Restraint of Human Skin Fibroblast Motility, Migration, and Cell Surface Actin Dynamics, by Pannexin 1 and P2X7 Receptor Signaling

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
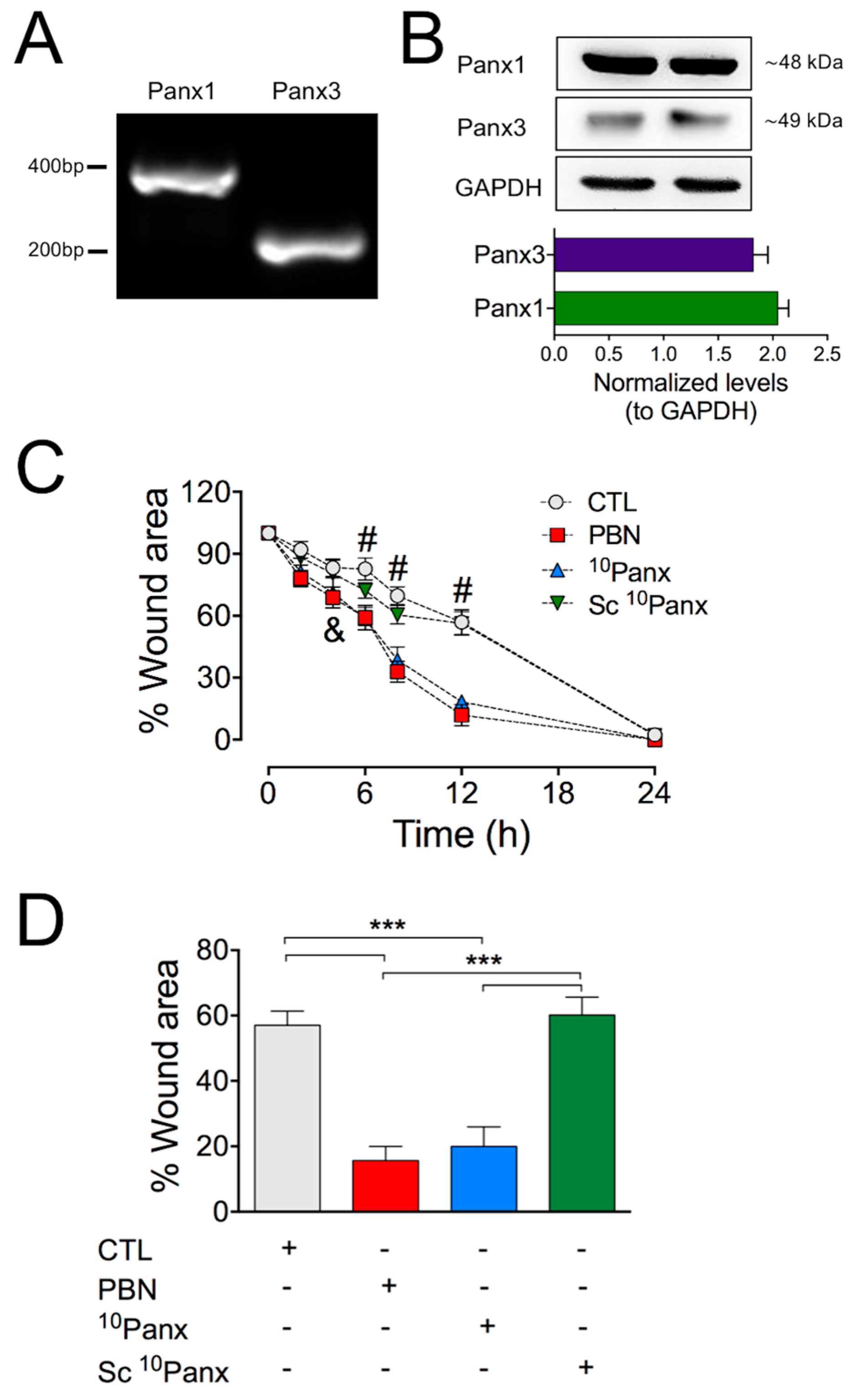
2.1. Inhibition or Blockage of Panx1 Channels Accelerates the In Vitro Wound Healing of HDFs
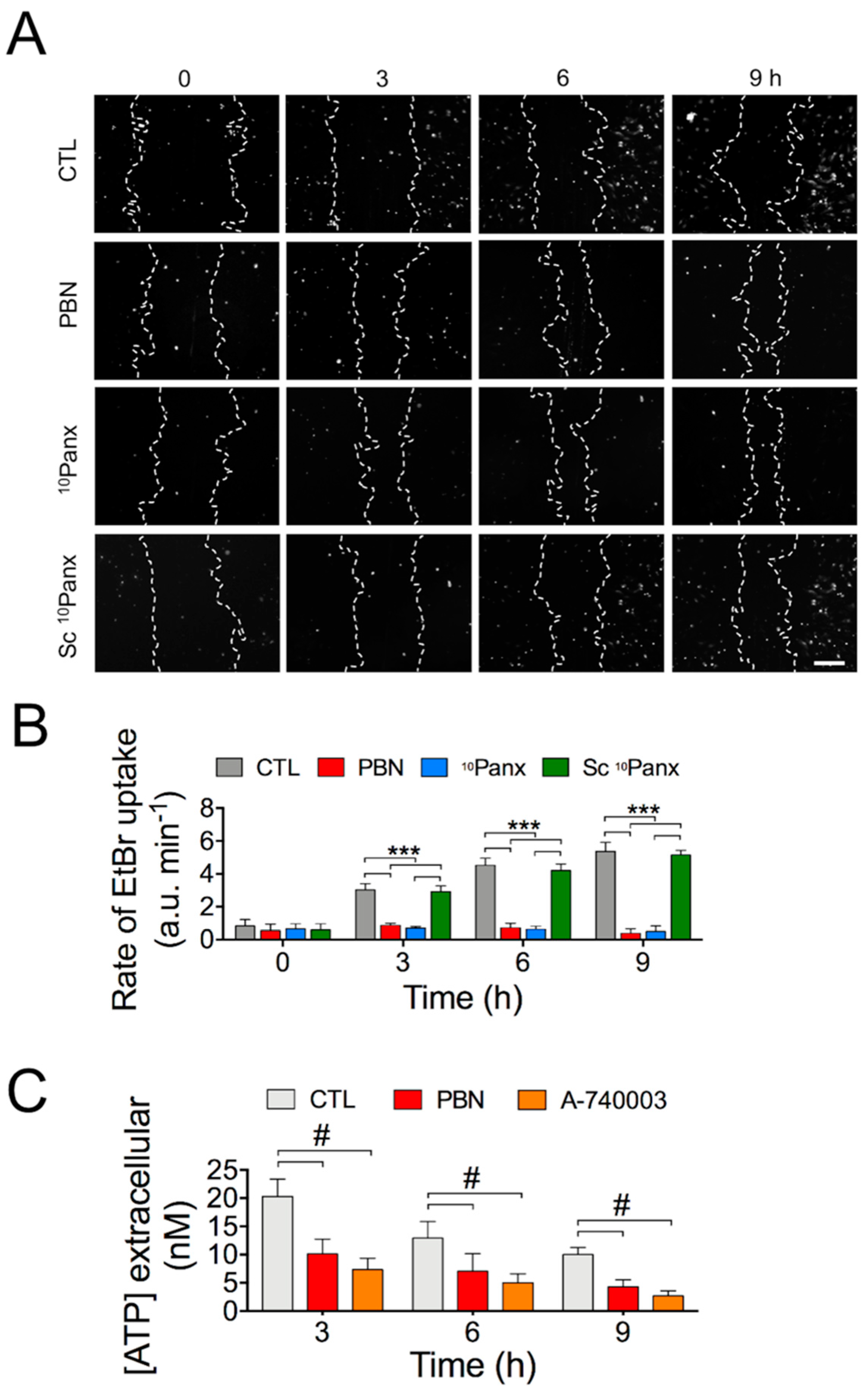
2.2. Activity of Panx1 Channels during Wound Healing in HDF Cultures
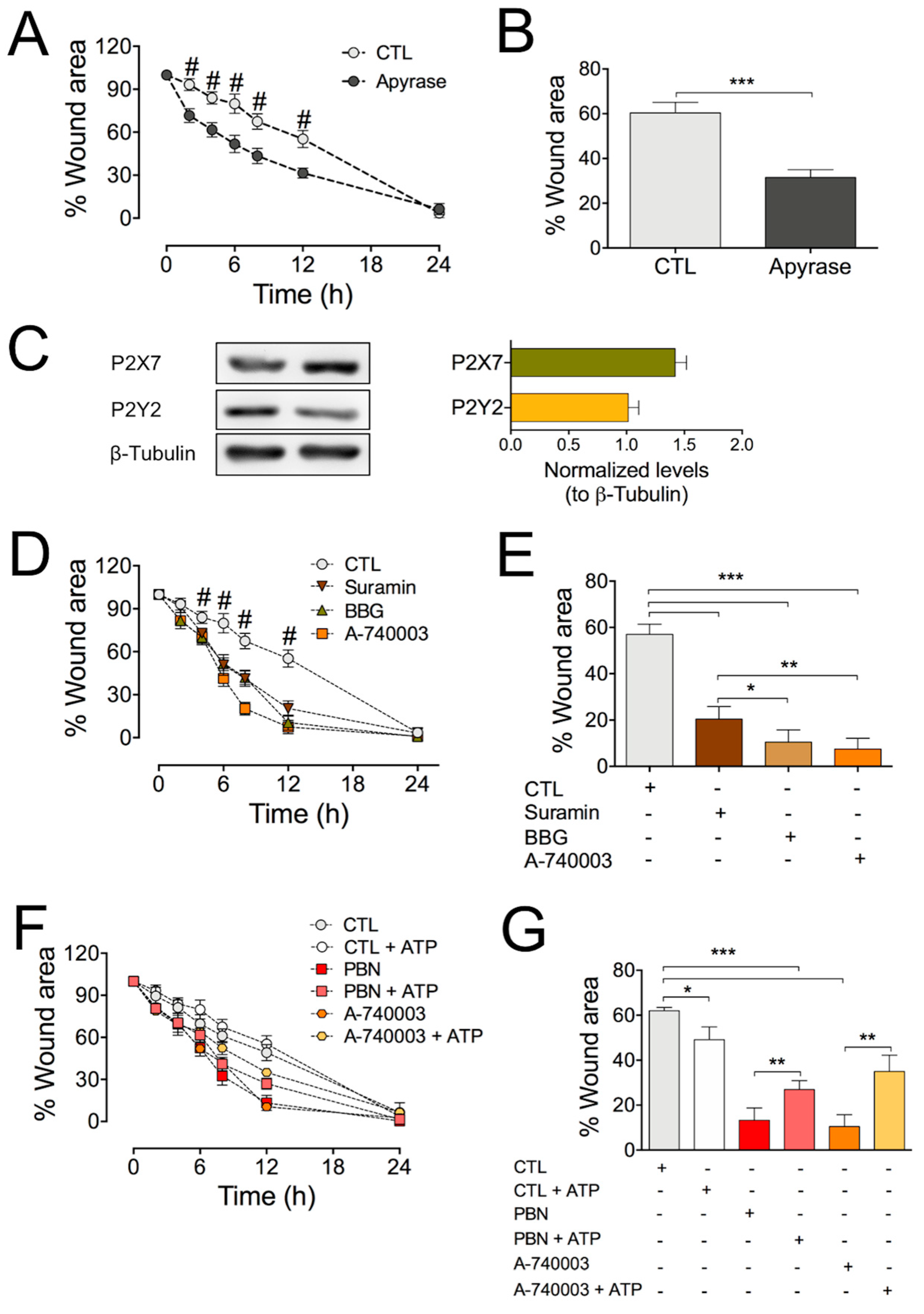
2.3. Blocking the Purinergic P2X7 Receptor Significantly Accelerates Wound Healing in HDF Cultures
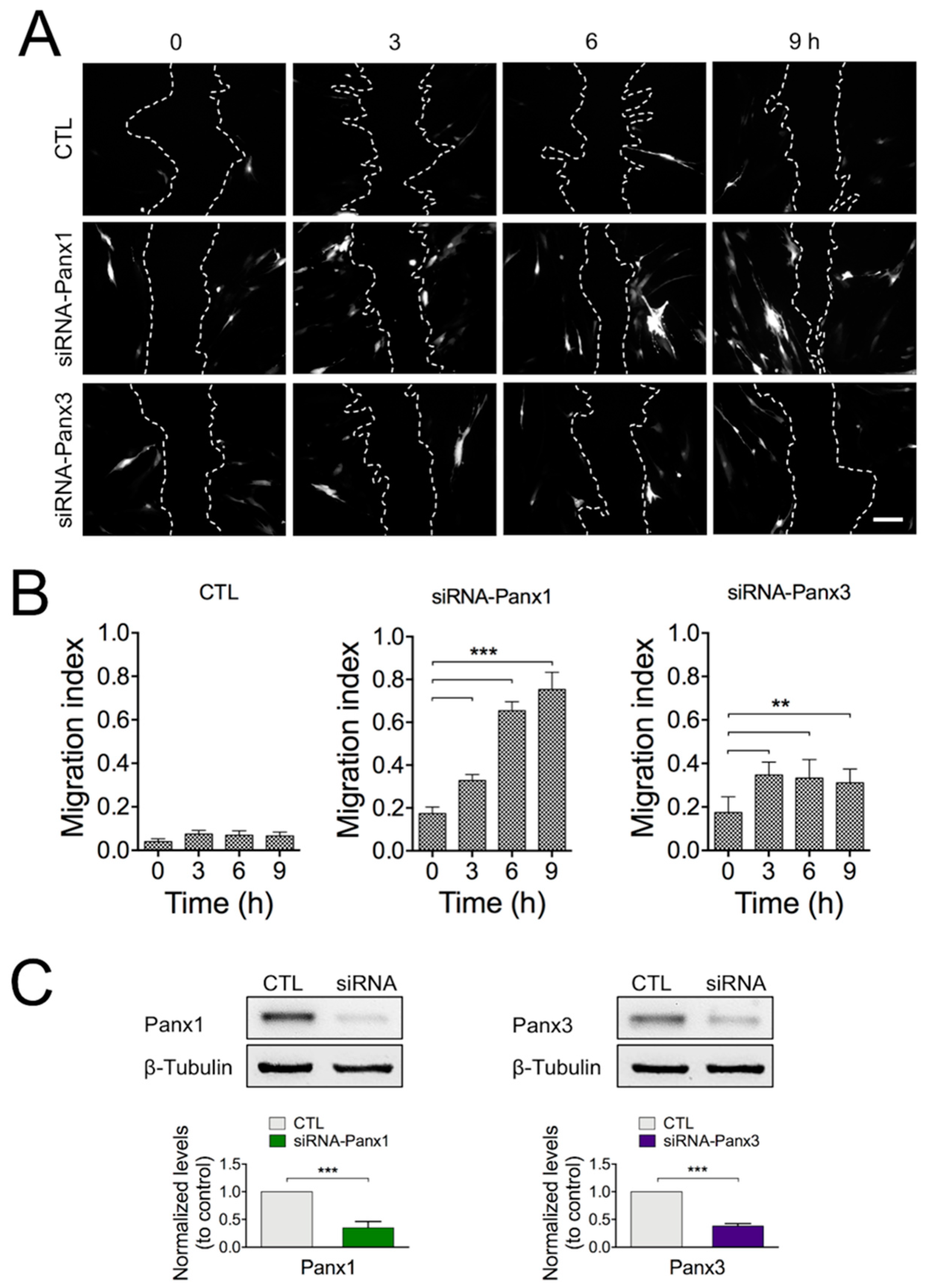
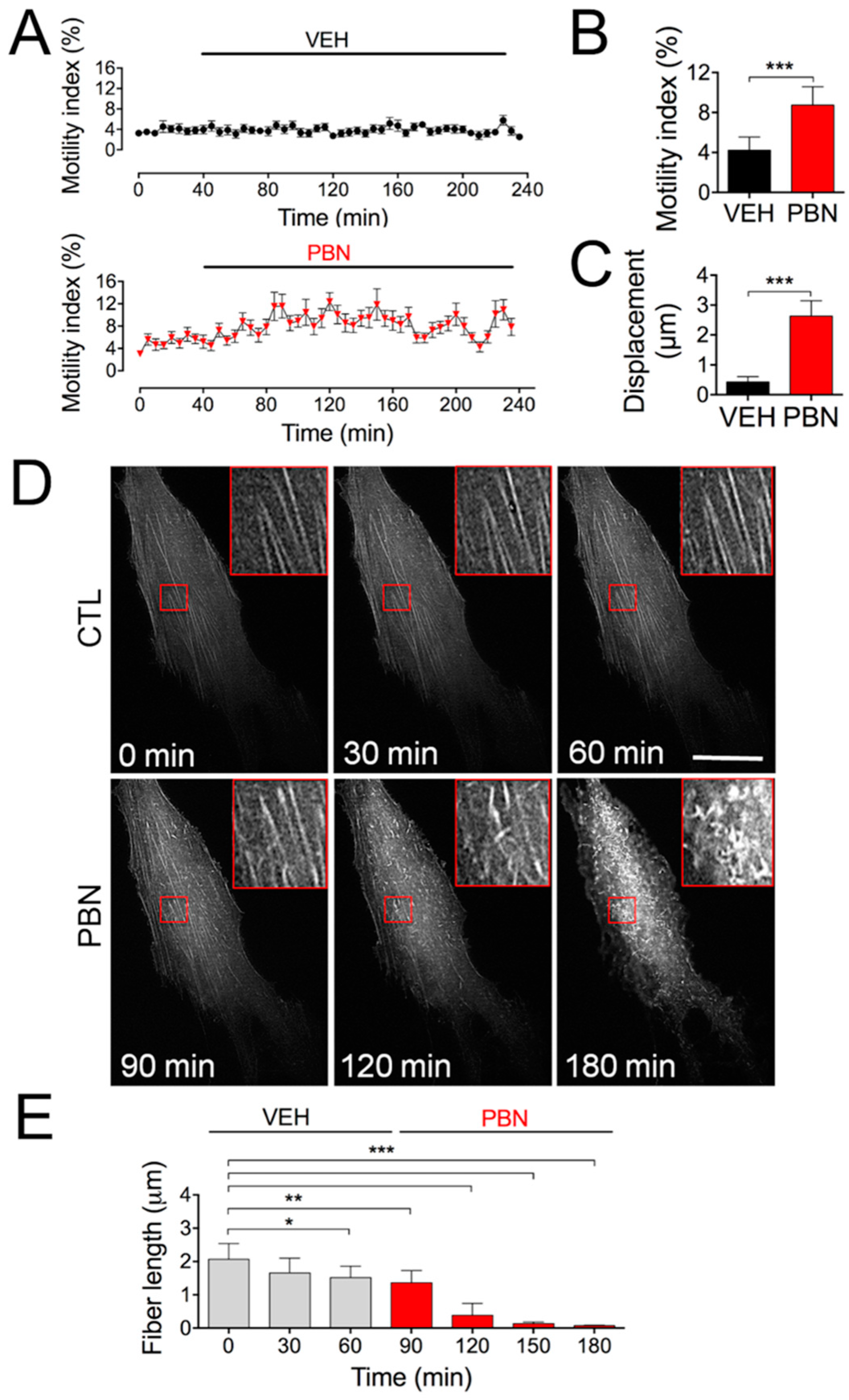
2.4. Blocking Panx1 Channels Increases Cell Motility and Triggers Fast Rearrangements of Cell Surface Actin Filaments
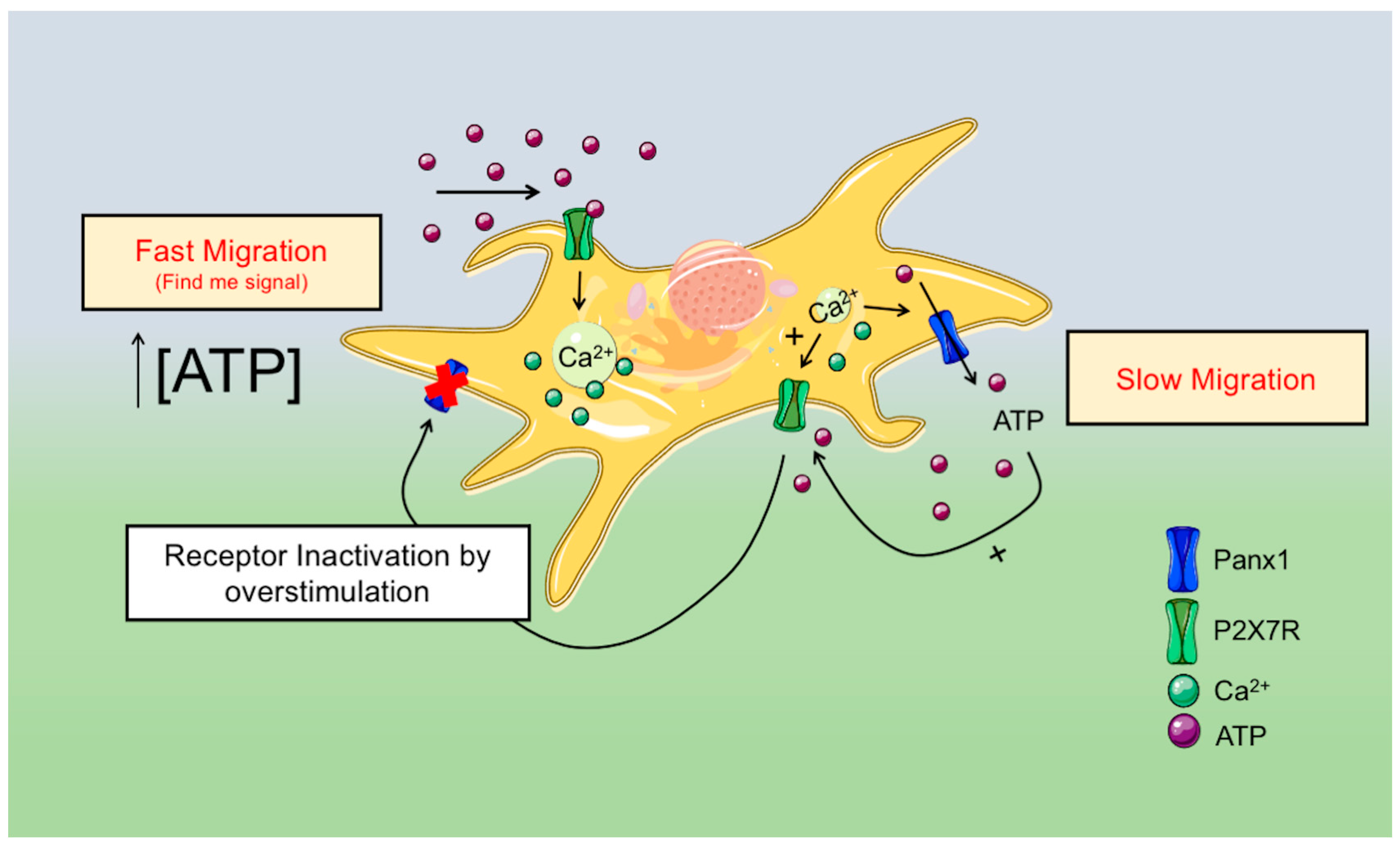
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Human and Mice Fibroblast Cell Cultures
4.3. Determination of the Amount of Pannexins mRNA
4.4. Determination of Relative Amount of Proteins
4.5. In Vitro Scratch Wound Assay
4.6. Knockdown of Panx1 and Panx3 Expression by siRNA
4.7. Measurement of ATP Concentrations
4.8. Dye Uptake Assays
4.9. Single-Fibroblasts Motility Assay
4.10. Cell Surface Imaging of Actin Fibers
4.11. Data Analysis and Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Panx | Pannexin |
| Conxs | Connexins |
| Panx1 | Pannexin 1 |
| Panx3 | Pannexin 3 |
| P2X7R | Purinergic receptor P2X7 |
| P2Y2R | P2Y2 receptor (P2Y2R) |
| MDF | Mouse dermal fibroblast |
| HDF | Human dermal fibroblast |
| MDFpanx1−/− | Pannexin1 knock-out mouse dermal fibroblast |
| siRNA | Small interfering RNA |
| PBN | Probenecid |
| WT | Wild type |
| Panx1KO | Pannexin 1 knock-out |
| F-actin | Filamentous actin |
| Arp3 | Actin-related protein 3 |
References
- Bao, L.; Locovei, S.; Dahl, G. Pannexin membrane channels are mechanosensitive conduits for ATP. FEBS Lett. 2004, 572, 65–68. [Google Scholar] [CrossRef] [Green Version]
- Chekeni, F.B. Pannexin 1 channels mediate ‘find-me’ signal release and membrane permeability during apoptosis. Nature 2010, 467, 863–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penuela, S.; Gehi, R.; Laird, D.W. The biochemistry and function of pannexin channels. Biochim. Biophys. Acta Biomembr. 2013, 1828, 15–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boassa, D.; Ambrosi, C.; Qiu, F.; Dahl, G.; Gaietta, G.; Sosinsky, G. Pannexin1 channels contain a glycosylation site that targets the hexamer to the plasma membrane. J. Biol. Chem. 2007, 282, 31733–31743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surprenant, A.; Rassendren, F.; Kawashima, E.; North, R.A.; Buell, G. The cytolytic P2Z receptor for extracellular ATP identified as a P2X receptor (P2X7). Science 1996, 272, 735–738. [Google Scholar] [CrossRef]
- Locovei, S.; Wang, J.; Dahl, G. Activation of pannexin 1 channels by ATP through P2Y receptors and by cytoplasmic calcium. FEBS Lett. 2006, 580, 239–244. [Google Scholar] [CrossRef] [Green Version]
- Locovei, S.; Scemes, E.; Qiu, F.; Spray, D.C.; Dahl, G. Pannexin1 is part of the pore forming unit of the P2X7 receptor death complex. FEBS Lett. 2007, 581, 483–488. [Google Scholar] [CrossRef] [Green Version]
- Iglesias, R. P2X7 receptor-Pannexin1 complex: Pharmacology and signaling. Am. J. Physiol.-Cell Physiol. 2008, 295, C752–C760. [Google Scholar] [CrossRef] [Green Version]
- Burnstock, G.; Knight, G.E.; Greig, A.V.H. Purinergic Signaling in Healthy and Diseased Skin. J. Investig. Dermatol. 2011, 132, 526–546. [Google Scholar] [CrossRef] [Green Version]
- Singer, A.J.; Clark, R.A. Cutaneous Wound Healing. N. Engl. J. Med. 1999, 341, 738–746. [Google Scholar] [CrossRef]
- Gurtner, G.C.; Werner, S.; Barrandon, Y.; Longaker, M.T. Wound repair and regeneration. Nature 2008, 453, 314–321. [Google Scholar] [CrossRef]
- Nobes, C.D.; Hall, A. Rho GTPases control polarity, protrusion, and adhesion during cell movement. J. Cell Biol. 1999, 144, 1235–1244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kretz, M. Altered connexin expression and wound healing in the epidermis of connexin-deficient mice. J. Cell Sci. 2003, 116, 3443–3452. [Google Scholar] [CrossRef] [Green Version]
- Brandner, J.M.; Houdek, P.; Hüsing, B.; Kaiser, C.; Moll, I. Connexins 26, 30, and 43: Differences among spontaneous, chronic, and accelerated human wound healing. J. Investig. Dermatol. 2004, 122, 1310–1320. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.M.; Lincoln, J.; Cook, J.E.; Becker, D.L. Abnormal connexin expression underlies delayed wound healing in diabetic skin. Diabetes 2007, 56, 2809–2817. [Google Scholar] [CrossRef] [Green Version]
- Wright, C.S.; Van Steensel, M.A.; Hodgins, M.B.; Martin, P.E. Connexin mimetic peptides improve cell migration rates of human epidermal keratinocytes and dermal fibroblasts in vitro. Wound Repair Regen. 2009, 17, 240–249. [Google Scholar] [CrossRef]
- Churko, J.M. Human dermal fibroblasts derived from oculodentodigital dysplasia patients suggest that patients may have wound-healing defects. Hum. Mutat. 2011, 32, 456–466. [Google Scholar] [CrossRef] [Green Version]
- Wright, C.S.; Pollok, S.; Flint, D.J.; Brandner, J.M.; Martin, P.E.M. The connexin mimetic peptide Gap27 increases human dermal fibroblast migration in hyperglycemic and hyperinsulinemic conditions in vitro. J. Cell. Physiol. 2012, 227, 77–87. [Google Scholar] [CrossRef]
- Penuela, S. Pannexin 1 and pannexin 3 are glycoproteins that exhibit many distinct characteristics from the connexin family of gap junction proteins. J. Cell Sci. 2007, 120, 3772–3783. [Google Scholar] [CrossRef] [Green Version]
- Celetti, S.J.; Cowan, K.N.; Penuela, S.; Shao, Q.; Churko, J.; Laird, D.W. Implications of pannexin 1 and pannexin 3 for keratinocyte differentiation. J. Cell Sci. 2010, 123, 1363–1372. [Google Scholar] [CrossRef] [Green Version]
- Penuela, S.; Kelly, J.J.; Churko, J.M.; Barr, K.J.; Berger, A.C.; Laird, D.W. Panx1 regulates cellular properties of keratinocytes and dermal fibroblasts in skin development and wound healing. J. Investig. Dermatol. 2014, 134, 2026–2035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P. Pannexin-3 Deficiency Delays Skin Wound Healing in Mice due to Defects in Channel Functionality. J. Investig. Dermatol. 2019, 139, 909–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinheiro, A.R. Histamine induces ATP release from human subcutaneous fibroblasts, via pannexin-1 hemichannels, leading to Ca2+ mobilization and cell proliferation. J. Biol. Chem. 2013, 288, 27571–27583. [Google Scholar] [CrossRef] [Green Version]
- Cowan, K.N.; Langlois, S.; Penuela, S.; Cowan, B.J.; Laird, D.W. Pannexin1 and Pannexin3 Exhibit Distinct Localization Patterns in Human Skin Appendages and are Regulated during Keratinocyte Differentiation and Carcinogenesis. Cell Commun. Adhes. 2012, 19, 45–53. [Google Scholar] [CrossRef] [Green Version]
- Baranova, A. The mammalian pannexin family is homologous to the invertebrate innexin gap junction proteins. Genomics 2004, 83, 706–716. [Google Scholar] [CrossRef]
- Bruzzone, R.; Hormuzdi, S.G.; Barbe, M.T.; Herb, A.; Monyer, H. Pannexins, a family of gap junction proteins expressed in brain. Proc. Natl. Acad. Sci. USA 2003, 100, 13644–13649. [Google Scholar] [CrossRef] [Green Version]
- Saéz, P.J.; Vargas, P.; Shoji, K.F.; Harcha, P.A.; Lennon-Duméni, A.M.; Saéz, J.C. ATP promotes the fast migration of dendritic cells through the activity of pannexin 1 channels and P2X7 receptors. Sci. Signal. 2017, 10, 7107. [Google Scholar] [CrossRef] [Green Version]
- Wicki-Stordeur, L.E.; Swayne, L.A. Panx1 regulates neural stem and progenitor cell behaviours associated with cytoskeletal dynamics and interacts with multiple cytoskeletal elements. Cell Commun. Signal. 2013, 11, 62. [Google Scholar] [CrossRef]
- Silverman, W.; Locovei, S.; Dahl, G. Probenecid, a gout remedy, inhibits pannexin 1 channels. AJP Cell Physiol. 2008, 295, C761–C767. [Google Scholar] [CrossRef] [Green Version]
- Pelegrin, P.; Surprenant, A. Pannexin-1 mediates large pore formation and interleukin-1beta release by the ATP-gated P2X7 receptor. Eur. Mol. Biol. Organ. J. 2006, 25, 5071–5082. [Google Scholar] [CrossRef] [Green Version]
- García, I.E. Keratitis-ichthyosis-deafness syndrome-associated Cx26 mutants produce nonfunctional gap junctions but hyperactive hemichannels when co-expressed with wild type Cx43. J. Investig. Dermatol. 2015, 135, 1338–1347. [Google Scholar] [CrossRef] [Green Version]
- Solini, A.; Chiozzi, P.; Morelli, A.; Fellin, R.; Di Virgilio, F. Human primary fibroblasts in vitro express a purinergic P2X7 receptor coupled to ion fluxes, microvesicle formation and IL-6 release. J. Cell Sci. 1999, 112, 297–305. [Google Scholar]
- Gentile, D. Searching novel therapeutic targets for scleroderma: P2X7-receptor is UP-regulated and promotes a fibrogenic phenotype in systemic sclerosis fibroblasts. Front. Pharmacol. 2017, 8, 638. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, K.A. Agonists and Antagonists for P2 Receptors. In Foundation Symposium; John Wiley: Chichester, NY, USA, 1999; p. 58. [Google Scholar]
- Jin, H. P2Y2R activation by nucleotides promotes skin wound-healing process. Exp. Dermatol. 2014, 23, 480–485. [Google Scholar] [CrossRef]
- Klepeis, V.E.; Weinger, I.; Kaczmarek, E.; Trinkaus-Randall, V. P2Y receptors play a critical role in epithelial cell communication and migration. J. Cell. Biochem. 2004, 93, 1115–1133. [Google Scholar] [CrossRef]
- Riedl, J. Lifeact: A versatile marker to visualize F-actin. Nat. Methods 2008, 5, 605–607. [Google Scholar] [CrossRef]
- Hata, S.; Okamura, K.; Hatta, M.; Ishikawa, H.; Yamazaki, J. Proteolytic and non-proteolytic activation of keratinocyte-derived latent TGF-β1 induces fibroblast differentiation in a wound-healing model using rat skin. J. Pharmacol. Sci. 2014, 124, 230–243. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y. ATP release guides neutrophil chemotaxis via P2Y2 and A3 receptors. Science 2006, 314, 1792–1795. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y. Purinergic signaling: A fundamental mechanism in neutrophil activation. Sci. Signal. 2010, 3, ra45. [Google Scholar] [CrossRef] [Green Version]
- Bao, Y.; Chen, Y.; Ledderose, C.; Li, L.; Junger, W.G. Pannexin 1 channels link chemoattractant receptor signaling to local excitation and global inhibition responses at the front and back of polarized neutrophils. J. Biol. Chem. 2013, 288, 22650–22657. [Google Scholar] [CrossRef] [Green Version]
- Xu, X. Probenecid disrupts a novel pannexin 1-collapsin response mediator protein 2 interaction and increases microtubule stability. Front. Cell. Neurosci. 2018, 12, 124. [Google Scholar] [CrossRef] [Green Version]
- Wei, L.; Yang, X.; Shi, X.; Chen, Y. Pannexin-1 silencing inhibits the proliferation of U87-MG cells. Mol. Med. Rep. 2015, 11, 3487–3492. [Google Scholar] [CrossRef]
- Orellana, J.A. ATP and glutamate released via astroglial connexin 43 hemichannels mediate neuronal death through activation of pannexin 1 hemichannels. J. Neurochem. 2011, 118, 826–840. [Google Scholar] [CrossRef] [Green Version]
- Khammanit, R.; Chantakru, S.; Kitiyanant, Y.; Saikhun, J. Effect of serum starvation and chemical inhibitors on cell cycle synchronization of canine dermal fibroblasts. Theriogenology 2008, 70, 27–34. [Google Scholar] [CrossRef]
- Thoumine, O.; Ott, A. Influence of adhesion and cytoskeletal integrity on fibroblast traction. Cell Motil. Cytoskelet. 1996, 35, 269–280. [Google Scholar] [CrossRef]
- Yin, J.; Xu, K.; Zhang, J.; Kumar, A.; Yu, F.S.X. Wound-induced ATP release and EGF receptor activation in epithelial cell. J. Cell Sci. 2007, 120, 5818–5825. [Google Scholar] [CrossRef] [Green Version]
- Corriden, R.; Insel, P.A. Basal release of ATP: An autocrine-paracrine mechanism for cell regulation. Sci. Signal. 2010, 3, re1. [Google Scholar] [CrossRef] [Green Version]
- Romanov, R.A.; Bystrova, M.F.; Rogachevskaya, O.A.; Sadovnikov, V.B.; Shestopalov, V.I.; Kolesnikov, S.S. The ATP permeability of pannexin 1 channels in a heterologous system and in mammalian taste cells is dispensable. J. Cell Sci. 2012, 125, 5514–5523. [Google Scholar] [CrossRef] [Green Version]
- Pellegatti, P.; Falzoni, S.; Pinton, P.; Rizzuto, R.; Di Virgilio, F. A novel recombinant plasma membrane-targeted luciferase reveals a new pathway for ATP secretion. Mol. Biol. Cell 2005, 16, 3659–3665. [Google Scholar] [CrossRef] [Green Version]
- Brandao-Burch, A.; Key, M.L.; Patel, J.J.; Arnett, T.R.; Orriss, I.R. The P2X7 receptor is an important regulator of extracellular ATP levels. Front. Endocrinol. 2012, 3, 41. [Google Scholar] [CrossRef] [Green Version]
- Qiu, F.; Dahl, G. A permeant regulating its permeation pore: Inhibition of pannexin 1 channels by ATP. Am. J. Physiol. Cell Physiol. 2009, 296, C250–C255. [Google Scholar] [CrossRef] [PubMed]
- Di Virgilio, F.; Schmalzing, G.; Markwardt, F. The Elusive P2X7 Macropore. Trends Cell Biol. 2018, 28, 392–404. [Google Scholar] [CrossRef] [PubMed]
- Karasawa, A.; Michalski, K.; Mikhelzon, P.; Kawate, T. The P2X7 receptor forms a dye-permeable pore independent of its intracellular domain but dependent on membrane lipid composition. eLife 2017, 6, e31186. [Google Scholar] [CrossRef] [PubMed]
- Boyce, A.K.J.; Wicki-Stordeur, L.E.; Swayne, L.A. Powerful partnership: Crosstalk between pannexin 1 and the cytoskeleton. Front. Physiol. 2014, 5, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyce, A.K.J.; Swayne, L.A. P2X7 receptor cross-Talk regulates ATP-induced pannexin 1 internalization. Biochem. J. 2017, 474, 2133–2144. [Google Scholar] [CrossRef]
- Boucher, I.; Rich, C.; Lee, A.; Marcincin, M.; Trinkaus-Randall, V. The P2Y2 receptor mediates the epithelial injury response and cell migration. Am. J. Physiol. Cell Physiol. 2010, 299, C411–C421. [Google Scholar] [CrossRef] [Green Version]
- Hashikawa, T. Regulation of adenosine receptor engagement by ecto-adenosine deaminase. FASEB J. 2004, 18, 131–133. [Google Scholar] [CrossRef]
- Sohail, M.A. Adenosine induces loss of actin stress fibers and inhibits contraction in hepatic stellate cells via Rho inhibition. Hepatology 2009, 49, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Mogilner, A. Comparison of cell migration mechanical strategies in three-dimensional matrices: A computational study. Interface Focus 2016, 6, 20060640. [Google Scholar] [CrossRef] [Green Version]
- Svitkina, T. The actin cytoskeleton and actin-based motility. Cold Spring Harb. Perspect. Biol. 2018, 10, a018267. [Google Scholar] [CrossRef] [Green Version]
- Bhalla-Gehi, R.; Penuela, S.; Churko, J.M.; Shao, Q.; Laird, D.W. Pannexin1 and pannexin3 delivery, cell surface dynamics, and cytoskeletal interactions. J. Biol. Chem. 2010, 285, 9147–9160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meijerman, I.; Blom, W.M.; Nagelkerke, J.F. The effect of extracellular ATP on the actin cytoskeleton of hepatocytes. Biochem. Soc. Trans. 1996, 24, 560S. [Google Scholar] [CrossRef]
- Meijerman, I.; Blom, W.M.; De Bont, H.J.G.M.; Mulder, G.J.; Nagelkerke, J.F. Nuclear accumulation of G-actin in isolated rat hepatocytes by adenine nucleotides. Biochem. Biophys. Res. Commun. 1997, 240, 697–700. [Google Scholar] [CrossRef]
- Joseph, J.; Grierson, I.; Hitchings, R.A. Exogenous ATP causes the contraction of intact fibroblasts in vitro. Exp. Cell Res. 1988, 176, 1–12. [Google Scholar] [CrossRef]
- Ehrlich, H.P.; Rajaratnam, J.B.; Griswold, T.R. ATP-induced cell contraction in dermal fibroblasts: Effects of cAMP and myosin light-chain kinase. J. Cell. Physiol. 1986, 128, 223–230. [Google Scholar] [CrossRef]
- Sun, S.X.; Walcott, S.; Wolgemuth, C.W. Cytoskeletal cross-linking and bundling in motor-independent contraction. Curr. Biol. 2010, 20, R649–R654. [Google Scholar] [CrossRef] [Green Version]
- Jahraus, A. ATP-dependent membrane assembly of F-actin facilitates membrane fusion. Mol. Biol. Cell 2001, 12, 155–170. [Google Scholar] [CrossRef] [Green Version]
- Kellerman, P.S.; Norenberg, S.; Guse, N. Exogenous adenosine triphosphate (ATP) preserves proximal tubule microfilament structure and function in vivo in a maleic acid model of ATP depletion. J. Clin. Investig. 1993, 92, 1940–1949. [Google Scholar] [CrossRef] [Green Version]
- Bao, B.A.; Lais, C.P.; Nauss, C.C.; Morgan, J.R. Pannexin1 drives multicellular aggregate compaction via a signaling cascade that remodels the actin cytoskeleton. J. Biol. Chem. 2012, 287, 8407–8416. [Google Scholar] [CrossRef] [Green Version]
- Anselmi, F. ATP release through connexin hemichannels and gap junction transfer of second messengers propagate Ca2+ signals across the inner ear. Proc. Natl. Acad. Sci. USA 2008, 105, 18770–18775. [Google Scholar] [CrossRef] [Green Version]
- Weinstein-Oppenheimer, C.R. Design of a hybrid biomaterial for tissue engineering: Biopolymer-scaffold integrated with an autologous hydrogel carrying mesenchymal stem-cells. Mater. Sci. Eng. C 2017, 79, 821–830. [Google Scholar] [CrossRef]
- Rittié, L.; Fisher, G.J. Isolation and culture of skin fibroblasts. Methods Mol. Med. 2005, 117, 83–98. [Google Scholar]
- Liang, C.C.; Park, A.Y.; Guan, J.L. In vitro scratch assay: A convenient and inexpensive method for analysis of cell migration in vitro. Nat. Protoc. 2007, 2, 329. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flores-Muñoz, C.; Maripillán, J.; Vásquez-Navarrete, J.; Novoa-Molina, J.; Ceriani, R.; Sánchez, H.A.; Abbott, A.C.; Weinstein-Oppenheimer, C.; Brown, D.I.; Cárdenas, A.M.; et al. Restraint of Human Skin Fibroblast Motility, Migration, and Cell Surface Actin Dynamics, by Pannexin 1 and P2X7 Receptor Signaling. Int. J. Mol. Sci. 2021, 22, 1069. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031069
Flores-Muñoz C, Maripillán J, Vásquez-Navarrete J, Novoa-Molina J, Ceriani R, Sánchez HA, Abbott AC, Weinstein-Oppenheimer C, Brown DI, Cárdenas AM, et al. Restraint of Human Skin Fibroblast Motility, Migration, and Cell Surface Actin Dynamics, by Pannexin 1 and P2X7 Receptor Signaling. International Journal of Molecular Sciences. 2021; 22(3):1069. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031069
Chicago/Turabian StyleFlores-Muñoz, Carolina, Jaime Maripillán, Jacqueline Vásquez-Navarrete, Joel Novoa-Molina, Ricardo Ceriani, Helmuth A. Sánchez, Ana C. Abbott, Caroline Weinstein-Oppenheimer, Donald I. Brown, Ana María Cárdenas, and et al. 2021. "Restraint of Human Skin Fibroblast Motility, Migration, and Cell Surface Actin Dynamics, by Pannexin 1 and P2X7 Receptor Signaling" International Journal of Molecular Sciences 22, no. 3: 1069. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031069