
Folinate Supplementation Ameliorates Methotrexate Induced Mitochondrial Formate Depletion In Vitro and In Vivo


 ,
,
Abstract
:1. Introduction
2. Results
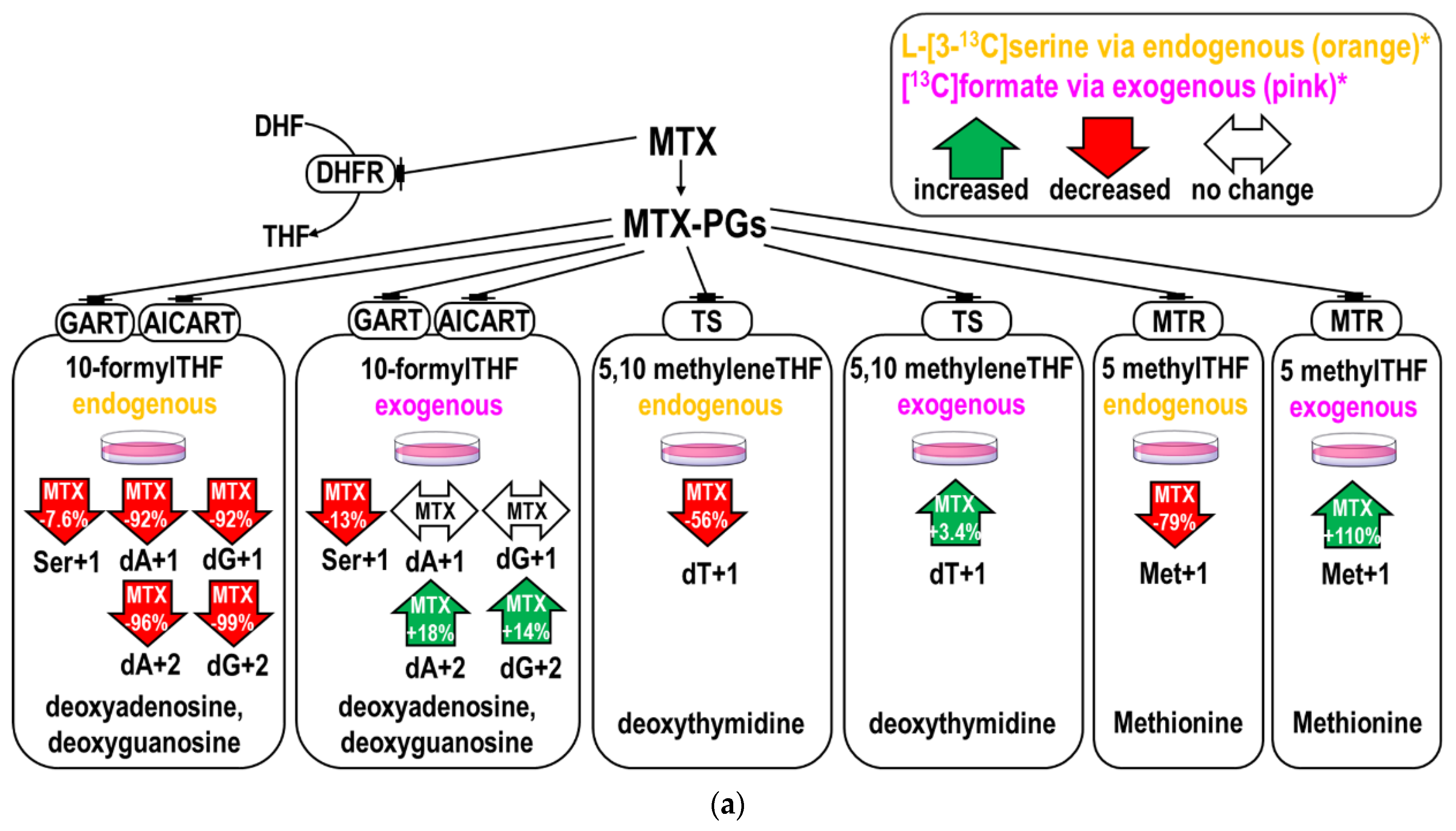
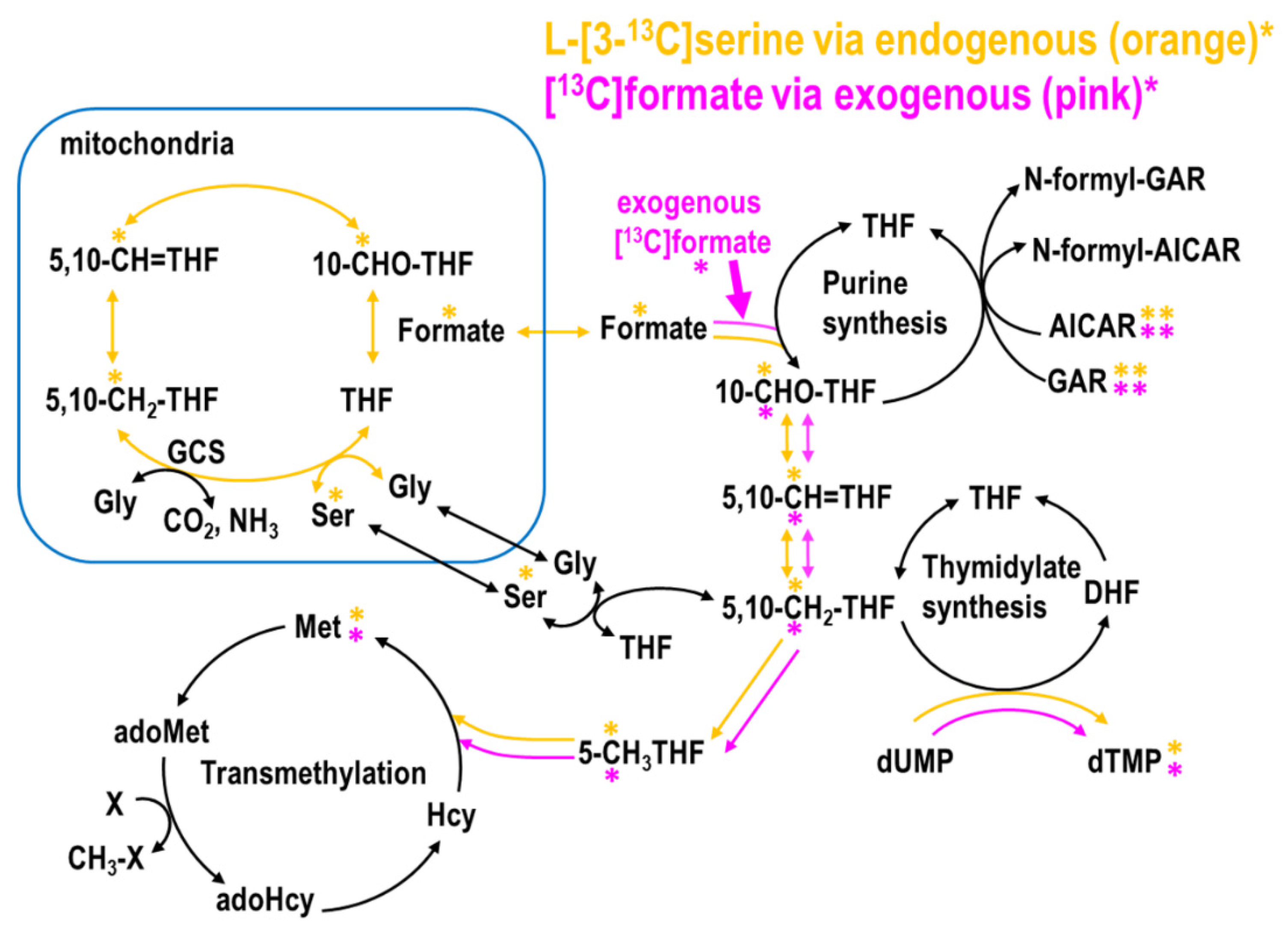
2.1. MTX Inhibited Endogenous and Promoted Exogenous Formate Utilization for Nucleotide and Methionine Biosyntheses In Vitro
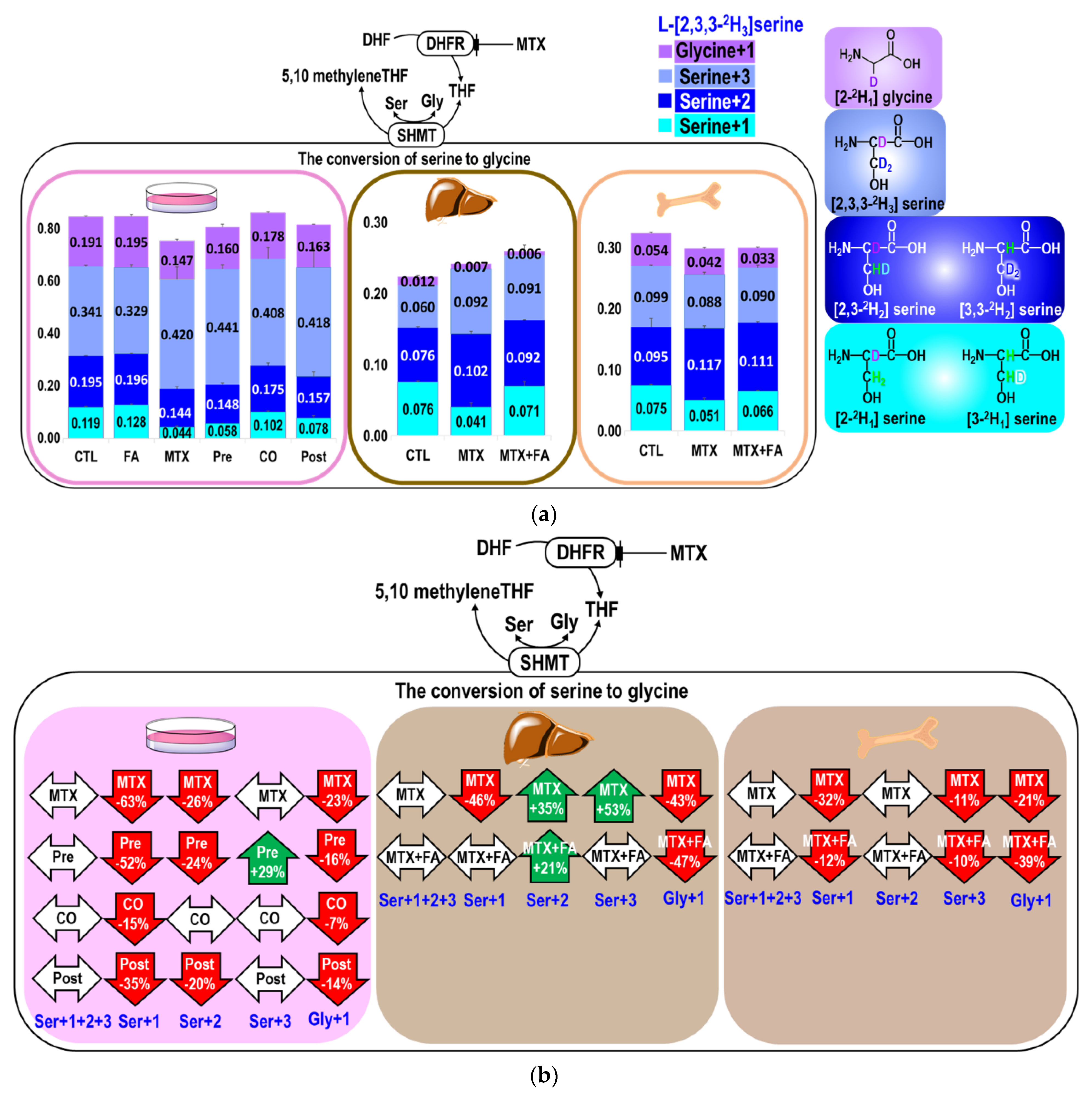
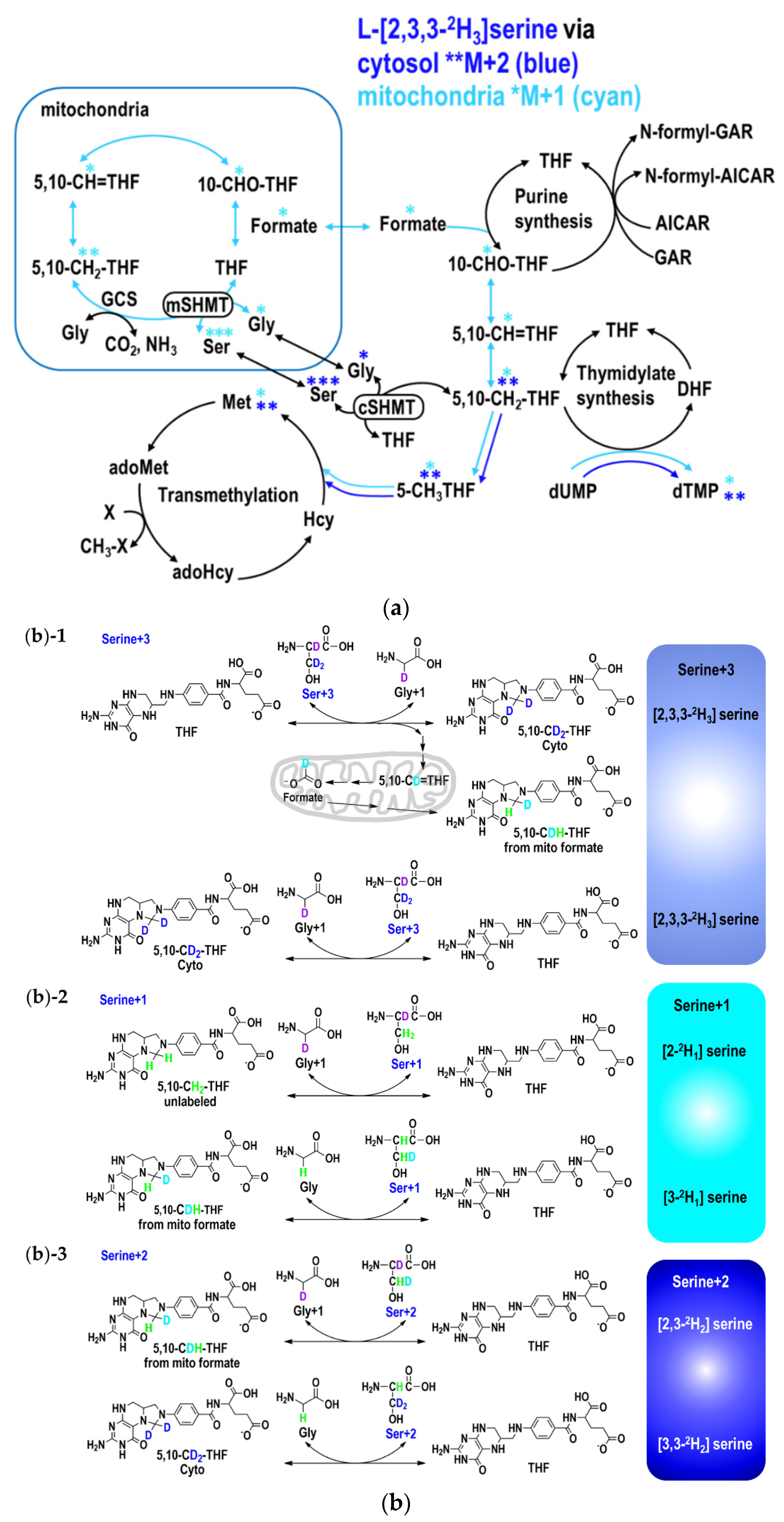
2.2. MTX Interferes with the Conversion between Serine and Glycine
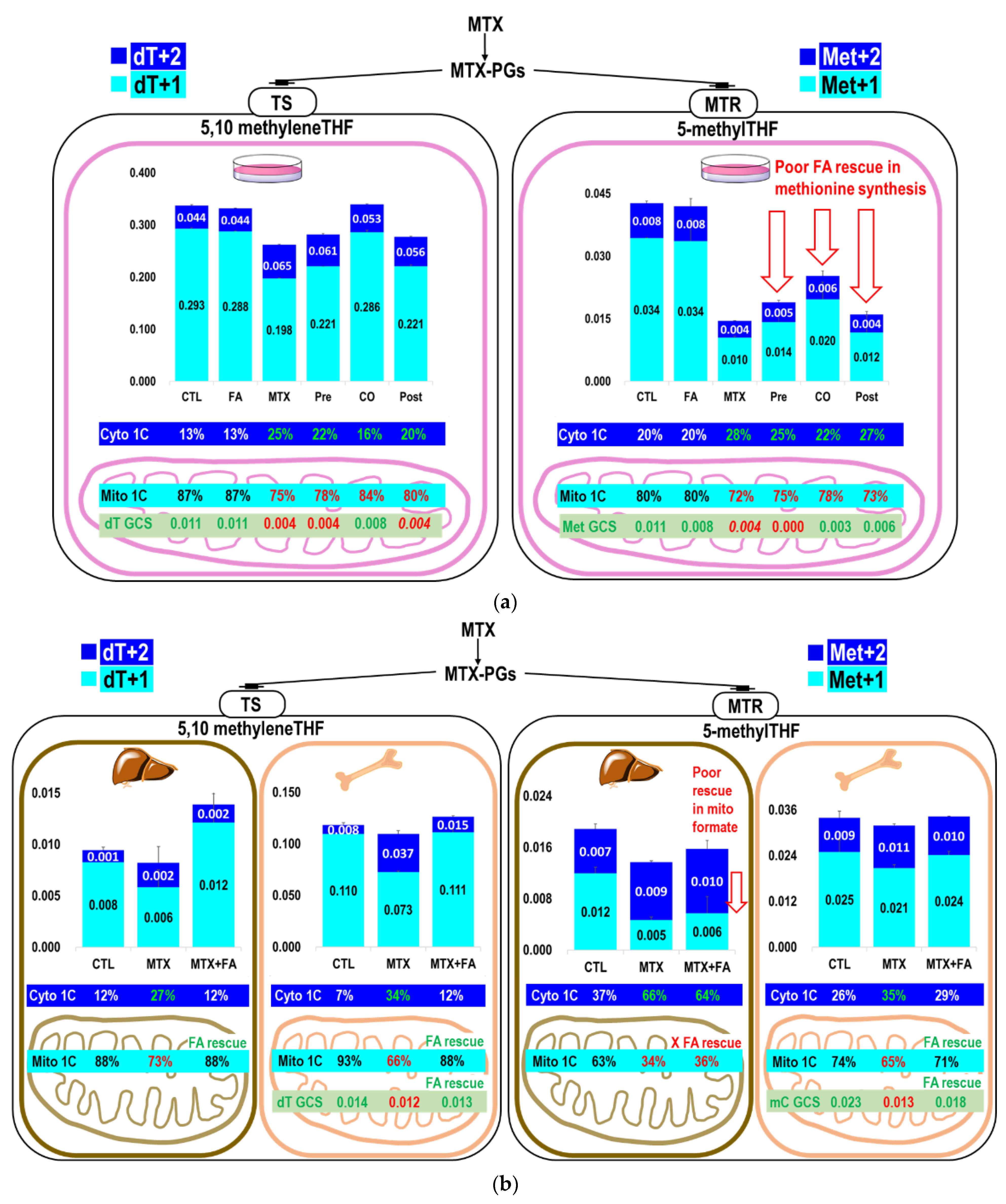
2.3. The Partitioning of 1C Metabolic Fuxes via MethyleneTHF between Mitochondria- and Cytosolic-Derived Formate
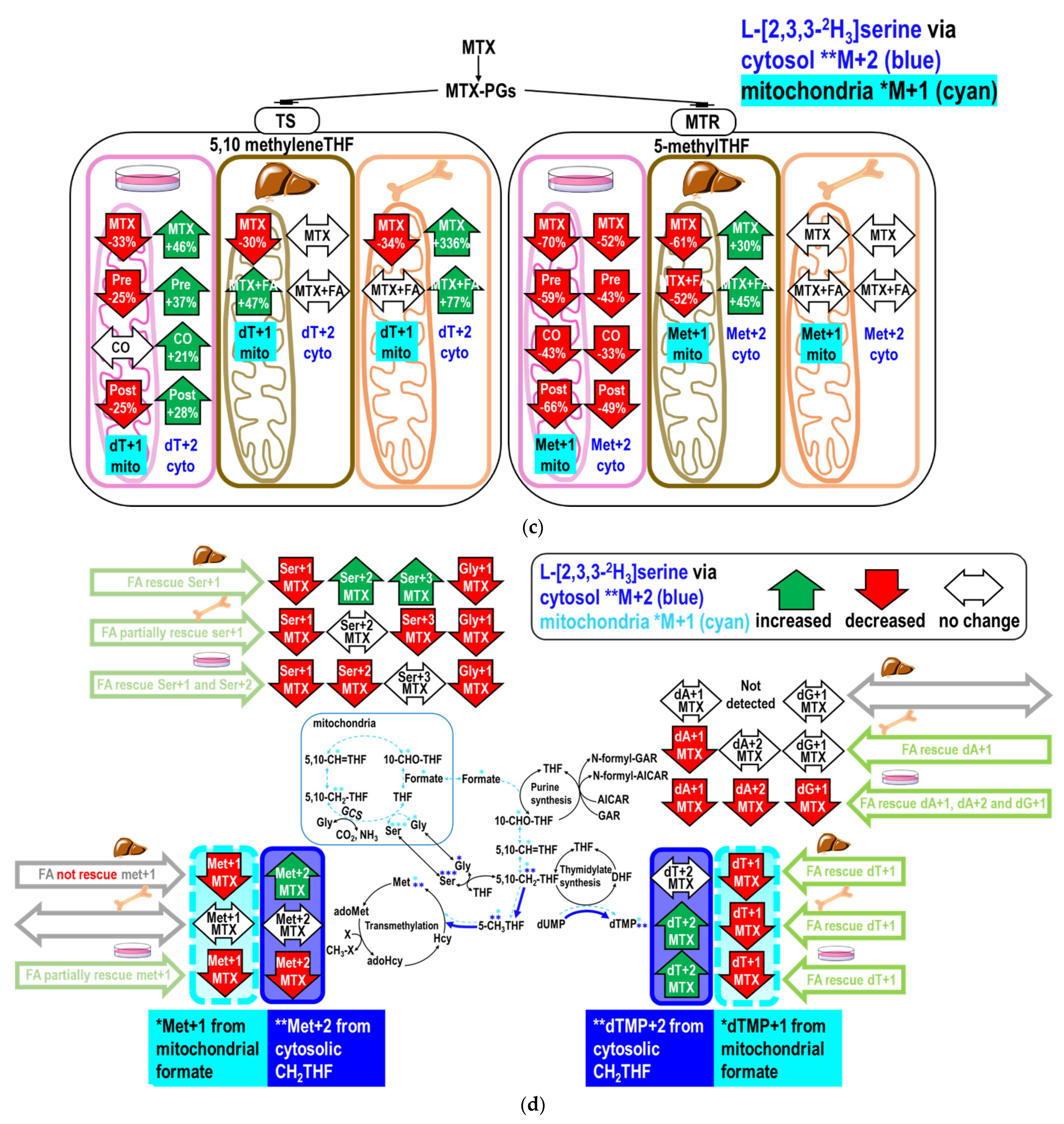
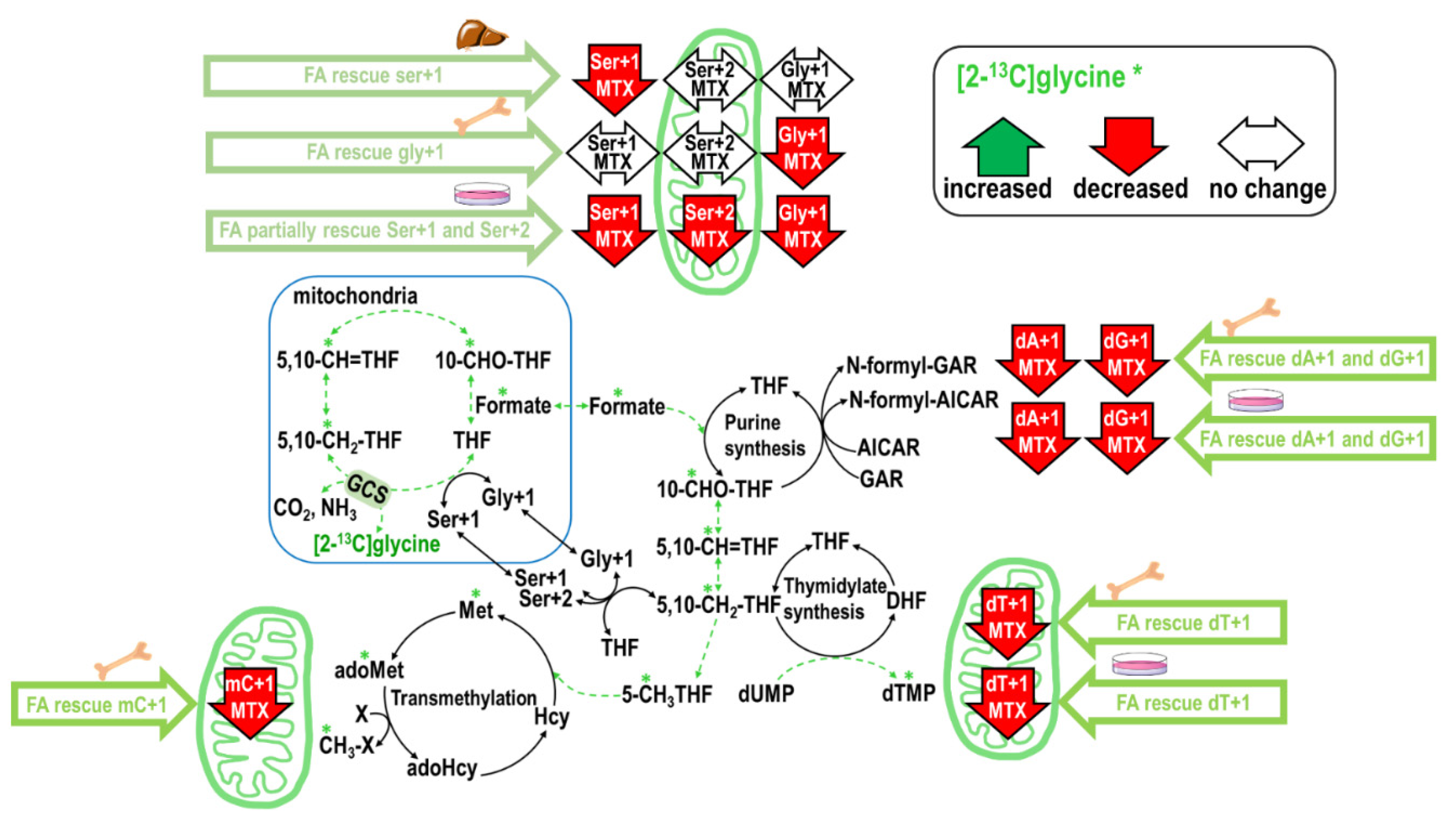
2.4. Folinate Supplementation Rescued the MTX-Inhibited De Novo Thymidylate Biosynthesis from Glycine via GCS
3. Discussion
3.1. Low-Dose MTX Depletes Mitochondrial Formate Production
3.2. Low-Dose MTX Changes the Partitioning between Mitochondrial and Cytosolic 1C Metabolic Fluxes
3.3. Mitochondria Formate Supply for Folate-Mediated dTMP and Methionine Synthesis in Bone Marrow and Liver
3.4. Inhibition of 1C Supply by MTX Is Specific to Tissue, Pathway, and Cellular Compartments
3.5. MTX Inhibits Formate Supply from the Mitochondria Glycine Cleavage System
3.6. Folinate Rescues 1C Supply for Nucleotides but Failed to Rescue Hepatic Transmethylation During MTX Therapy
4. Materials and Methods
4.1. Cell Model, Culture Condition, and Treatment Protocol
4.2. Effects of Low-Dose MTX on Endogenous and Exogenous Formate Utilization In Vitro
4.3. Partitioning between Mitochondrial and Cytosolic 1C Metabolism
4.4. Tracing the Metabolic Fate of Mitochondria-Derived Formate from Glycine Cleavage System
4.5. Animal Study Design
4.6. Gas Chromatography/Mass Spectrometry Analysis
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Morrow, G.P.; MacMillan, L.; Lamarre, S.G.; Young, S.K.; MacFarlane, A.J.; Brosnan, M.E.; Brosnan, J.T. In vivo kinetics of formate metabolism in folate-deficient and folate-replete rats. J. Biol. Chem. 2015, 290, 2244–2250. [Google Scholar] [CrossRef] [Green Version]
- Tibbetts, A.S.; Appling, D.R. Compartmentalization of Mammalian folate-mediated one-carbon metabolism. Annu. Rev. Nutr. 2010, 30, 57–81. [Google Scholar] [CrossRef] [PubMed]
- Pietzke, M.; Meiser, J.; Vazquez, A. Formate metabolism in health and disease. Mol. Metab. 2019, 33, 23–37. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Lin, T.Y.; Lee, G.; Paddock, M.N.; Momb, J.; Cheng, Z.; Li, Q.; Fei, D.L.; Stein, B.D.; Ramsamooj, S.; et al. Mitochondrial One-Carbon Pathway Supports Cytosolic Folate Integrity in Cancer Cells. Cell 2018, 175, 1546–1560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ducker, G.S.; Chen, L.; Morscher, R.J.; Ghergurovich, J.M.; Esposito, M.; Teng, X.; Kang, Y.; Rabinowitz, J.D. Reversal of Cytosolic One-Carbon Flux Compensates for Loss of the Mitochondrial Folate Pathway. Cell Metab. 2016, 23, 1140–1153. [Google Scholar] [CrossRef] [Green Version]
- Burgos-Barragan, G.; Wit, N.; Meiser, J.; Dingler, F.A.; Pietzke, M.; Mulderrig, L.; Pontel, L.B.; Rosado, I.V.; Brewer, T.F.; Cordell, R.L.; et al. Mammals divert endogenous genotoxic formaldehyde into one-carbon metabolism. Nature 2017, 548, 549–554. [Google Scholar] [CrossRef] [Green Version]
- Selhub, J.; Seyoum, E.; Pomfret, E.A.; Zeisel, S.H. Effects of choline deficiency and methotrexate treatment upon liver folate content and distribution. Cancer Res. 1991, 51, 16–21. [Google Scholar]
- Meiser, J.; Tumanov, S.; Maddocks, O.; Labuschagne, C.F.; Athineos, D.; Van Den Broek, N.; Mackay, G.M.; Gottlieb, E.; Blyth, K.; Vousden, K.; et al. Serine one-carbon catabolism with formate overflow. Sci. Adv. 2016, 2, e1601273. [Google Scholar] [CrossRef] [Green Version]
- Tedeschi, P.M.; Johnson-Farley, N.; Lin, H.; Shelton, L.M.; Ooga, T.; Mackay, G.; Van Den Broek, N.; Bertino, J.R.; Vazquez, A. Quantification of folate metabolism using transient metabolic flux analysis. Cancer Metab. 2015, 3, 6. [Google Scholar]
- Bao, X.R.; Ong, S.E.; Goldberger, O.; Peng, J.; Sharma, R.; Thompson, D.A.; Vafai, S.B.; Cox, A.G.; Marutani, E.; Ichinose, F.; et al. Mitochondrial dysfunction remodels one-carbon metabolism in human cells. eLife 2016, 5, e10575. [Google Scholar] [CrossRef]
- Garcia-Martinez, L.F.; Appling, D.R. Characterization of the folate-dependent mitochondrial oxidation of carbon 3 of serine. Biochemistry 1993, 32, 4671–4676. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Kim, H.; Fu, Z.; Qiu, Y.; Yang, Z.; Wang, J.; Zhang, D.; Tong, X.; Yin, L.; Li, J.; et al. Deficiency of the Mitochondrial NAD Kinase Causes Stress-Induced Hepatic Steatosis in Mice. Gastroenterology 2018, 154, 224–237. [Google Scholar] [CrossRef] [PubMed]
- Hess, J.A.; Khasawneh, M.K. Cancer metabolism and oxidative stress: Insights into carcinogenesis and chemotherapy via the non-dihydrofolate reductase effects of methotrexate. BBA Clin. 2015, 3, 152–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabassum, H.; Parvez, S.; Pasha, S.T.; Banerjee, B.D.; Raisuddin, S. Protective effect of lipoic acid against methotrexate-induced oxidative stress in liver mitochondria. Food Chem. Toxicol. 2010, 48, 1973–1979. [Google Scholar] [CrossRef]
- Kolli, V.K.; Natarajan, K.; Isaac, B.; Selvakumar, D.; Abraham, P. Mitochondrial dysfunction and respiratory chain defects in a rodent model of methotrexate-induced enteritis. Hum. Exp. Toxicol. 2014, 33, 1051–1065. [Google Scholar] [CrossRef]
- Chiang, E.P.; Wang, Y.C.; Tang, F.Y. Folate restriction and methylenetetrahydrofolate reductase 677T polymorphism decreases adoMet synthesis via folate-dependent remethylation in human-transformed lymphoblasts. Leukemia 2007, 21, 651–658. [Google Scholar] [CrossRef]
- Herbig, K.; Chiang, E.P.; Lee, L.R.; Hills, J.; Shane, B.; Stover, P.J. Cytoplasmic serine hydroxymethyltransferase mediates competition between folate-dependent deoxyribonucleotide and S-adenosylmethionine biosyntheses. J. Biol. Chem. 2002, 277, 38381–38389. [Google Scholar]
- Anderson, D.D.; Woeller, C.F.; Chiang, E.P.; Shane, B.; Stover, P.J. Serine hydroxymethyltransferase anchors de novo thymidylate synthesis pathway to nuclear lamina for DNA synthesis. J. Biol. Chem. 2012, 287, 7051–7062. [Google Scholar]
- Wang, Y.C.; Tang, F.Y.; Chen, S.Y.; Chen, Y.M.; Chiang, E.P. Glycine-N methyltransferase expression in HepG2 cells is involved in methyl group homeostasis by regulating transmethylation kinetics and DNA methylation. J. Nutr. 2011, 141, 777–782. [Google Scholar] [CrossRef] [Green Version]
- Anguera, M.C.; Field, M.S.; Perry, C.; Ghandour, H.; Chiang, E.P.; Selhub, J.; Shane, B.; Stover, P.J. Regulation of folate-mediated one-carbon metabolism by 10-formyltetrahydrofolate dehydrogenase. J. Biol. Chem. 2006, 281, 18335–18342. [Google Scholar]
- Tan, Y.L.; Sou, N.L.; Tang, F.Y.; Ko, H.A.; Yeh, W.T.; Peng, J.H.; Chiang, E.I. Tracing Metabolic Fate of Mitochondrial Glycine Cleavage System Derived Formate In vitro and In vivo. Int. J. Mol. Sci. 2020, 21, 8808. [Google Scholar] [CrossRef] [PubMed]
- Baggott, J.E.; Gorman, G.S.; Tamura, T. 13C enrichment of carbons 2 and 8 of purine by folate-dependent reactions after [13C]formate and [2-13C]glycine dosing in adult humans. Metabolism 2007, 56, 708–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brosnan, M.E.; MacMillan, L.; Stevens, J.R.; Brosnan, J.T. Division of labour: How does folate metabolism partition between one-carbon metabolism and amino acid oxidation? Biochem. J. 2015, 472, 135–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, D.D.; Quintero, C.M.; Stover, P.J. Identification of a de novo thymidylate biosynthesis pathway in mammalian mitochondria. Proc. Natl. Acad. Sci. USA 2011, 108, 15163–15168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.C.; Wu, M.T.; Tang, F.Y.; Chen, D.Y.; Ko, H.A.; Shane, B.; Huang, W.N.; Chiang, E.P. MTHFR C677T polymorphism increases MTX sensitivity via the inhibition of S-adenosylmethionine and de novo purine synthesis. Clin. Sci. 2019, 133, 253–267. [Google Scholar] [CrossRef] [PubMed]
- Stempak, J.M.; Sohn, K.J.; Chiang, E.P.; Shane, B.; Kim, Y.I. Cell and stage of transformation-specific effects of folate deficiency on methionine cycle intermediates and DNA methylation in an in vitro model. Carcinogenesis 2005, 26, 981–990. [Google Scholar] [CrossRef]
- Sohn, K.J.; Jang, H.; Campan, M.; Weisenberger, D.J.; Dickhout, J.; Wang, Y.C.; Cho, R.C.; Yates, Z.; Lucock, M.; Chiang, E.P.; et al. The methylenetetrahydrofolate reductase C677T mutation induces cell-specific changes in genomic DNA methylation and uracil misincorporation: A possible molecular basis for the site-specific cancer risk modification. Int. J. Cancer 2009, 124, 1999–2005. [Google Scholar] [PubMed] [Green Version]
- Horne, D.W. Effects of nitrous oxide inactivation of vitamin B12 and of methionine on folate coenzyme metabolism in rat liver, kidney, brain, small intestine and bone marrow. Biofactors 1989, 2, 65–68. [Google Scholar]
- Deacon, R.; Chanarin, I.; Lumb, M.; Perry, J. Role of folate dependent transformylases in synthesis of purine in bone marrow of man and in bone marrow and liver of rats. J. Clin. Pathol. 1985, 38, 1349–1352. [Google Scholar]
- Wang, Y.C.; Chiang, E.P. Low-dose methotrexate inhibits methionine S-adenosyltransferase in vitro and in vivo. Mol. Med. 2012, 18, 423–432. [Google Scholar] [CrossRef]
- Chen, S.Y.; Lin, J.R.; Darbha, R.; Lin, P.; Liu, T.Y.; Chen, Y.M. Glycine N-methyltransferase tumor susceptibility gene in the benzo(a)pyrene-detoxification pathway. Cancer Res. 2004, 64, 3617–3623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.Y.; Guan, D.Y.; Yang, M.; Wang, H.; Shen, Z.H. All-trans-retinoic acid intensifies endoplasmic reticulum stress in N-acetylglucosaminyltransferase V repressed human hepatocarcinoma cells by perturbing homocysteine metabolism. J. Cell. Biochem. 2010, 109, 468–477. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.C.; Chen, Y.M.; Lin, Y.J.; Liu, S.P.; Chiang, E.P. GNMT expression increases hepatic folate contents and folate-dependent methionine synthase-mediated homocysteine remethylation. Mol. Med. 2011, 17, 486–494. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.C.; Wu, M.T.; Lin, Y.J.; Tang, F.Y.; Ko, H.A.; Chiang, E.P. Regulation of Folate-Mediated One-Carbon Metabolism by Glycine N-Methyltransferase (GNMT) and Methylenetetrahydrofolate Reductase (MTHFR). J. Nutr. Sci. Vitaminol. 2015, 61, 148–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, E.P.; Wang, Y.C.; Chen, W.W.; Tang, F.Y. Effects of insulin and glucose on cellular metabolic fluxes in homocysteine transsulfuration, remethylation, S-adenosylmethionine synthesis, and global deoxyribonucleic acid methylation. J. Clin. Endocrinol. Metab. 2009, 94, 1017–1025. [Google Scholar] [CrossRef] [Green Version]
- Weisman, M.H.; Furst, D.E.; Park, G.S.; Kremer, J.M.; Smith, K.M.; Wallace, D.J.; Caldwell, J.R.; Dervieux, T. Risk genotypes in folate-dependent enzymes and their association with methotrexate-related side effects in rheumatoid arthritis. Arthritis Rheum. 2006, 54, 607–612. [Google Scholar] [CrossRef] [Green Version]
- Gregory, J.F., 3rd; Cuskelly, G.J.; Shane, B.; Toth, J.P.; Baumgartner, T.G.; Stacpoole, P.W. Primed, constant infusion with [2H3]serine allows in vivo kinetic measurement of serine turnover, homocysteine remethylation, and transsulfuration processes in human one-carbon metabolism. Am. J. Clin. Nutr. 2000, 72, 1535–1541. [Google Scholar] [CrossRef]
- Chang, H.Y.; Tzen, J.T.; Lin, S.J.; Wu, Y.T.; Chiang, E.P. Long-term prednisolone treatments increase bioactive vitamin B6 synthesis in vivo. J. Pharmacol. Exp. Ther. 2011, 337, 102–109. [Google Scholar]
- Sandberg, J.S.; Goldin, A. The use of leucovorin orally in normal and leukemic L1210 mice to prevent the toxicity and gastrointestinal lesions caused by high doses of methotrexate. Cancer Res. 1970, 30, 1276–1280. [Google Scholar]
- Chiang, E.P.; Selhub, J.; Bagley, P.J.; Dallal, G.; Roubenoff, R. Pyridoxine supplementation corrects vitamin B6 deficiency but does not improve inflammation in patients with rheumatoid arthritis. Arthritis Res. Ther. 2005, 7, R1404–R1411. [Google Scholar] [CrossRef] [Green Version]
- Chiang, E.P.; Smith, D.E.; Selhub, J.; Dallal, G.; Wang, Y.C.; Roubenoff, R. Inflammation causes tissue-specific depletion of vitamin B6. Arthritis Res. Ther. 2005, 7, R1254–R1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, D.Y.; Chih, H.M.; Lan, J.L.; Chang, H.Y.; Chen, W.W.; Chiang, E.P. Blood lipid profiles and peripheral blood mononuclear cell cholesterol metabolism gene expression in patients with and without methotrexate treatment. BMC Med. 2011, 9, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Ede, A.E.; Laan, R.F.; Rood, M.J.; Huizinga, T.W.; van de Laar, M.A.; van Denderen, C.J.; Westgeest, T.A.; Romme, T.C.; de Rooij, D.J.; Jacobs, M.J.; et al. Effect of folic or folinic acid supplementation on the toxicity and efficacy of methotrexate in rheumatoid arthritis: A forty-eight week, multicenter, randomized, double-blind, placebo-controlled study. Arthritis Rheum. 2001, 44, 1515–1524. [Google Scholar] [CrossRef]
- Welty, F.K.; Lichtenstein, A.H.; Barrett, P.H.; Dolnikowski, G.G.; Ordovas, J.M.; Schaefer, E.J. Production of apolipoprotein B-67 in apolipoprotein B-67/B-100 heterozygotes: Technical problems associated with leucine contamination in stable isotope studies. J. Lipid Res. 1997, 38, 1535–1543. [Google Scholar] [CrossRef]
- Wang, Y.C.; Lin, W.L.; Lin, Y.J.; Tang, F.Y.; Chen, Y.M.; Chiang, E.P. A novel role of the tumor suppressor GNMT in cellular defense against DNA damage. Int. J. Cancer 2014, 134, 799–810. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (A) Purine synthesis from L-[3-13C]serine derived formate and exogenous [13C]formate 1,2. | ||||||
| Endogenous Formate from L-[3-13C]serine | Exogenous [13C]formate Incorporation | |||||
| dA+1 3 | dA+2 3 | dA (MIA) 4,6 | dA+1 3 | dA+2 3 | dA (MIA) 4,6 | |
| CTL 4 | 0.310 ± 0.003 | 0.179 ± 0.003 | 1.155 ± 0.021 | 0.319 ± 0.002 | 0.121 ± 0.003 | 0.756 ± 0.025 |
| MTX 4 | 0.025 ± 0.014 | 0.008 ± 0.002 | 0.648 ± 0.154 | 0.324 ± 0.005 | 0.142 ±0.003 | 0.876 ± 0.007 |
| p-value2 | 0.001 | 0.000 | 0.044 | 0.281 | 0.022 | 0.023 |
| %change5 | −92.06% | −95.54% | −50.73% | 1.65% | +17.72% | +11.94% |
| dG+1 3 | dG+2 3 | dG (MIA) 4,6 | dG+1 3 | dG+2 3 | dG (MIA) 4,6 | |
| CTL 4 | 0.321 ± 0.015 | 0.171 ± 0.007 | 1.066 ± 0.042 | 0.323 ± 0.008 | 0.119 ± 0.001 | 0.736 ± 0.013 |
| MTX 4 | 0.025 ± 0.010 | 0.001 ± 0.002 | 0.120 ± 0.169 | 0.336 ± 0.006 | 0.135 ± 0.002 | 0.802 ± 0.001 |
| p-value2 | 0.002 | 0.001 | 0.017 | 0.199 | 0.011 | 0.019 |
| %change5 | −92.34% | −99.14% | −94.59% | 4.27% | +13.70% | +6.64% |
| (B) Relative incorporation of L-[3-13C]serine derived formate and exogenous [13C]formate in purines 1,2. | ||||||
| Endogenous Formate from L-[3-13C]serine | Exogenous [13C]formate Incorporation | |||||
| Ser+1 3 | dA+1 3 | dA+1/Ser+1 7 | Ser+1 3 | dA+1 3 | ||
| CTL 4 | 0.515 ± 0.016 | 0.310 ± 0.003 | 0.602 ± 0.006 | 0.189 ± 0.007 | 0.319 ± 0.002 | |
| MTX 4 | 0.475 ± 0.008 | 0.025 ± 0.014 | 0.052 ± 0.029 | 0.166 ± 0.000 | 0.324 ± 0.005 | |
| p-value2 | 0.087 | 0.001 | 0.001 | 0.046 | 0.281 | |
| %change5 | −7.60% | −92.06% | −55.05% | −12.50% | 1.65% | |
| Ser+1 3 | dG+1 3 | dG+1/Ser+1 7 | Ser+1 3 | dG+1 3 | ||
| CTL 4 | 0.515 ± 0.016 | 0.321 ± 0.015 | 0.624 ± 0.029 | 0.189 ± 0.007 | 0.323 ± 0.008 | |
| MTX 4 | 0.475 ± 0.008 | 0.025 ± 0.010 | 0.052 ± 0.021 | 0.166 ± 0.000 | 0.336 ± 0.006 | |
| p-value2 | 0.087 | 0.002 | 0.002 | 0.046 | 0.199 | |
| %change5 | −7.60% | −92.34% | −57.21% | −12.50% | 4.27% | |
| (C) Thymidine synthesis from L-[3-13C]serine derived formate and exogenous [13C]formate 1,2. | ||||||
| Endogenous Formate from L-[3-13C]serine | Exogenous [13C]formate Incorporation | |||||
| Ser+1 3 | dT+1 3 | dT+1/Ser+1 7 | Ser+1 3 | dT+1 3 | ||
| CTL 4 | 0.515 ± 0.016 | 0.452 ± 0.003 | 0.879 ± 0.006 | 0.189 ± 0.007 | 0.384 ± 0.002 | |
| MTX 4 | 0.475 ± 0.008 | 0.198 ± 0.022 | 0.417 ± 0.047 | 0.166 ± 0.000 | 0.397 ± 0.001 | |
| t-Test 2 | 0.087 | 0.004 | 0.005 | 0.046 | 0.017 | |
| %change 5 | −7.60% | −56.14% | −46.14% | −12.50% | +3.42% | |
| (A) Relative incorporations of L-[3-13C]serine derived formate and exogenous [13C]formate in methionine 1,2. | |||||
| Endogenous Formate from L-[3-13C]serine | Exogenous [13C]formate Incorporation | ||||
| Ser+1 6 | Met+1 6 | Met+1/Ser+1 7 | Ser+1 6 | Met+1 6 | |
| CTL 4 | 0.515 ± 0.016 | 0.014 ± 0.003 | 0.027 ± 0.006 | 0.189 ± 0.007 | 0.012 ± 0.004 |
| MTX 4 | 0.475 ± 0.008 | 0.003 ± 0.003 | 0.006 ± 0.007 | 0.166 ± 0.000 | 0.024 ± 0.001 |
| t-Test | 0.087 | 0.074 | 0.083 | 0.046 | 0.038 |
| %change 5 | −7.60% | −78.90% | −2.1% | −12.50% | +110.00% |
| (B) Partitioning of 5,10 methyleneTHF between methionine and thymidylate syntheses using endogenous formate (from L-[3-13C]serine) and exogenous [13C]formate in vitro 1,2. | |||||
| L-[3-13C]serine | Ser+1 6 | dT+1 6 | Met+1 6 | dT+1/Met+1 8 | |
| CTL 4 | 0.515 ± 0.016 a | 0.452 ± 0.003 a | 0.014 ± 0.003 a | 32.568 ± 0.208 a | |
| MTX 4 | 0.475 ± 0.008b | 0.198 ± 0.022 b | 0.003 ± 0.003 b | 67.634 ± 7.554 b | |
| [13C]formate | Ser+1 6 | dT+1 6 | Met+1 6 | dT+1/Met+1 8 | |
| CTL 4 | 0.189 ± 0.007 a | 0.384 ± 0.002 a | 0.012 ± 0.004 a | 33.410 ± 0.139 a | |
| MTX 4 | 0.166 ± 0.000 b | 0.397 ± 0.001 b | 0.024 ± 0.001 b | 16.450 ± 0.024 b | |
| (C) Relative incorporation of the methylene group from L-[2,3,3-2H3]serine into methionine and thymidylate syntheses with MTX and folinate rescue in vitro 1,3. | |||||
| L-[2,3,3-2H3]serine | Via 5-methylTHF | Via 5,10 methyleneTHF | |||
| Ser+1+2+3 6 | Met+1+2 6 | Met from Ser 7 | dT+1+2 6 | dT from Ser 7 | |
| CTL 4 | 0.656 ± 0.001 a | 0.043 ± 0.000 a | 0.065 ± 0.000 a | 0.337 ± 0.003 a | 0.514 ± 0.006 a |
| FA 4 | 0.653 ± 0.005 a | 0.042 ± 0.007 ac | 0.064 ± 0.011 ac | 0.331 ± 0.001 a | 0.507 ± 0.005 ac |
| MTX 4 | 0.608 ± 0.041 a | 0.015 ± 0.000 bc | 0.024 ± 0.002 b | 0.262 ± 0.002 b | 0.432 ± 0.027 bc |
| MTX+FA Pre 4 | 0.647 ± 0.010 a | 0.019 ± 0.002 b | 0.029 ± 0.003 bc | 0.281 ± 0.002 c | 0.435 ± 0.004 b |
| MTX+FA CO 4 | 0.685 ± 0.037 a | 0.025 ± 0.005 b | 0.037 ± 0.010bc | 0.339 ± 0.005 a | 0.496 ± 0.034 ab |
| MTX+FA Post 4 | 0.653 ± 0.063 a | 0.016 ± 0.005 b | 0.025 ± 0.010 b | 0.277 ± 0.002 c | 0.426 ± 0.043ab |
| (D) Partitioning of 5,10 methyleneTHF 1C between methionine and thymidylate syntheses from L-[2,3,3-2H3]serine in vitro 1,3. | |||||
| From Mito | From Cyto | From Both | |||
| L-[2,3,3-2H3]serine | dT+1/Met+1 8 | dT+2/Met+2 8 | dT+1+2/Met+1+2 8 | ||
| CTL 4 | 10.152 ± 0.103 a | 5.253 ± 0.182 a | 7.890 ± 0.148 a | ||
| FA 4 | 10.331 ± 1.643 a | 5.208 ± 0.096 a | 8.013 ± 1.340 ac | ||
| MTX 4 | 17.220 ± 0.893 b | 16.032 ± 0.252 b | 18.082 ± 0.610 b | ||
| MTX+FA Pre 4 | 17.025 ± 2.420b | 12.669 ± 0.370 c | 14.956 ± 1.642 b | ||
| MTX+FA CO 4 | 16.768 ± 3.576 ab | 9.478 ± 0.161 d | 13.725 ± 2.733bc | ||
| MTX+FA Post 4 | 19.548 ± 7.623 ab | 13.162 ± 0.190 c | 18.099 ± 5.308 ab | ||
| Ser+1+2+3 2 | Ser+1 2 | Ser+2 2 | Ser+3 2 | Gly+1 2 | ||
|---|---|---|---|---|---|---|
| Marrow | CTL 3 | 0.270 ± 0.013 a | 0.075 ± 0.001 a | 0.095 ± 0.014 a | 0.099 ± 0.002 a | 0.054 ± 0.002 a |
| MTX 3 | 0.256 ± 0.010 a | 0.051 ± 0.003 b | 0.117 ± 0.004 a | 0.088 ± 0.003 b | 0.042 ± 0.002 b | |
| MTX+FA 3 | 0.267 ± 0.007 a | 0.066 ± 0.000 c | 0.111 ± 0.002 a | 0.090 ± 0.004b | 0.033 ± 0.001 c | |
| Liver | CTL 3 | 0.212 ± 0.012 a | 0.076 ± 0.002 a | 0.076 ± 0.002 a | 0.060 ± 0.008 a | 0.012 ± 0.002 a |
| MTX 3 | 0.235 ± 0.017 a | 0.041 ± 0.006 b | 0.102 ± 0.004 b | 0.092 ± 0.007 b | 0.007 ± 0.001b | |
| MTX+FA 3 | 0.254 ± 0.021 a | 0.071 ± 0.006 a | 0.092 ± 0.001 b | 0.091 ± 0.015 ab | 0.006 ± 0.001b |
| (A) Relative incorporations of L-[2,3,3-2H3]serine in dTMP and methionine 1. | ||||||
| via 5-methylTHF | via 5,10 methyleneTHF | |||||
| Met+1+2 2 | Met from Ser 4 | dT+1+2 2 | dT from Ser 4 | |||
| Marrow | CTL 3 | 0.034 ± 0.005 a | 0.125 ± 0.012 a | 0.118 ± 0.003 a | 0.439 ± 0.031 a | |
| MTX 3 | 0.032 ± 0.001 a | 0.125 ± 0.003 a | 0.110 ± 0.004 a | 0.428 ± 0.002 a | ||
| MTX+FA 3 | 0.034 ± 0.001 a | 0.128 ± 0.007 a | 0.126 ± 0.007 a | 0.472 ± 0.014 a | ||
| Liver | CTL 3 | 0.019 ± 0.002 a | 0.089 ± 0.013 a | 0.009 ± 0.001 a | 0.044 ± 0.002 a | |
| MTX 3 | 0.014 ± 0.000 b | 0.058 ± 0.003 b | 0.008 ± 0.003 a | 0.034 ± 0.011 a | ||
| MTX+FA 3 | 0.016 ± 0.001 ab | 0.063 ± 0.010 ab | 0.014 ± 0.004 a | 0.054 ± 0.012 a | ||
| (B) Utilization of L-[2,3,3-2H3]serine for dTMP and methionine in vivo 1. | ||||||
| via 5,10 methyleneTHF | via 5-methylTHF | |||||
| dT+1 (Mito) 2 | dT+2 (Cyto) 2 | dT% 5 | Met+1 (Mito) 2 | Met+2 (Cyto) 2 | met% 5 | |
| Marrow | ||||||
| CTL 3 | 0.110 ± 0.005 a | 0.008 ± 0.002 a | 92.8 ± 2.3 a | 0.025 ± 0.003 a | 0.009 ± 0.002 a | 73.8 ± 1.5 a |
| MTX 3 | 0.073 ± 0.001 b | 0.037 ± 0.003 b | 66.2 ± 1.6 b | 0.021 ± 0.001 a | 0.011 ± 0.000 a | 64.8 ± 1.9 b |
| MTX+FA 3 | 0.111 ± 0.008 a | 0.015 ± 0.001 c | 88.1 ± 1.4 a | 0.024 ± 0.001 a | 0.010 ± 0.000 a | 70.5 ± 0.7 a |
| Liver | ||||||
| CTL 3 | 0.008 ± 0.001 a | 0.001 ± 0.000 a | 87.9 ± 1.9 a | 0.012 ± 0.001 a | 0.007 ± 0.001 a | 63.4 ± 0.9 a |
| MTX 3 | 0.006 ± 0.002a* | 0.002 ± 0.002 a | 72.7 ± 8.4a* | 0.005 ± 0.000 b | 0.009 ± 0.000 b | 34.3 ± 2.8 b |
| MTX+FA 3 | 0.012 ± 0.003a* | 0.002 ± 0.001 a | 88.3 ± 6.4 a | 0.006 ± 0.003 b | 0.010 ± 0.001 b | 36.0 ± 13.4b |
| (C) Partitioning of 5,10 methyleneTHF in methionine and dTMP syntheses from L-[2,3,3-2H3]serine in vivo 1. | ||||||
| Mito | Cyto | Total | ||||
| dT+1/Met+1 6 | dT+2/Met+2 6 | dT+1+2/Met+1+2 6 | ||||
| Marrow | CTL 3 | 4.390 ± 0.201 a | 0.953 ± 0.278 a | 3.486 ± 0.075 a | ||
| MTX 3 | 3.507 ± 0.046 b | 3.301 ± 0.278 b | 3.435 ± 0.128 a | |||
| MTX+FA 3 | 4.601 ± 0.326 a | 1.489 ± 0.092 a | 3.684 ± 0.203 a | |||
| Liver | CTL 3 | 0.693 ± 0.051 a | 0.166 ± 0.042 a | 0.500 ± 0.048 a | ||
| MTX 3 | 1.239 ± 0.360 ab | 0.264 ± 0.176 a | 0.598 ± 0.239 a | |||
| MTX+FA 3 | 2.111 ± 0.487 b | 0.176 ± 0.137 a | 0.882 ± 0.265 a | |||
| (A) Nucleotide synthesis from [2-13C]glycine in vitro 1,2. | |||||||
| [2-13C]glycine | Ser+1 3 | Gly+1 3 | dA+1 3 | dG+1 3 | dT+1 3 | Ser+2 3 | |
| CTL 4 | 0.262 ±0.006 a | 0.329 ± 0.005 a | 0.176 ± 0.001 a | 0.220 ± 0.004 a | 0.011 ± 0.001 a | 0.008 ± 0.001 a | |
| FA 4 | 0.268 ± 0.006 a | 0.278 ± 0.034 a b | 0.175 ± 0.002 a | 0.212 ± 0.008 ad | 0.011 ± 0.000 a | 0.008 ± 0.002 a | |
| MTX 4 | 0.151 ± 0.001 b | 0.269 ± 0.009 b | 0.095 ± 0.001 b | 0.142 ± 0.000 b | 0.004 ± 0.002 b c | 0.000 ± 0.000 b | |
| MTX+FA Pre 4 | 0.164 ± 0.000 c | 0.160 ±0.032 c | 0.112 ± 0.002 c | 0.156 ± 0.003 c | 0.004 ± 0.000 c | 0.006 ± 0.006 a b c | |
| MTX+FA CO 4 | 0.222 ± 0.006d | 0.248 ± 0.070 a b c | 0.153 ± 0.001d | 0.201 ± 0.001d | 0.008 ± 0.002 a b | 0.003 ± 0.001 c | |
| MTX+FA Post 4 | 0.179 ± 0.000 e | 0.265 ± 0.010 b | 0.084 ± 0.002 e | 0.126 ± 0.001 e | 0.004 ± 0.003 b c | 0.000 ± 0.000 b | |
| (B) Utilization of glycine for serine synthesis in bone marrow and liver 1. | |||||||
| In Protein Lysates | In Cytosol Free Amino Acid | ||||||
| Ser+1 3 | Gly+1 3 | Ser+1 3 | Gly+1 3 | ||||
| Marrow | CTL 4 | 0.068 ± 0.001 a | 0.057 ± 0.001 a | 0.070 ± 0.004 a | 0.068 ± 0.002 a | ||
| MTX 4 | 0.075 ± 0.005 a b | 0.038 ± 0.006 b | 0.069 ± 0.003 a | 0.070 ± 0.003 a | |||
| MTX+FA 4 | 0.084 ± 0.002 b | 0.073 ± 0.000 c | 0.074 ± 0.003 a | 0.061 ± 0.003 b | |||
| Liver | CTL 4 | 0.125 ± 0.001 a | 0.020 ± 0.001 a | 0.097 ± 0.009 a | 0.111 ± 0.003 a | ||
| MTX 4 | 0.115 ± 0.004 b | 0.061 ± 0.042 a | 0.102 ± 0.004 a | 0.110 ± 0.014 a b | |||
| MTX+FA 4 | 0.125 ± 0.002 a b | 0.032 ± 0.035 a | 0.105 ± 0.003 a | 0.127 ± 0.002 b | |||
| (C) Nucleotide synthesis from [2-13C]glycine in bone marrow 1. | |||||||
| Ser+1 3 | Gly+1 3 | dT+1 3 | mC+1 3 | Ser+2 3 | dA+1 3 | dG+1 3 | |
| CTL 4 | 0.068 ± 0.001 a | 0.057 ± 0.001 a | 0.014 ± 0.000 a | 0.023 ± 0.002 a | 0.0013 ± 0.000 a | 0.112 ± 0.003 a | 0.113 ± 0.001 a |
| MTX 4 | 0.075 ± 0.005 a b | 0.038 ± 0.006 b | 0.012 ± 0.000 b | 0.013 ± 0.000 b | 0.0008 ± 0.001 a | 0.097 ± 0.003 b | 0.095 ± 0.002 b |
| MTX+FA 4 | 0.084 ± 0.002 b | 0.073 ± 0.000 c | 0.013 ± 0.001 a | 0.018 ± 0.002 a | 0.0019 ± 0.000 a | 0.107 ± 0.003 a | 0.105 ± 0.005 a b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sou, N.-L.; Huang, Y.-H.; Chen, D.-Y.; Chen, Y.-M.; Tang, F.-Y.; Ko, H.-A.; Fan, Y.-H.; Lin, Y.-Y.; Wang, Y.-C.; Chih, H.-M.; et al. Folinate Supplementation Ameliorates Methotrexate Induced Mitochondrial Formate Depletion In Vitro and In Vivo. Int. J. Mol. Sci. 2021, 22, 1350. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031350
Sou N-L, Huang Y-H, Chen D-Y, Chen Y-M, Tang F-Y, Ko H-A, Fan Y-H, Lin Y-Y, Wang Y-C, Chih H-M, et al. Folinate Supplementation Ameliorates Methotrexate Induced Mitochondrial Formate Depletion In Vitro and In Vivo. International Journal of Molecular Sciences. 2021; 22(3):1350. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031350
Chicago/Turabian StyleSou, Nga-Lai, Yu-Hsuan Huang, Der-Yuan Chen, Yi-Ming Chen, Feng-Yao Tang, Hsin-An Ko, Yi-Hsuan Fan, Yi-Ying Lin, Yi-Cheng Wang, Hui-Ming Chih, and et al. 2021. "Folinate Supplementation Ameliorates Methotrexate Induced Mitochondrial Formate Depletion In Vitro and In Vivo" International Journal of Molecular Sciences 22, no. 3: 1350. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031350