
A Novel Multiprotein Bridging Factor 1-Like Protein Induces Cyst Wall Protein Gene Expression and Cyst Differentiation in Giardia lamblia


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
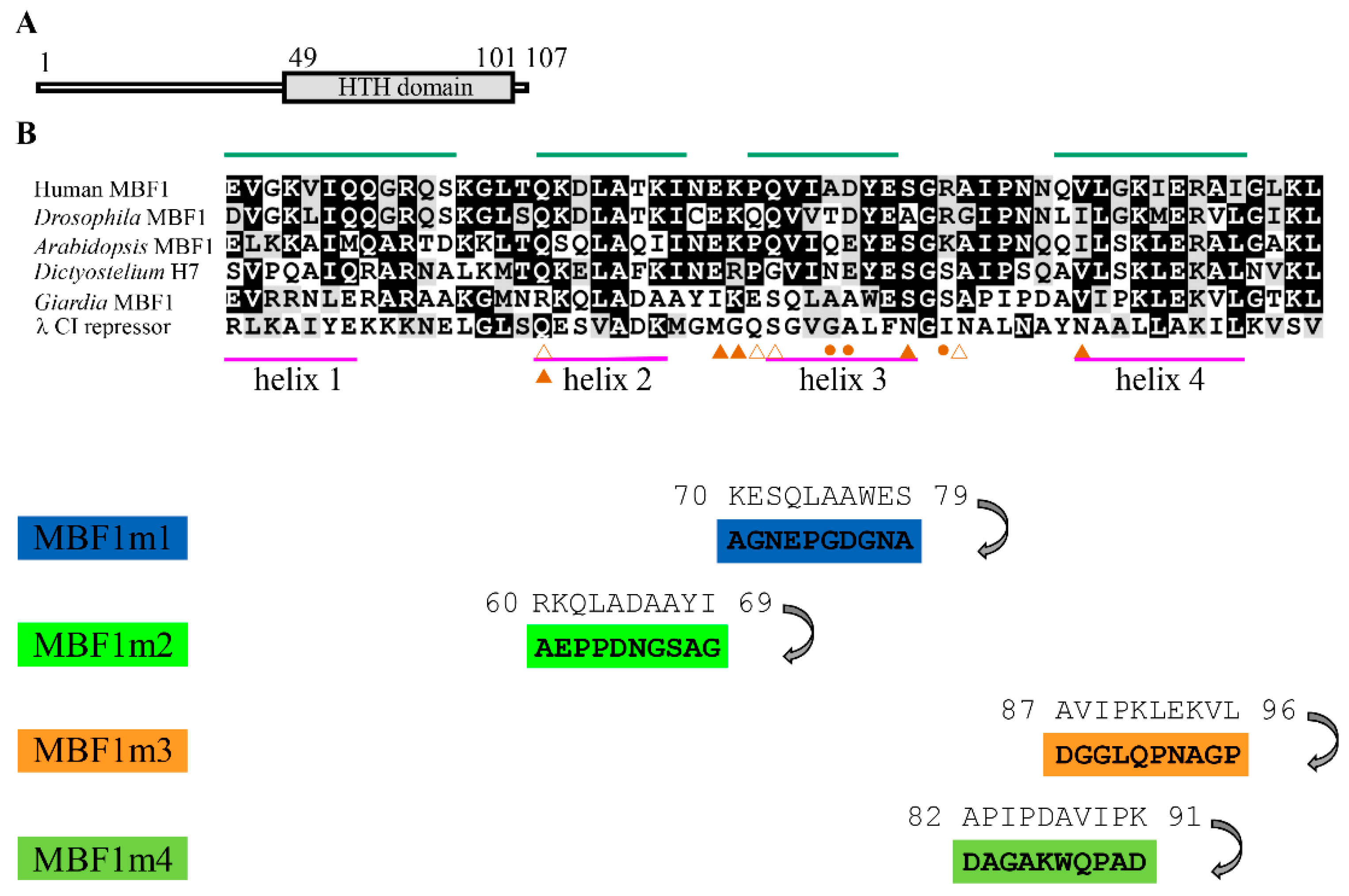
2.1. Analysis of the mbf1 Gene
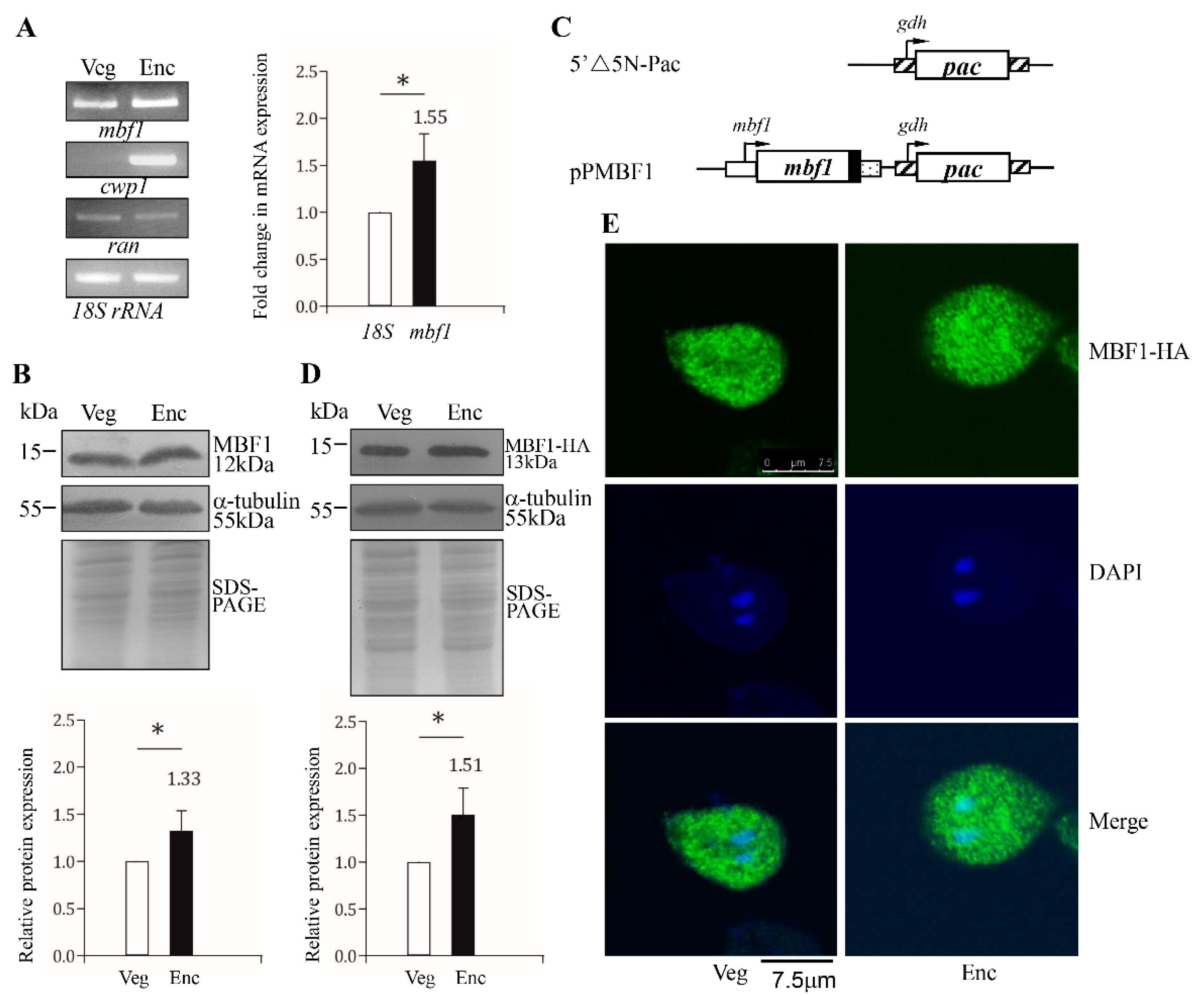
2.2. Encystation-Induced Expression and Localization of the MBF1 Protein
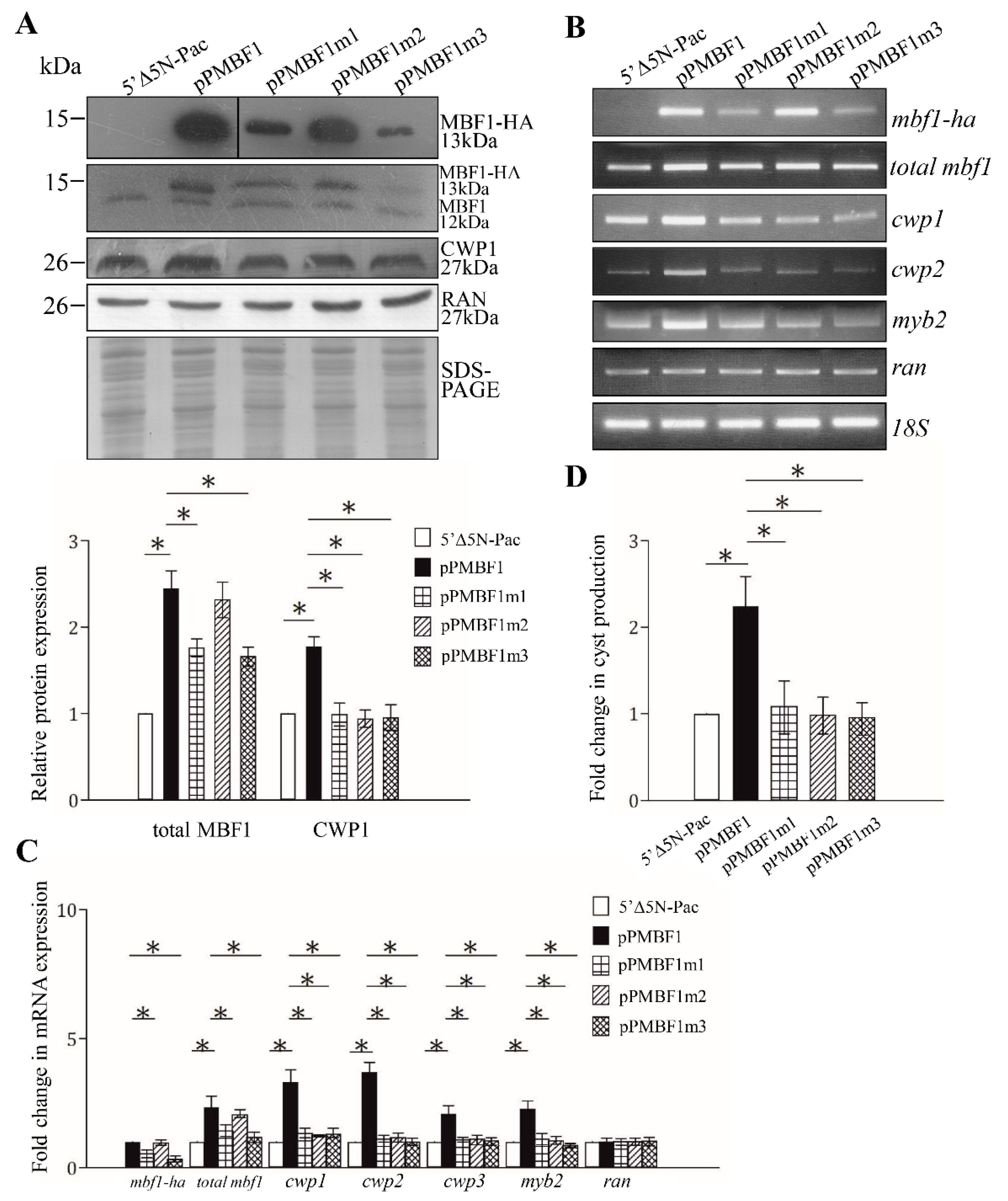
2.3. MBF1 Can Induce the Expression of the cwp1-3, and myb2 Genes
2.4. Effect of the MBF1 Mutants
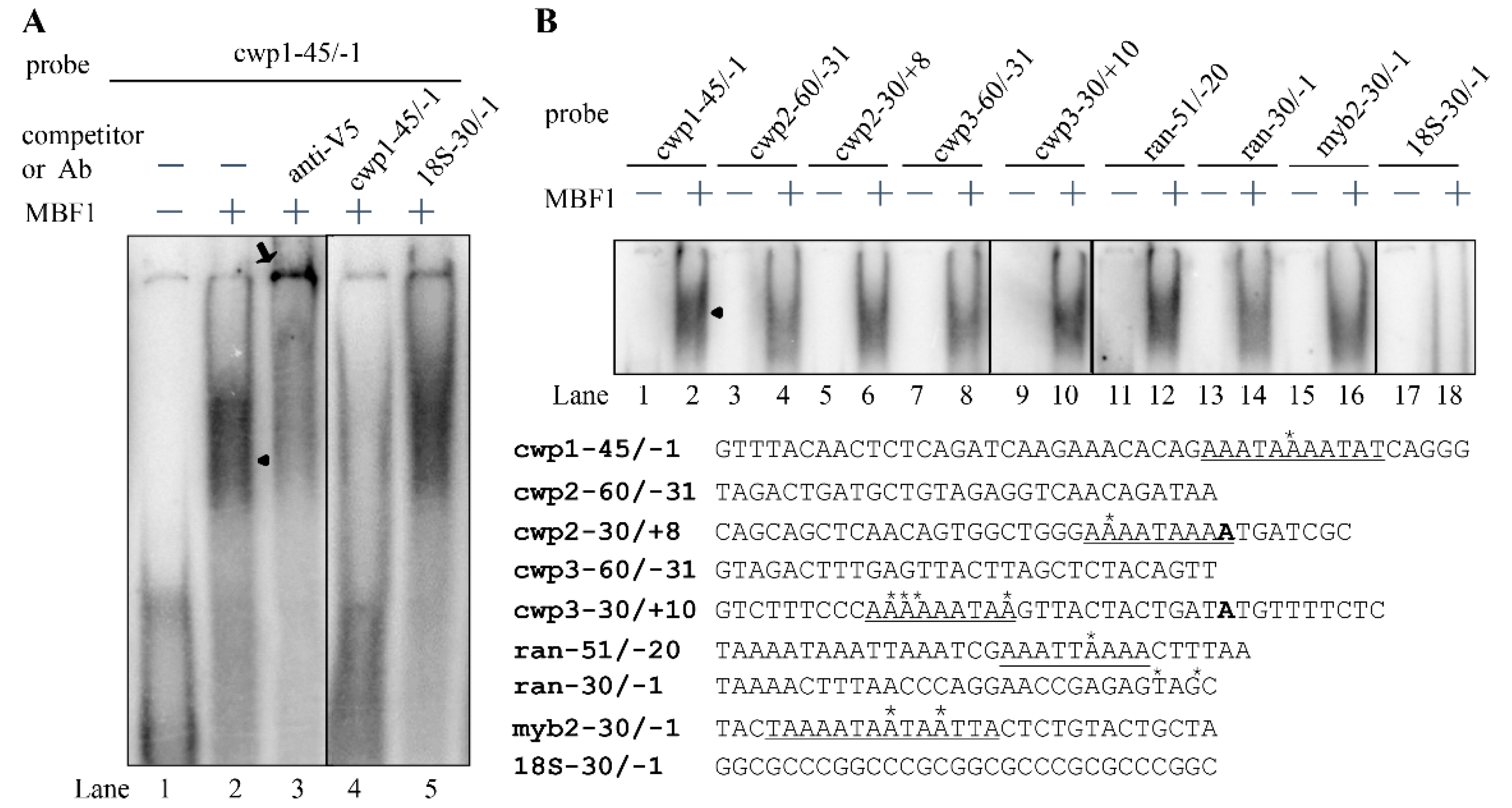
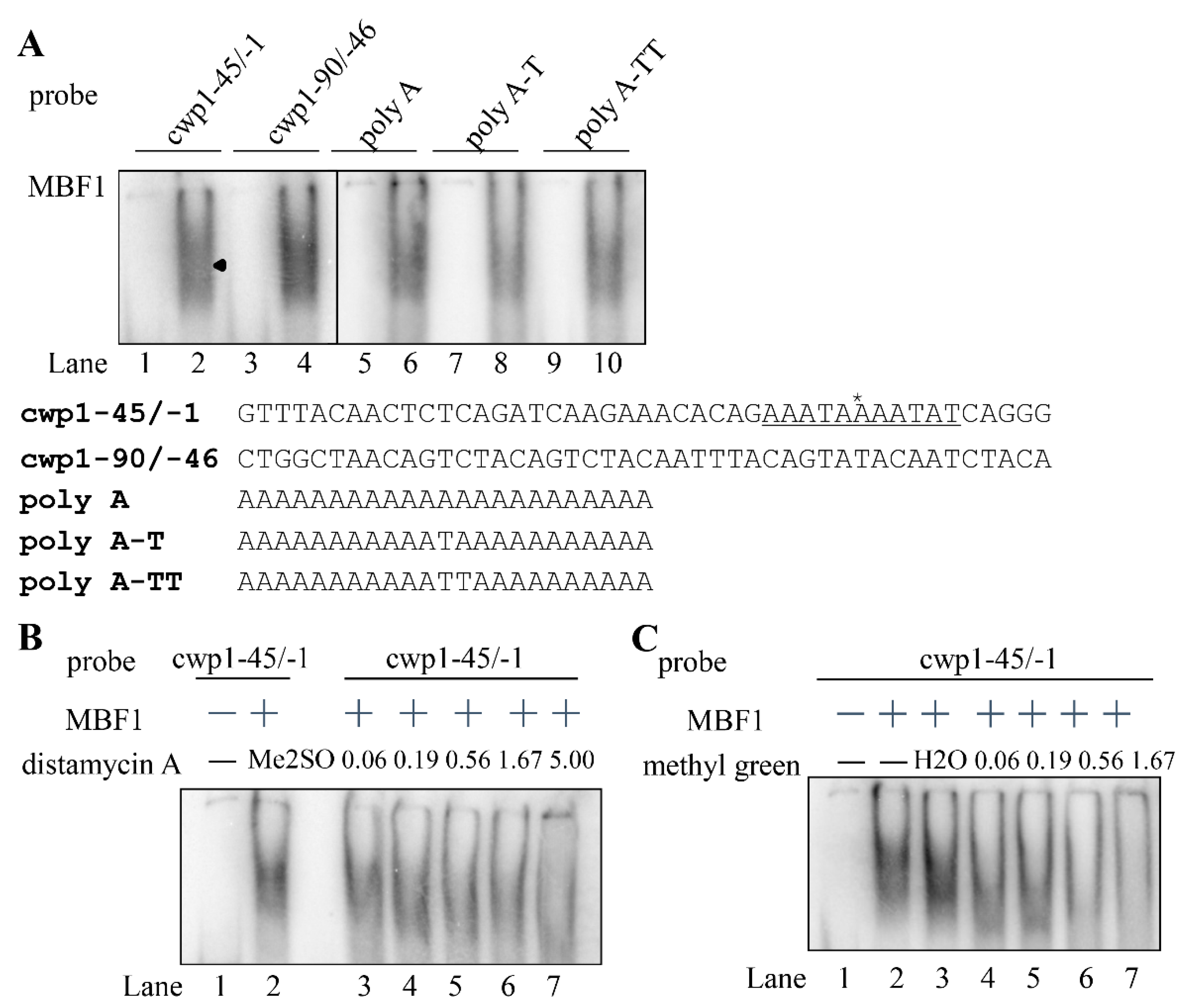
2.5. MBF1 Has DNA Binding Activity
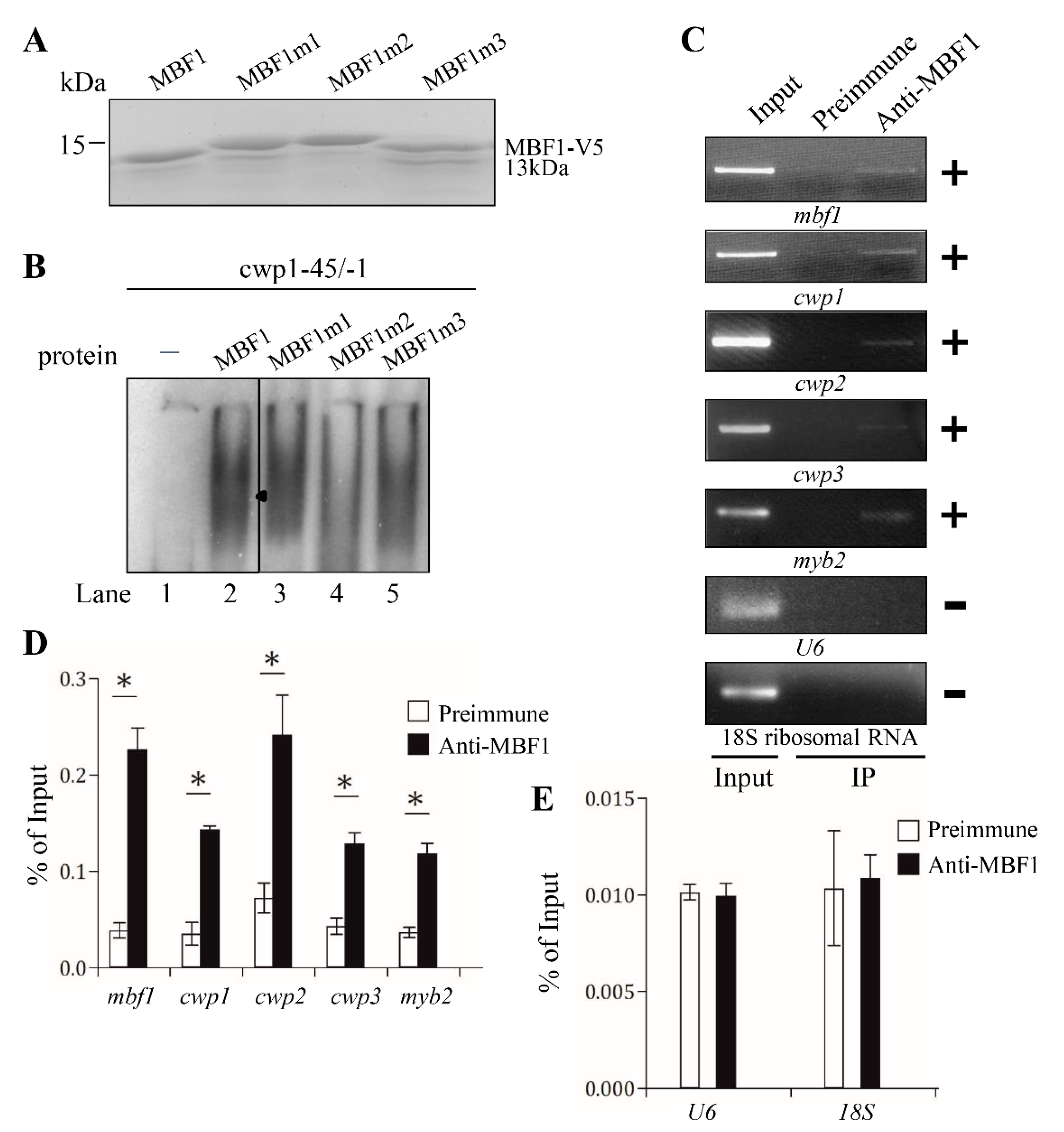
2.6. Analysis of DNA Binding Activity of MBF1 Mutants
2.7. In Vivo Association of MBF1 with the cwp1-3, myb2, and mbf1 Its Own Promoters
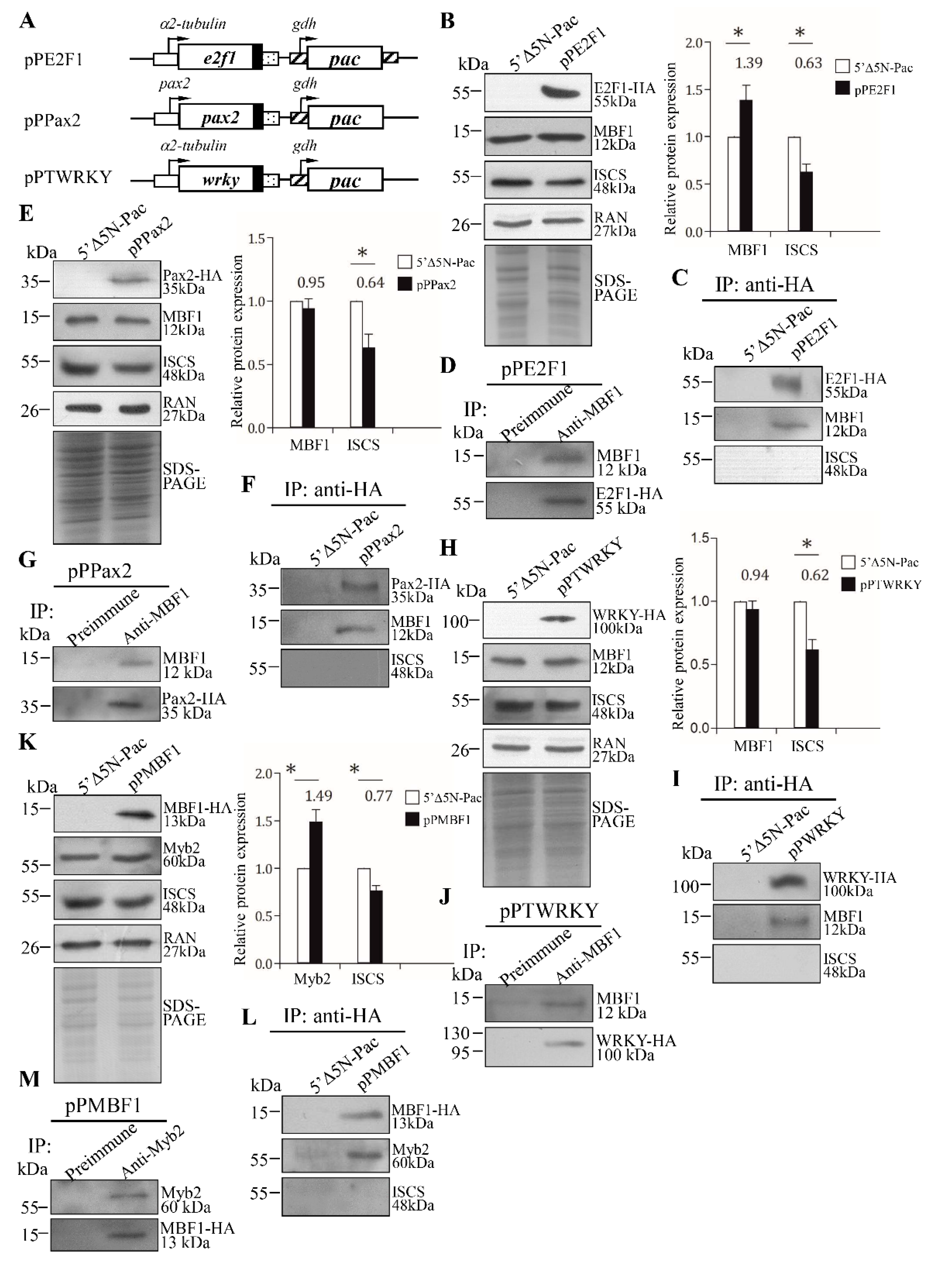
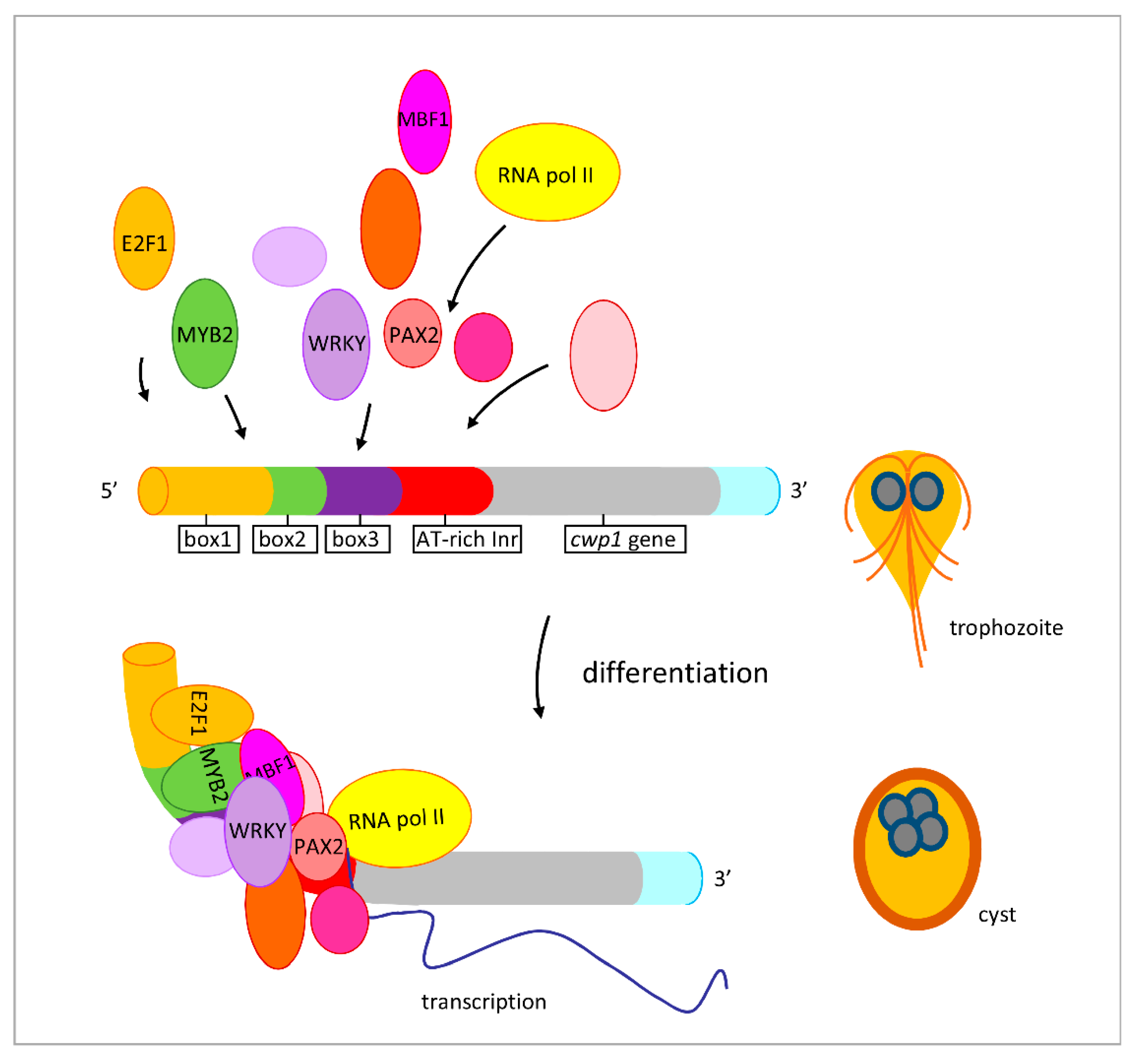
2.8. Interaction of MBF1 and E2F1, Pax2, or WRKY in a Complex
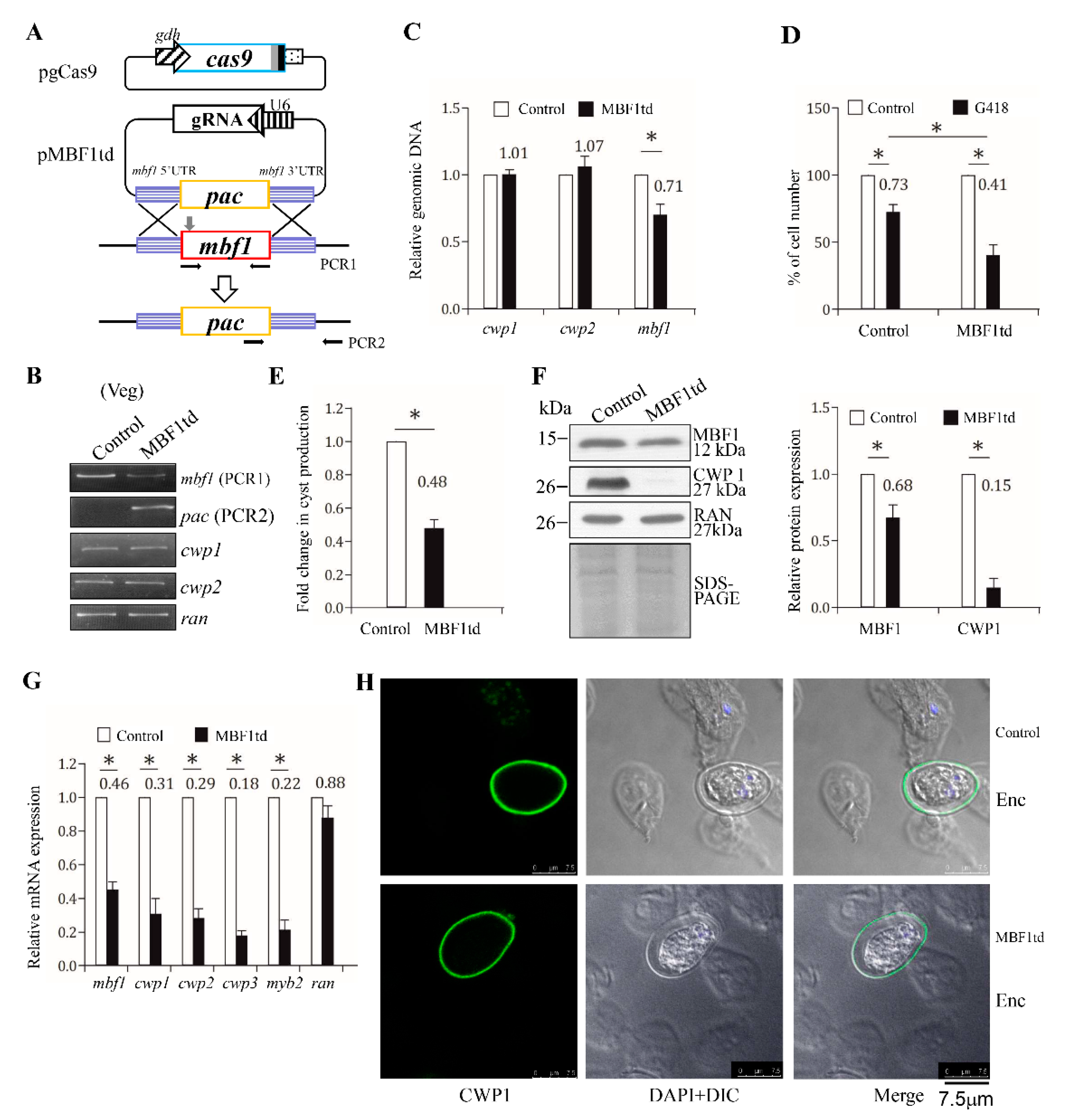
2.9. Reduced Expression of cwp1-3 and myb2 Genes by Targeted Disruption of the mbf1 Gene
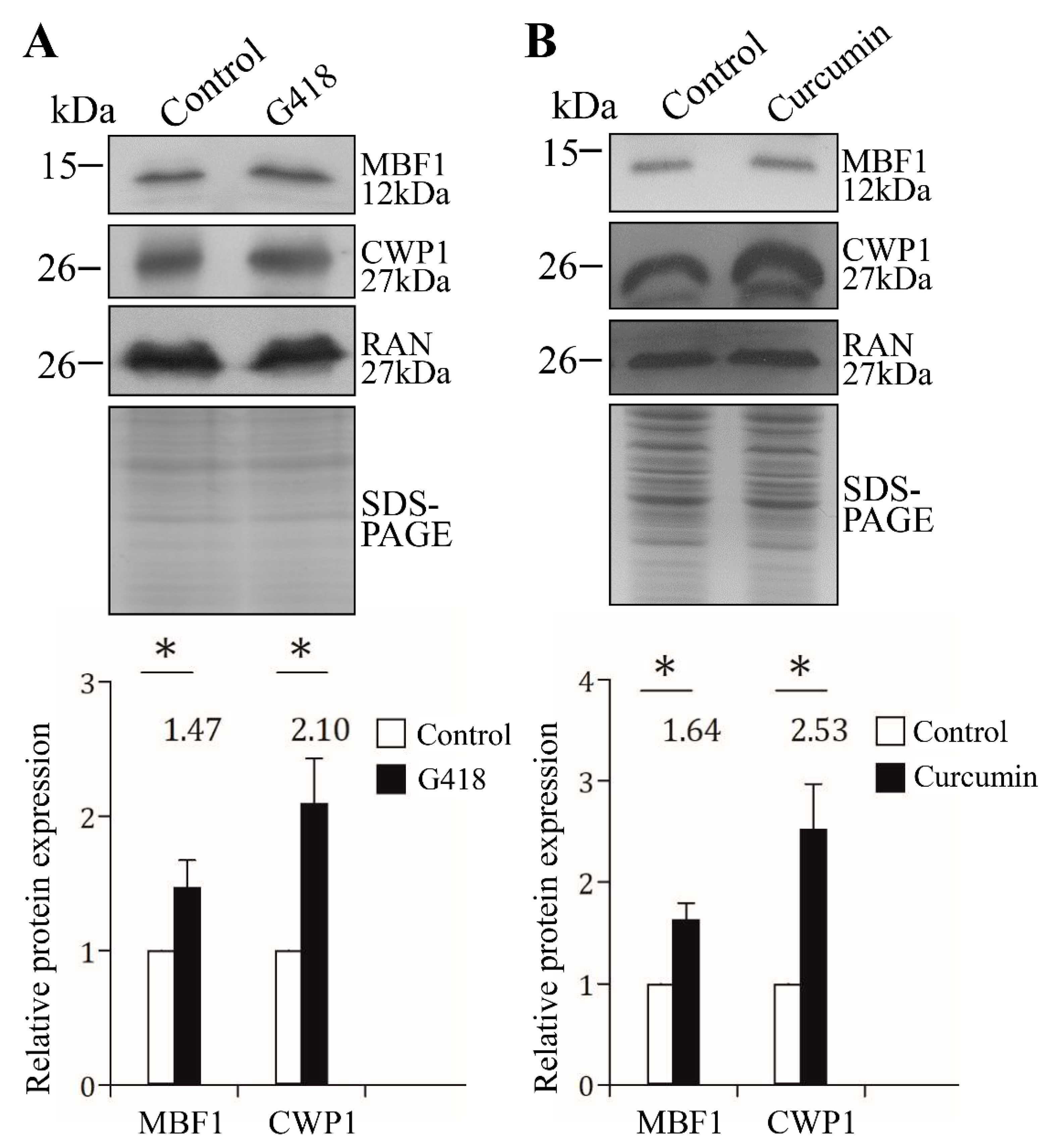
2.10. G418 and Curcumin Increased the Levels of MBF1 and CWP1 Proteins
3. Discussion
4. Materials and Methods
4.1. G. Lamblia Culture
4.2. Cyst Count
4.3. Isolation and Analysis of the mbf1 Gene
4.4. Genomic DNA Extraction, PCR and Quantitative Real-Time PCR Analysis
4.5. RNA Extraction, RT-PCR and Quantitative Real-Time PCR Analysis
4.6. Plasmid Construction
4.7. Transfection and Western Blot Analysis
4.8. Expression and Purification of Recombinant MBF1 Protein
4.9. Generation of Anti-MBF1 Antibody
4.10. Immunofluorescence Assay
4.11. Electrophoretic Mobility Shift Assay
4.12. Co-Immunoprecipitation Assay
4.13. ChIP Assays
4.14. Microarray Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adam, R.D. Biology of Giardia lamblia. Clin. Microbiol. Rev. 2001, 14, 447–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Einarsson, E.; Ma’ayeh, S.; Svärd, S.G. An update on Giardia and giardiasis. Curr. Opin. Microbiol. 2016, 34, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Ankarklev, J.; Jerlstrom-Hultqvist, J.; Ringqvist, E.; Troell, K.; Svard, S.G. Behind the smile: Cell biology and disease mechanisms of Giardia species. Nat. Rev. Microbiol. 2010, 8, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Hanevik, K.; Dizdar, V.; Langeland, N.; Hausken, T. Development of functional gastrointestinal disorders after Giardia lamblia infection. BMC Gastroenterol. 2009, 9, 27. [Google Scholar] [CrossRef] [Green Version]
- Muhsen, K.; Levine, M.M. A systematic review and meta-analysis of the association between Giardia lamblia and endemic pediatric diarrhea in developing countries. Clin. Infect. Dis. 2012, 55, S271–S293. [Google Scholar] [CrossRef] [PubMed]
- Bartelt, L.A.; Platts-Mills, J.A. Giardia: A pathogen or commensal for children in high-prevalence settings? Curr. Opin. Infect. Dis. 2016, 29, 502–507. [Google Scholar] [CrossRef]
- Halliez, M.C.; Buret, A.G. Extra-intestinal and long term consequences of Giardia duodenalis infections. World J. Gastroenterol. 2013, 19, 8974–8985. [Google Scholar] [CrossRef]
- Wensaas, K.A.; Langeland, N.; Hanevik, K.; Mørch, K.; Eide, G.E.; Rortveit, G. Irritable bowel syndrome and chronic fatigue 3 years after acute giardiasis: Historic cohort study. Gut 2012, 61, 214–219. [Google Scholar] [CrossRef]
- Hanevik, K.; Wensaas, K.A.; Rortveit, G.; Eide, G.E.; Mørch, K.; Langeland, N. Irritable bowel syndrome and chronic fatigue 6 years after giardia infection: A controlled prospective cohort study. Clin. Infect. Dis. 2014, 59, 1394–1400. [Google Scholar] [CrossRef]
- Morrison, H.G.; McArthur, A.G.; Gillin, F.D.; Aley, S.B.; Adam, R.D.; Olsen, G.J.; Best, A.A.; Cande, W.Z.; Chen, F.; Cipriano, M.J. Genomic minimalism in the early diverging intestinal parasite Giardia lamblia. Science 2017, 317, 1921–1926. [Google Scholar] [CrossRef] [Green Version]
- Knodler, L.A.; Svard, S.G.; Silberman, J.D.; Davids, B.J.; Gillin, F.D. Developmental gene regulation in Giardia lamblia: First evidence for an encystation-specific promoter and differential 5’ mRNA processing. Mol. Microbiol. 1999, 34, 327–340. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Díaz, H.; Carrero, J.C.; Argüello-García, R.; Laclette, J.P.; Morales-Montor, J. Cyst and encystment in protozoan parasites: Optimal targets for new life-cycle interrupting strategies? Trends Parasitol. 2011, 27, 450–458. [Google Scholar] [CrossRef] [PubMed]
- Faso, C.; Hehl, A.B. Membrane trafficking and organelle biogenesis in Giardia lamblia: Use it or lose it. Int. J. Parasitol. 2011, 41, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Lujan, H.D.; Mowatt, M.R.; Conrad, J.T.; Bowers, B.; Nash, T.E. Identification of a novel Giardia lamblia cyst wall protein with leucine-rich repeats. Implications for secretory granule formation and protein assembly into the cyst wall. J. Biol. Chem. 1995, 270, 29307–29313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mowatt, M.R.; Lujan, H.D.; Cotten, D.B.; Bowers, B.; Yee, J.; Nash, T.E.; Stibbs, H.H. Developmentally regulated expression of a Giardia lamblia cyst wall protein gene. Mol. Microb. 1995, 15, 955–963. [Google Scholar] [CrossRef]
- Sun, C.H.; McCaffery, J.M.; Reiner, D.S.; Gillin, F.D. Mining the Giardia lamblia genome for new cyst wall proteins. J. Biol. Chem. 2003, 278, 21701–21708. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.H.; Palm, D.; McArthur, A.G.; Svard, S.G.; Gillin, F.D. A Novel Myb-Related Protein Involved in Transcriptional Activation of Encystation Genes in Giardia lamblia. Mol. Microbiol. 2002, 46, 971–984. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.H.; Su, L.H.; Gillin, F.D. Novel plant-GARP-like transcription factors in Giardia lamblia. Mol. Biochem. Parasitol. 2006, 146, 45–57. [Google Scholar] [CrossRef]
- Wang, C.H.; Su, L.H.; Sun, C.H. A novel ARID/Bright-like protein involved in transcriptional activation of cyst wall protein 1 gene in Giardia lamblia. J. Biol. Chem. 2007, 282, 8905–8914. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.-C.; Su, L.-H.; Lee, G.A.; Chiu, P.-W.; Cho, C.-C.; Wu, J.-Y.; Sun, C.-H. Regulation of cyst wall protein promoters by Myb2 in Giardia lamblia. J. Biol. Chem. 2008, 283, 31021–31029. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.J.; Cho, C.C.; Kao, Y.Y.; Sun, C.H. A novel WRKY-like protein involved in transcriptional activation of cyst wall protein genes in Giardia lamblia. J. Biol. Chem. 2009, 284, 17975–17988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.T.; Pan, Y.J.; Cho, C.C.; Lin, B.C.; Su, L.H.; Huang, Y.-C.; Sun, C.-H. A novel pax-like protein involved in transcriptional activation of cyst wall protein genes in Giardia lamblia. J. Biol. Chem. 2010, 285, 32213–32226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, L.-H.; Pan, Y.-J.; Huang, Y.-C.; Cho, C.-C.; Chen, C.-W.; Huang, S.-W.; Chuang, S.-F.; Sun, C.-H. A Novel E2F-like Protein Involved in Transcriptional Activation of Cyst Wall Protein Genes in Giardia lamblia. J. Biol. Chem. 2011, 286, 34101–34120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuang, S.F.; Su, L.H.; Cho, C.C.; Pan, Y.J.; Sun, C.H. Functional Redundancy of Two Pax-like Proteins in Transcriptional Activation of Cyst Wall Protein Genes in Giardia lamblia. PLoS ONE 2012, 7, e30614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brennan, R.G.; Matthews, B.W. The helix-turn-helix DNA binding motif. J. Biol. Chem. 1989, 264, 1903–1906. [Google Scholar] [CrossRef]
- Aravind, L.; Anantharaman, V.; Balaji, S.; Babu, M.M.; Iyer, L.M. The many faces of the helix-turn-helix domain: Transcription regulation and beyond. FEMS Microbiol. Rev. 2005, 29, 231–262. [Google Scholar] [CrossRef]
- Marrero Coto, J.; Ehrenhofer-Murray, A.E.; Pons, T.; Siebers, B. Functional analysis of archaeal MBF1 by complementation studies in yeast. Biol. Direct. 2011, 6, 18. [Google Scholar] [CrossRef] [Green Version]
- Harrison, S.C.; Aggarwal, A.K. DNA recognition by proteins with the helix-turn-helix motif. Annu. Rev. Biochem. 1990, 59, 933–969. [Google Scholar] [CrossRef]
- Kabe, Y.; Goto, M.; Shima, D.; Imai, T.; Wada, T.; Morohashi, K.; Shirakawa, M.; Hirose, S.; Handa, H. The role of human MBF1 as a transcriptional coactivator. J. Biol. Chem. 1999, 274, 34196–34202. [Google Scholar] [CrossRef] [Green Version]
- Brendel, C.; Gelman, L.; Auwerx, J. Multiprotein bridging factor-1 (MBF-1) is a cofactor for nuclear receptors that regulate lipid metabolism. Mol. Endocrinol. 2002, 16, 1367–1377. [Google Scholar] [CrossRef]
- De Koning, B.; Blombach, F.; Wu, H.; Brouns, S.J.; van der Oost, J. Role of multiprotein bridging factor 1 in archaea: Bridging the domains? Biochem. Soc. Trans. 2009, 37, 52–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takemaru, K.; Harashima, S.; Ueda, H.; Hirose, S. Yeast coactivator MBF1 mediates GCN4-dependent transcriptional activation. Mol. Cell. Biol. 1998, 18, 4971–4976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dragoni, I.; Mariotti, M.; Consalez, G.G.; Soria, M.R.; Maier, J.A. EDF-1, a novel gene product down-regulated in human endothelial cell differentiation. J. Biol. Chem. 1998, 273, 31119–31124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.Q.; Takemaru, K.; Goto, M.; Ueda, H.; Handa, H.; Hirose, S. Transcriptional activation through interaction of MBF2 with TFIIA. Genes Cells 1997, 2, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Li, F.Q.; Ueda, H.; Hirose, S. Mediators of activation of fushi tarazu gene transcription by BmFTZ-F1. Mol. Cell. Biol. 1994, 14, 3013–3021. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.P.; Chen, R.Y.; Zhang, Q.X.; Peng, H.; Wang, K.J. Differential gene expression profile from haematopoietic tissue stem cells of red claw crayfish, Cherax quadricarinatus, in response to WSSV infection. Dev. Comp. Immunol. 2011, 35, 716–724. [Google Scholar] [CrossRef]
- Sugikawa, Y.; Ebihara, S.; Tsuda, K.; Niwa, Y.; Yamazaki, K. Transcriptional coactivator MBF1s from Arabidopsis predominantly localize in nucleolus. J. Plant Res. 2005, 118, 431–437. [Google Scholar] [CrossRef]
- Jindra, M.; Gaziova, I.; Uhlirova, M.; Okabe, M.; Hiromi, Y.; Hirose, S. Coactivator MBF1 preserves the redox-dependent AP-1 activity during oxidative stress in Drosophila. EMBO J. 2004, 23, 3538–3547. [Google Scholar] [CrossRef] [Green Version]
- Mauro, M.F.D.; Iglesias, M.J.; Arce, D.P.; Valle, E.M.; Arnold, R.B.; Tsuda, K.; Yamazaki, K.; Casalongué, C.A.; Godoy, A.V. MBF1s regulate ABA-dependent germination of Arabidopsis seeds. Plant Signal. Behav. 2012, 7, 188–192. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wei, X.; Huang, J.; Wei, J. Modification and functional adaptation of the MBF1 gene family in the lichenized fungus Endocarpon pusillum under environmental stress. Sci. Rep. 2017, 7, 16333. [Google Scholar] [CrossRef] [Green Version]
- Blombach, F.; Launay, H.; Snijders, A.P.; Zorraquino, V.; Wu, H.; de Koning, B.; Brouns, S.J.; Ettema, T.J.; Camiloni, C.; Cavalli, A.; et al. Archaeal MBF1 binds to 30S and 70S ribosomes via its helix-turn-helix domain. Biochem. J. 2014, 462, 373–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Zhou, J.; Yang, Q.; Grayhack, E.J. Multi-protein bridging factor 1(Mbf1), Rps3 and Asc1 prevent stalled ribosomes from frameshifting. eLife 2018, 7, e39637. [Google Scholar] [CrossRef] [PubMed]
- Millership, J.J.; Waghela, P.; Cai, X.; Cockerham, A.; Zhu, G. Differential expression and interaction of transcription co-activator MBF1 with TATA-binding protein (TBP) in the apicomplexan Cryptosporidium parvum. Microbiology 2004, 150, 1207–1213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Z.Q.; Gan, S.W.; Tung, S.Y.; Ho, C.C.; Su, L.H.; Sun, C.H. Development of CRISPR/Cas9-mediated gene disruption systems in Giardia lamblia. PLoS ONE 2019, 14, e0213594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finn, R.D.; Tate, J.; Mistry, J.; Coggill, P.C.; Sammut, J.S.; Hotz, H.R.; Ceric, G.; Forslund, K.; Eddy, S.R.; Sonnhammer, E.L.; et al. The Pfam protein families database. Nucleic Acids Res. 2008, 36, D281–D288. [Google Scholar] [CrossRef] [Green Version]
- Pabo, C.O.; Lewis, M. The operator-binding domain of lambda repressor: Structure and DNA recognition. Nature 1982, 29, 8443–8447. [Google Scholar]
- Buchan, D.W.A.; Minneci, F.; Nugent, T.C.O.; Bryson, K.; Jones, D.T. Scalable web services for the PSIPRED Protein Analysis Workbench. Nucleic Acids Res. 2013, 41, W340–W348. [Google Scholar] [CrossRef]
- Jordan, S.R.; Pabo, C.O. Structure of the lambda complex at 2.5 A resolution: Details of the repressor-operator interactions. Science 1988, 242, 893–899. [Google Scholar] [CrossRef]
- Singer, S.M.; Yee, J.; Nash, T.E. Episomal and integrated maintenance of foreign DNA in Giardia lamblia. Mol. Biochem. Parasitol. 1988, 92, 59–69. [Google Scholar] [CrossRef]
- Suzuki, N.; Sejima, H.; Tam, R.; Schlauch, K.; Mittler, R. Identification of the MBF1 heat-response regulon of Arabidopsis thaliana. Plant J. 2011, 66, 844–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, C.H.; Tai, J.H. Identification and characterization of a ran gene promoter in the protozoan pathogen Giardia lamblia. J. Biol. Chem. 1999, 274, 19699–19706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeda, Y.; Kim, J.G.; Caday, C.G.; Steers, E., Jr.; Ohlendorf, D.H.; Anderson, W.F.; Matthews, B.W. Different interactions used by Cro repressor in specific and nonspecific DNA binding. J. Biol. Chem. 1986, 261, 8608–8616. [Google Scholar] [CrossRef]
- Albright, R.A.; Matthews, B.W. How Cro and lambda-repressor distinguish between operators: The structural basis underlying a genetic switch. Proc. Natl. Acad. Sci. USA 1998, 95, 3431–3436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dickinson, L.A.; Joh, T.; Kohwi, Y.; Kohwi-Shigematsu, T. A tissue-specific MAR/SAR DNA-binding protein with unusual binding site recognition. Cell 1992, 70, 631–645. [Google Scholar] [CrossRef]
- Kim, S.K.; Nordén, B. Methyl green. A DNA major-groove binding drug. FEBS Lett. 1993, 315, 61–64. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.H.; Weng, S.C.; Wu, J.H.; Tung, S.Y.; Su, L.H.; Lin, M.H.; Lee, G.A. DNA topoisomerase IIIbeta promotes cyst generation by inducing cyst wall protein gene expression in Giardia lamblia. Open Biol. 2020, 10, 190228. [Google Scholar] [CrossRef] [Green Version]
- Gao, K.; Chi, Y.; Zhang, X.; Zhang, H.; Li, G.; Sun, W.; Takeda, M.; Yao, J. A novel TXNIP-based mechanism for Cx43-mediated regulation of oxidative drug injury. J. Cell Mol. Med. 2015, 19, 2469–2480. [Google Scholar] [CrossRef]
- Kocyigit, A.; Guler, E.M. Curcumin induce DNA damage and apoptosis through generation of reactive oxygen species and reducing mitochondrial membrane potential in melanoma cancer cells. Cell. Mol. Biol. 2017, 63, 97–105. [Google Scholar] [CrossRef]
- Larasati, Y.A.; Yoneda-Kato, N.; Nakamae, I.; Yokoyama, T.; Meiyanto, E.; Kato, J.Y. Curcumin targets multiple enzymes involved in the ROS metabolic pathway to suppress tumor cell growth. Sci. Rep. 2018, 8, 2039. [Google Scholar] [CrossRef]
- Su, L.H.; Lee, G.A.; Huang, Y.C.; Chen, Y.H.; Sun, C.H. Neomycin and puromycin affect gene expression in Giardia lamblia stable transfection. Mol. Biochem. Parasitol. 2007, 156, 124–135. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Gutiérrez, F.; Palomo-Ligas, L.; Hernández-Hernández, J.M.; Pérez-Rangel, A.; Aguayo-Ortiz, R.; Hernández-Campos, A.; Castillo, R.; González-Pozos, S.; Cortés-Zárate, R.; Ramírez-Herrera, M.A.; et al. Curcumin alters the cytoskeleton and microtubule organization on trophozoites of Giardia lamblia. Acta Trop. 2017, 72, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Yee, J.; Mowatt, M.R.; Dennis, P.P.; Nash, T.E. Transcriptional analysis of the glutamate dehydrogenase gene in the primitive eukaryote, Giardia lamblia. Identification of a primordial gene promoter. J. Biol. Chem. 2000, 275, 11432–11439. [Google Scholar] [PubMed] [Green Version]
- Elmendorf, H.G.; Singer, S.M.; Pierce, J.; Cowan, J.; Nash, T.E. Initiator and upstream elements in the alpha2-tubulin promoter of Giardia lamblia. Mol. Biochem. Parasitol. 2001, 113, 157–169. [Google Scholar] [CrossRef]
- Narayanan, A.; Paul, L.N.; Tomar, S.; Patil, D.N.; Kumar, P.; Yernool, D.A. Structure-function studies of DNA binding domain of response regulator KdpE reveals equal affinity interactions at DNA half-sites. PLoS ONE 2012, 7, e30102. [Google Scholar] [CrossRef] [Green Version]
- Nakai, K.; Kanehisa, M. A knowledge base for predicting protein localization sites in eukaryotic cells. Genomics 1992, 14, 897–911. [Google Scholar] [CrossRef]
- Görlich, D.; Kutay, U. Transport between the cell nucleus and the cytoplasm. Annu. Rev. Cell. Dev. Biol. 1999, 15, 607–660. [Google Scholar] [CrossRef]
- Keister, D.B. Axenic culture of Giardia lamblia in TYI-S-33 medium supplemented with bile. Trans. R. Soc. Trop. Med. Hyg. 1983, 77, 487–488. [Google Scholar] [CrossRef]
- Gillin, F.D.; Boucher, S.E.; Rossi, S.S.; Reiner, D.S. Giardia lamblia: The roles of bile, lactic acid, and pH in the completion of the life cycle in vitro. Exp. Parasitol. 1996, 69, 164–174. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2001. [Google Scholar]
- Chen, Y.H.; Su, L.H.; Sun, C.H. Incomplete nonsense-mediated mRNA decay in Giardia lamblia. Int. J. Parasitol. 2008, 38, 1305–1317. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, S.-W.; Lin, Z.-Q.; Tung, S.-Y.; Su, L.-H.; Ho, C.-C.; Lee, G.A.; Sun, C.-H. A Novel Multiprotein Bridging Factor 1-Like Protein Induces Cyst Wall Protein Gene Expression and Cyst Differentiation in Giardia lamblia. Int. J. Mol. Sci. 2021, 22, 1370. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031370
Huang S-W, Lin Z-Q, Tung S-Y, Su L-H, Ho C-C, Lee GA, Sun C-H. A Novel Multiprotein Bridging Factor 1-Like Protein Induces Cyst Wall Protein Gene Expression and Cyst Differentiation in Giardia lamblia. International Journal of Molecular Sciences. 2021; 22(3):1370. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031370
Chicago/Turabian StyleHuang, Shao-Wei, Zi-Qi Lin, Szu-Yu Tung, Li-Hsin Su, Chun-Che Ho, Gilbert Aaron Lee, and Chin-Hung Sun. 2021. "A Novel Multiprotein Bridging Factor 1-Like Protein Induces Cyst Wall Protein Gene Expression and Cyst Differentiation in Giardia lamblia" International Journal of Molecular Sciences 22, no. 3: 1370. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031370