
Influence of Risk Factors for Male Infertility on Sperm Protein Composition

 , and
, and
Abstract
:1. Introduction
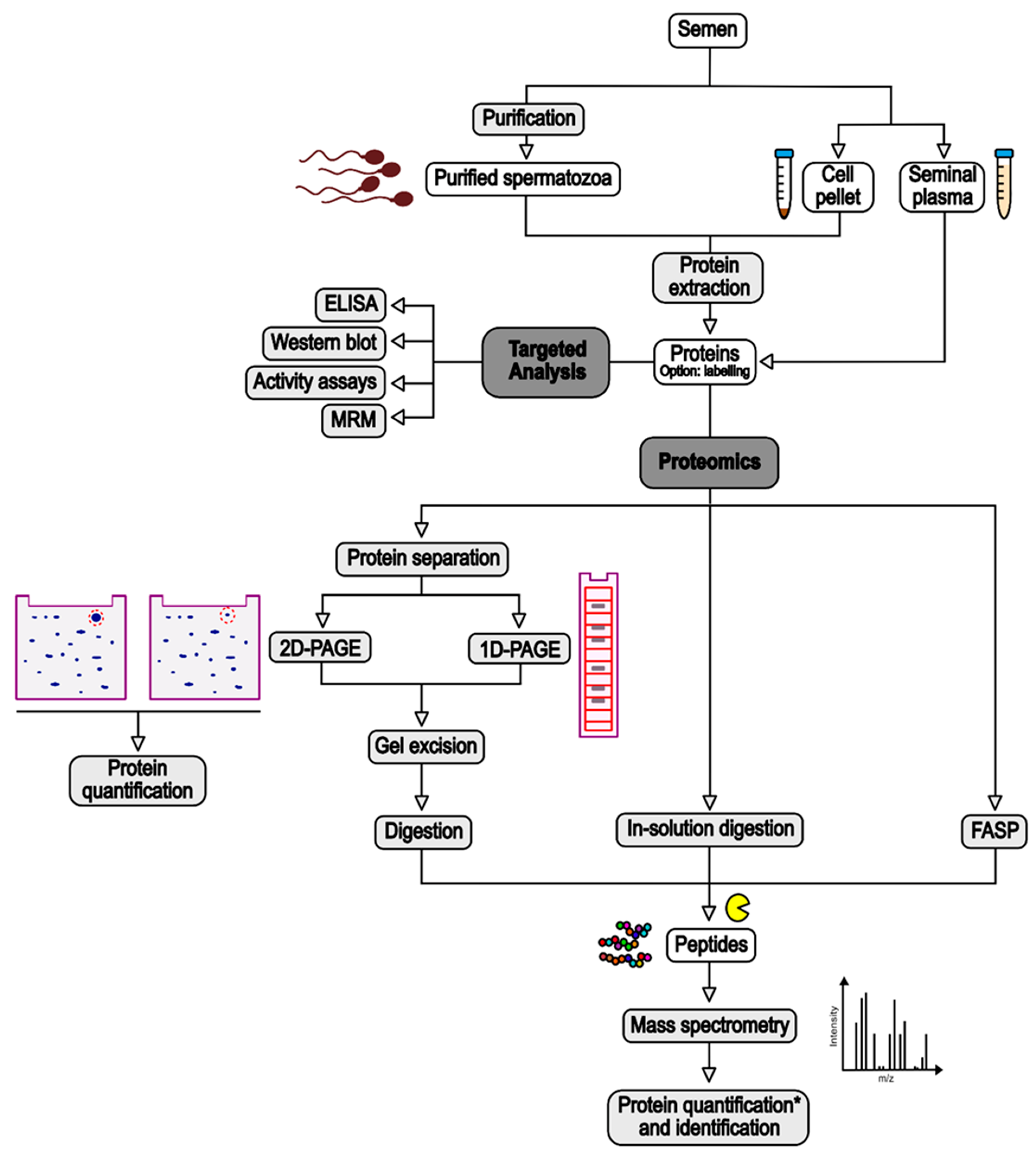
2. Methods
3. Experimental Strategies for Identification of Sperm Proteins Deregulated by Risk Factors
3.1. Sample Preparation
3.2. Proteomics
3.3. Targeted Analyses
4. Impact of Infertility Risk Factors on Sperm Proteins
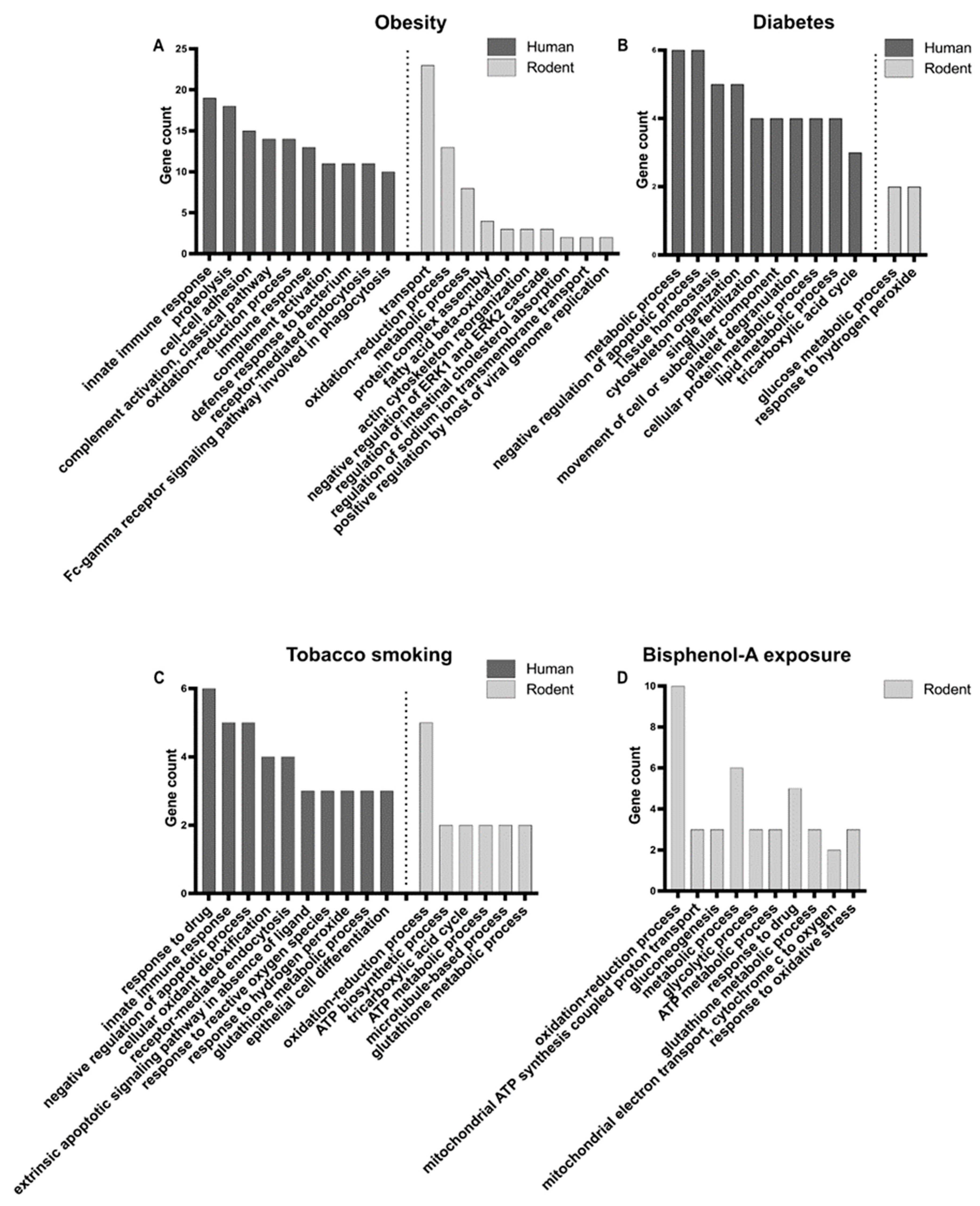
4.1. Obesity
4.2. Diabetes
4.3. Tobacco Smoking
4.4. Exposure to Bisphenol-A
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wistuba, J.; Stukenbrog, J.B.; Luetjens, C.M. Mammalian spermatogenesis. Funct. Dev. Embryol. 2007, 2, 99–117. [Google Scholar]
- Dimitriadis, F.; Adonakis, G.; Kaponis, A.; Mamoulakis, C.; Takenaka, A.; Sofikitis, N. Pre-Testicular, Testicular, and Post-Testicular Causes of Male Infertility. In Endocrinology of the Testis and Male Reproduction; Simoni, M., Huhtaniemi, I., Eds.; Springer: Cham, Switzerland, 2017; pp. 1–47. [Google Scholar]
- Zegers-Hochschild, F.; Adamson, G.D.; Dyer, S.; Racowsky, C.; de Mouzon, J.; Sokol, R.; Rienzi, L.; Sunde, A.; Schmidt, L.; Cooke, I.D.; et al. The International Glossary on Infertility and Fertility Care. Fertil. Steril. 2017, 108, 393–406. [Google Scholar] [CrossRef] [Green Version]
- Jungwirth, A.; Diemer, T.; Kopa, Z.; Krausz, C.; Minhas, S.; Tournaye, H. EAU Guidelines on Male Infertility. Eur. Urol. 2018, 7, 226–241. [Google Scholar]
- Irvine, D.S. Epidemiology and aetiology of male infertility. Hum. Reprod. 1998, 13, 33–44. [Google Scholar] [CrossRef] [Green Version]
- Hwang, K.; Walters, R.C.; Lipshultz, L.I. Contemporary concepts in the evaluation and management of male infertility. Nat. Reviews. Urol. 2011, 8, 86–94. [Google Scholar] [CrossRef] [Green Version]
- Barratt, C.; Björndahl, L.; De Jonge, C.J.; Lamb, D.J.; Osorio Martini, F.; McLachlan, R.; Oates, R.D.; van der Poel, S.; St John, B.; Sigman, M.; et al. The diagnosis of male infertility: An analysis of the evidence to support the development of global WHO guidance-challenges and future research opportunities. Hum. Reprod. Update 2017, 23, 660–680. [Google Scholar] [CrossRef]
- World Health Organization (WHO). WHO Laboratory Manual for the Examination and Processing of Human Semen, 6th ed.; World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- Nieschlag, E.; Behre, H.M.; Nieschlag, S. Andrology: Male Reproductive Health and Dysfunction; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Barbăroșie, C.; Agarwal, A.; Henkel, R. Diagnostic value of advanced semen analysis in evaluation of male infertility. Andrologia 2021, 53, e13625. [Google Scholar] [CrossRef]
- Hwang, K.; Lamb, D.J. Molecular Mechanisms of Antioxidants in Male Infertility. In Male Infertility; Parekattil, S., Agarwal, A., Eds.; Springer: New York, NY, USA, 2012; pp. 45–54. [Google Scholar]
- Sheehan, M.M.; Ramasamy, R.; Lamb, D.J. Molecular mechanisms involved in varicocele-associated infertility. J. Assist. Reprod. Genet. 2014, 31, 521–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bracke, A.; Peeters, K.; Punjabi, U.; Hoogewijs, D.; Dewilde, S. A search for molecular mechanisms underlying male idiopathic infertility. Reprod. Biomed. Online 2018, 36, 327–339. [Google Scholar] [CrossRef] [Green Version]
- E Silva, K.S.F. Molecular genetics of male infertility: A mini-review. Trends Res. 2018, 1, 1–2. [Google Scholar]
- Ashrafzadeh, A.; Karsani, S.A.; Nathan, S. Mammalian sperm fertility related proteins. Int. J. Med Sci. 2013, 10, 1649–1657. [Google Scholar] [CrossRef] [Green Version]
- Brewis, I.A.; Gadella, B.M. Sperm surface proteomics: From protein lists to biological function. Mol. Hum. Reprod. 2010, 16, 68–79. [Google Scholar] [CrossRef] [Green Version]
- Codina, M.; Estanyol, J.M.; Fidalgo, M.J.; Ballescà, J.L.; Oliva, R. Advances in sperm proteomics: Best-practise methodology and clinical potential. Expert Rev. Proteom. 2015, 12, 255–277. [Google Scholar] [CrossRef]
- Amaral, A.; Castillo, J.; Ramalho-Santos, J.; Oliva, R. The combined human sperm proteome: Cellular pathways and implications for basic and clinical science. Hum. Reprod. Update 2014, 20, 40–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panner Selvam, M.K.; Finelli, R.; Agarwal, A.; Henkel, R. Proteomics and metabolomics—Current and future perspectives in clinical andrology. Andrologia 2020, 53, e13711. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Panner Selvam, M.K.; Baskaran, S. Proteomic Analyses of Human Sperm Cells: Understanding the Role of Proteins and Molecular Pathways Affecting Male Reproductive Health. Int. J. Mol. Sci. 2020, 21, 1621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliva, A.; Spira, A.; Multigner, L. Contribution of environmental factors to the risk of male infertility. Hum. Reprod. 2001, 16, 1768–1776. [Google Scholar] [CrossRef] [PubMed]
- Durairajanayagam, D. Lifestyle causes of male infertility. Arab. J. Urol. 2018, 16, 10–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunson, D.B.; Baird, D.D.; Colombo, B. Increased infertility with age in men and women. Obstet. Gynecol. 2004, 103, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Biedenharn, K.R.; Fedor, J.M.; Agarwal, A. Lifestyle factors and reproductive health: Taking control of your fertility. Reprod. Biol. Endocrinol. 2013, 11, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Mahat, R.K.; Arora, M.; Bhale, D.V.; Holkar, S.; Kumar, S.; Yadav, T. Risk Factors and Causes of Male Infertility—A Review. Biochem. Anal. Biochem. 2016, 5, 271. [Google Scholar]
- Ricci, E.; Viganò, P.; Cipriani, S.; Somigliana, E.; Chiaffarino, F.; Bulfoni, A.; Parazzini, F. Coffee and caffeine intake and male infertility: A systematic review. Nutr. J. 2017, 16, 37. [Google Scholar] [CrossRef] [Green Version]
- Condorelli, R.A.; La Vignera, S.; Mongioì, L.M.; Alamo, A.; Calogero, A.E. Diabetes Mellitus and Infertility: Different Pathophysiological Effects in Type 1 and Type 2 on Sperm Function. Front. Endocrinol. 2018, 9, 268. [Google Scholar] [CrossRef] [Green Version]
- Donkin, I.; Barrès, R. Sperm epigenetics and influence of environmental factors. Molecular Metabilism 2018, 14, 1–11. [Google Scholar] [CrossRef]
- Fariello, R.M.; Pariz, J.R.; Spaine, D.M.; Gozzo, F.C.; Pilau, E.J.; Fraietta, R.; Bertolla, R.P.; Andreoni, C.; Cedenho, A.P. Effect of smoking on the functional aspects of sperm and seminal plasma protein profiles in patients with varicocele. Hum. Reprod. 2012, 27, 3140–3149. [Google Scholar] [CrossRef] [Green Version]
- Antoniassi, M.P.; Belardin, L.B.; Camargo, M.; Intasqui, P.; Carvalho, V.M.; Cardozo, K.; Bertolla, R.P. Seminal plasma protein networks and enriched functions in varicocele: Effect of smoking. Andrologia 2020, 52, e13562. [Google Scholar] [CrossRef]
- Ferigolo, P.C.; Ribeiro de Andrade, M.B.; Camargo, M.; Carvalho, V.M.; Cardozo, K.H.M.; Bertolla, R.P.; Fraietta, R. Sperm functional aspects and enriched proteomic pathways of seminal plasma of adult men with obesity. Andrology 2019, 7, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, M.G.; Silva, K.M.; Aristizabal, V.; Ortiz, P.; Paranzini, C.S.; Melchert, A.; Amaro, J.L.; Souza, F.F. Effects of Obesity and Diabetes on Sperm Cell Proteomics in Rats. J. Proteome Res. 2021, 20, 2628–2642. [Google Scholar] [CrossRef] [PubMed]
- Kriegel, T.M.; Heidenreich, F.; Kettner, K.; Pursche, T.; Hoflack, B.; Grunewald, S.; Poenicke, K.; Glander, H.J.; Paasch, U. Identification of diabetes- and obesity- associated proteomic changes in human spermatozoa by difference gel electrophoresis. Reprod. BioMed Online 2009, 19, 660–670. [Google Scholar] [CrossRef] [Green Version]
- Paasch, U.; Heidenreich, F.; Pursche, T.; Kuhlisch, E.; Kettner, K.; Grunewald, S.; Kratzsch, J.; Dittmar, G.; Glander, H.J.; Hoflack, B.; et al. Identification of increased amounts of eppin protein complex components in sperm cells of diabetic and obese individuals by difference gel electrophoresis. Mol. Cell. Proteom. MCP 2011, 10, M110.007187. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Guo, Y.; Song, N.; Fan, Y.; Li, K.; Teng, X.; Guo, Q.; Ding, Z. Proteomic pattern changes associated with obesity-induced asthenozoospermia. Andrology 2015, 3, 247–259. [Google Scholar] [CrossRef]
- Peng, Y.; Zhao, W.; Qu, F.; Jing, J.; Hu, Y.; Liu, Y.; Ding, Z. Proteomic alterations underlie an association with teratozoospermia in obese mice sperm. Reprod. Biol. Endocrinol. 2019, 17, 82. [Google Scholar] [CrossRef] [Green Version]
- Pini, T.; Parks, J.; Russ, J.; Dzieciatkowska, M.; Hansen, K.C.; Schoolcraft, W.B.; Katz-Jaffe, M. Obesity significantly alters the human sperm proteome, with potential implications for fertility. J. Assist. Reprod. Genet. 2020, 37, 777–787. [Google Scholar] [CrossRef] [PubMed]
- An, T.; Wang, Y.F.; Liu, J.X.; Pan, Y.Y.; Liu, Y.F.; He, Z.C.; Mo, F.F.; Li, J.; Kang, L.H.; Gu, Y.J.; et al. Comparative analysis of proteomes between diabetic and normal human sperm: Insights into the effects of diabetes on male reproduction based on the regulation of mitochondria-related proteins. Mol. Reprod. Dev. 2018, 85, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Xu, W.; Miao, M.; Zhu, Z.; Dai, J.; Chen, Z.; Fang, P.; Wu, J.; Nie, D.; Wang, L.; et al. Alteration of sperm protein profile induced by cigarette smoking. Acta Biochim. Biophys. Sin. 2015, 47, 504–515. [Google Scholar] [CrossRef] [Green Version]
- Antoniassi, M.P.; Intasqui, P.; Camargo, M.; Zylbersztejn, D.S.; Carvalho, V.M.; Cardozo, K.H.; Bertolla, R.P. Analysis of the functional aspects and seminal plasma proteomic profile of sperm from smokers. BJU Int. 2016, 118, 814–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.S.; Kwon, W.S.; Yoon, S.J.; Park, Y.J.; Ryu, B.Y.; Pang, M.G. A novel approach to assessing bisphenol-A hazards using an in vitro model system. BMC Genom. 2016, 17, 577. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.S.; Kwon, W.S.; Karmakar, P.C.; Yoon, S.J.; Ryu, B.Y.; Pang, M.G. Gestational Exposure to Bisphenol A Affects the Function and Proteome Profile of F1 Spermatozoa in Adult Mice. Environ. Health Perspect. 2017, 125, 238–245. [Google Scholar] [CrossRef]
- Rahman, M.S.; Kwon, W.S.; Ryu, D.Y.; Khatun, A.; Karmakar, P.C.; Ryu, B.Y.; Pang, M.G. Functional and Proteomic Alterations of F1 Capacitated Spermatozoa of Adult Mice Following Gestational Exposure to Bisphenol A. J. Proteome Res. 2018, 17, 524–535. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.S.; Lee, J.S.; Kwon, W.S.; Pang, M.G. Sperm proteomics: Road to male fertility and contraception. Int. J. Endocrinol. 2013, 2013, 360986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panner Selvam, M.K.; Agarwal, A.; Dias, T.R.; Martins, A.D.; Samanta, L. Presence of Round Cells Proteins do not Interfere with Identification of Human Sperm Proteins from Frozen Semen Samples by LC-MS/MS. Int. J. Mol. Sci. 2019, 20, 314. [Google Scholar] [CrossRef] [Green Version]
- Panner Selvam, M.K.; Agarwal, A.; Dias, T.R.; Martins, A.D.; Baskaran, S.; Samanta, L. Molecular Pathways Associated with Sperm Biofunction Are Not Affected by the Presence of Round Cell and Leukocyte Proteins in Human Sperm Proteome. J. Proteome Res. 2019, 18, 1191–1197. [Google Scholar] [CrossRef] [PubMed]
- Rundlett, K.L.; Armstrong, D.W. Mechanism of signal suppression by anionic surfactants in capillary electrophoresis-electrospray ionization mass spectrometry. Anal. Chem. 1996, 68, 3493–3497. [Google Scholar] [CrossRef] [PubMed]
- Kachuk, C.; Stephen, K.; Doucette, A. Comparison of sodium dodecyl sulfate depletion techniques for proteome analysis by mass spectrometry. J. Chromatogr. 2015, 1418, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Aslanian, A.; Yates, J.R. Mass spectrometry for proteomics. Curr. Opin. Chem. Biol. 2008, 12, 483–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, P.C.; Noirel, J.; Ow, S.Y.; Fazeli, A. A review of current proteomics technologies with a survey on their widespread use in reproductive biology investigations. Theriogenology 2012, 77, 738–765.e52. [Google Scholar] [CrossRef] [PubMed]
- Verheggen, K.; Raeder, H.; Berven, F.S.; Martens, L.; Barsnes, H.; Vaudel, M. Anatomy and evolution of database search engines-a central component of mass spectrometry based proteomic workflows. Mass Spectrom. Rev. 2020, 39, 292–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palomba, A.; Abbondio, M.; Fiorito, G.; Uzzau, S.; Pagnozzi, D.; Tanca, A. Comparative Evaluation of MaxQuant and Proteome Discoverer MS1-Based Protein Quantification Tools. J. Proteome Res. 2021, 20, 3497–3507. [Google Scholar] [CrossRef] [PubMed]
- El-Melegy, N.T.; Ali, M.E. Apoptotic markers in semen of infertile men: Association with cigarette smoking. Int. Braz. J. Urol. Off. J. Braz. Soc. Urol. 2011, 37, 495–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, W.; Xu, Y.; Liu, Y.; Zhang, Z.; Lu, L.; Ding, Z. Obesity or Overweight, a Chronic Inflammatory Status in Male Reproductive System, Leads to Mice and Human Subfertility. Front. Physiol. 2018, 8, 1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, X.; Jing, X.; Wu, X.; Wang, Z.; Li, Q. Potential effect of smoking on semen quality through DNA damage and the downregulation of Chk1 in sperm. Mol. Med. Rep. 2016, 14, 753–761. [Google Scholar] [CrossRef] [Green Version]
- Butler, J.E. Enzyme-Linked Immunosorbent Assay. J. Immunoass. 2000, 21, 165–209. [Google Scholar] [CrossRef]
- Mahmood, T.; Yang, P.C. Western blot: Technique, theory, and trouble shooting. North Am. J. Med Sci. 2012, 4, 429–434. [Google Scholar]
- Kitteringham, N.R.; Jenkins, R.E.; Lane, C.S.; Elliott, V.L.; Park, B.K. Multiple reaction monitoring for quantitative biomarker analysis in proteomics and metabolomics. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2009, 877, 1229–1239. [Google Scholar] [CrossRef]
- Shi, L.; Zhang, Q.; Xu, B.; Jiang, X.; Dai, Y.; Zhang, C.Y.; Zen, K. Sustained high protein-tyrosine phosphatase 1B activity in the sperm of obese males impairs the sperm acrosome reaction. J. Biol. Chem. 2014, 289, 8432–8441. [Google Scholar] [CrossRef] [Green Version]
- Gerhard, I.; Fröhlich, E.; Eggert-Kruse, W.; Klinga, K.; Runnebaum, B. Relationship of sperm acrosin activity to semen and clinical parameters in infertile patients. Andrologia 1989, 21, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Kumosani, T.A.; Elshal, M.F.; Al-Jonaid, A.A.; Abduljabar, H.S. The influence of smoking on semen quality, seminal microelements and Ca2+-ATPase activity among infertile and fertile men. Clin. Biochem. 2008, 41, 1199–1203. [Google Scholar] [CrossRef]
- Ghaffari, M.A.; Rostami, M. The effect of cigarette smoking on human sperm creatine kinase activity: As an ATP buffering system in sperm. Int. J. Fertil. Steril. 2013, 6, 258–265. [Google Scholar]
- Barbonetti, A.; Castellini, C.; Di Giammarco, N.; Santilli, G.; Francavilla, S.; Francavilla, F. In vitro exposure of human spermatozoa to bisphenol A induces pro-oxidative/apoptotic mitochondrial dysfunction. Reprod. Toxicol. 2016, 66, 61–67. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Obesity and Overweight. 9 June 2021. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 7 July 2021).
- Eisenberg, M.L.; Kim, S.; Chen, Z.; Sundaram, R.; Schisterman, E.F.; Buck Louis, G.M. The relationship between male BMI and waist circumference on semen quality: Data from the LIFE study. Hum. Reprod. 2014, 29, 193–200. [Google Scholar] [CrossRef] [Green Version]
- McPherson, N.; Michelle, L. Male obesity and subfertility, is it really about increased adiposity? Asian J. Androl. 2015, 17, 450–458. [Google Scholar] [CrossRef]
- Cui, X.; Jing, X.; Wu, X.; Yan, M. Protective effect of resveratrol on spermatozoa function in male infertility induced by excess weight and obesity. Mol. Med. Rep. 2016, 14, 4659–4665. [Google Scholar] [CrossRef] [Green Version]
- Kahn, B.E.; Brannigan, R.E. Obesity and male infertility. Curr. Opin. Urol. 2017, 27, 441–445. [Google Scholar] [CrossRef]
- Liu, Y.; Ding, Z. Obesity, a serious etiologic factor for male subfertility in modern society. Reproduction 2017, 154, R123–R131. [Google Scholar] [CrossRef] [Green Version]
- Ramaraju, G.A.; Teppala, S.; Prathigudupu, K.; Kalagara, M.; Thota, S.; Kota, M.; Cheemakurthi, R. Association between obesity and sperm quality. Andrologia 2018, 50, e12888. [Google Scholar] [CrossRef]
- Kashou, A.H.; du Plessis, S.S.; Agarwal, A. The Role of Obesity in ROS Generation and Male Infertility. In Studies on Men's Health and Fertility, Oxidative Stress in Applied Basic Research and Clinical Practice; Agarwal, A., Aitken, R., Alvarez, J., Eds.; Humana Press: Totowa, NJ, USA, 2012; pp. 571–590. [Google Scholar]
- Leisegang, K.; Sengupta, P.; Agarwal, A.; Henkel, R. Obesity and male infertility: Mechanisms and management. Andrologia 2021, 53, e13617. [Google Scholar] [CrossRef] [PubMed]
- Miki, K.; Qu, W.; Goulding, E.H.; Willis, W.D.; Bunch, D.O.; Strader, L.F.; Perreault, S.D.; Eddy, E.M.; O'Brien, D.A. Glyceraldehyde 3-phosphate dehydrogenase-S, a sperm-specific glycolytic enzyme, is required for sperm motility and male fertility. Proc. Natl. Acad. Sci. USA 2004, 101, 16501–16506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Widgren, E.E.; Richardson, R.T.; O’Rand, M.G. Characterization of an eppin protein complex from human semen and spermatozoa. Biol. Reprod. 2007, 77, 476–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, X.; Cui, Y.; Zhang, X.; Lou, J.; Zhou, J.; Bei, H.; Wei, R. Proteomic profile of human spermatozoa in healthy and asthenozoospermic individuals. Reprod. Biol. Endocrinol. 2018, 16, 16. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Jiang, W.; Yu, W.; Niu, X.; Liu, F.; Zhou, T.; Zhang, H.; Li, Y.; Zhu, H.; Zhou, Z.; et al. Proteomics analysis of asthenozoospermia and identification of glucose-6-phosphate isomerase as an important enzyme for sperm motility. J. Proteom. 2019, 208, 103478. [Google Scholar] [CrossRef] [PubMed]
- Cho, H. Protein tyrosine phosphatase 1B (PTP1B) and obesity. Vitam. Horm. 2013, 91, 405–424. [Google Scholar]
- Lampiao, F.; Agarwal, A.; du Plessis, S.S. The role of insulin and leptin in male reproduction. Arch. Med. Sci. 2009, 5, S48–S54. [Google Scholar]
- Leisegang, K.; Bouic, P.J.; Menkveld, R.; Henkel, R.R. Obesity is associated with increased seminal insulin and leptin alongside reduced fertility parameters in a controlled male cohort. Reprod. Biol. Endocrinol. 2014, 12, 34. [Google Scholar] [CrossRef] [Green Version]
- Binder, N.K.; Sheedy, J.R.; Hannan, N.J.; Gardner, D.K. Male obesity is associated with changed spermatozoa Cox4i1 mRNA level and altered seminal vesicle fluid composition in a mouse model. Mol. Hum. Reprod. 2015, 21, 424–434. [Google Scholar] [CrossRef]
- Liu, X.; Li, Q.; Wang, W.; Liu, F. Aberrant expression of sperm specific glycolytic enzymes are associated with poor sperm quality. Mol. Med. Rep. 2019, 19, 2471–2478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Lamirande, E.; Yoshida, K.; Yoshiike, T.M.; Iwamoto, T.; Gagnon, C. Semenogelin, the main protein of semen coagulum, inhibits human sperm capacitation by interfering with the superoxide anion generated during this process. J. Androl. 2001, 22, 672–679. [Google Scholar]
- Thakkar, J.K.; East, J.; Franson, R.C. Modulation of phospholipase A2 activity associated with human sperm membranes by divalent cations and calcium antagonists. Biol. Reprod. 1984, 30, 679–686. [Google Scholar] [CrossRef] [Green Version]
- Fry, M.R.; Ghosh, S.S.; East, J.M.; Franson, R.C. Role of human sperm phospholipase A2 in fertilization: Effects of a novel inhibitor of phospholipase A2 activity on membrane perturbations and oocyte penetration. Biol. Reprod. 1992, 47, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Batruch, I.; Lecker, I.; Kagedan, D.; Smith, C.R.; Mullen, B.J.; Grober, E.; Lo, K.C.; Diamandis, E.P.; Jarvi, K.A. Proteomic analysis of seminal plasma from normal volunteers and post-vasectomy patients identifies over 2000 proteins and candidate biomarkers of the urogenital system. J. Proteome Res. 2011, 10, 941–953. [Google Scholar] [CrossRef]
- Akerlöf, E.; Jörnvall, H.; Slotte, H.; Pousette, A. Identification of apolipoprotein A1 and immunoglobulin as components of a serum complex that mediates activation of human sperm motility. Biochemistry 1991, 30, 8986–8990. [Google Scholar] [CrossRef]
- Jorsaraei, S.G.A.; Shibahara, H.; Hirano, Y.; Suzuki, T.; Tahmasbpour, M.E.; Zeynalzadeh, M.; Suzuki, M. The Leptin concentrations in seminal plasma of men and its relationship to semen parameters. Iran. J. Reprod. Med. 2010, 8, 95–100. [Google Scholar]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxidative Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, P.F.; Sousa, M.; Silva, B.M.; Monteiro, M.P.; Alves, M.G. Obesity, energy balance and spermatogenesis. Reproduction 2017, 153, 173–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, T.R.; Samanta, L.; Agarwal, A.; Pushparaj, P.N.; Panner Selvam, M.K.; Sharma, R. Proteomic Signatures Reveal Differences in Stress Response, Antioxidant Defense and Proteasomal Activity in Fertile Men with High Seminal ROS Levels. Int. J. Mol. Sci. 2019, 20, 203. [Google Scholar] [CrossRef] [Green Version]
- Raad, G.; Hazzouri, M.; Bottini, S.; Trabucchi, M.; Azoury, J.; Grandjean, V. Paternal obesity: How bad is it for sperm quality and progeny health? Basic Clin. Androl. 2017, 27, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization (WHO). Diabetes. 13 April 2021. Available online: https://www.who.int/news-room/fact-sheets/detail/diabetes (accessed on 15 April 2021).
- La Vignera, S.; Condorelli, R.; Vicari, E.; D'Agata, R.; Calogero, A.E. Diabetes mellitus and sperm parameters. J. Androl. 2012, 33, 145–153. [Google Scholar] [CrossRef]
- Baccetti, B.; la Marca, A.; Piomboni, P.; Capitani, S.; Bruni, E.; Petraglia, F.; De Leo, V. Insulin-dependent diabetes in men is associated with hypothalamo-pituitary derangement and with impairment in semen quality. Hum. Reprod. 2002, 17, 2673–2677. [Google Scholar] [CrossRef] [PubMed]
- Agbaje, I.M.; Rogers, D.A.; McVicar, C.M.; McClure, N.; Atkinson, A.B.; Mallidis, C.; Lewis, S.E.M. Insulin dependent diabetes mellitus: Implications for male reproductive function. Hum. Reprod. 2007, 22, 1871–1877. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.K.; Tomarz, S.; Chaudhari, A.R.; Sinqh, R.; Verma, N. Type 2 diabetes mellitus affects male fertility potential. Indian J. Physiol. Pharmacol. 2014, 58, 403–406. [Google Scholar] [PubMed]
- Ding, G.L.; Liu, Y.; Liu, M.E.; Pan, J.X.; Guo, M.X.; Sheng, J.Z.; Huang, H.F. The effects of diabetes on male fertility and epigenetic regulation during spermatogenesis. Asian J. Androl. 2015, 17, 948–953. [Google Scholar]
- Martínez-Heredia, J.; de Mateo, S.; Vidal-Taboada, J.M.; Ballescà, J.L.; Oliva, R. Identification of proteomic differences in asthenozoospermic sperm samples. Hum. Reprod. 2008, 23, 783–791. [Google Scholar] [CrossRef] [Green Version]
- Miki, K.; Willis, W.D.; Brown, P.R.; Goulding, E.H.; Fulcher, K.D.; Eddy, E.M. Targeted disruption of the Akap4 gene causes defects in sperm flagellum and motility. Dev. Biol. 2002, 248, 331–342. [Google Scholar] [CrossRef] [Green Version]
- Keller, B.O.; Sui, J.; Young, A.B.; Whittal, R.M. Interferences and contaminants encountered in modern mass spectrometry. Anal. Chim. Acta 2008, 627, 71–81. [Google Scholar] [CrossRef]
- Zhao, W.; Li, Z.; Ping, P.; Wang, G.; Yuan, X.; Sun, F. Outer dense fibers stabilize the axoneme to maintain sperm motility. J. Cell. Mol. Med. 2018, 22, 1755–1768. [Google Scholar] [CrossRef]
- Dunleavy, J.; O'Bryan, M.K.; Stanton, P.G.; O’Donnell, L. The cytoskeleton in spermatogenesis. Reproduction 2019, 157, R53–R72. [Google Scholar] [CrossRef] [Green Version]
- Sonesson, A.; Malm, J.; Rylander, L.; Giwercman, A.; Hillarp, A. Serum amyloid P component: A new biomarker for low sperm concentration? Asian J. Androl. 2021, 23, 450–455. [Google Scholar] [CrossRef]
- Mountjoy, J.R.; Xu, W.; McLeod, D.; Hyndman, D.; Oko, R. RAB2A: A major subacrosomal protein of bovine spermatozoa implicated in acrosomal biogenesis. Biol. Reprod. 2008, 79, 223–232. [Google Scholar] [CrossRef] [Green Version]
- Kwon, W.S.; Rahman, M.S.; Lee, J.S.; Yoon, S.J.; Park, Y.J.; Pang, M.G. Discovery of predictive biomarkers for litter size in boar spermatozoa. Mol. Cell. Proteom. MCP 2015, 14, 1230–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piomboni, P.; Focarelli, R.; Stendardi, A.; Ferramosca, A.; Zara, V. The role of mitochondria in energy production for human sperm motility. Int. J. Androl. 2012, 35, 109–124. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization (WHO). Tobacco. 26 July 2021. Available online: https://www.who.int/news-room/fact-sheets/detail/tobacco (accessed on 3 August 2021).
- World Health Organization. WHO Global Report on Trends in Prevalence of Tobacco Use 2000–2025, 3rd ed.; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- Dai, J.B.; Wang, Z.X.; Qiao, Z.D. The hazardous effects of tobacco smoking on male fertility. Asian J. Androl. 2015, 17, 954–960. [Google Scholar]
- Harlev, A.; Agarwal, A.; Gunes, S.O.; Shetty, A.; du Plessis, S.S. Smoking and Male Infertility: An Evidence-Based Review. World J. Men’s Health 2015, 33, 143–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunes, S.; Metin Mahmutoglu, A.; Arslan, M.A.; Henkel, R. Smoking-induced genetic and epigenetic alterations in infertile men. Andrologia 2018, 50, e13124. [Google Scholar] [CrossRef]
- Rehman, R.; Zahid, N.; Amjad, S.; Baig, M.; Gazzaz, Z.J. Relationship Between Smoking Habit and Sperm Parameters Among Patients Attending an Infertility Clinic. Front. Physiol. 2019, 10, 1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogl, T.; Eisenblätter, M.; Völler, T.; Zenker, S.; Hermann, S.; van Lent, P.; Faust, A.; Geyer, C.; Petersen, B.; Roebrock, K.; et al. Alarmin S100A8/S100A9 as a biomarker for molecular imaging of local inflammatory activity. Nat. Commun. 2014, 5, 4593. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.P.; Meng, Q.Y.; Wang, Q.; Zhang, L.J.; Mao, Y.L.; Sun, Z.X. Effect of smoking on semen quality of infertile men in Shandong, China. Asian J. Androl. 2000, 2, 143–146. [Google Scholar]
- Kiziler, A.R.; Aydemir, B.; Onaran, I.; Alici, B.; Ozkara, H.; Gulyasar, T.; Akyolcu, M.C. High levels of cadmium and lead in seminal fluid and blood of smoking men are associated with high oxidative stress and damage in infertile subjects. Biol. Trace Elem. Res. 2007, 120, 82–91. [Google Scholar] [CrossRef]
- Elshal, M.F.; El-Sayed, I.H.; Elsaied, M.A.; El-Masry, S.A.; Kumosani, T.A. Sperm head defects and disturbances in spermatozoal chromatin and DNA integrities in idiopathic infertile subjects: Association with cigarette smoking. Clin. Biochem. 2009, 42, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Chen, J.; Liu, D.; Zhou, H.; Xiao, W.; Xia, X.; Huang, Z. Cigarette smoking is associated with human semen quality in synergy with functional NRF2 polymorphisms. Biol. Reprod. 2013, 89, 5. [Google Scholar] [CrossRef]
- Ou, Z.; Wen, Q.; Deng, Y.; Yu, Y.; Chen, Z.; Sun, L. Cigarette smoking is associated with high level of ferroptosis in seminal plasma and affects semen quality. Reprod. Biol. Endocrinol. 2020, 18, 55. [Google Scholar] [CrossRef] [PubMed]
- Aboulmaouahib, S.; Madkour, A.; Kaarouch, I.; Sefrioui, O.; Saadani, B.; Copin, H.; Benkhalifa, M.; Louanjli, N.; Cadi, R. Impact of alcohol and cigarette smoking consumption in male fertility potential: Looks at lipid peroxidation, enzymatic antioxidant activities and sperm DNA damage. Andrologia 2018, 50, e12926. [Google Scholar] [CrossRef] [PubMed]
- El Mulla, K.F.; Köhn, F.M.; El Beheiry, A.H.; Schill, W.B. The effect of smoking and varicocele on human sperm acrosin activity and acrosome reaction. Hum. Reprod. 1995, 10, 3190–3194. [Google Scholar] [CrossRef] [PubMed]
- Ito, C.; Toshimori, K. Acrosome markers of human sperm. Anat. Sci. Int. 2016, 91, 128–142. [Google Scholar] [CrossRef]
- Agarwal, A.; Loughlin, K.R. Acrosin activity in patients with idiopathic infertility. Archives of andrology 1991, 27, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Chaudhury, K.; Das, T.; Chakravarty, B.; Bhattacharyya, A.K. Acrosin activity as a potential marker for sperm membrane characteristics in unexplained male infertility. Fertil. Steril. 2005, 83, 104–109. [Google Scholar] [CrossRef]
- Tavilani, H.; Goodarzi, M.T.; Vaisi-raygani, A.; Salimi, S.; Hassanzadeh, T. Activity of antioxidant enzymes in seminal plasma and their relationship with lipid peroxidation of spermatozoa. Int. Braz. J. Urol. Off. J. Braz. Soc. Urol. 2008, 34, 485–491. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Xu, W.; Dai, J.; Chen, X.; Zhao, X.; Fang, P.; Yang, F.; Tang, M.; Wang, Z.; Wang, L.; et al. The alteration of protein profile induced by cigarette smoking via oxidative stress in mice epididymis. Int. J. Biochem. Cell Biol. 2013, 45, 571–582. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Endocrine Disruptors. Available online: https://ec.europa.eu/info/policies/endocrine-disruptors_en (accessed on 27 September 2021).
- Environmental Protection Agency. What is Endocrine Disruption? Available online: https://www.epa.gov/endocrine-disruption/what-endocrine-disruption (accessed on 27 September 2021).
- Diamanti-Kandarakis, E.; Bourguignon, J.P.; Giudice, L.C.; Hauser, R.; Prins, G.S.; Soto, A.M.; Zoeller, R.T.; Gore, A.C. Endocrine-disrupting chemicals: An Endocrine Society scientific statement. Endocr. Rev. 2009, 30, 293–342. [Google Scholar] [CrossRef]
- Sharma, A.; Mollier, J.; Brocklesby, R.; Caves, C.; Jayasena, C.N.; Minhas, S. Endocrine-disrupting chemicals and male reproductive health. Reprod. Med. Biol. 2020, 19, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Staples, C.A.; Dome, P.B.; Klecka, G.M.; Oblock, S.T.; Harris, L.R. A review of the environmental fate, effects, and exposures of bisphenol A. Chemosphere 1998, 36, 2149–2173. [Google Scholar] [CrossRef]
- Kang, J.H.; Kondo, F.; Katayama, Y. Human exposure to bisphenol A. Toxicology 2006, 226, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Vandenberg, L.N.; Hauser, R.; Marcus, M.; Olea, N.; Welshons, W.V. Human exposure to bisphenol A (BPA). Reprod. Toxicol. 2007, 24, 139–177. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.S.; Kwon, W.S.; Lee, J.S.; Yoon, S.J.; Ryu, B.Y.; Pang, M.G. Bisphenol-A affects male fertility via fertility-related proteins in spermatozoa. Sci. Rep. 2015, 5, 9169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, W.H.; Mohamed, E.A.; Pang, W.K.; Kang, K.H.; Ryu, D.Y.; Rahman, M.S.; Pang, M.G. Effect of endocrine disruptors on the ratio of X and Y chromosome-bearing live spermatozoa. Reprod. Toxicol. 2018, 82, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Kang, H.; Peng, Z.; Wang, H.F.; Weng, S.Q.; Zeng, X.H. Physiologically detectable bisphenol A impairs human sperm functions by reducing protein-tyrosine phosphorylation. Ecotoxicol. Environ. Saf. 2021, 221, 112418. [Google Scholar] [CrossRef] [PubMed]
- Tainaka, H.; Takahashi, H.; Umezawa, M.; Tanaka, H.; Nishimune, Y.; Oshio, S.; Takeda, K. Evaluation of the testicular toxicity of prenatal exposure to bisphenol A based on microarray analysis combined with MeSH annotation. J. Toxicol. Sci. 2012, 37, 539–548. [Google Scholar] [CrossRef] [Green Version]
- Xie, M.; Bu, P.; Li, F.; Lan, S.; Wu, H.; Yuan, L.; Wang, Y. Neonatal bisphenol A exposure induces meiotic arrest and apoptosis of spermatogenic cells. Oncotarget 2016, 7, 10606–10615. [Google Scholar] [CrossRef] [Green Version]
- Murata, M.; Kang, J.H. Bisphenol A (BPA) and cell signaling pathways. Biotechnol. Adv. 2018, 36, 311–327. [Google Scholar] [CrossRef] [PubMed]
- Hengstler, J.G.; Foth, H.; Gebel, T.; Kramer, P.J.; Lilienblum, W.; Schweinfurth, H.; Völkel, W.; Wollin, K.M.; Gundert-Remy, U. Critical evaluation of key evidence on the human health hazards of exposure to bisphenol A. Crit. Rev. Toxicol. 2011, 41, 263–291. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.S.; Pang, W.K.; Ryu, D.Y.; Park, Y.J.; Ryu, B.Y.; Pang, M.G. Multigenerational impacts of gestational bisphenol A exposure on the sperm function and fertility of male mice. J. Hazard. Mater. 2021, 416, 125791. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.S.; Pang, W.K.; Ryu, D.Y.; Park, Y.J.; Pang, M.G. Multigenerational and transgenerational impact of paternal bisphenol A exposure on male fertility in a mouse model. Hum. Reprod. 2020, 35, 1740–1752. [Google Scholar] [CrossRef] [PubMed]
- Selvaraj, V.; Asano, A.; Page, J.L.; Nelson, J.L.; Kothapalli, K.S.; Foster, J.A.; Brenna, J.T.; Weiss, R.S.; Travis, A.J. Mice lacking FABP9/PERF15 develop sperm head abnormalities but are fertile. Dev. Biol. 2010, 348, 177–189. [Google Scholar] [CrossRef] [Green Version]
- Pelloni, M.; Paoli, D.; Majoli, M.; Pallotti, F.; Carlini, T.; Lenzi, A.; Lombardo, F. Molecular study of human sperm RNA: Ropporin and CABYR in asthenozoospermia. J. Endocrinol. Investig. 2018, 41, 781–787. [Google Scholar] [CrossRef]
- Xiao, X.; Yang, W.X. Actin-based dynamics during spermatogenesis and its significance. J. Zhejiang Univ. Sci. B 2007, 8, 498–506. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.J.; Ou, J.X.; Chen, G.W.; Wu, J.P.; Shi, H.J.; Wai-Sum, O.; Martin-DeLeon, P.A.; Chen, H. Does prohibitin expression regulate sperm mitochondrial membrane potential, sperm motility, and male fertility? Antioxid. Redox Signal. 2012, 17, 513–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, X.; Chan, C.; Waters, T.; Chi, L.; Chan, D.; Li, T.C. Lifestyle and demographic factors associated with human semen quality and sperm function. Syst. Biol. Reprod. Med. 2018, 64, 358–367. [Google Scholar] [CrossRef] [Green Version]
- Breitbart, H.; Cohen, G.; Rubinstein, S. Role of actin cytoskeleton in mammalian sperm capacitation and the acrosome reaction. Reproduction 2005, 129, 263–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petit, F.M.; Serres, C.; Bourgeon, F.; Pineau, C.; Auer, J. Identification of sperm head proteins involved in zona pellucida binding. Hum. Reprod. 2013, 28, 852–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, C.; Cardoso, M.F.; Sousa, M.; Viana, P.; Gonçalves, A.; Silva, J.; Barros, A. Quantitative study of caspase-3 activity in semen and after swim-up preparation in relation to sperm quality. Hum. Reprod. 2005, 20, 1307–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammadi, P.; Mesbah-Namin, S.A.; Movahedin, M. Attenuation of aquaporin-3 may be contributing to low sperm motility and is associated with activated caspase-3 in asthenozoospermic individuals. Andrologia 2021, 53, e14119. [Google Scholar] [CrossRef]
- Paranko, J.; Longo, F.; Potts, J.; Krohne, G.; Franke, W.W. Widespread occurrence of calicin, a basic cytoskeletal protein of sperm cells, in diverse mammalian species. Differ. Res. Biol. Divers. 1988, 38, 21–27. [Google Scholar] [CrossRef]
- Rosenberg, M.E.; Silkensen, J. Clusterin: Physiologic and pathophysiologic considerations. Int. J. Biochem. Cell Biol. 1995, 27, 633–645. [Google Scholar] [CrossRef]
- Saewu, A.; Kadunganattil, S.; Raghupathy, R.; Kongmanas, K.; Diaz-Astudillo, P.; Hermo, L.; Tanphaichitr, N. Clusterin in the mouse epididymis: Possible roles in sperm maturation and capacitation. Reproduction 2017, 154, 867–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirchhoff, C.; Pera, I.; Rust, W.; Ivell, R. Major human epididymis-specific gene product, HE3, is the first representative of a novel gene family. Mol. Reprod. Dev. 1994, 37, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Maiorino, M.; Ursini, F. Oxidative stress, spermatogenesis and fertility. Biol. Chem. 2002, 383, 591–597. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, H.; Takeo, T.; Tojo, H.; Sakoh, K.; Berger, T.; Nakagata, N.; Mak, T.W.; Kondoh, G. Lipocalin 2 binds to membrane phosphatidylethanolamine to induce lipid raft movement in a PKA-dependent manner and modulates sperm maturation. Development 2014, 141, 2157–2164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, K.; Meinhardt, A.; Zhang, B.; Grzmil, P.; Adham, I.M.; Hoyer-Fender, S. The Small Heat Shock Protein ODF1/HSPB10 Is Essential for Tight Linkage of Sperm Head to Tail and Male Fertility in Mice. Mol. Cell. Biol. 2012, 32, 216–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klinefelter, G.R.; Welch, J.E.; Perreault, S.D.; Moore, H.D.; Zucker, R.M.; Suarez, J.D.; Roberts, N.L.; Bobseine, K.; Jeffay, S. Localization of the sperm protein SP22 and inhibition of fertility in vivo and in vitro. J. Androl. 2002, 23, 48–63. [Google Scholar] [CrossRef]
- An, C.N.; Jiang, H.; Wang, Q.; Yuan, R.P.; Liu, J.M.; Shi, W.L.; Zhang, Z.Y.; Pu, X.P. Down-regulation of DJ-1 protein in the ejaculated spermatozoa from Chinese asthenozoospermia patients. Fertil. Steril. 2011, 96, 19–23.e2. [Google Scholar] [CrossRef]
- Takakubo, F.; Dahl, H.H. The expression pattern of the pyruvate dehydrogenase E1 alpha subunit genes during spermatogenesis in adult mouse. Exp. Cell Res. 1992, 199, 39–49. [Google Scholar] [CrossRef]
- Leone, M.G.; Haq, H.A.; Saso, L. Lipocalin type prostaglandin D-synthase: Which role in male fertility? Contraception 2002, 65, 293–295. [Google Scholar] [CrossRef]
- Guan, Y.; Leu, N.A.; Ma, J.; Chmátal, L.; Ruthel, G.; Bloom, J.C.; Lampson, M.A.; Schimenti, J.C.; Luo, M.; Wang, P.J. SKP1 drives the prophase I to metaphase I transition during male meiosis. Sci. Adv. 2020, 6, eaaz2129. [Google Scholar] [CrossRef] [Green Version]
- Chiriva-Internati, M.; Gagliano, N.; Donetti, E.; Costa, F.; Grizzi, F.; Franceschini, B.; Albani, E.; Levi-Setti, P.E.; Gioia, M.; Jenkins, M.; et al. Sperm protein 17 is expressed in the sperm fibrous sheath. J. Transl. Med. 2009, 7, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandal, A.; Klotz, K.L.; Shetty, J.; Jayes, F.L.; Wolkowicz, M.J.; Bolling, L.C.; Coonrod, S.A.; Black, M.B.; Diekman, A.B.; Haystead, T.A.; et al. SLLP1, a unique, intra-acrosomal, non-bacteriolytic, c lysozyme-like protein of human spermatozoa. Biol. Reprod. 2003, 68, 1525–1537. [Google Scholar] [CrossRef]
- Sachdev, M.; Mandal, A.; Mulders, S.; Digilio, L.C.; Panneerdoss, S.; Suryavathi, V.; Pires, E.; Klotz, K.L.; Hermens, L.; Herrero, M.B.; et al. Oocyte specific oolemmal SAS1B involved in sperm binding through intra-acrosomal SLLP1 during fertilization. Dev. Biol. 2012, 363, 40–51. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Obesity | |||||||
|---|---|---|---|---|---|---|---|
| References | Species | Investigated Groups | Sperm Preparation | Working Sample | Method for Protein Identification | Method for Protein Quantification | Results * |
| Ferigolo et al. 2019 [31] | Human | 27 obese men (BMI ≥ 33 kg/m2), 20 eutrophic men (18.5 kg/m2 ≤BMI ≤ 25 kg/m2); the samples from 4 men were pooled and 4 pools were obtained per group. No differences in sperm volume, concentration, and progressive motility, but significant differences in non-progressive motility and morphology. | Centrifugation | Seminal plasma | In-solution digestion; LC-MS/MS (hybrid quadrupole-Orbitrap) | Label-free; Maxquant software; Ibaq; all fold changes were considered. | 485 proteins identified; 70 differentially expressed proteins: 50 more abundant, 19 less abundant, and 1 exclusive in the obese group. |
| Carvalho et al. 2021 [32] | Rat | 10 rats fed with a control diet and 10 rats fed with a hyperglycidic diet for 266 days. No differences in sperm concentration but differences in sperm morphology between groups. | Washes with centrifugation | Pellet after centrifugation (no purification) | SDS-PAGE, LC-MS/MS (QTOF) | Label-free, calculation of the emPAI. | 144 proteins identified; 15 differentially expressed proteins: 8 more abundant and 7 less abundant in obese rats. |
| Kriegel et al. 2009 [33] | Human | 5 normospermic men (mean BMI = 22 kg/m2), 2 non-diabetic obese men (mean BMI = 33 kg/m2); 3 semen samples for each. No significant differences in sperm parameters between the 2 groups. | 50–90% gradient | Progressive spermatozoa | 2D-PAGE (DIGE); MALDI-TOF-MS | Fluorescent labelling; image acquisition and identification of differentially expressed proteins with a fluorescence imager. Spots with a fold change set to 2.0 were excised and analysed. | 2700 fluorescent protein spots detected; 9 differentially expressed proteins: 2 more abundant, 6 less abundant, and one more or less abundant according to the gel spot, in obese men. |
| Paasch et al. 2011 [34] | Human | 21 normospermic and clinically healthy men (mean BMI = 22.5 kg/m2), 13 non-diabetic obese men (mean BMI = 34 kg/m2); 3 semen samples for each. Sperm progressive motility and morphology were significantly different between the groups. | 50–90% gradient | Progressive spermatozoa | 2D-PAGE (DIGE); MALDI-TOF-MS | Fluorescent labelling; image acquisition and identification of differentially expressed proteins with a fluorescence imager. Spots with a fold change ≤ −1.6 or ≥1.6 were excised and analysed. | 3187 fluorescent protein spots detected; 7 differentially expressed protein: 6 more abundant and 1 less abundant in obese men. |
| Liu et al. 2015 [35] | Human | 3 normospermic fertile men (mean BMI = 24 kg/m2), 3 obese men (mean BMI = 33.5 kg/m2) with severe asthenozoospermia (normal concentration and morphology, but progressive motility <20%). | 45% Percoll gradient | Spermatozoa purified from seminal plasma | FASP, LC-MS/MS (LTQ-Orbitrap) | Label free; Maxquant software; all fold changes were considered. | 1975 proteins identified; 127 differentially expressed proteins: 22 more abundant and 105 less abundant in obesity-associated asthenozoospermia. |
| Peng et al. 2019 [36] | Mouse | 6 mice fed with control diet and 6 mice fed with high-fat diet for 10 weeks. No differences in sperm concentration but significant differences in sperm motility, progressive motility and morphology. | 45% Percoll gradient | Spermatozoa purified from seminal plasma | FASP, LC-MS/MS (LTQ-Orbitrap) | Label-free; Maxquant software; only proteins with a fold change of ≥1.3 or ≤0.7 were considered. | 1562 proteins identified; 160 differentially expressed proteins: 60 more abundant and 100 less abundant in obese mice. |
| Pini et al. 2020 [37] | Human | 5 men with healthy weight (BMI ≤ 25 kg/m2), 5 obese men (BMI ≥ 33 kg/m2); all normospermic. | 45–90% gradient | Progressive spermatozoa | FASP, LC-MS/MS (hybrid quadrupole-Orbitrap) | Label-free quantification with normalised weighted spectra (NWS); only proteins with a fold change of ≥1.5 or ≤0.5 were considered. | 2034 proteins identified; 27 differentially expressed proteins: 3 more abundant and 24 less abundant in obese men. |
| Diabetes | |||||||
| References | Species | Investigated groups | Sperm Preparation | Working Sample | Method for Protein Identification | Method for Protein Quantification | Results |
| Carvalho et al. 2021 [32] | Rat | 10 control rats and 5 rats injected with streptozotocin to induce diabetes. No differences in sperm concentration but differences in sperm morphology between groups. | Washes with centrifugation | Pellet after centrifugation (no purification) | SDS-PAGE, LC-MS/MS (QTOF) | Label-free, calculation of the emPAI. | 144 proteins identified, 15 differentially expressed proteins: 3 more abundant and 12 less abundant in diabetic men. |
| Kriegel et al. 2009 [33] | Human | 5 normospermic donors, 2 type-1 diabetic patients; 3 semen samples for each. No significant differences in sperm parameters between the 2 groups. | 50–90% gradient | Progressive spermatozoa | 2D-PAGE (DIGE); MALDI-TOF-MS | Fluorescent labelling; Image acquisition and identification of differentially expressed proteins with a fluorescence imager. Spots with a fold change set to 2.0 were excised and analysed. | 2700 fluorescent protein spots detected; 7 differentially expressed proteins: 4 more abundant, 2 less abundant, and one more or less abundant according to the gel spot, in diabetic men. |
| Paasch et al. 2011 [34] | Human | 21 normospermic and clinically healthy individuals, 8 type-1 diabetic individuals, and 7 type-2 diabetic individuals; 3 semen samples for each. Sperm progressive motility and morphology were significantly different between the groups. | 50–90% gradient | Progressive spermatozoa | 2D-PAGE (DIGE); MALDI-TOF-MS | Fluorescent labelling; Image acquisition and identification of differentially expressed proteins with a fluorescence imager. Spots with a fold change ≤−1.6 or ≥1.6 were excised and analysed. | 3187 fluorescent protein spots detected; 8 differentially expressed protein in type-1 diabetic patients: 6 more abundant and 2 less abundant; 39 differentially expressed proteins in type-2 diabetic patients: 12 more abundant, 25 less abundant, and 2 more or less abundant according to the gel spot. |
| An et al. 2018 [38] | Human | 6 healthy men and 6 type-2 diabetic men. Sperm volume and concentration were significantly different between the groups. | Centrifugation | Pellet after centrifugation (no purification) | Gel-free digestion; LC-MS/MS (hybrid quadrupole-Orbitrap) | Itraq labelling; only proteins with a fold change of >1.2 or <0.83 were considered. | 1114 proteins identified; 357 differentially expressed: 38 less abundant and 319 more abundant in diabetic men. |
| Tobacco smoking | |||||||
| References | Species | Investigated groups | Sperm Preparation | Working Sample | Method for Protein Identification | Method for Protein Quantification | Results |
| Chen et al. 2015 [39] | Mouse | 3 mice exposed daily to cigarette smoke for 2 weeks, and 3 non-exposed mice. No differences in sperm motility between the groups. | Swim up | Spermatozoa | 2D-PAGE, MALDI-TOF-MS | Image acquisition and spot density analysis. Spots with a fold change set to 2.0 were excised and analysed. | More than 1000 protein spots detected; 22 differentially expressed proteins: 10 more abundant and 12 less abundant proteins in exposed mice. |
| Antoniassi et al. 2016 [40] | Human | 20 non-smoking normospermic men and 20 smoking patients (≥ 10 cigarettes/day). The samples were distributed into 4 pools per group. No significant differences in sperm parameters between the 2 groups. | Centrifugation | Seminal plasma | In-solution digestion; LC-MS/MS (hybrid quadrupole-Orbitrap) | Label-free; Maxquant software; Ibaq; all fold changes were considered. | 422 proteins identified, 25 differentially expressed proteins: 1 absent, 6 more abundant, and 18 less abundant in smokers. |
| Bisphenol-A exposure | |||||||
| References | Species | Investigated groups | Sperm Preparation | Working Sample | Method for Protein Identification | Method for Protein Quantification | Results |
| Rahman et al. 2016 [41] | Mouse | Spermatozoa from 3 mice were exposed or not to various concentrations (0.0001–100 µM) of BPA. | Swim up | Spermatozoa | 2D-PAGE, ESI-QTOF | Image acquisition and spot density analysis. Spots with significant density changes were excised and analysed. | 399 spots detected; 23 differentially expressed proteins: 8 more abundant and 15 less abundant in spermatozoa exposed to the upper concentration of BPA. |
| Rahman et al. 2017 [42] | Mouse | Male mice (F1; n = 3) generated from females which were orally exposed or not to various concentrations (0.05–50 mg/kg per day) of BPA during pregnancy. | Swim up | Spermatozoa | 2D-PAGE, ESI-QTOF | Image acquisition and spot density analysis. Spots with significant density changes were excised and analysed. | 284 spots detected, 6 differentially expressed proteins: 2 more abundant and 4 less abundant in males generated from females exposed to BPA. |
| Rahman et al. 2018 [43] | Mouse | Male mice (F1; n = 3) generated from females which were orally exposed or not to various concentrations (0.05–50 mg/kg per day) of BPA during pregnancy. | Capacitation, wash and swim up | Capacitated spermatozoa | 2D-PAGE, ESI-QTOF | Image acquisition and spot density analysis. Spots with significant density changes were excised and analysed. | 285 spots detected, 15 differentially expressed proteins: 4 more abundant and 11 less abundant in males generated from females exposed to the upper concentration of BPA. |
| Protein (Gene Name) | Risk Factors | Role in Sperm Fertility | References |
|---|---|---|---|
| Acrosin (ACR) | Diabetes, Tobacco smoking | Acrosomal enzyme involved in acrosome reaction and zone pellucida penetration. | [123] |
| Actin (ACTB) | Obesity, BPA exposure | Cytoskeletal protein, plays a role in sperm motility and the acrosome reaction. | [146,149] |
| Fructose-bisphosphate aldolase A (ALDOA) | Diabetes, Tobacco smoking | Glycolytic enzyme involved in zone pellucida binding. | [150] |
| Apolipoprotein A-I (APOA1) | Obesity, Tobacco smoking | Sterol acceptor in the seminal fluid involved in sperm motility activation. | [87,88] |
| ATP synthase subunit beta (ATP5B) | Obesity, Diabetes | Involved in ATP synthesis during oxidative phosphorylation, which is important to sustain sperm motility. | [108] |
| Caspase 3 (CASP3) | Tobacco smoking, BPA exposure | Functional enzyme of the apoptosis process. Its high expression is related to athenospermia and teratospermia. | [151,152] |
| Calicin (CCIN) | Obesity, Diabetes | Cytoskeletal protein of the sperm head. | [153] |
| Clusterin (CLU) | Obesity, Diabetes | Seminal plasma protein involved in sperm maturation and capacitation. | [154,155] |
| Epididymal secretory protein E3-beta (EDDM3B) | Obesity, Diabetes | Epididymal protein, may be involved in sperm maturation. | [156] |
| Glyceraldehyde-3-phosphate dehydrogenase (GAPDHS) | Obesity, Diabetes, BPA exposure | Glycolytic enzyme involved in sperm motility. | [75,83] |
| Glutathione peroxidase 4 (GPX4) | Tobacco smoking, BPA exposure | Antioxidant enzyme implicated during spermatogenesis and sperm maturation. | [157] |
| Neutrophil gelatinase-associated lipocalin (LCN2) | Obesity, Tobacco smoking | Modulates sperm capacitation. | [158] |
| Lactotransferrin (LTF) | Obesity, Diabetes | Bound to eppin in the EPC and involved in protection of spermatozoa and motility regulation. | [34,76] |
| Outer dense fiber protein 1 (ODF1) | Obesity, Diabetes, Tobacco smoking | Chaperone protein composing the outer dense fibres (ODF) of the sperm tail, involved in sperm structure and motility. | [159] |
| Outer dense fiber protein 2 (ODF2) | Obesity, Tobacco smoking, diabetes, BPA | Major component of the outer dense fibres (ODF) of the sperm tail, involved in sperm motility. | [103] |
| Protein/nucleic acid deglycase DJ-1 (PARK7) | Obesity, Diabetes | Oxidative stress response protein involved in fertilisation process and sperm motility. | [160,161] |
| Pyruvate dehydrogenase E1 component subunit a, testis-specific form, mitochondrial (PDHA2) | Diabetes, Tobacco smoking | Involved in the development of spermatogenic cells. | [162] |
| Prolactin-inducible protein (PIP) | Obesity, Diabetes | May influence sperm viscosity. | [100] |
| Phospholipase A2, membrane associated (PLA2B) | Obesity, Tobacco smoking | Involved in membrane fusion events occurring during the acrosome reaction and fertilisation. | [85] |
| Prostaglandin-H2 D-isomerase (PTGDS) | Obesity, Diabetes | Carrier protein for thyroid hormone and retinoids involved in spermatogenesis and sperm maturation. | [163] |
| Ras-related protein Rab-2A (RAB2A) | Obesity, Diabetes, BPA exposure | Acrosome biogenesis. | [106] |
| Semenogelin-1 (SEMG1) | Obesity, Diabetes | Seminal protein which inhibits premature sperm capacitation. | [76] |
| Alpha-1-antitrypsin (SERPINA1) | Obesity, Diabetes | Serine protease inhibitor involved in the liquefaction cascade of the ejaculated sperm. | [158] |
| S-phase kinase-associated protein 1 (SKP1) | Obesity, Diabetes | Regulates meiosis during spermatogenesis. | [164] |
| Sperm surface protein Sp17 (SPA17) | Diabetes, BPA exposure | Fibrous sheath protein involved in the binding to the oocyte zona pellucida. | [165] |
| Sperm acrosome membrane-associated protein 3 (SPACA3) | Obesity, Diabetes | Acrosomal protein involved in the binding to the oocyte. | [166,167] |
| Triosephosphate isomerase (TPI1) | Diabetes, BPA exposure | Glycolytic enzyme involved in the binding to the oocyte zone pellucida. | [149] |
| Tubulin beta chain (TUBB4B) | Obesity, Diabetes | Cytoskeletal protein of the sperm tail. | [104] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bisconti, M.; Simon, J.-F.; Grassi, S.; Leroy, B.; Martinet, B.; Arcolia, V.; Isachenko, V.; Hennebert, E. Influence of Risk Factors for Male Infertility on Sperm Protein Composition. Int. J. Mol. Sci. 2021, 22, 13164. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222313164
Bisconti M, Simon J-F, Grassi S, Leroy B, Martinet B, Arcolia V, Isachenko V, Hennebert E. Influence of Risk Factors for Male Infertility on Sperm Protein Composition. International Journal of Molecular Sciences. 2021; 22(23):13164. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222313164
Chicago/Turabian StyleBisconti, Marie, Jean-François Simon, Sarah Grassi, Baptiste Leroy, Baptiste Martinet, Vanessa Arcolia, Vladimir Isachenko, and Elise Hennebert. 2021. "Influence of Risk Factors for Male Infertility on Sperm Protein Composition" International Journal of Molecular Sciences 22, no. 23: 13164. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222313164