Markert, U.R. The Placenta in Toxicology. Part IV. Toxicol. Pathol. 2013, 42, 345–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Booth, A.G.; Olaniyan, R.O.; Vanderpuye, O.A. An improved method for the preparation of human placental syncytiotrophoblast microvilli. Placenta 1980, 1, 327–336. [Google Scholar] [CrossRef]
- Kelley, L.K.; Smith, C.H.; King, B.F. Isolation and partial characterization of the basal cell membrane of human placental trophoblast. Biochim. Biophys. Acta (BBA) Biomembr. 1983, 734, 91–98. [Google Scholar] [CrossRef]
- Pu, Y.; Gingrich, J.; Veiga-Lopez, A. A 3-dimensional microfluidic platform for modeling human extravillous trophoblast invasion and toxicological screening. Lab. Chip 2020, 21, 546–557. [Google Scholar] [CrossRef] [PubMed]
- Kreuder, A.-E.; Bolaños-Rosales, A.; Palmer, C.; Thomas, A.; Geiger, M.-A.; Lam, T.; Amler, A.-K.; Markert, U.R.; Lauster, R.; Kloke, L. Inspired by the human placenta: A novel 3D bioprinted membrane system to create barrier models. Sci. Rep. 2020, 10, 15606. [Google Scholar] [CrossRef] [PubMed]
- Blundell, C.; Yi, Y.-S.; Ma, L.; Tess, E.R.; Farrell, M.J.; Georgescu, A.; Aleksunes, L.; Huh, D. Placental Drug Transport-on-a-Chip: A Microengineered In Vitro Model of Transporter-Mediated Drug Efflux in the Human Placental Barrier. Adv. Healthc. Mater. 2017, 7, 1700786. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Human placental structure (above) and localization of drug transporters in human placenta (below). The blue arrows indicate the efflux direction, while the yellow arrows indicate the uptake direction in regard to the specifically shown cell type or compartment. The actual drug transport direction (discharge from placenta into the maternal circulation or uptake into placenta and fetal circulation) depends on the localization of the transporters. The transport direction is indicated by blue arrows for discharge and yellow arrows for uptake. Transporters in the fetal capillary are indicated by pink double-headed arrows because they may work in both directions depending on their exact localization in endothelium. The symbol “Φ” indicates a localization where the respective transporter does not exist. No symbols and no arrows mean that information is not available. The question mark symbol “?” shows that conflicting information exists.
Figure 1.
Human placental structure (above) and localization of drug transporters in human placenta (below). The blue arrows indicate the efflux direction, while the yellow arrows indicate the uptake direction in regard to the specifically shown cell type or compartment. The actual drug transport direction (discharge from placenta into the maternal circulation or uptake into placenta and fetal circulation) depends on the localization of the transporters. The transport direction is indicated by blue arrows for discharge and yellow arrows for uptake. Transporters in the fetal capillary are indicated by pink double-headed arrows because they may work in both directions depending on their exact localization in endothelium. The symbol “Φ” indicates a localization where the respective transporter does not exist. No symbols and no arrows mean that information is not available. The question mark symbol “?” shows that conflicting information exists.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yamashita, M.; Markert, U.R. Overview of Drug Transporters in Human Placenta. Int. J. Mol. Sci. 2021, 22, 13149. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222313149
AMA Style
Yamashita M, Markert UR. Overview of Drug Transporters in Human Placenta. International Journal of Molecular Sciences. 2021; 22(23):13149. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222313149
Chicago/Turabian StyleYamashita, Michiko, and Udo R. Markert. 2021. "Overview of Drug Transporters in Human Placenta" International Journal of Molecular Sciences 22, no. 23: 13149. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222313149
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.