
Modulation of Prostanoids Profile and Counter-Regulation of SDF-1α/CXCR4 and VIP/VPAC2 Expression by Sitagliptin in Non-Diabetic Rat Model of Hepatic Ischemia-Reperfusion Injury


 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
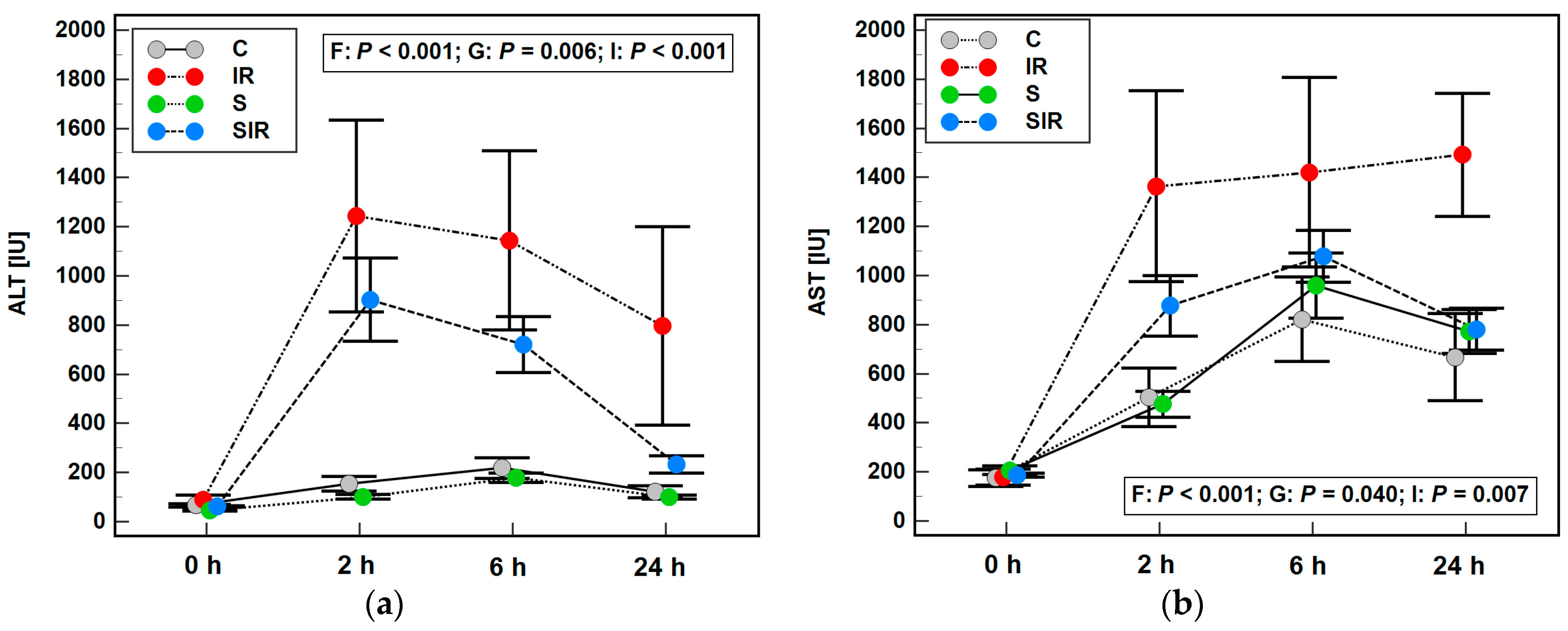
2.1. A Rat Model of IR and Its Validation
2.2. Effect of IR Injury and Sitagliptin on Liver Profile of Eicosanoids
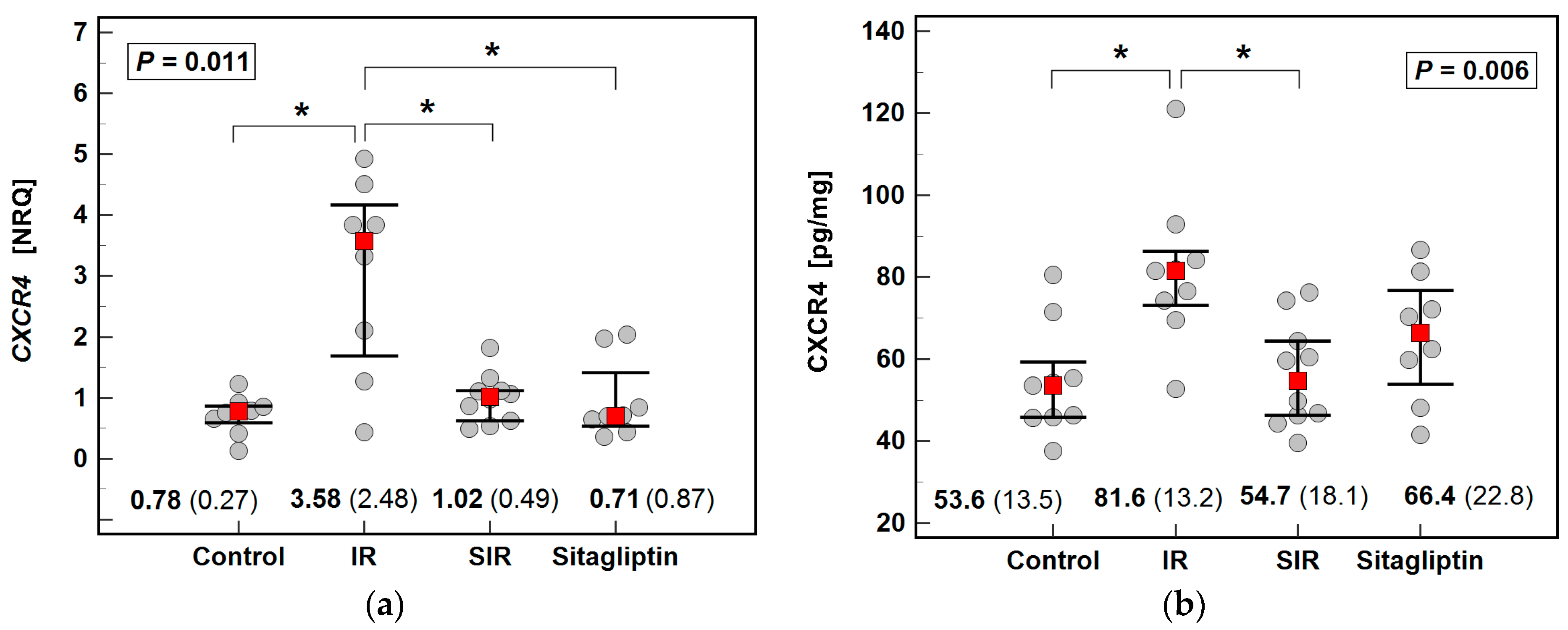
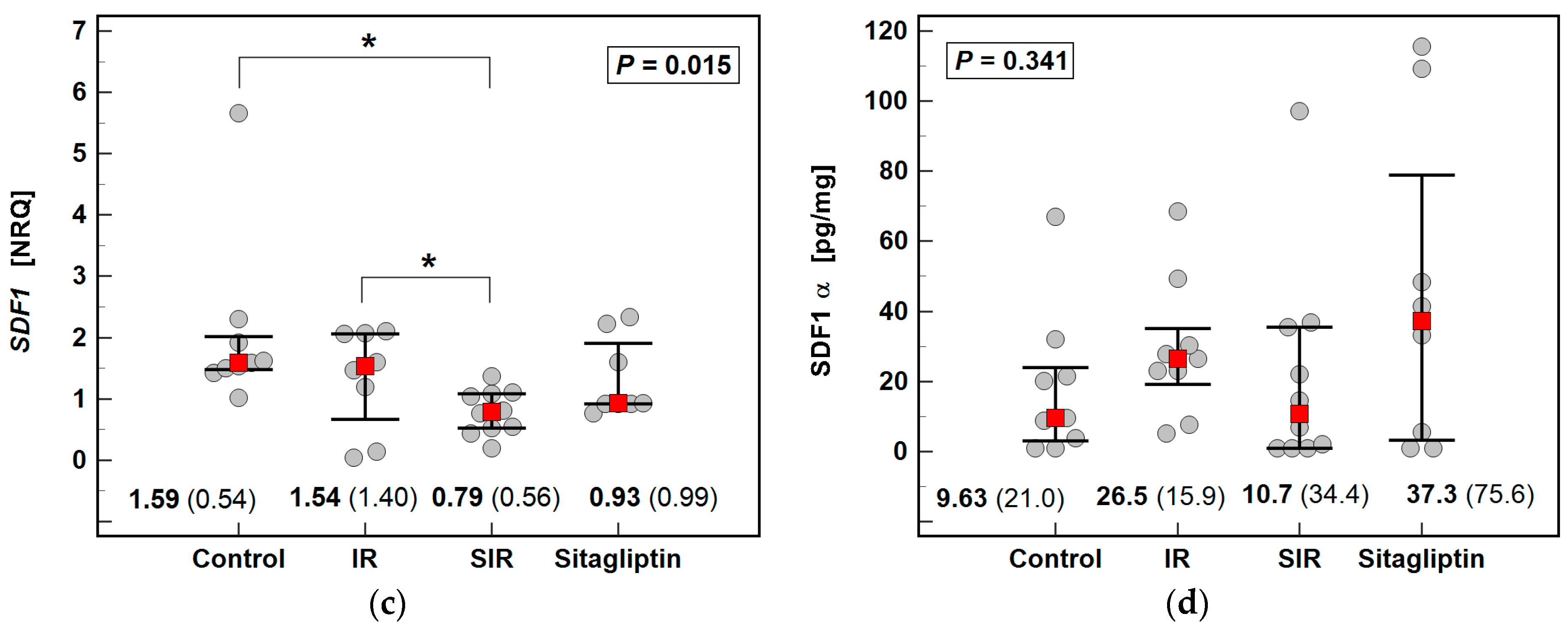
2.3. Effect of IR Injury and Sitagliptin on Liver Expression of CXCR4/SDF1
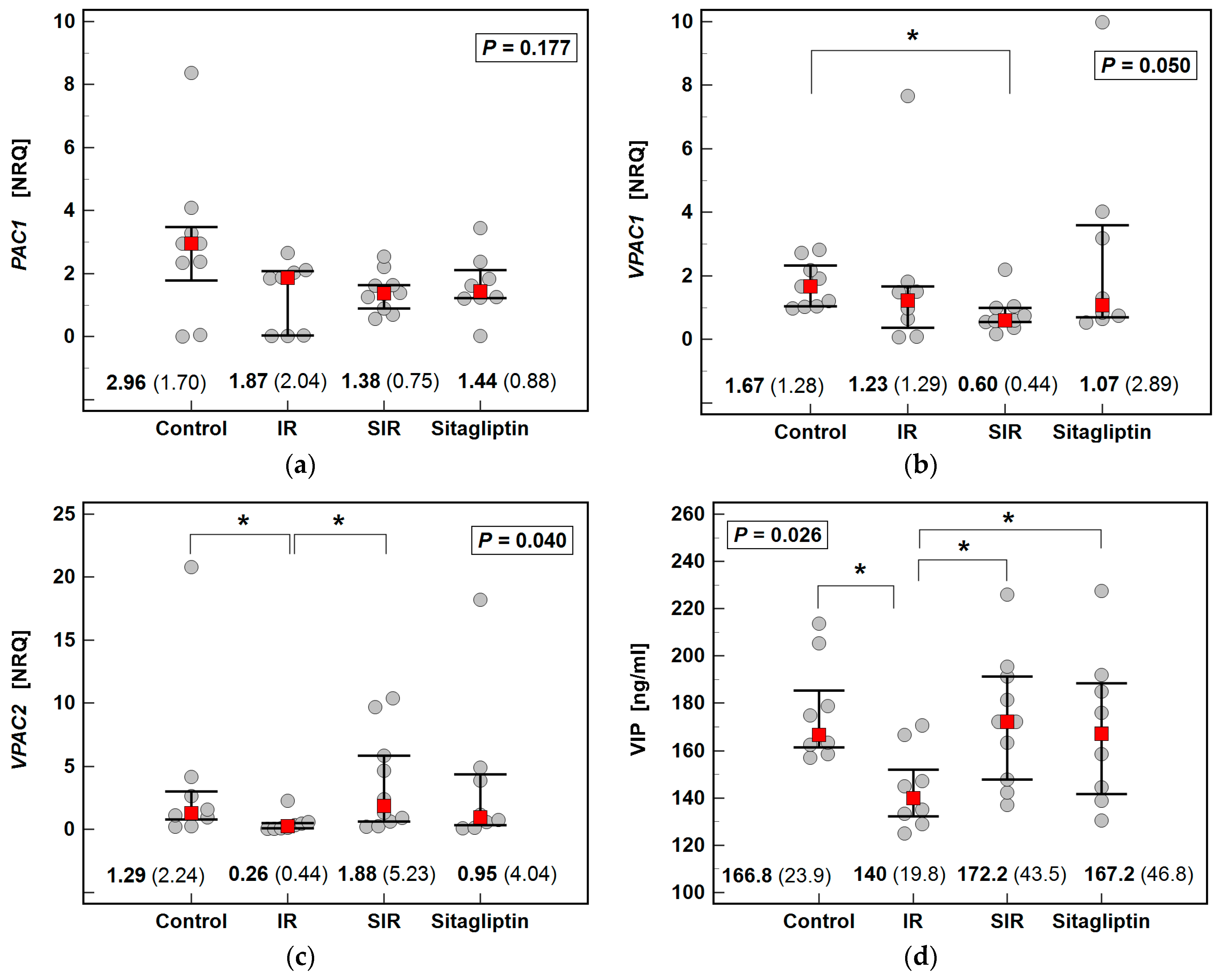
2.4. Effect of IR Injury and Sitagliptin on Liver Expression of PAC1, VPAC1, VPAC2/VIP
2.5. Effect of IR Injury and Sitagliptin on Liver Expression of GLP1R/GLP1
2.6. Relationship between Eicosanoids, DPP4 Ligands, and Their Receptors, and Mediators of Inflammation and Oxidative, Nitrosative, and Halogenative Stress (Univariate Analysis)
2.6.1. Eicosanoids
2.6.2. CXCR4/SDF1
2.6.3. PAC1, VPAC1, VPAC2/VIP
2.6.4. GLP1R/GLP1
2.7. Independent Predictors of Liver Eicosanoids and DPP4 Ligands and Their Receptors (Multivariate Analysis)
2.7.1. Independent Predictors of Liver Eicosanoids
2.7.2. Independent Predictors of Liver Expression of DPP4 Ligands and Their Receptors
3. Discussion
4. Materials and Methods
4.1. Experimental Setting
4.1.1. Animals
4.1.2. Chemicals
4.1.3. The IR Procedure
4.2. Analytical Methods
4.2.1. Metabolomic Analysis of Eicosanoids
Chemicals and Reagents
Sample Preparation
LC-MS Analysis
4.2.2. Immunoassays
4.2.3. Protein Determination
4.2.4. Transcriptional Analysis
4.2.5. Inflammatory Mediators and Markers of Oxidative, Nitrosative, and Halogenative Stress
4.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rampes, S.; Ma, D. Hepatic ischemia-reperfusion injury in liver transplant setting: Mechanisms and protective strategies. J. Biomed. Res. 2019, 33, 221–234. [Google Scholar] [CrossRef]
- Dar, W.A.; Sullivan, E.; Bynon, J.S.; Eltzschig, H.; Ju, C. Ischaemia reperfusion injury in liver transplantation: Cellular and molecular mechanisms. Liver Int. 2019, 39, 788–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smyth, E.M.; Grosser, T.; Wang, M.; Yu, Y.; FitzGerald, G.A. Prostanoids in health and disease. J Lipid Res. 2009, 50, S423–S428. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Wu, L.; Chen, J.; Dong, L.; Chen, C.; Wen, Z.; Hu, J.; Fleming, I.; Wang, D.W. Metabolism pathways of arachidonic acids: Mechanisms and potential therapeutic targets. Signal. Transduct. Target. Ther. 2021, 6, 94. [Google Scholar] [CrossRef]
- Cheng, H.; Huang, H.; Guo, Z.; Chang, Y.; Li, Z. Role of prostaglandin E2 in tissue repair and regeneration. Theranostics 2021, 11, 8836–8854. [Google Scholar] [CrossRef]
- Hamada, T.; Tsuchihashi, S.; Avanesyan, A.; Duarte, S.; Moore, C.; Busuttil, R.W.; Coito, A.J. Cyclooxygenase-2 deficiency enhances Th2 immune responses and impairs neutrophil recruitment in hepatic ischemia/reperfusion injury. J. Immunol. 2008, 180, 1843–1853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abo-Youssef, A.M.; Messiha, B.A.S. The Protective Effect of Dexamethasone, Aspirin and Bromocriptine on Hepatic Ischemia/Reperfusion Injury in Rats. Res. J. Pharmacol. 2014, 8, 6–13. [Google Scholar]
- Taghizadieh, M.; Hajipour, B.; Asl, N.A.; Khodadadi, A.; Somi, M.H.; Banei, M. Combination effect of melatonin and dexamethasone on liver ischemia/reperfusion injury. Bratisl. Lek. Listy 2016, 117, 47–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishizawa, N.; Ito, Y.; Eshima, K.; Ohkubo, H.; Kojo, K.; Inoue, T.; Raouf, J.; Jakobsson, P.J.; Uematsu, S.; Akira, S.; et al. Inhibition of microsomal prostaglandin E synthase-1 facilitates liver repair after hepatic injury in mice. J. Hepatol. 2018, 69, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, Y.; Ding, H.; Shi, X.; Ren, H. Mesenchymal stem cell-secreted prostaglandin E2 ameliorates acute liver failure via attenuation of cell death and regulation of macrophage polarization. Stem Cell Res. Ther. 2021, 12, 15. [Google Scholar] [CrossRef] [PubMed]
- Kuzumoto, Y.; Sho, M.; Ikeda, N.; Hamada, K.; Mizuno, T.; Akashi, S.; Tsurui, Y.; Kashizuka, H.; Nomi, T.; Kubo, A.; et al. Significance and therapeutic potential of prostaglandin E2 receptor in hepatic ischemia/reperfusion injury in mice. Hepatology 2005, 42, 608–617. [Google Scholar] [CrossRef] [PubMed]
- Sherif, I.O.; Al-Shaalan, N.H. Vildagliptin attenuates hepatic ischemia/reperfusion Iinjury via the TLR4/NF-κB signaling pathway. Oxid. Med. Cell. Longev. 2018, 2018, 3509091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song-Chol, M.; Hye-Sun, H. Dipeptidyl peptidase-4 inhibitor sitagliptin prevents hepatic injury via liver ischemia/reperfusion in rats. Int. J. Clin. Exp. Physiol. 2018, 5, 123–126. [Google Scholar] [CrossRef]
- Abdel-Gaber, S.A.; Geddawy, A.; Moussa, R.A. The hepatoprotective effect of sitagliptin against hepatic ischemia reperfusion-induced injury in rats involves Nrf-2/HO-1 pathway. Pharmacol. Rep. 2019, 71, 1044–1049. [Google Scholar] [CrossRef]
- Trocha, M.; Nowak, B.; Merwid-Ląd, A.; Szuba, A.; Dzięgiel, P.; Pieśniewska, M.; Gomułkiewicz, A.; Wiśniewski, J.; Piasecki, T.; Gziut, M.; et al. The impact of sitagliptin, inhibitor of dipeptidyl peptidase-4 (DPP-4), on the ADMA-DDAH-NO pathway in ischemic and reperfused rat livers. Adv. Clin. Exp. Med. 2018, 27, 1483–1490. [Google Scholar] [CrossRef] [PubMed]
- Trocha, M.; Krzystek-Korpacka, M.; Merwid-Ląd, A.; Nowak, B.; Pieśniewska, M.; Dzięgiel, P.; Gomułkiewicz, A.; Kowalski, P.; Diakowska, D.; Szeląg, A.; et al. Sitagliptin-dependent differences in the intensity of oxidative stress in rat livers subjected to ischemia and reperfusion. Oxid. Med. Cell. Longev. 2019, 2019, 2738605. [Google Scholar] [CrossRef] [Green Version]
- Trocha, M.; Fleszar, M.G.; Fortuna, P.; Lewandowski, Ł.; Gostomska-Pampuch, K.; Sozański, T.; Merwid-Ląd, A.; Krzystek-Korpacka, M. Sitagliptin Modulates Oxidative, Nitrative and Halogenative Stress and Inflammatory Response in Rat Model of Hepatic Ischemia-Reperfusion. Antioxidants 2021, 10, 1168. [Google Scholar] [CrossRef] [PubMed]
- Mun, S.C.; Hong, H.S. The effect of sitagliptin on hepatic ischemic reperfusion injury in rats. Med. J. Dr. DY Patil Vidyapeeth 2020, 13, 156–160. [Google Scholar] [CrossRef]
- Bradic, J.; Milosavljevic, I.; Bolevich, S.; Litvitskiy, P.F.; Jeremic, N.; Bolevich, S.; Zivkovic, V.; Srejovic, I.; Jeremic, J.; Jovicic, N.; et al. Dipeptidyl peptidase 4 inhibitors attenuate cardiac ischaemia-reperfusion injury in rats with diabetes mellitus type 2. Clin. Exp. Pharmacol. Physiol. 2021, 48, 575–584. [Google Scholar] [CrossRef]
- Rankovic, M.; Jeremic, N.; Srejovic, I.; Radonjic, K.; Stojanovic, A.; Glisic, M.; Bolevich, S.; Bolevich, S.; Jakovljevic, V. Dipeptidyl peptidase-4 inhibitors as new tools for cardioprotection. Heart Fail. Rev. 2021, 26, 437–450. [Google Scholar] [CrossRef] [PubMed]
- Glorie, L.L.; Verhulst, A.; Matheeussen, V.; Baerts, L.; Magielse, J.; Hermans, N.; D’Haese, P.C.; De Meester, I.; De Beuf, A. DPP4 inhibition improves functional outcome after renal ischemia-reperfusion injury. Am. J. Physiol. Renal Physiol. 2012, 303, F681–F688. [Google Scholar] [CrossRef]
- Nuransoy, A.; Beytur, A.; Polat, A.; Samdanci, E.; Sagir, M.; Parlakpinar, H. Protective effect of sitagliptin against renal ischemia reperfusion injury in rats. Ren. Fail. 2015, 37, 687–693. [Google Scholar] [CrossRef] [Green Version]
- Chang, M.W.; Chen, C.H.; Chen, Y.C.; Wu, Y.C.; Zhen, Y.Y.; Leu, S.; Tsai, T.H.; Ko, S.F.; Sung, P.H.; Yang, C.C.; et al. Sitagliptin protects rat kidneys from acute ischemia-reperfusion injury via upregulation of GLP-1 and GLP-1 receptors. Acta Pharmacol. Sin. 2015, 36, 119–130. [Google Scholar] [CrossRef] [Green Version]
- El-Sahar, A.E.; Safar, M.M.; Zaki, H.F.; Attia, A.S.; Ain-Shoka, A.A. Sitagliptin attenuates transient cerebral ischemia/reperfusion injury in diabetic rats: Implication of the oxidative-inflammatory-apoptotic pathway. Life Sci. 2015, 126, 81–86. [Google Scholar] [CrossRef]
- Abdel-Aziz, A.M.; Naguib Abdel Hafez, S.M. Sitagliptin protects male albino rats with testicular ischaemia/reperfusion damage: Modulation of VCAM-1 and VEGF-A. Andrologia 2020, 52, e13472. [Google Scholar] [CrossRef]
- Khedr, R.M.; Ahmedt, A.A.E.; Kamel, R.; Raafat, E.M. Sitagliptin attenuates intestinal ischemia/reperfusion injury via cAMP/PKA, PI3K/Akt pathway in a glucagon-like peptide 1 receptor-dependent manner. Life Sci. 2018, 211, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Khedr, R.; Ahmed, A.; Kamel, R.; Rafaat, E. Antioxidant effects of sitagliptin in a rat model of intestinal ischemia/reperfusion injury. J. Adv. Pharm. Res. 2021, 5, 234–240. [Google Scholar] [CrossRef]
- Hata, A.N.; Breyer, R.M. Pharmacology and signaling of prostaglandin receptors: Multiple roles in inflammation and immune modulation. Pharmacol. Ther. 2004, 103, 147–166. [Google Scholar] [CrossRef]
- Hossain, M.A.; Wakabayashi, H.; Izuishi, K.; Okano, K.; Yachida, S.; Maeta, H. The role of prostaglandins in liver ischemia-reperfusion injury. Curr. Pharm. Des. 2006, 12, 2935–2951. [Google Scholar] [CrossRef]
- Nakamura, K.; Kageyama, S.; Ito, T.; Hirao, H.; Kadono, K.; Aziz, A.; Dery, K.J.; Everly, M.J.; Taura, K.; Uemoto, S.; et al. Antibiotic pretreatment alleviates liver transplant damage in mice and humans. J. Clin. Investig. 2019, 129, 3420–3434. [Google Scholar] [CrossRef] [Green Version]
- Qiu, H.; Liu, J.Y.; Wei, D.; Li, N.; Yamoah, E.N.; Hammock, B.D.; Chiamvimonvat, N. Cardiac-generated prostanoids mediate cardiac myocyte apoptosis after myocardial ischaemia. Cardiovasc. Res. 2012, 95, 336–345. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Yang, B.; Wang, J.; Zhao, L.; Luo, W.; Jiang, Q.; Yang, J. Time course change of COX2-PGI2/TXA2 following global cerebral ischemia reperfusion injury in rat hippocampus. Behav. Brain Funct. 2014, 10, 42. [Google Scholar] [CrossRef] [Green Version]
- Pushpakom, S.; Iorio, F.; Eyers, P.A.; Escott, K.J.; Hopper, S.; Wells, A.; Doig, A.; Guilliams, T.; Latimer, J.; McNamee, C.; et al. Drug repurposing: Progress, challenges and recommendations. Nat. Rev. Drug Discov. 2019, 18, 41–58. [Google Scholar] [CrossRef]
- Lee, Y.S.; Jun, H.S. Anti-Inflammatory Effects of GLP-1-Based Therapies beyond Glucose Control. Mediat. Inflamm. 2016, 2016, 3094642. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.H.; Lin, C.C. Sitagliptin attenuates inflammatory responses in lipopolysaccharide-stimulated cardiomyocytes via nuclear factor-κB pathway inhibition. Exp. Ther. Med. 2016, 11, 2609–2615. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Li, J.J.; Li, S.N.; Feng, J.; Liu, T.; Wang, F.; Dai, W.Q.; Xia, Y.J.; Lu, J.; Zhou, Y.Q.; et al. 15-Deoxy-Δ12,14-prostaglandin J2 alleviates hepatic ischemia-reperfusion injury in mice via inducing antioxidant response and inhibiting apoptosis and autophagy. Acta Pharmacol. Sin. 2017, 38, 672–687. [Google Scholar] [CrossRef] [Green Version]
- Sharawy, M.H.; El-Kashef, D.H.; Shaaban, A.A.; El-Agamy, D.S. Anti-fibrotic activity of sitagliptin against concanavalin A-induced hepatic fibrosis. Role of Nrf2 activation/NF-κB inhibition. Int. Immunopharmacol. 2021, 100, 108088. [Google Scholar] [CrossRef]
- Chang, X.M.; Xiao, F.; Pan, Q.; Wang, X.X.; Guo, L.X. Sitagliptin attenuates endothelial dysfunction independent of its blood glucose controlling effect. Korean J. Physiol. Pharmacol. 2021, 25, 425–437. [Google Scholar] [CrossRef]
- Gilbert, M.P.; Pratley, R.E. GLP-1 Analogs and DPP-4 Inhibitors in Type 2 Diabetes Therapy: Review of Head-to-Head Clinical Trials. Front. Endocrinol. 2020, 11, 178. [Google Scholar] [CrossRef]
- Matsubara, M.; Kanemoto, S.; Leshnower, B.G.; Albone, E.F.; Hinmon, R.; Plappert, T.; Gorman, J.H., 3rd; Gorman, R.C. Single dose GLP-1-Tf ameliorates myocardial ischemia/reperfusion injury. J. Surg Res. 2011, 165, 38–45. [Google Scholar] [CrossRef] [Green Version]
- Abdel-latif, R.G.; Heeba, G.H.; Taye, A.; Khalifa, M.M.A. Lixisenatide, a novel GLP-1 analog, protects against cerebral ischemia/reperfusion injury in diabetic rats. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2018, 391, 705–717. [Google Scholar] [CrossRef] [PubMed]
- Joo, K.W.; Kim, S.; Ahn, S.Y.; Chin, H.J.; Chae, D.W.; Lee, J.; Han, J.S.; Na, K.Y. Dipeptidyl peptidase IV inhibitor attenuates kidney injury in rat remnant kidney. BMC Nephrol. 2013, 14, 98. [Google Scholar] [CrossRef] [Green Version]
- Gupta, N.A.; Kolachala, V.L.; Jiang, R.; Abramowsky, C.; Romero, R.; Fifadara, N.; Anania, F.; Knechtle, S.; Kirk, A. The glucagon-like peptide-1 receptor agonist Exendin 4 has a protective role in ischemic injury of lean and steatotic liver by inhibiting cell death and stimulating lipolysis. Am. J. Pathol. 2012, 181, 1693–1701. [Google Scholar] [CrossRef] [Green Version]
- Basalay, M.V.; Davidson, S.M.; Yellon, D.M. Neuroprotection in Rats Following Ischaemia-Reperfusion Injury by GLP-1 Analogues-Liraglutide and Semaglutide. Cardiovasc. Drugs Ther. 2019, 33, 661–667. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Verma, S. Mechanisms by Which Glucagon-Like-Peptide-1 Receptor Agonists and Sodium-Glucose Cotransporter-2 Inhibitors Reduce Cardiovascular Risk in Adults with Type 2 Diabetes Mellitus. Can. J. Diabetes 2020, 44, 93–102. [Google Scholar] [CrossRef] [Green Version]
- Jin, T.; Weng, J. Hepatic functions of GLP-1 and its based drugs: Current disputes and perspectives. Am. J. Physiol. Endocrinol. Metab. 2016, 311, E620–E627. [Google Scholar] [CrossRef] [PubMed]
- Müller, T.D.; Finan, B.; Bloom, S.R.; D’Alessio, D.; Drucker, D.J.; Flatt, P.R.; Fritsche, A.; Gribble, F.; Grill, H.J.; Habener, J.F.; et al. Glucagon-like peptide 1 (GLP-1). Mol. Metab. 2019, 30, 72–130. [Google Scholar] [CrossRef] [PubMed]
- Klune, J.R.; Bartels, C.; Luo, J.; Yokota, S.; Du, Q.; Geller, D.A. IL-23 mediates murine liver transplantation ischemia-reperfusion injury via IFN-γ/IRF-1 pathway. Am. J. Physiol. Gastrointest. Liver Physiol. 2018, 315, G991–G1002. [Google Scholar] [CrossRef] [PubMed]
- Couvineau, A.; Laburthe, M. VPAC receptors: Structure, molecular pharmacology and interaction with accessory proteins. Br. J. Pharmacol. 2012, 166, 42–50. [Google Scholar] [CrossRef]
- Ji, H.; Zhang, Y.; Liu, Y.; Shen, X.D.; Gao, F.; Nguyen, T.T.; Busuttil, R.W.; Waschek, J.A.; Kupiec-Weglinski, J.W. Vasoactive intestinal peptide attenuates liver ischemia/reperfusion injury in mice via the cyclic adenosine monophosphate-protein kinase a pathway. Liver Transpl. 2013, 19, 945–956. [Google Scholar] [CrossRef] [Green Version]
- Jungraithmayr, W.; De Meester, I.; Matheeussen, V.; Inci, I.; Augustyns, K.; Scharpé, S.; Weder, W.; Korom, S. Inhibition of CD26/DPP IV attenuates ischemia/reperfusion injury in orthotopic mouse lung transplants: The pivotal role of vasoactive intestinal peptide. Peptides 2010, 31, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Erden, S.H.; Tunçel, N.; Aydyn, Y.; Sahintürk, V.; Koşar, M.; Tunçel, M. The effect of vasoactive intestinal peptide (VIP) and inhibition of nitric oxide on renal tissue injury of rats exposed to hemorrhagic ischemia and retransfusion: A possible interaction mechanism among mast cells and tissue histamine. Ann. N. Y. Acad. Sci. 1998, 865, 570–581. [Google Scholar] [CrossRef] [PubMed]
- Matheeussen, V.; Jungraithmayr, W.; De Meester, I. Dipeptidyl peptidase 4 as a therapeutic target in ischemia/reperfusion injury. Pharmacol. Ther. 2012, 136, 267–282. [Google Scholar] [CrossRef] [PubMed]
- Ochiai, K.; Muramatsu, H.; Yamamoto, S.; Ando, H.; Muramatsu, T. The role of midkine and pleiotrophin in liver regeneration. Liver Int. 2004, 24, 484–491. [Google Scholar] [CrossRef]
- Yazihan, N.; Kocak, M.K.; Akcil, E.; Erdem, O.; Sayal, A. Role of midkine in cadmium-induced liver, heart and kidney damage. Hum. Exp. Toxicol. 2011, 30, 391–397. [Google Scholar] [CrossRef]
- Miao, J.; Ding, M.; Zhang, A.; Xiao, Z.; Qi, W.; Luo, N.; Di, W.; Tao, Y.; Fang, Y. Pleiotrophin promotes microglia proliferation and secretion of neurotrophic factors by activating extracellular signal-regulated kinase 1/2 pathway. Neurosci. Res. 2012, 74, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Christman, K.L.; Fang, Q.; Kim, A.J.; Sievers, R.E.; Fok, H.H.; Candia, A.F.; Colley, K.J.; Herradon, G.; Ezquerra, L.; Deuel, T.F.; et al. Pleiotrophin induces formation of functional neovasculature invivo. Biochem. Biophys. Res. Commun. 2005, 332, 1146–1152. [Google Scholar] [CrossRef] [PubMed]
- Horiba, M.; Kadomatsu, K.; Yasui, K.; Lee, J.K.; Takenaka, H.; Sumida, A.; Kamiya, K.; Chen, S.; Sakuma, S.; Muramatsu, T.; et al. Midkine plays a protective role against cardiac ischemia/reperfusioninjury through a reduction of apoptotic reaction. Circulation 2006, 114, 1713–1720. [Google Scholar] [CrossRef] [Green Version]
- Sato, W.; Kadomatsu, K.; Yuzawa, Y.; Muramatsu, H.; Hotta, N.; Matsuo, S.; Muramatsu, T. Midkine is involved in neutrophil infiltrationinto the tubulointerstitium in ischemic renal injury. J. Immunol. 2001, 167, 3463–3469. [Google Scholar] [CrossRef] [Green Version]
- Weckbach, L.T.; Groesser, L.; Borgolte, J.; Pagel, J.I.; Pogoda, F.; Schymeinsky, J.; Muller-Hocker, J.; Shakibaei, M.; Muramatsu, T.; Deindl, E.; et al. Midkine acts as proangiogenic cytokine in hypoxia-induced angiogenesis. Am. J. Physiol. Heart Circ. Physiol. 2012, 303, H429–H438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beckers, P.A.J.; Gielis, J.F.; Van Schil, P.E.; Adriaensen, D. Lung ischemia reperfusion injury: The therapeutic role of dipeptidyl peptidase 4 inhibition. Ann. Transl. Med. 2017, 5, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liepelt, A.; Tacke, F. Stromal cell-derived factor-1 (SDF-1) as a target in liver diseases. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 311, G203–G209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chetram, M.A.; Hinton, C.V. ROS-mediated regulation of CXCR4 in cancer. Front. Biol. 2013, 8, 273–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, H.M.; Peng, K.Y.; Chan, C.K.; Sun, C.Y.; Chen, Y.Y.; Chang, H.M.; Huang, C.L.; Liu, P.C.; Chen, P.Y.; Wang, K.C.; et al. FGF23 ameliorates ischemia-reperfusion induced acute kidney injury via modulation of endothelial progenitor cells: Targeting SDF-1/CXCR4 signaling. Cell Death Dis. 2021, 12, 409. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Chemaly, E.; Liang, L.; Kho, C.; Lee, A.; Park, J.; Altman, P.; Schecter, A.D.; Hajjar, R.J.; Tarzami, S.T. Effects of CXCR4 gene transfer on cardiac function after ischemia-reperfusion injury. Am. J. Pathol. 2010, 176, 1705–1715. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Fan, W.; Wang, H.; Bao, L.; Li, G.; Li, T.; Song, S.; Li, H.; Hao, J.; Sun, J. Resveratrol Protects PC12 Cell against 6-OHDA Damage via CXCR4 Signaling Pathway. Evid. Based Complement. Alternat. Med. 2015, 2015, 730121. [Google Scholar] [CrossRef] [Green Version]
- Wilson, G.C.; Freeman, C.M.; Kuethe, J.W.; Quillin, R.C., 3rd; Nojima, H.; Schuster, R.; Blanchard, J.; Edwards, M.J.; Caldwell, C.C.; Lentsch, A.B. CXC chemokine receptor-4 signaling limits hepatocyte proliferation after hepatic ischemia-reperfusion in mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 308, G702–G709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mo, H.; Wu, Q.; Miao, J.; Luo, C.; Hong, X.; Wang, Y.; Tang, L.; Hou, F.F.; Liu, Y.; Zhou, L. C-X-C Chemokine Receptor Type 4 Plays a Crucial Role in Mediating Oxidative Stress-Induced Podocyte Injury. Antioxid. Redox Signal. 2017, 27, 345–362. [Google Scholar] [CrossRef]
- Remm, F.; Kränkel, N.; Lener, D.; Drucker, D.J.; Sopper, S.; Brenner, C. Sitagliptin Accelerates Endothelial Regeneration after Vascular Injury Independent from GLP1 Receptor Signaling. Stem Cells Int. 2018, 2018, 5284963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, G.; Liu, P.; Shi, Y.; Li, S.; Liu, Y.; Zhu, W. Sitagliptin Stimulates Endothelial Progenitor Cells to Induce Endothelialization in Aneurysm Necks Through the SDF-1/CXCR4/NRF2 Signaling Pathway. Front. Endocrinol. 2019, 10, 823. [Google Scholar] [CrossRef] [PubMed]
- Seemann, S.; Lupp, A. Administration of AMD3100 in endotoxemia is associated with pro-inflammatory, pro-oxidative, and pro-apoptotic effects in vivo. J. Biomed. Sci. 2016, 23, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellemans, J.; Vandesompele, J. qPCR data analysis—Unlocking the secret to successful results. In PCR Troubleshooting and Optimization: The Essential Guide, 1st ed.; Kennedy, S., Oswald, N., Eds.; Caister Academic Press: Poole, UK, 2011; pp. 1–13. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite | Median Prostanoid Concentration (pg/mg) (IQR) | p | |||
|---|---|---|---|---|---|
| Control, n = 9 | IR, n = 9 | SIR, n = 10 | Sitagliptin, n = 8 | ||
| 6-ketoPGF1α (PGI2) | 68.0 (72.5) 2,4 | 247.5 (70.5) 1,3 | 121.8 (48.3) 2,4 | 211 (141.5) 1,3 | 0.001 |
| PGE2 | 154.7 (127) 2 | 263.3 (74.4) 1,3 | 184.7 (39) 2 | 225.4 (160) | 0.050 |
| PGF2 | 58.7 (35.5) | 55.1 (17.7) | 57.4 (27.7) | 62.4 (25.8) | 0.962 |
| PGD2 | 571 (849) 4 | 1142 (434) 4 | 1180 (493) 4 | 1555 (144) 1,2,3 | 0.002 |
| 15-deoxy-12,14-PGJ2 (PGD2) | 1.04 (1.47) 3,4 | 1.15 (1.87) 3,4 | 2.88 (2.48) 1,2 | 3.2 (1.23) 1,2 | 0.004 |
| 13,14-dihydro-PGE1 (PGE1) | 0.066 (0.11) 2 | 0.15 (0.12) 1,3 | 0.053 (0.06) 2 | 0.082 (0.10) | 0.043 |
| TXB2 (TXA2) | 6.19 (4.42) | 3.6 (2.85) | 3.44 (5.0) | 4.61 (3.0) | 0.559 |
| LTB4 (LTA4) | 41.9 (102) | 136.2 (66.9) | 90.6 (124.1) | 124.1 (102.5) | 0.495 |
| 6-ketoPGF1α (PGI2)/TXB2 (TXA2) | 15.7 (34.4) 2,4 | 75.2 (72.9) 1,3 | 21.6 (45.4) 2 | 46.3 (39.2) 1 | 0.015 |
| Dependent Variable | Explanator Variables | Regression Coefficient (β), p | rp | VIF | R2; ANOVA |
|---|---|---|---|---|---|
| 6-ketoPGF1α (PGI2) | (Constant) | 100.4 | R2 = 0.832; F = 19.1, p < 0.0001 | ||
| 13,14-dihydro-PGE1 | 321.3, p = 0.003 | 0.53 | 1.07 | ||
| PGE2 | 0.26, p = 0.028 | 0.41 | 1.63 | ||
| Cxcr4 (log) | 56.7, p = 0.022 | 0.42 | 1.32 | ||
| 3-NT | 1.56, p < 0.001 | 0.64 | 1.18 | ||
| Nox4 (log) | −22.0, p = 0.055 | −0.36 | 1.64 | ||
| VIP | −0.73, p = 0.025 | −0.42 | 1.39 | ||
| IL-1β | 0.07, p = 0.002 | 0.55 | 1.21 | ||
| PGE2 | (Constant) | 23.11 | R2 = 0.744; F = 30.0, p < 0.0001 | ||
| 6-ketoPGF1α (PGI2) | 0.33, p < 0.001 | 0.57 | 1.17 | ||
| PGF2 | 1.57, p < 0.001 | 0.63 | 1.26 | ||
| LTB4 | 0.33, p = 0.006 | 0.47 | 1.33 | ||
| PGF2 | (Constant) | 3.10 | R2 = 0.613; F = 26.1, p < 0.0001 | ||
| PGE2 | 0.18, p < 0.001 | 0.69 | 1.05 | ||
| TXB2 (log) | 33.9, p = 0.002 | 0.52 | 1.05 | ||
| PGD2 | (Constant) | 126.7 | R2 = 0.729; F = 20.8, p < 0.0001 | ||
| 15-deoxy-12,14-PGJ2 | 145.5, p < 0.001 | 0.59 | 1.10 | ||
| PGE2 | 2.27, p = 0.007 | 0.46 | 1.40 | ||
| SDF1α | 4.86, p = 0.026 | 0.39 | 1.50 | ||
| Sitagliptin | 405, p = 0.005 | 0.48 | 1.19 | ||
| 15-deoxy-12,14-PGJ2 | (Constant) | 0.20 | R2 = 0.453; F = 13.7, p < 0.001 | ||
| PGD2 | 0.002, p < 0.001 | 0.60 | 1.00 | ||
| Sitagliptin | 1.25, p = 0.006 | 0.45 | 1.00 | ||
| 13,14-dihydro-PGE1 | (Constant) | 0.08 | R2 = 0.161; F = 6.54, p = 0.015 | ||
| IR | 0.07, p = 0.015 | 0.40 | 1.00 | ||
| TXB2 (log) | (Constant) | 0.24 | R2 = 0.279; F = 12.8, p = 0.001 | ||
| PGF2 | 0.01, p = 0.001 | 0.53 | 1.00 | ||
| LTB4 | (Constant) | −42.2 | R2 = 0.624; F = 17.7, p < 0.0001 | ||
| 15-deoxy-12,14-PGJ2 | 17.6, p = 0.002 | 0.50 | 1.02 | ||
| PGE2 | 0.42, p = 0.001 | 0.53 | 1.40 | ||
| SDF1α | 0.72, p = 0.027 | 0.38 | 1.42 |
| Dependent Variable | Explanatory Variables | Regression Coefficient (β), p | rp | VIF | R2; ANOVA |
|---|---|---|---|---|---|
| CXCR4 | (Constant) | 48.1 | R2 = 0.390; F = 10.2, p < 0.001 | ||
| 6-ketoPGF1α (PGI2) | 0.07, p = 0.034 | 0.36 | 1.31 | ||
| IR | 15.8, p = 0.027 | 0.38 | 1.31 | ||
| Cxcr4 (log) | (Constant) | −0.11 | R2 = 0.376; F = 19.9, p < 0.0001 | ||
| IR | 0.50, p < 0.001 | 0.61 | 1.00 | ||
| Sdf1 (log) | (Constant) | 0.002 | R2 = 0.817; F = 46.1, p < 0.0001 | ||
| Nox4 (log) | 0.16, p < 0.001 | 0.60 | 1.13 | ||
| Vpac1 (log) | 0.40, p < 0.001 | 0.65 | 1.57 | ||
| Nampt (log) | 0.62, p < 0.001 | 0.62 | 1.41 | ||
| SDF1α | (Constant) | −12.8 | R2 = 0.436; F = 12.7, p < 0.001 | ||
| LTB4 | 0.16, p = 0.028 | 0.37 | 1.54 | ||
| PGD2 | 0.02, p = 0.030 | 0.37 | 1.54 | ||
| Vpac1 (log) | (Constant) | 0.003 | R2 = 0.893; F = 85.8, p < 0.0001 | ||
| Mdk (log) | 0.38, p < 0.001 | 0.82 | 1.27 | ||
| Nox2 (log) | −0.18, p = 0.043 | −0.36 | 1.13 | ||
| Sdf1 (log) | 0.62, p < 0.001 | 0.83 | 1.24 | ||
| Vpac2 (log) | (Constant) | 0.17 | R2 = 0.231; F = 9.88, p = 0.004 | ||
| IR | −0.78, p = 0.004 | −0.48 | 1.00 | ||
| VIP | (Constant) | 200.0 | R2 = 0.388; F = 6.55, p = 0.002 | ||
| PGE2 | −0.09, p = 0.073 | −0.32 | 1.13 | ||
| 3-BT | −1.95, p = 0.040 | −0.36 | 1.01 | ||
| IR | −23.1, p = 0.021 | −0.40 | 1.12 | ||
| Glp1r (log) | (Constant) | −0.02 | R2 = 0.829; F = 77.3, p < 0.0001 | ||
| Nox1 (log) | 1.02, p < 0.001 | 0.89 | 1.10 | ||
| Nox2 (log) | −0.67, p = 0.030 | −0.37 | 1.10 | ||
| GLP1 | (Constant) | 49.9 | R2 = 0.315; F = 7.37, p = 0.002 | ||
| IFNγ | −0.004, p =0.019 | −0.40 | 1.00 | ||
| IR 1 | 14.4, p =0.006 | 0.47 | 1.00 |
| Gene | Forward Sequence | Reverse Sequence |
|---|---|---|
| Gapdh | TGACTCTACCCACGG-CAAGTTCAA | ACGACATACTCAGCACCAG-CATCA |
| Cxcr4 | GCTGGAGAGCGAGCATTG | TAGATGGTGGGCAGGAAGATCC |
| Sdf1 | CTCAACACTCCAAACTGTGCCC | GTCCAGGTACTCTTGGATCCAC |
| Pac1 | GGCTGTGCTGAGGCTCTATTTTG | AGGATGATGATGATGCCGATGA |
| Vpac1 | GATGTGGGACAACCTCACCTG | TAACCATGAATGGGGGCAAAC |
| Vpac2 | GGTGAGCAGCATCCACCCAG | TCACTAGTGCAGTTTTTGCTTA |
| Glp1r | GGGTATCTGGCTGCATAAGGACAAC | AAGGATGGCTGAAGCGATGAC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krzystek-Korpacka, M.; Fleszar, M.G.; Fortuna, P.; Gostomska-Pampuch, K.; Lewandowski, Ł.; Piasecki, T.; Kosyk, B.; Szeląg, A.; Trocha, M. Modulation of Prostanoids Profile and Counter-Regulation of SDF-1α/CXCR4 and VIP/VPAC2 Expression by Sitagliptin in Non-Diabetic Rat Model of Hepatic Ischemia-Reperfusion Injury. Int. J. Mol. Sci. 2021, 22, 13155. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222313155
Krzystek-Korpacka M, Fleszar MG, Fortuna P, Gostomska-Pampuch K, Lewandowski Ł, Piasecki T, Kosyk B, Szeląg A, Trocha M. Modulation of Prostanoids Profile and Counter-Regulation of SDF-1α/CXCR4 and VIP/VPAC2 Expression by Sitagliptin in Non-Diabetic Rat Model of Hepatic Ischemia-Reperfusion Injury. International Journal of Molecular Sciences. 2021; 22(23):13155. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222313155
Chicago/Turabian StyleKrzystek-Korpacka, Małgorzata, Mariusz G. Fleszar, Paulina Fortuna, Kinga Gostomska-Pampuch, Łukasz Lewandowski, Tomasz Piasecki, Bogna Kosyk, Adam Szeląg, and Małgorzata Trocha. 2021. "Modulation of Prostanoids Profile and Counter-Regulation of SDF-1α/CXCR4 and VIP/VPAC2 Expression by Sitagliptin in Non-Diabetic Rat Model of Hepatic Ischemia-Reperfusion Injury" International Journal of Molecular Sciences 22, no. 23: 13155. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222313155