
The Expression of the Cancer-Associated lncRNA Snhg15 Is Modulated by EphrinA5-Induced Signaling

 ,
,  ,
,  , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
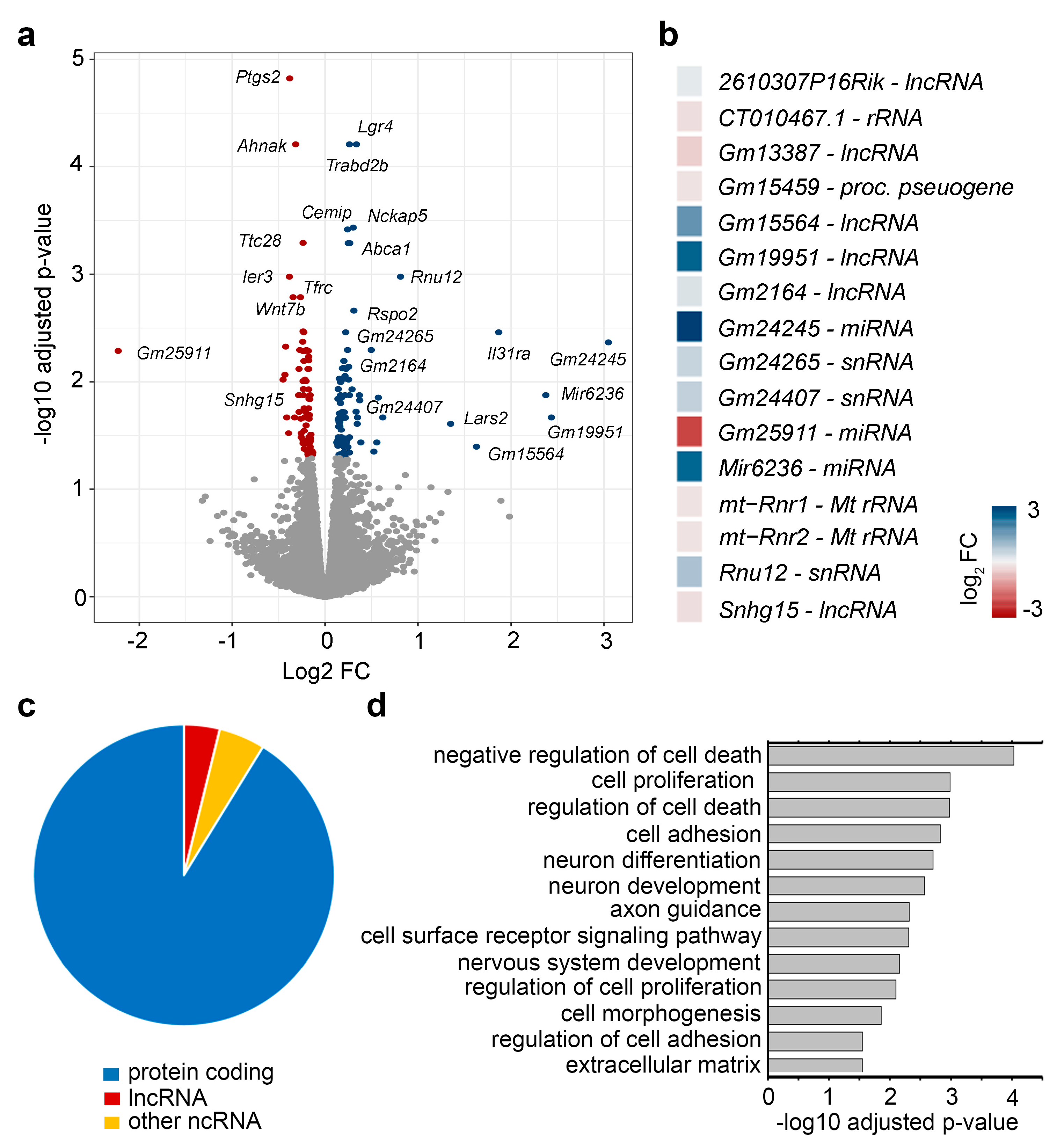
2.1. EphrinA5-Fc Stimulation Induces Changes in the Expression of Non-Coding RNAs in Addition to Protein-Coding Genes
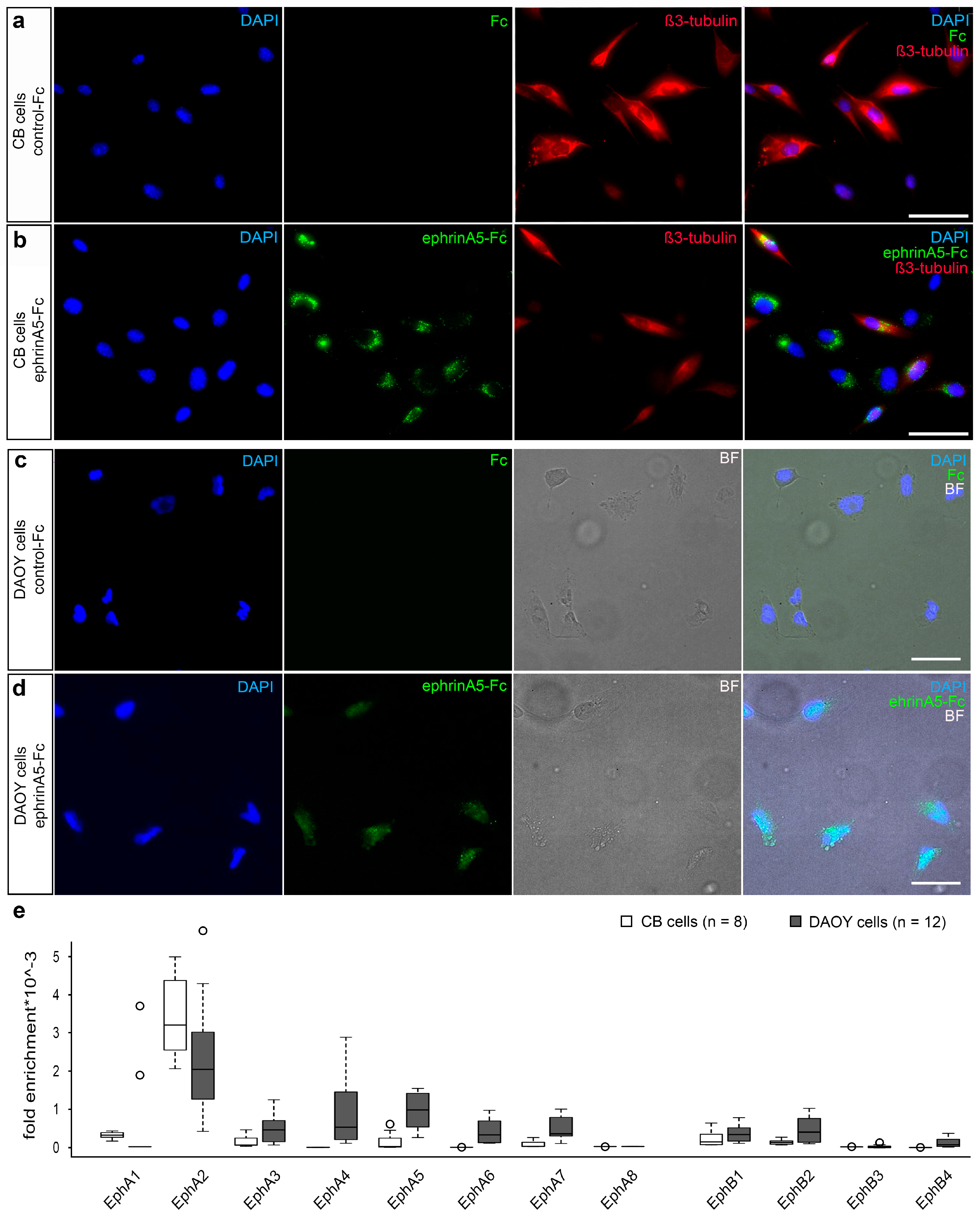
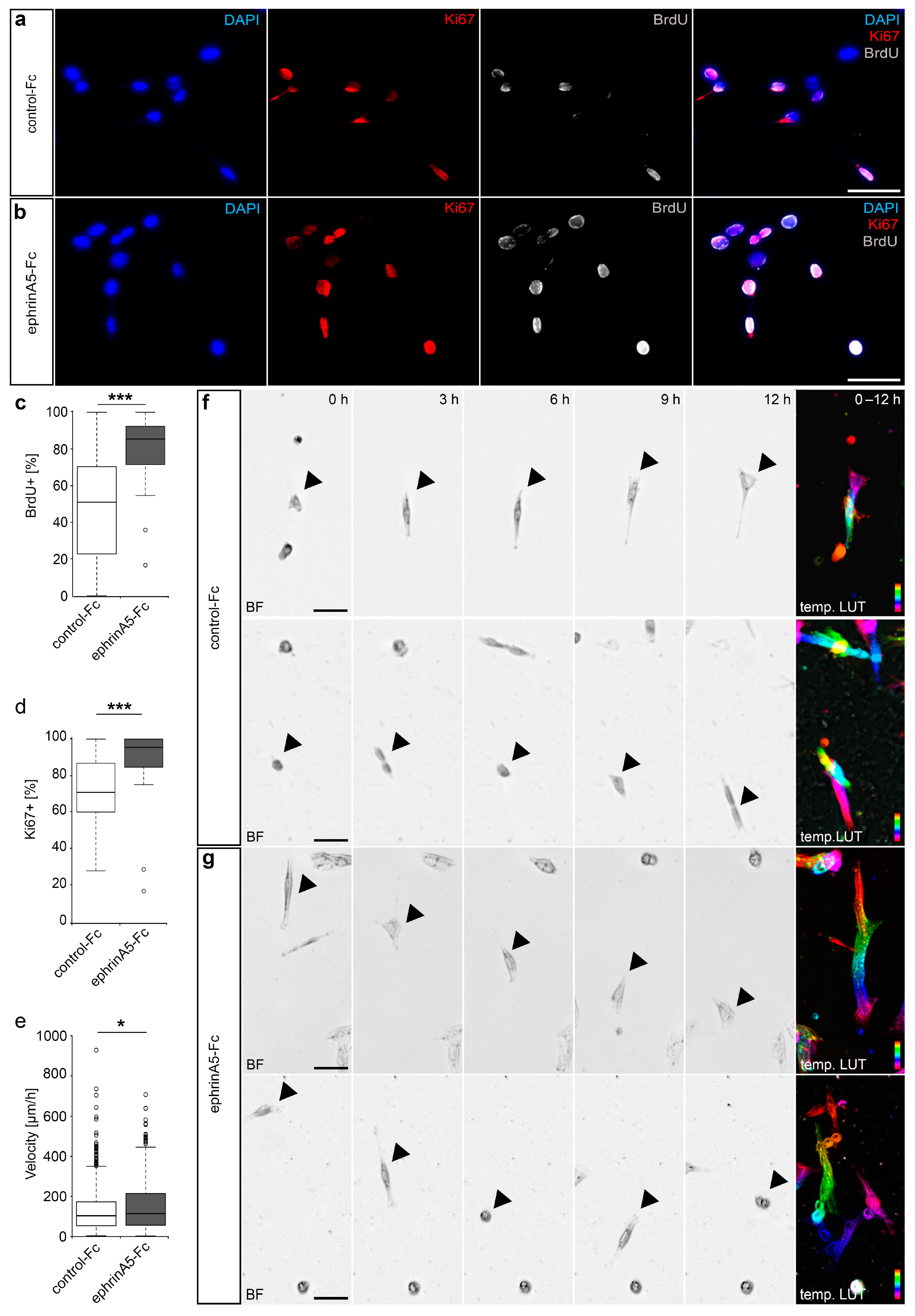
2.2. EphrinA5-Fc Treatment of CB Cells Stimulates Their Motility and Proliferation
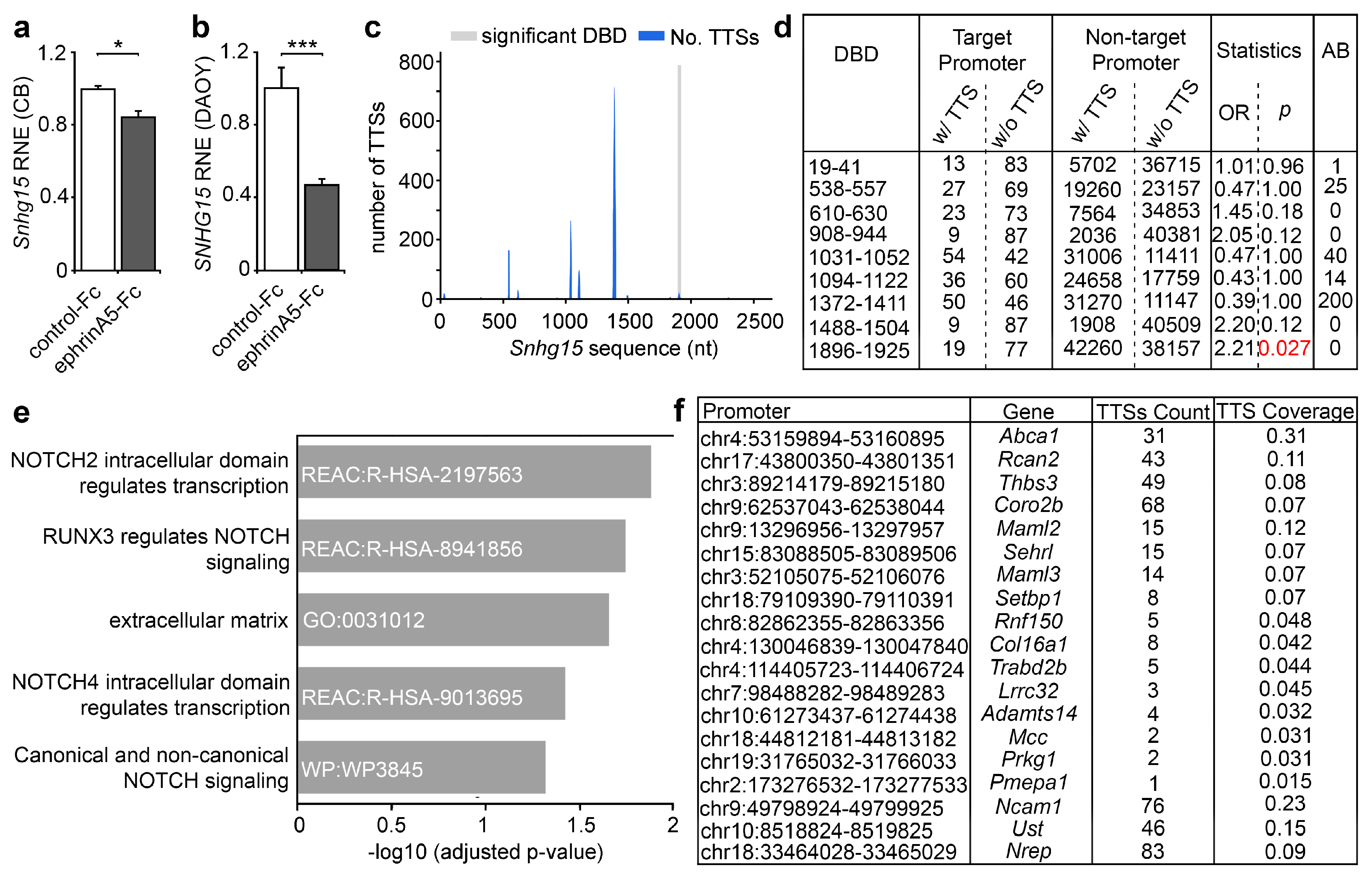
2.3. The lncRNA Snhg15 Is Regulated by EphrinA5-Fc and Is Associated with Proliferation of Cancer Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Treatment with Recombinant EphrinA5-Fc
4.2. Immunocytochemistry
4.3. Migration Assay
4.4. Microscopy and Data Analysis
4.5. Nuclear Enrichment and RNA Isolation of CB Cells
4.6. cDNA Synthesis and Quantitative PCR
4.7. RNA Sequencing
4.8. Alignment, Quality Control, Batch Effect Correction, and Differential Expression Analysis
4.9. LncRNA Binding Prediction
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferluga, S.; Debinski, W. Ephs and Ephrins in malignant gliomas. Growth Factors 2014, 32, 190–201. [Google Scholar] [CrossRef] [Green Version]
- Zimmer, G.; Kästner, B.; Weth, F.; Bolz, J. Multiple Effects of Ephrin-A5 on Cortical Neurons Are Mediated by Src Family Kinases. J. Neurosci. 2007, 27, 5643–5653. [Google Scholar] [CrossRef]
- Zimmer, G.; Garcez, P.; Rudolph, J.; Niehage, R.; Weth, F.; Lent, R.; Bolz, J. Ephrin-A5 acts as a repulsive cue for migrating cortical interneurons. Eur. J. Neurosci. 2008, 28, 62–73. [Google Scholar] [CrossRef]
- Gerstmann, K.; Zimmer, G. The role of the Eph/ephrin family during cortical development and cerebral malformations. Med. Res. Arch. 2018, 6. [Google Scholar] [CrossRef]
- Rudolph, J.; Zimmer, G.; Steinecke, A.; Barchmann, S.; Bolz, J. Ephrins guide migrating cortical interneurons in the basal telencephalon. Cell Adhes. Migr. 2010, 4, 400–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmer, G.; Rudolph, J.; Landmann, J.; Gerstmann, K.; Steinecke, A.; Gampe, C.; Bolz, J. Bidirectional EphrinB3/EphA4 Signaling Mediates the Segregation of Medial Ganglionic Eminence- and Preoptic Area-Derived Interneurons in the Deep and Superficial Migratory Stream. J. Neurosci. 2011, 31, 18364–18380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerstmann, K.; Pensold, D.; Symmank, J.; Khundadze, M.; Hübner, C.A.; Bolz, J.; Zimmer, G. Thalamic afferents influence cortical progenitors via ephrin A5-EphA4 interactions. Development 2015, 142, 140–150. [Google Scholar] [CrossRef] [Green Version]
- Pasquale, E.B. Eph receptors and ephrins in cancer: Bidirectional signalling and beyond. Nat. Rev. Cancer 2010, 10, 165–180. [Google Scholar] [CrossRef] [Green Version]
- Liang, L.-Y.; Patel, O.; Janes, P.W.; Murphy, J.M.; Lucet, I.S. Eph receptor signalling: From catalytic to non-catalytic functions. Oncogene 2019, 38, 6567–6584. [Google Scholar] [CrossRef] [Green Version]
- Vecera, M.; Sana, J.; Lipina, R.; Smrcka, M.; Slaby, O. Long Non-Coding RNAs in Gliomas: From Molecular Pathology to Diagnostic Biomarkers and Therapeutic Targets. Int. J. Mol. Sci. 2018, 19, 2754. [Google Scholar] [CrossRef] [Green Version]
- Beermann, J.; Piccoli, M.-T.; Viereck, J.; Thum, T. Non-coding RNAs in development and disease: Background, mechanisms, and therapeutic approaches. Physiol. Rev. 2016, 96, 1297–1325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iyer, M.K.; Niknafs, Y.S.; Malik, R.; Singhal, U.; Sahu, A.; Hosono, Y.; Barrette, T.R.; Prensner, J.R.; Evans, J.R.; Zhao, S. The landscape of long noncoding RNAs in the human transcriptome. Nat. Genet. 2015, 47, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Tong, J.; Ma, X.; Yu, H.; Yang, J. SNHG15: A promising cancer-related long noncoding RNA. Cancer Manag. Res. 2019, 11, 5961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmer-Bensch, G. Emerging roles of long non-coding RNAs as drivers of brain evolution. Cells 2019, 8, 1399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuo, C.C.; Hanzelmann, S.; Cetin, N.S.; Frank, S.; Zajzon, B.; Derks, J.P.; Akhade, V.S.; Ahuja, G.; Kanduri, C.; Grummt, I.; et al. Detection of RNA-DNA binding sites in long noncoding RNAs. Nucleic Acids Res. 2019. [Google Scholar] [CrossRef] [Green Version]
- Wapinski, O.; Chang, H.Y. Long noncoding RNAs and human disease. Trends Cell Biol. 2011, 21, 354–361. [Google Scholar]
- Yoon, J.-H.; Abdelmohsen, K.; Gorospe, M. Posttranscriptional gene regulation by long noncoding RNA. J. Mol. Biol. 2013, 425, 3723–3730. [Google Scholar] [CrossRef] [Green Version]
- Saeinasab, M.; Bahrami, A.R.; González, J.; Marchese, F.P.; Martinez, D.; Mowla, S.J.; Matin, M.M.; Huarte, M. SNHG15 is a bifunctional MYC-regulated noncoding locus encoding a lncRNA that promotes cell proliferation, invasion and drug resistance in colorectal cancer by interacting with AIF. J. Exp. Clin. Cancer Res. 2019, 38, 1–16. [Google Scholar]
- Wu, D.-M.; Wang, S.; Wen, X.; Han, X.-R.; Wang, Y.-J.; Shen, M.; Fan, S.-H.; Zhang, Z.-F.; Shan, Q.; Li, M.-Q. LncRNA SNHG15 acts as a ceRNA to regulate YAP1-Hippo signaling pathway by sponging miR-200a-3p in papillary thyroid carcinoma. Cell Death Dis. 2018, 9, 1–15. [Google Scholar] [CrossRef]
- Ma, Z.; Huang, H.; Wang, J.; Zhou, Y.; Pu, F.; Zhao, Q.; Peng, P.; Hui, B.; Ji, H.; Wang, K. Long non-coding RNA SNHG15 inhibits P15 and KLF2 expression to promote pancreatic cancer proliferation through EZH2-mediated H3K27me3. Oncotarget 2017, 8, 84153. [Google Scholar] [CrossRef] [Green Version]
- Tani, H.; Torimura, M. Development of cytotoxicity-sensitive human cells using overexpression of long non-coding RNAs. J. Biosci. Bioeng. 2015, 119, 604–608. [Google Scholar] [CrossRef] [PubMed]
- Beck, Z.T.; Xing, Z.; Tran, E.J. LncRNAs: Bridging environmental sensing and gene expression. RNA Biol. 2016, 13, 1189–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briggs, J.A.; Wolvetang, E.J.; Mattick, J.S.; Rinn, J.L.; Barry, G. Mechanisms of long non-coding RNAs in mammalian nervous system development, plasticity, disease, and evolution. Neuron 2015, 88, 861–877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behesti, H.; Marino, S. Cerebellar granule cells: Insights into proliferation, differentiation, and role in medulloblastoma pathogenesis. Int. J. Biochem. Cell Biol. 2009, 41, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Gilbertson, R.J.; Ellison, D.W. The origins of medulloblastoma subtypes. Annu. Rev. Pathol. Mech. Dis. 2008, 3, 341–365. [Google Scholar] [CrossRef]
- Casciati, A.; Tanori, M.; Manczak, R.; Saada, S.; Tanno, B.; Giardullo, P.; Porcù, E.; Rampazzo, E.; Persano, L.; Viola, G.; et al. Human Medulloblastoma Cell Lines: Investigating on Cancer Stem Cell-Like Phenotype. Cancers 2020, 12, 226. [Google Scholar] [CrossRef] [Green Version]
- Himanen, J.-P.; Chumley, M.J.; Lackmann, M.; Li, C.; Barton, W.A.; Jeffrey, P.D.; Vearing, C.; Geleick, D.; Feldheim, D.A.; Boyd, A.W.; et al. Repelling class discrimination: Ephrin-A5 binds to and activates EphB2 receptor signaling. Nat. Neurosci. 2004, 7, 501–509. [Google Scholar] [CrossRef]
- Wykosky, J.; Gibo, D.M.; Stanton, C.; Debinski, W. EphA2 as a novel molecular marker and target in glioblastoma multiforme. Mol. Cancer Res. 2005, 3, 541–551. [Google Scholar] [CrossRef] [Green Version]
- Ricci, B.; Millner, T.O.; Pomella, N.; Zhang, X.; Guglielmi, L.; Badodi, S.; Ceric, D.; Gemma, C.; Cognolato, E.; Zhang, Y. Polycomb-mediated repression of EphrinA5 promotes growth and invasion of glioblastoma. Oncogene 2020, 39, 2523–2538. [Google Scholar] [CrossRef] [Green Version]
- Pasquale, E.B. Eph receptor signalling casts a wide net on cell behaviour. Nat. Rev. Mol. Cell Biol. 2005, 6, 462–475. [Google Scholar] [CrossRef]
- Klein, R. Eph/ephrin signaling in morphogenesis, neural development and plasticity. Curr. Opin. Cell Biol. 2004, 16, 580–589. [Google Scholar] [CrossRef] [PubMed]
- Knöll, B.; Drescher, U. Ephrin-As as receptors in topographic projections. Trends Neurosci. 2002, 25, 145–149. [Google Scholar] [CrossRef]
- Cramer, K.S.; Miko, I.J. Eph-ephrin signaling in nervous system development. F1000Research 2016, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cal, S.; López-Otín, C. ADAMTS proteases and cancer. Matrix Biol. 2015, 44, 77–85. [Google Scholar] [PubMed]
- Guan, F.; Wang, X.; He, F. Promotion of cell migration by neural cell adhesion molecule (NCAM) is enhanced by PSA in a polysialyltransferase-specific manner. PLoS ONE 2015, 10, e0124237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imayoshi, I.; Sakamoto, M.; Yamaguchi, M.; Mori, K.; Kageyama, R. Essential roles of Notch signaling in maintenance of neural stem cells in developing and adult brains. J. Neurosci. 2010, 30, 3489–3498. [Google Scholar] [CrossRef] [Green Version]
- Meurette, O.; Mehlen, P. Notch signaling in the tumor microenvironment. Cancer Cell 2018, 34, 536–548. [Google Scholar] [CrossRef] [Green Version]
- Venkatesh, V.; Nataraj, R.; Thangaraj, G.S.; Karthikeyan, M.; Gnanasekaran, A.; Kaginelli, S.B.; Kuppanna, G.; Kallappa, C.G.; Basalingappa, K.M. Targeting Notch signalling pathway of cancer stem cells. Stem Cell Investig. 2018, 5. [Google Scholar] [CrossRef] [Green Version]
- Zimmer, G.; Schanuel, S.M.; Bürger, S.; Weth, F.; Steinecke, A.; Bolz, J.; Lent, R. Chondroitin Sulfate Acts in Concert with Semaphorin 3A to Guide Tangential Migration of Cortical Interneurons in the Ventral Telencephalon. Cereb. Cortex 2010, 20, 2411–2422. [Google Scholar] [CrossRef] [Green Version]
- Knöll, B.; Drescher, U. Src family kinases are involved in EphA receptor-mediated retinal axon guidance. J. Neurosci. 2004, 24, 6248–6257. [Google Scholar] [CrossRef] [Green Version]
- Dudanova, I.; Klein, R. The axon’s balancing act: Cis-and trans-interactions between Ephs and ephrins. Neuron 2011, 71, 1–3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peuckert, C.; Wacker, E.; Rapus, J.; Levitt, P.; Bolz, J. Adaptive changes in gene expression patterns in the somatosensory cortex after deletion of ephrinA5. Mol. Cell. Neurosci. 2008, 39, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Meier, C.; Anastasiadou, S.; Knöll, B. Ephrin-A5 suppresses neurotrophin evoked neuronal motility, ERK activation and gene expression. PLoS ONE 2011, 6, e26089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iida, H.; Honda, M.; Kawai, H.; Yamashita, T.; Shirota, Y.; Wang, B.; Miao, H.; Kaneko, S. Ephrin-A1 expression contributes to the malignant characteristics of α-fetoprotein producing hepatocellular carcinoma. Gut 2005, 54, 843–851. [Google Scholar] [CrossRef] [Green Version]
- Miao, H.; Wang, B. EphA receptor signaling—Complexity and emerging themes. In Seminars in Cell & Developmental Biology; Elsevier: Amsterdam, The Netherlands, 2012; pp. 16–25. [Google Scholar]
- Duxbury, M.S.; Ito, H.; Zinner, M.J.; Ashley, S.W.; Whang, E.E. EphA2: A determinant of malignant cellular behavior and a potential therapeutic target in pancreatic adenocarcinoma. Oncogene 2004, 23, 1448–1456. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Yuan, X.; Li, Z.; Tu, H.; Li, D.; Qing, J.; Wang, H.; Zhang, L. RNA interference targeting EphA2 inhibits proliferation, induces apoptosis, and cooperates with cytotoxic drugs in human glioma cells. Surg. Neurol. 2008, 70, 562–568; discussion 568–569. [Google Scholar] [CrossRef]
- Amato, K.R.; Wang, S.; Hastings, A.K.; Youngblood, V.M.; Santapuram, P.R.; Chen, H.; Cates, J.M.; Colvin, D.C.; Ye, F.; Brantley-Sieders, D.M.; et al. Genetic and pharmacologic inhibition of EPHA2 promotes apoptosis in NSCLC. J. Clin. Investig. 2014, 124, 2037–2049. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Li, J.; Ma, X.; Yang, Y.; Sun, W.; Jin, W.; Wang, L.; He, Y.; Yang, F.; Yi, Z.; et al. Inhibition of HDACs-EphA2 Signaling Axis with WW437 Demonstrates Promising Preclinical Antitumor Activity in Breast Cancer. EBioMedicine 2018, 31, 276–286. [Google Scholar] [CrossRef]
- Landen, C.N., Jr.; Chavez-Reyes, A.; Bucana, C.; Schmandt, R.; Deavers, M.T.; Lopez-Berestein, G.; Sood, A.K. Therapeutic EphA2 gene targeting in vivo using neutral liposomal small interfering RNA delivery. Cancer Res. 2005, 65, 6910–6918. [Google Scholar] [CrossRef] [Green Version]
- Binda, E.; Visioli, A.; Giani, F.; Lamorte, G.; Copetti, M.; Pitter, K.L.; Huse, J.T.; Cajola, L.; Zanetti, N.; DiMeco, F. The EphA2 receptor drives self-renewal and tumorigenicity in stem-like tumor-propagating cells from human glioblastomas. Cancer Cell 2012, 22, 765–780. [Google Scholar] [CrossRef] [Green Version]
- Miao, H.; Gale, N.W.; Guo, H.; Qian, J.; Petty, A.; Kaspar, J.; Murphy, A.J.; Valenzuela, D.M.; Yancopoulos, G.; Hambardzumyan, D. EphA2 promotes infiltrative invasion of glioma stem cells in vivo through cross-talk with Akt and regulates stem cell properties. Oncogene 2015, 34, 558–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wozniak, M.A.; Modzelewska, K.; Kwong, L.; Keely, P.J. Focal adhesion regulation of cell behavior. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2004, 1692, 103–119. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.; Kinch, M.S.; Burridge, K. Rho-stimulated contractility contributes to the fibroblastic phenotype of Ras-transformed epithelial cells. Mol. Biol. Cell 1997, 8, 2329–2344. [Google Scholar] [CrossRef] [Green Version]
- Sturm, D.; Witt, H.; Hovestadt, V.; Khuong-Quang, D.-A.; Jones, D.T.; Konermann, C.; Pfaff, E.; Tönjes, M.; Sill, M.; Bender, S. Hotspot mutations in H3F3A and IDH1 define distinct epigenetic and biological subgroups of glioblastoma. Cancer Cell 2012, 22, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Xue, Y.; Liu, X.; Qu, C.; Cai, H.; Wang, P.; Li, Z.; Li, Z.; Liu, Y. SNHG15 affects the growth of glioma microvascular endothelial cells by negatively regulating miR-153. Oncol. Rep. 2017, 38, 3265–3277. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Li, J.; Li, F.; Li, M.; Shao, Y.; Wu, L. SNHG15 functions as a tumor suppressor in thyroid cancer. J. Cell. Biochem. 2019, 120, 6120–6126. [Google Scholar] [CrossRef]
- Liu, Y.; Li, J.; Li, M.; Li, F.; Shao, Y.; Wu, L. microRNA-510-5p promotes thyroid cancer cell proliferation, migration, and invasion through suppressing SNHG15. J. Cell. Biochem. 2019, 120, 11738–11744. [Google Scholar] [CrossRef] [PubMed]
- Rutishauser, U.; Acheson, A.; Hall, A.K.; Mann, D.M.; Sunshine, J. The neural cell adhesion molecule (NCAM) as a regulator of cell-cell interactions. Science 1988, 240, 53–57. [Google Scholar] [CrossRef]
- Livingston, B.D.; Jacobs, J.; Glick, M.; Troy, F.A. Extended polysialic acid chains (n greater than 55) in glycoproteins from human neuroblastoma cells. J. Biol. Chem. 1988, 263, 9443–9448. [Google Scholar] [CrossRef]
- Lehembre, F.; Yilmaz, M.; Wicki, A.; Schomber, T.; Strittmatter, K.; Ziegler, D.; Kren, A.; Went, P.; Derksen, P.W.; Berns, A. NCAM-induced focal adhesion assembly: A functional switch upon loss of E-cadherin. EMBO J. 2008, 27, 2603–2615. [Google Scholar] [CrossRef] [Green Version]
- Dalla-Torre, C.A.; Yoshimoto, M.; Lee, C.-H.; Joshua, A.M.; de Toledo, S.R.; Petrilli, A.S.; Andrade, J.A.; Chilton-MacNeill, S.; Zielenska, M.; Squire, J.A. Effects of THBS3, SPARC and SPP1 expression on biological behavior and survival in patients with osteosarcoma. BMC Cancer 2006, 6, 237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratzinger, S.; Grässel, S.; Dowejko, A.; Reichert, T.E.; Bauer, R.J. Induction of type XVI collagen expression facilitates proliferation of oral cancer cells. Matrix Biol. 2011, 30, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Wu, W.; Liu, W.; Ren, F. Mini-Review: GARP, a Putative Potential Molecule in Tumor Immunosuppressive Environment. J. Cancer Treat. Diagn. 2019, 3, 14–18. [Google Scholar] [CrossRef]
- Kitagawa, M. Notch signalling in the nucleus: Roles of Mastermind-like (MAML) transcriptional coactivators. J. Biochem. 2016, 159, 287–294. [Google Scholar] [CrossRef] [Green Version]
- Heynen, G.J.J.E.; Nevedomskaya, E.; Palit, S.; Jagalur Basheer, N.; Lieftink, C.; Schlicker, A.; Zwart, W.; Bernards, R.; Bajpe, P.K. Mastermind-Like 3 Controls Proliferation and Differentiation in Neuroblastoma. Mol. Cancer Res. 2016, 14, 411. [Google Scholar] [CrossRef] [Green Version]
- Köchert, K.; Ullrich, K.; Kreher, S.; Aster, J.C.; Kitagawa, M.; Jöhrens, K.; Anagnostopoulos, I.; Jundt, F.; Lamprecht, B.; Zimber-Strobl, U.; et al. High-level expression of Mastermind-like 2 contributes to aberrant activation of the NOTCH signaling pathway in human lymphomas. Oncogene 2011, 30, 1831–1840. [Google Scholar] [CrossRef] [Green Version]
- Fossale, E.; Wolf, P.; Espinola, J.A.; Lubicz-Nawrocka, T.; Teed, A.M.; Gao, H.; Rigamonti, D.; Cattaneo, E.; MacDonald, M.E.; Cotman, S.L.; et al. Membrane trafficking and mitochondrial abnormalities precede subunit c deposition in a cerebellar cell model of juvenile neuronal ceroid lipofuscinosis. BMC Neurosci. 2004, 5, 57. [Google Scholar] [CrossRef] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Ewels, P.A.; Peltzer, A.; Fillinger, S.; Patel, H.; Alneberg, J.; Wilm, A.; Garcia, M.U.; Di Tommaso, P.; Nahnsen, S. The nf-core framework for community-curated bioinformatics pipelines. Nat. Biotechnol. 2020, 38, 276–278. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pensold, D.; Gehrmann, J.; Pitschelatow, G.; Walberg, A.; Braunsteffer, K.; Reichard, J.; Ravaei, A.; Linde, J.; Lampert, A.; Costa, I.G.; et al. The Expression of the Cancer-Associated lncRNA Snhg15 Is Modulated by EphrinA5-Induced Signaling. Int. J. Mol. Sci. 2021, 22, 1332. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031332
Pensold D, Gehrmann J, Pitschelatow G, Walberg A, Braunsteffer K, Reichard J, Ravaei A, Linde J, Lampert A, Costa IG, et al. The Expression of the Cancer-Associated lncRNA Snhg15 Is Modulated by EphrinA5-Induced Signaling. International Journal of Molecular Sciences. 2021; 22(3):1332. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031332
Chicago/Turabian StylePensold, Daniel, Julia Gehrmann, Georg Pitschelatow, Asa Walberg, Kai Braunsteffer, Julia Reichard, Amin Ravaei, Jenice Linde, Angelika Lampert, Ivan G. Costa, and et al. 2021. "The Expression of the Cancer-Associated lncRNA Snhg15 Is Modulated by EphrinA5-Induced Signaling" International Journal of Molecular Sciences 22, no. 3: 1332. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031332