
Regulation of the CoA Biosynthetic Complex Assembly in Mammalian Cells

 , , , ,
, , , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
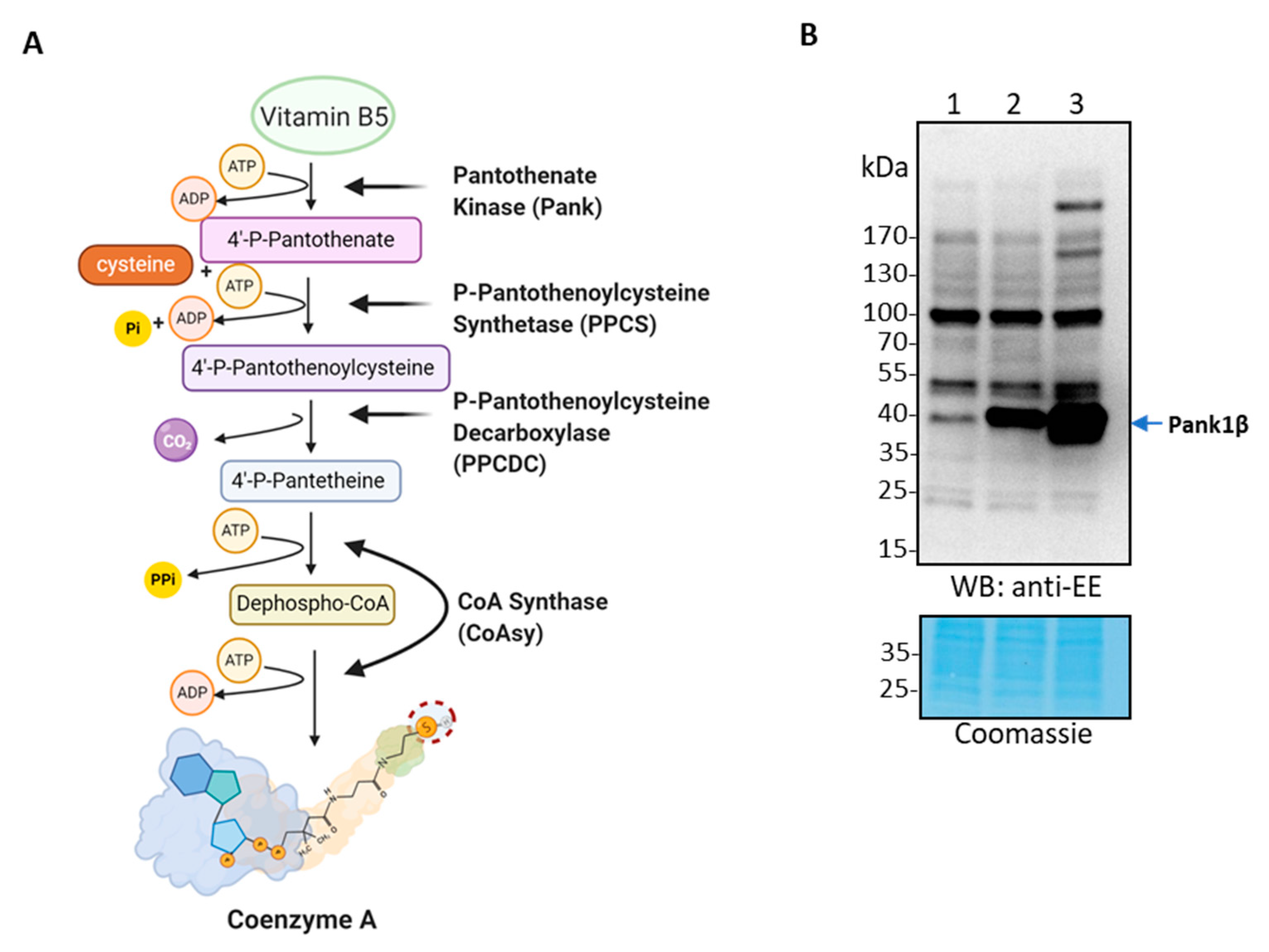
2.1. Analysis of the CoA Biosynthetic Complex in HEK293/Pank1β Cells Using Conventional Approaches
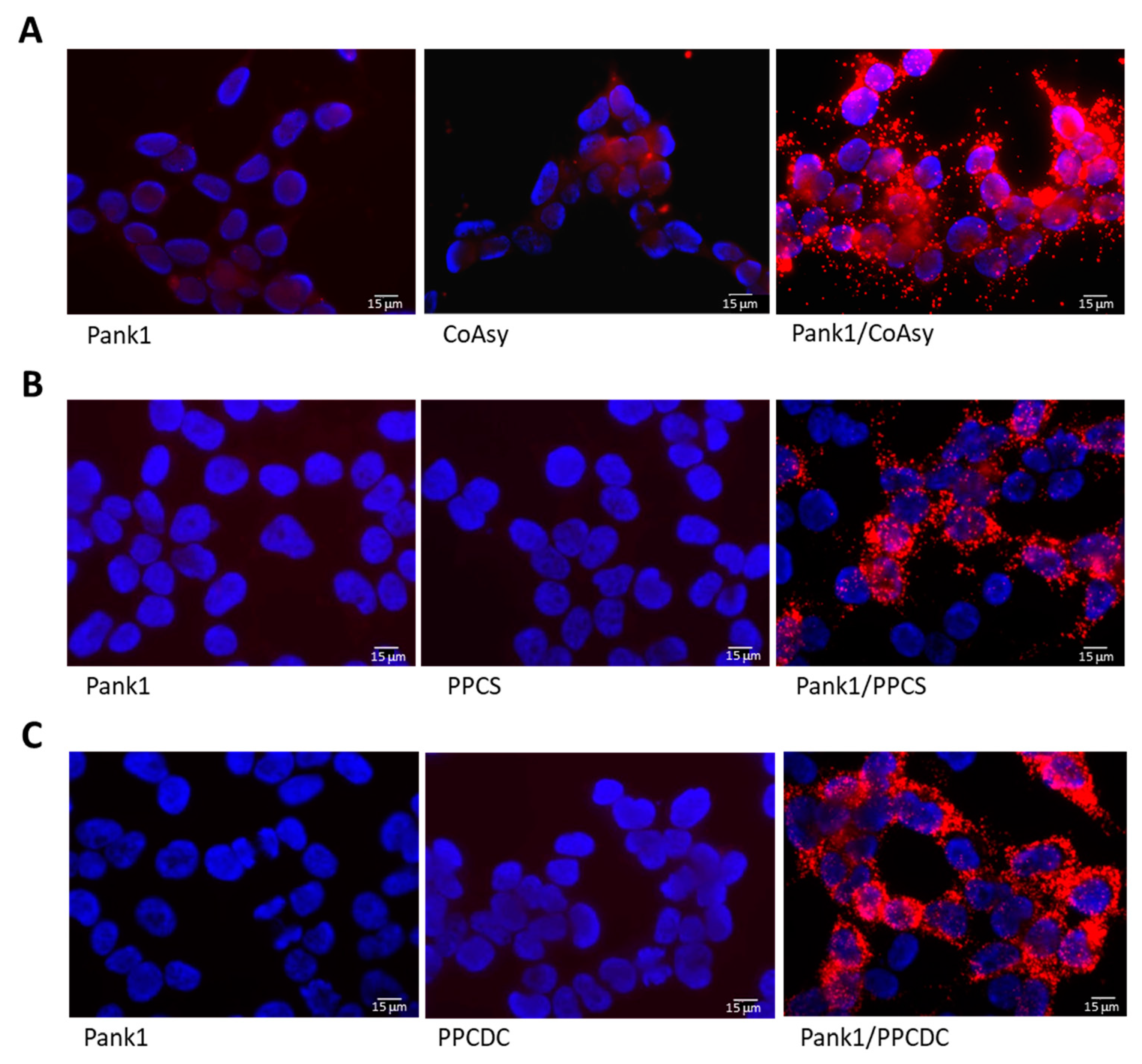
2.2. Validation of the Proximity Ligation Assay for Testing the Interaction between Enzymes of CoA Biosynthesis
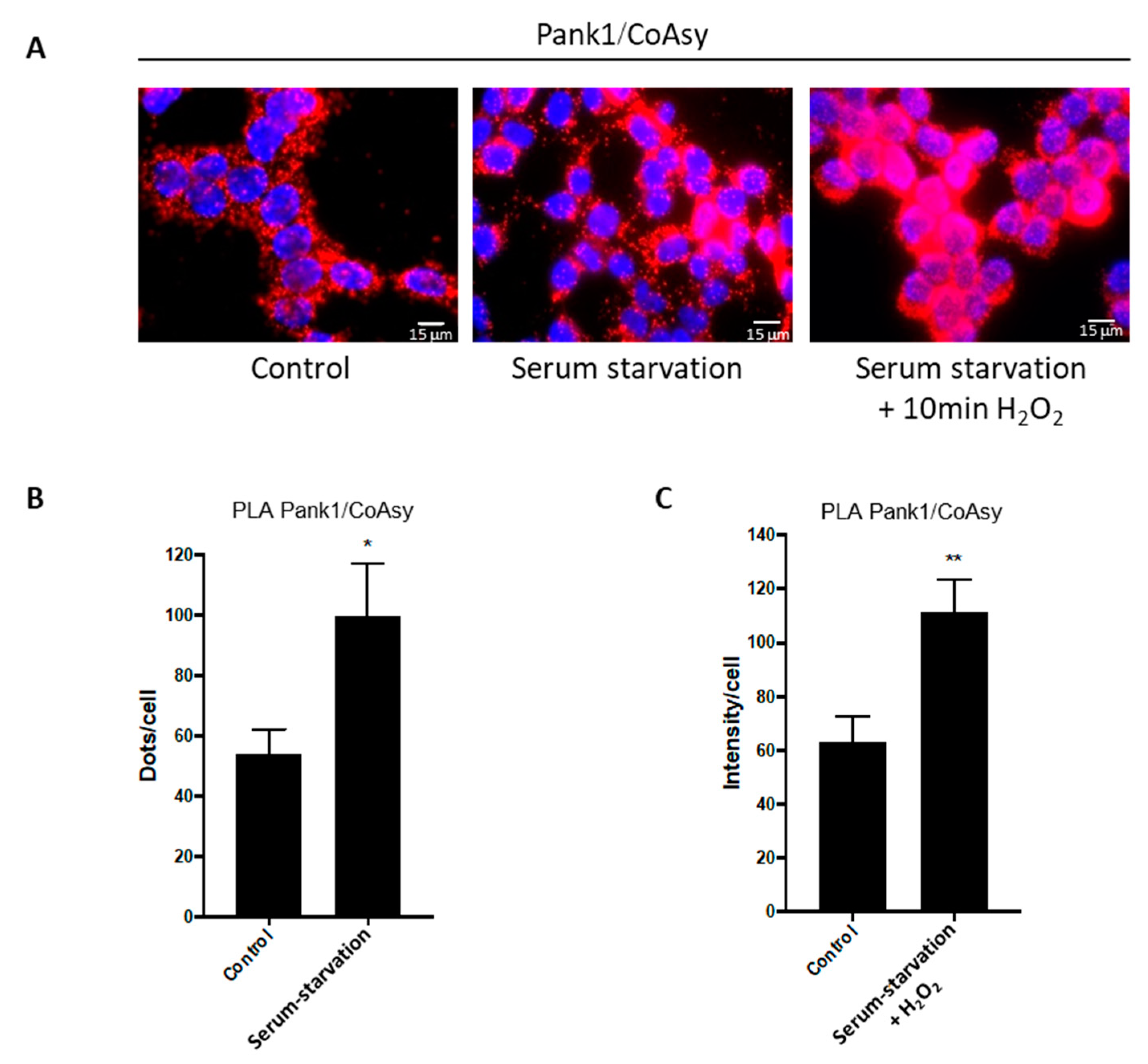
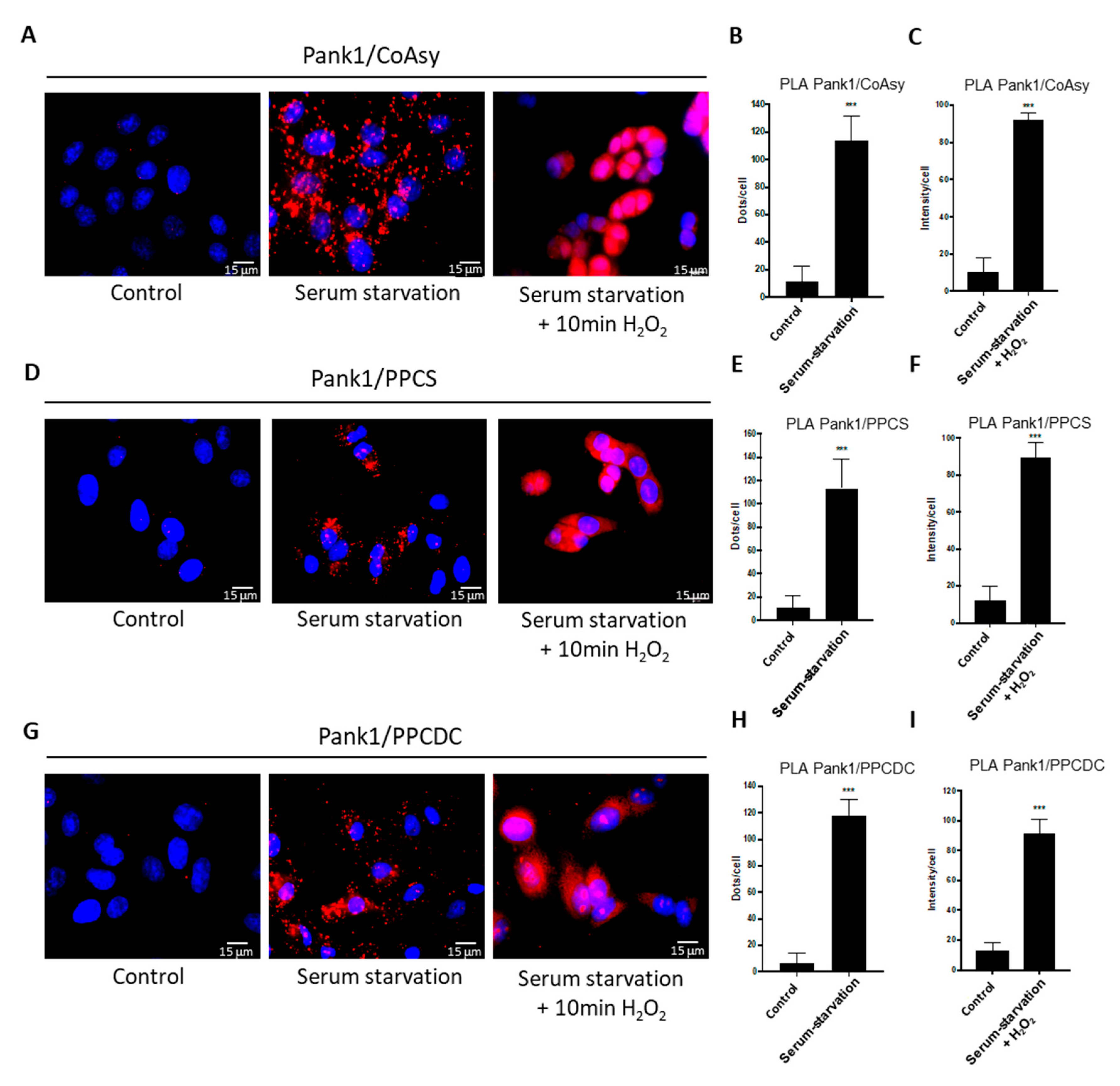
2.3. Oxidative Stress and Serum Starvation Induce the Association of CoA Biosynthetic Enzymes
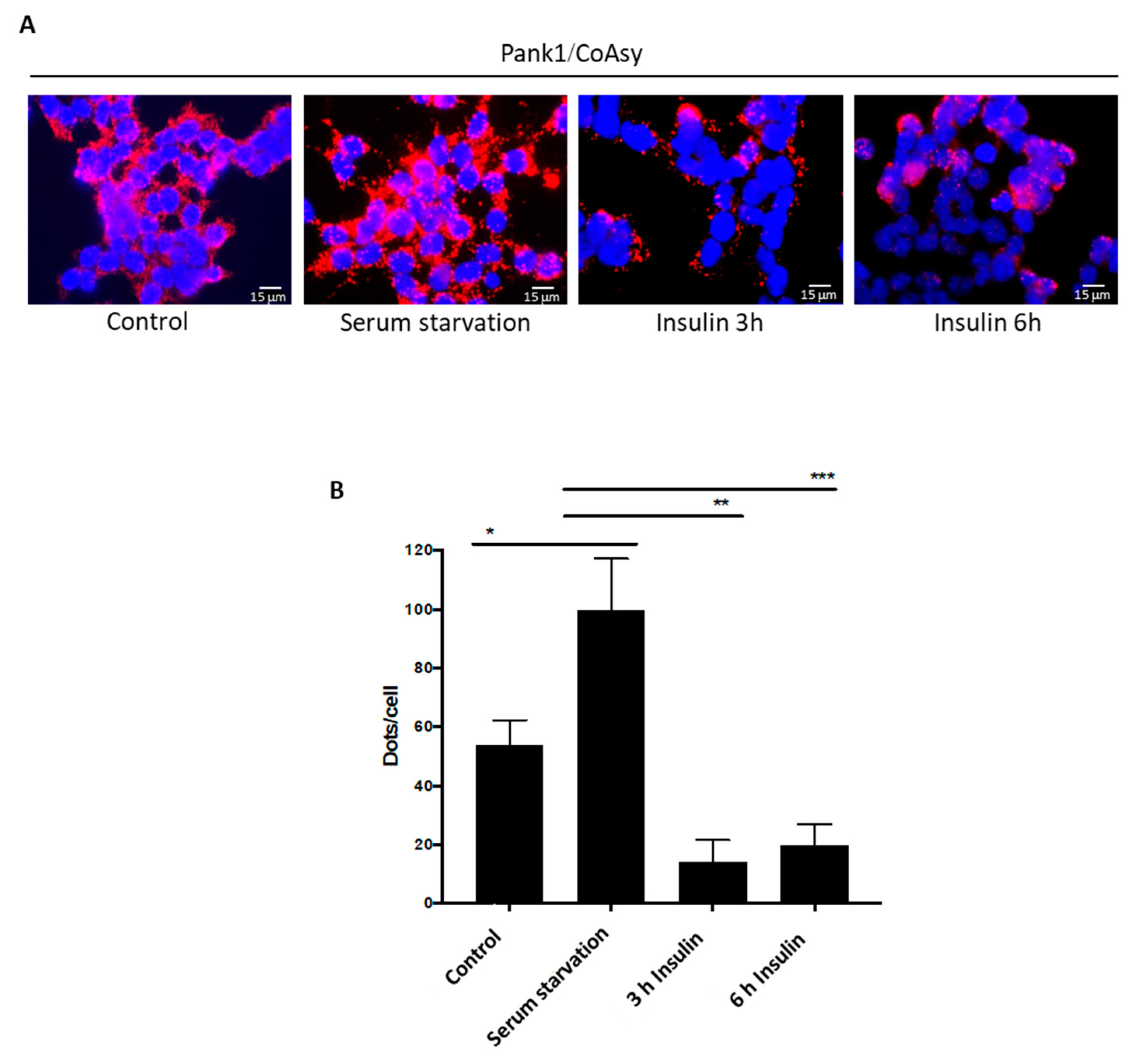
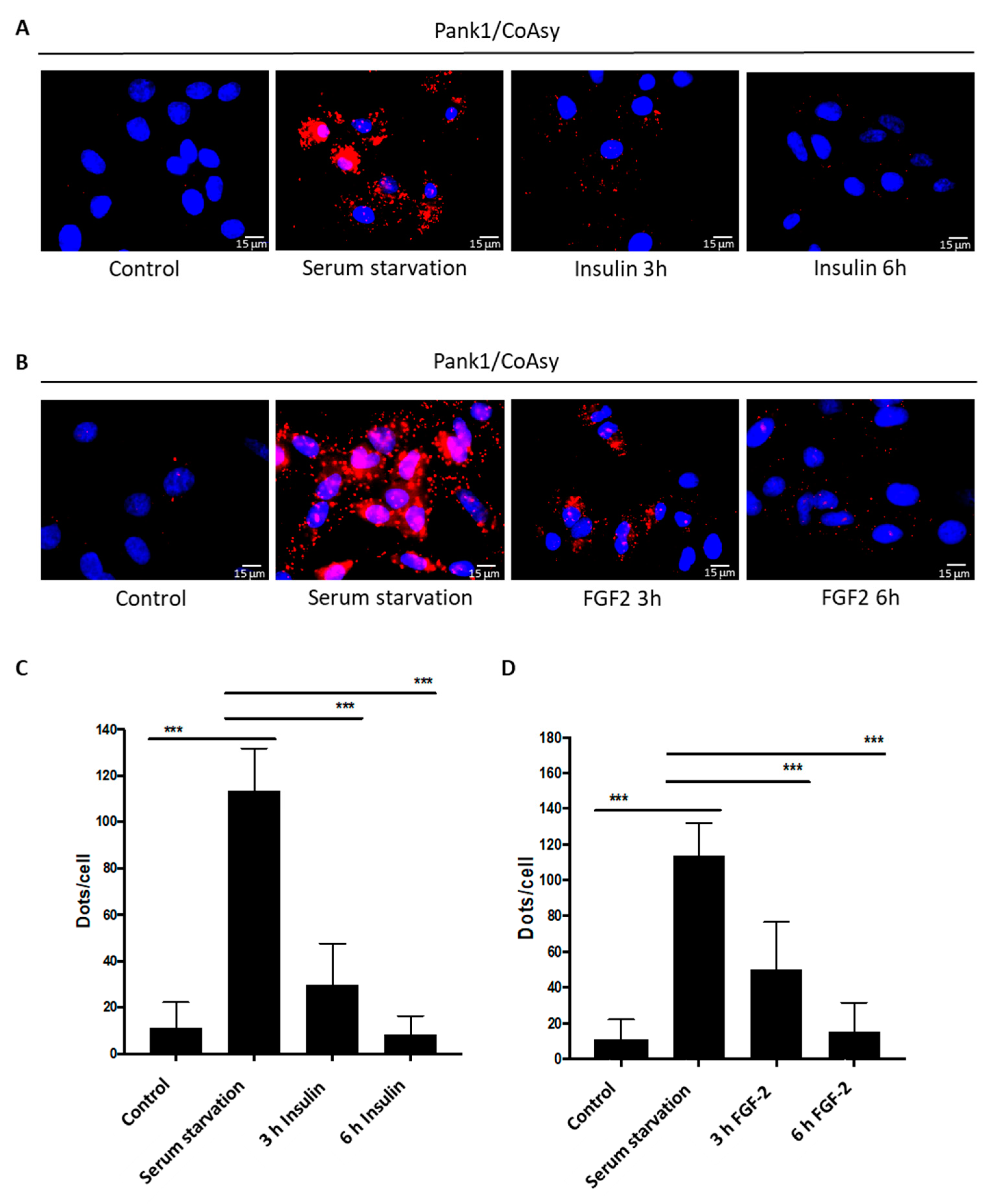
2.4. Treatment of Serum-Starved Cells with Insulin Promotes the Dissociation of CoA Biosynthetic Enzymes
2.5. Assembly of Endogenous CoA Biosynthetic Enzymes is Induced in A549 Cells in Response to Oxidative Stress
2.6. Growth Signaling Inhibits the Assembly of Endogenous CoA Biosynthetic Enzymes in A549 Cells
3. Discussion
4. Materials and Methods
4.1. Reagents and Chemicals
Antibodies
4.2. Cell Culture and Generation of the HEK293/Pank1β Cell Line
4.3. Treatment of HEK293/Pank1β and A549 Cells
4.4. SDS-PAGE and Western Blot Analysis
4.5. Proximity Ligation Assay
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| CoA | Coenzyme A |
| PanK | Pantothenate kinase |
| PPCS | 4′-phosphopantothenoylcysteine synthetase |
| PPCDC | 4′-phosphopantothenoylcysteine decarboxylase |
| CoAsy | CoA synthase |
| PLA | Proximity ligation assay |
| FGF | Fibroblast growth factor |
| NBIA | Neurodegeneration with brain iron accumulation |
References
- Leonardi, R.; Zhang, Y.M.; Rock, C.O.; Jackowski, S. Coenzyme A: Back in action. Prog. Lipid Res. 2005, 44, 125–153. [Google Scholar] [CrossRef] [PubMed]
- Martinez, D.L.; Tsuchiya, Y.; Gout, I. Coenzyme A biosynthetic machinery in mammalian cells. Biochem. Soc. Trans. 2014, 42, 1112–1117. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, Y.; Peak-Chew, S.Y.; Newell, C.; Miller-Aidoo, S.; Mangal, S.; Zhyvoloup, A.; Baković, J.; Malanchuk, O.; Pereira, G.C.; Kotiadis, V.; et al. Protein CoAlation: A redox-regulated protein modification by coenzyme A in mammalian cells. Biochem. J. 2017, 2, 2489–2508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuchiya, Y.; Zhyvoloup, A.; Baković, J.; Thomas, N.; Yi Kun Yu, B.; Das, S.; Orengo, C.; Newell, C.; Ward, J.; Saladino, G.; et al. Protein CoAlation and antioxidant function of coenzyme A in prokaryotic cells. Biochem. J. 2018, 475, 1909–1937. [Google Scholar] [CrossRef] [Green Version]
- Gout, I. Coenzyme A, protein CoAlation and redox regulation in mammalian cells. Biochem. Soc. Trans. 2018, 46, 721–728. [Google Scholar] [CrossRef] [Green Version]
- Gout, I. Coenzyme A: A protective thiol in bacterial antioxidant defence. Biochem. Soc. Trans. 2019, 47, 469–476. [Google Scholar] [CrossRef]
- Bakovic, J.; Yu, B.Y.K.; Silva, D.; Chew, S.P.; Kim, S.; Ahn, S.-H.; Palmer, L.; Aloum, L.; Stanzani, G.; Malanchuk, O.; et al. A key metabolic integrator, coenzyme A, modulates the activity of peroxiredoxin 5 via covalent modification. Mol. Cell. Biochem. 2019, 461, 91–102. [Google Scholar] [CrossRef] [Green Version]
- Aloum, L.; Brimson, C.A.; Zhyvoloup, A.; Baines, R.; Baković, J.; Filonenko, V.; Thompson, C.R.L.; Gout, I. Coenzyme A and protein CoAlation levels are regulated in response to oxidative stress and during morphogenesis in Dictyostelium discodeum. Biochem. Biophys. Res. Commun. 2019, 511, 294–299. [Google Scholar] [CrossRef]
- Tsuchiya, Y.; Byrne, D.P.; Burgess, S.G.; Bormann, J.; Bakovic, J.; Huang, Y.; Zhyvoloup, A.; Yu, B.Y.K.; Peak-Chew, S.; Tran, T.; et al. Covalent Aurora A regulation by the metabolic integrator coenzyme A. Redox Biol. 2019, 28, 1013–1018. [Google Scholar] [CrossRef]
- Tsuji, K.; Yoon, K.-S.; Ogo, S. Glyceraldehyde-3-phosphate dehydrogenase from Citrobacter sp. S-77 is post-translationally modified by CoA (protein CoAlation) under oxidative stress. FEBS Open Bio. 2019, 9, 53–73. [Google Scholar] [CrossRef] [Green Version]
- Leonardi, R.; Jackowski, S. Biosynthesis of pantothenic acid and coenzyme A. EcoSal Plus 2007, 2, 1–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strauss, E.; Kinsland, C.; Ge, Y.; McLafferty, F.W.; Begley, T.P. Phosphopantothenoylcysteine synthetase from Escherichia coli. Identification and characterization of the last unidentified coenzyme A biosynthetic enzyme in bacteria. J. Biol. Chem. 2001, 276, 13513–13516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rana, A.; Seinen, E.; Siudeja, K.; Muntendam, R.; Srinivasan, B.; van der Want, J.J.; Hayflick, S.; Reijngoud, D.-J.; Kayser, O.; Sibon, O.C.M. Pantethine rescues a Drosophila model for pantothenate kinase-associated neurodegeneration. Proc. Natl. Acad. Sci. USA 2010, 107, 6988–6993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calder, R.B.; Williams, R.S.B.; Ramaswamy, G.; Rock, C.O.; Campbell, E.; Unkles, S.E.; Kinghorn, J.R.; Jackowski, S. Cloning and characterization of a eukaryotic pantothenate kinase gene (panK) from Aspergillus nidulans. J. Biol. Chem. 1999, 274, 2014–2020. [Google Scholar] [CrossRef] [Green Version]
- Breus, O.; Panasyuk, G.; Gout, I.T.; Filonenko, V.; Nemazanyy, I. CoA Synthase is phosphorylated on tyrosines in mammalian cells, interacts with and is dephosphorylated by Shp2PTP. Mol. Cell. Biochem. 2010, 335, 195–202. [Google Scholar] [CrossRef]
- Gudkova, D.; Panasyuk, G.; Nemazanyy, I.; Zhyvoloup, A.; Monteil, P.; Filonenko, V.; Gout, I. EDC4 interacts with and regulates the dephospho-CoA kinase activity of CoA synthase. FEBS Lett. 2012, 586, 3590–3595. [Google Scholar] [CrossRef] [Green Version]
- Nemazanyy, I.; Panasyuk, G.; Zhyvoloup, A.; Panayotou, G.; Gout, I.T.; Filonenko, V. Specific interaction between S6K1 and CoA synthase: A potential link between the mTOR/S6K pathway, CoA biosynthesis and energy metabolism. FEBS Lett. 2004, 578, 357–362. [Google Scholar] [CrossRef] [Green Version]
- Zhyvoloup, A.; Nemazanyy, I.; Panasyuk, G.; Valovka, T.; Fenton, T.; Rebholz, H.; Wang, M.-L.L.; Foxon, R.; Lyzogubov, V.; Usenko, V.; et al. Subcellular localization and regulation of coenzyme A synthase. J. Biol. Chem. 2003, 278, 50316–50321. [Google Scholar] [CrossRef] [Green Version]
- Robishaw, J.D.; Berkich, D.; Neely, J.R. Rate-limiting step and control of coenzyme A synthesis in cardiac-muscle. J. Biol. Chem. 1982, 257, 967–972. [Google Scholar] [CrossRef]
- Tubbs, P.K.; Garland, P.B. Variations in tissue contents of coenzyme A thioesters and possible metabolic implications. Biochem. J. 1964, 93, 550–557. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.M.; Savage, C.R. Regulation of coenzyme A biosynthesis by glucagon and glucocorticoid in adult rat liver parenchymal cells. Biochem. J. 1980, 188, 175–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theodoulou, F.L.; Sibon, O.C.M.; Jackowski, S.; Gout, I. Coenzyme A and its derivatives: Renaissance of a textbook classic. Biochem. Soc. Trans. 2014, 42, 1025–1032. [Google Scholar] [CrossRef] [PubMed]
- Reibel, D.K.; Wyse, B.W.; Berkich, D.A.; Neely, J.R. Regulation of coenzyme A synthesis effects of diabetes and fasting in heart muscle: Effects of diabetes and fasting. Am. J. Physiol. 1981, 240, H606–H611. [Google Scholar] [CrossRef] [PubMed]
- Mcallister, R.A.; Fixter, L.M.; Campbell, E.H.G. The effect of tumour growth on liver pantothenate, CoA, and fatty acid synthetase activity in the mouse. Br. J. Cancer 1988, 57, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Westaway, S.K.; Levinson, B.; Johnson, M.; Gitschier, J.; Hayflick, S.J. A novel pantothenate kinase gene (PANK2) is defective in Hallervorden-Spatz syndrome. Nat. Genet. 2001, 28, 345–349. [Google Scholar] [CrossRef] [PubMed]
- Dusi, S.; Valletta, L.; Haack, T.B.; Tsuchiya, Y.; Venco, P.; Pasqualato, S.; Goffrini, P.; Tigano, M.; Demchenko, N.; Wieland, T.; et al. Exome sequence reveals mutations in CoA synthase as a cause of neurodegeneration with brain iron accumulation. Am. J. Hum. Genet. 2014, 94, 11–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iuso, A.; Wiersma, M.; Schüller, H.-J.; Pode-Shakked, B.; Marek-Yagel, D.; Grigat, M.; Schwarzmayr, T.; Berutti, R.; Alhaddad, B.; Kanon, B.; et al. Mutations in PPCS, encoding phosphopantothenoylcysteine synthetase, cause autosomal-recessive dilated cardiomyopathy. Am. J. Hum. Genet. 2018, 102, 1018–1030. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, A.; Gonzalez, A.; Munoz, I.; Serrano, R.; Abrie, J.A.; Strauss, E.; Arino, J. Moonlighting proteins Hal3 and Vhs3 form a heteromeric PPCDC with Ykl088w in yeast CoA biosynthesis. Nat. Chem. Biol. 2009, 5, 920–928. [Google Scholar] [CrossRef] [Green Version]
- Rock, C.O.; Calder, R.B.; Karim, M.A.; Jackowski, S. Pantothenate kinase regulation of the intracellular concentration of coenzyme A. J. Biol. Chem. 2000, 275, 1377–1383. [Google Scholar] [CrossRef] [Green Version]
- Rhee, H.-W.; Zou, P.; Udeshi, N.D.; Martell, J.D.; Mootha, V.K.; Carr, S.A.; Ting, A.Y. Proteomic Mapping of Mitochondria in Living Cells via Spatially Restricted Enzymatic Tagging. Science 2013, 339, 1328–1331. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.-C.; Kitagawa, M.; Tang, X.; Hou, M.-H.; Wu, J.; Qu, D.C.; Srinivas, V.; Liu, X.; Thompson, J.W.; Mathey-Prevot, B.; et al. CoA synthase regulates mitotic fidelity via CBP-mediated acetylation. Nat. Commun. 2018, 9, 1039. [Google Scholar] [CrossRef] [PubMed]
- Söderberg, O.; Gullberg, M.; Jarvius, M.; Ridderstråle, K.; Leuchowius, K.-J.; Jarvius, J.; Wester, K.; Hydbring, P.; Bahram, F.; Larsson, L.-G.; et al. Direct observation of individual endogenous protein complexes in situ by proximity ligation. Nat. Methods 2006, 3, 995–1000. [Google Scholar] [CrossRef] [PubMed]
- Nakhjavani, M.; Nikounezhad, N.; Ashtarinezhad, A.; Shirazi, F.H. Human lung carcinoma reaction against metabolic serum deficiency stress. Iran. J. Pharm. Res. 2016, 15, 817–823. [Google Scholar] [PubMed]
- White, E.Z.; Pennant, N.M.; Carter, J.R.; Hawsawi, O.; Odero-Marah, V.; Hinton, C. V Serum deprivation initiates adaptation and survival to oxidative stress in prostate cancer cells. Sci. Rep. 2020, 10, 12505. [Google Scholar] [CrossRef] [PubMed]
- Reczek, C.R.; Chandel, N.S. The two faces of reactive oxygen species in cancer. Annu. Rev. Cancer Biol. 2017, 1, 79–98. [Google Scholar] [CrossRef]
- Chandler, L.A.; Sosnowski, B.A.; Greenlees, L.; Aukerman, S.L.; Baird, A.; Pierce, G.F. Prevalent expression of fibroblast growth factor (FGF) receptors and FGF2 in human tumor cell lines. Int. J. Cancer 1999, 81, 451–458. [Google Scholar] [CrossRef]
- Pardo, O.E.; Wellbrock, C.; Khanzada, U.K.; Aubert, M.; Arozarena, I.; Davidson, S.; Bowen, F.; Parker, P.J.; Filonenko, V.V.; Gout, I.T.; et al. FGF-2 protects small cell lung cancer cells from apoptosis through a complex involving PKCepsilon, B-Raf and S6K2. EMBO J. 2006, 25, 3078–3088. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, H.; Xu, X.; Shi, H.; Yu, X.; Wang, X.; Yan, Y.; Fu, X.; Hu, H.; Li, X.; et al. bFGF inhibits ER stress induced by ischemic oxidative injury via activation of the PI3K/Akt and ERK1/2 pathways. Toxicol. Lett. 2012, 212, 137–146. [Google Scholar] [CrossRef]
- Nelson, D.L. Lehninger—Principles of Biochemistry, 4th ed.; Freeman, W.H., Ed.; Macmillan: New York, NY, USA, 2005. [Google Scholar]
- Idell-Wenger, J.A.; Grotyohann, L.W.; Neely, J.R. Coenzyme A and carnitine distribution in normal and ischemic hearts. J. Biol. Chem. 1978, 253, 4310–4318. [Google Scholar] [CrossRef]
- Horie, S.; Isobe, M.; Suga, T. Changes in CoA pools in hepatic peroxisomes of the rat, under various conditions. J. Biochem. 1986, 99, 1345–1352. [Google Scholar] [CrossRef]
- Williamson, J.R.; Corkey, B. Assay of citric acid cycle intermediates and related compounds- Update with tissue metabolite levels and intracellular distribution. In Methods in Enzymology—Biomembranes, Bioenergetics: Oxidative Phosphorylation; Academic Press: Cambridge, MA, USA, 1979; Volume 55, pp. 200–222. ISBN 0076-6879. [Google Scholar]
- Fiermonte, G.; Paradies, E.; Todisco, S.; Marobbio, C.M.T.; Palmieri, F. A novel member of solute carrier family 25 (SLC25A42) is a transporter of coenzyme A and adenosine 3’,5’-diphosphate in human mitochondria. J. Biol. Chem. 2009, 284, 18152–18159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, L.K.; Subramanian, C.; Yun, M.K.; Frank, M.W.; White, S.W.; Rock, C.O.; Lee, R.E.; Jackowski, S. A therapeutic approach to pantothenate kinase associated neurodegeneration. Nat. Commun. 2018, 9, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malanchuk, O.M.; Panasyuk, G.G.; Serbin, N.M.; Gout, I.T.; Filonenko, V.V. Generation and characterization of monoclonal antibodies specific to Coenzyme A. Biopolym. Cell 2015, 31, 187–192. [Google Scholar] [CrossRef] [Green Version]
- Thedieck, K.; Holzwarth, B.; Prentzell, M.T.; Boehlke, C.; Kläsener, K.; Ruf, S.; Sonntag, A.G.; Maerz, L.; Grellscheid, S.-N.; Kremmer, E.; et al. Inhibition of mTORC1 by astrin and stress granules prevents apoptosis in cancer cells. Cell 2013, 154, 859–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baković, J.; López Martínez, D.; Nikolaou, S.; Yu, B.Y.K.; Tossounian, M.-A.; Tsuchiya, Y.; Thrasivoulou, C.; Filonenko, V.; Gout, I. Regulation of the CoA Biosynthetic Complex Assembly in Mammalian Cells. Int. J. Mol. Sci. 2021, 22, 1131. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031131
Baković J, López Martínez D, Nikolaou S, Yu BYK, Tossounian M-A, Tsuchiya Y, Thrasivoulou C, Filonenko V, Gout I. Regulation of the CoA Biosynthetic Complex Assembly in Mammalian Cells. International Journal of Molecular Sciences. 2021; 22(3):1131. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031131
Chicago/Turabian StyleBaković, Jovana, David López Martínez, Savvas Nikolaou, Bess Yi Kun Yu, Maria-Armineh Tossounian, Yugo Tsuchiya, Christopher Thrasivoulou, Valeriy Filonenko, and Ivan Gout. 2021. "Regulation of the CoA Biosynthetic Complex Assembly in Mammalian Cells" International Journal of Molecular Sciences 22, no. 3: 1131. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031131