
IGF-1 Upregulates Biglycan and Decorin by Increasing Translation and Reducing ADAMTS5 Expression

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
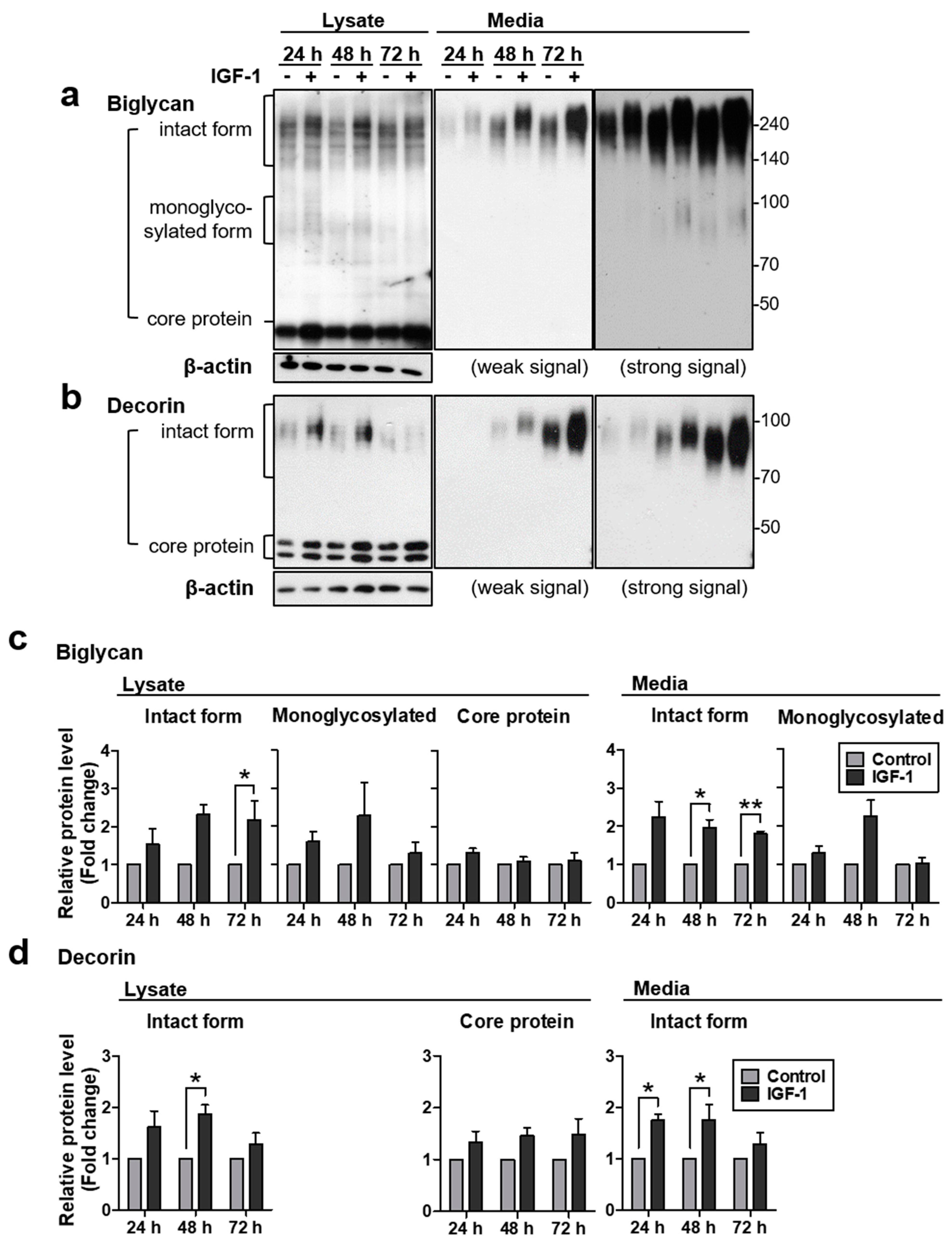
2.1. IGF-1 Increases Protein Levels of Biglycan and Decorin, but Not mRNA Expression in NHDFs
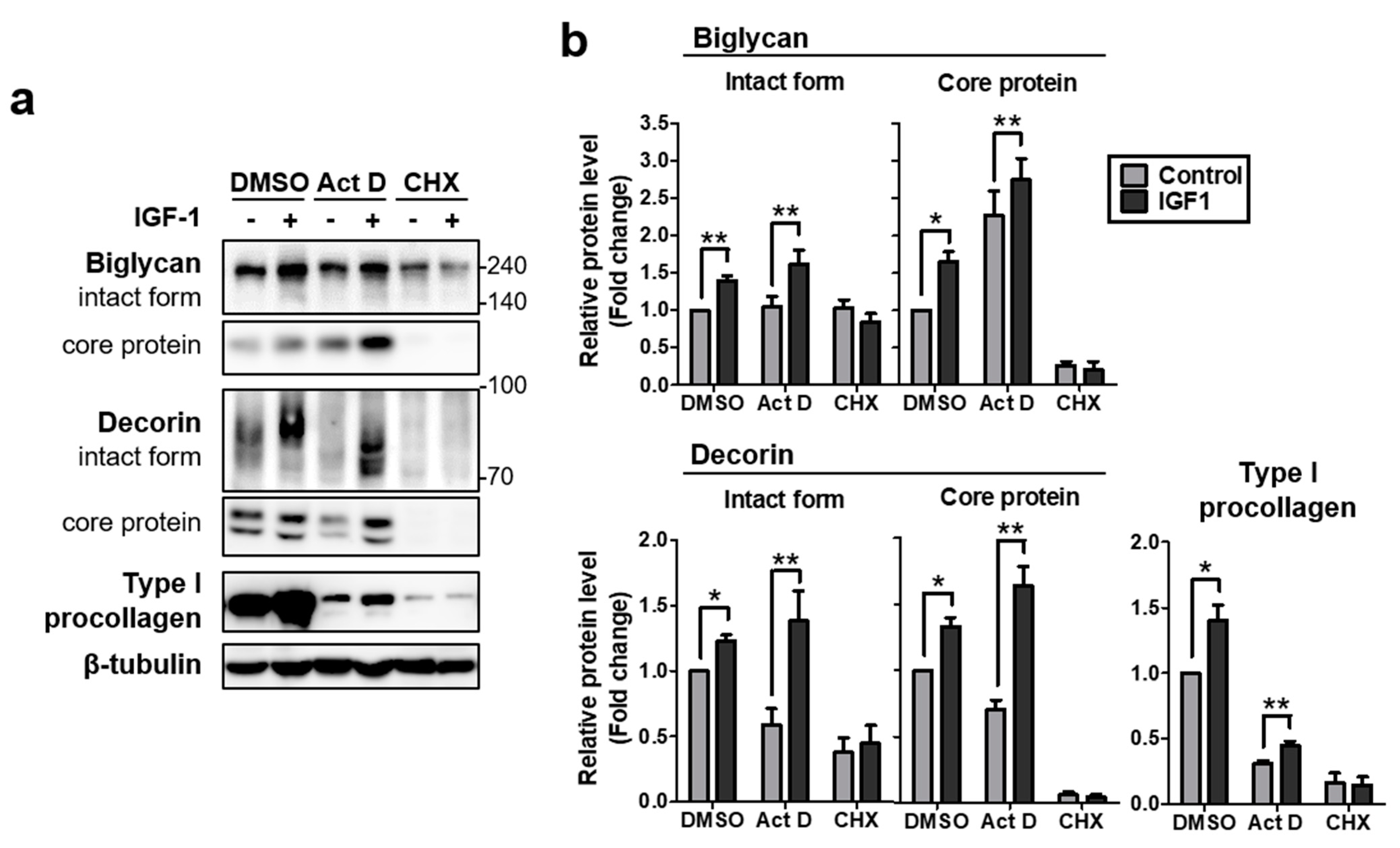
2.2. IGF-1 Increases Protein Levels of Biglycan and Decorin by Upregulating Protein Translation in NHDFs
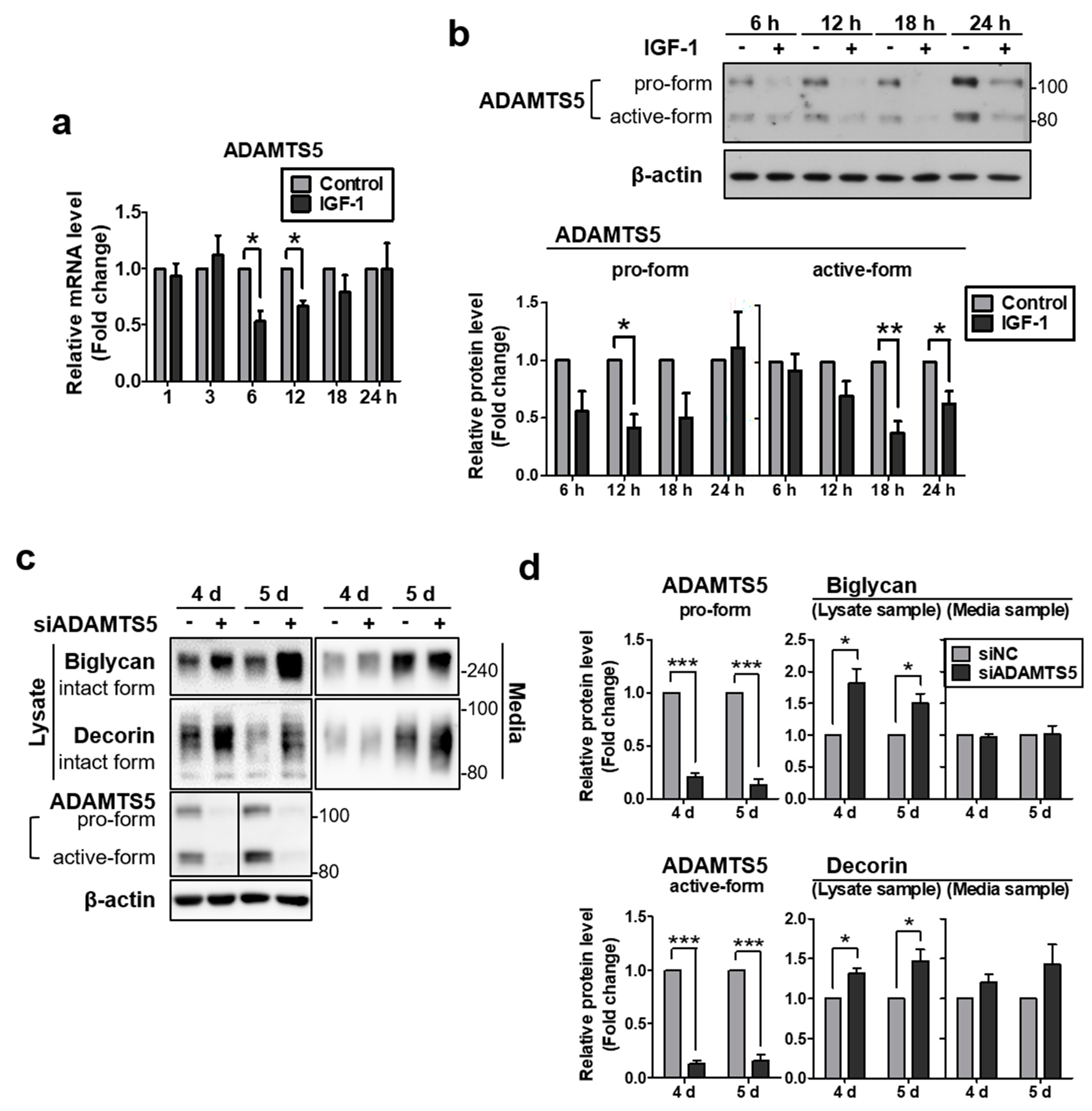
2.3. IGF-1 Downregulates the Expression of ADAMTS5, and Knockdown of ADAMTS5 Resulted in Augmented Biglycan and Decorin Protein Levels in NHDFs
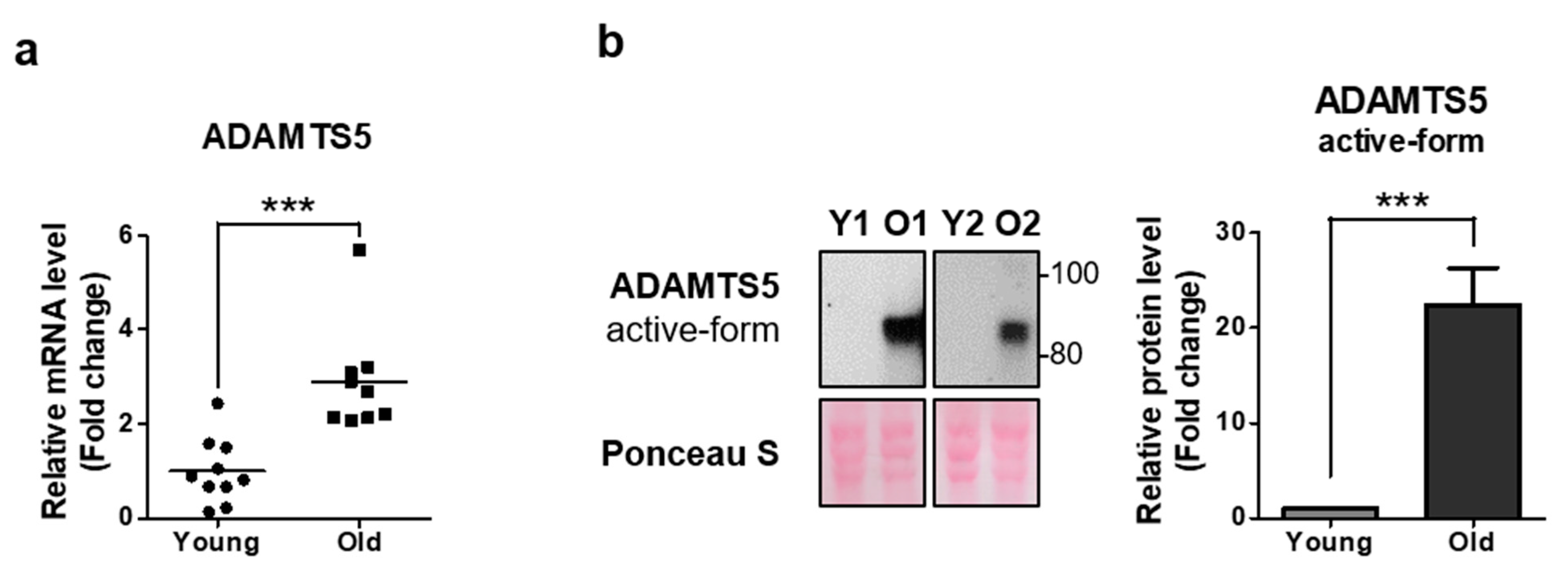
2.4. ADAMTS5 Expression Is Increased in the Dermis of Elderly People
3. Discussion
4. Materials and Methods
4.1. Study/Ethical Approval
4.2. Cell Culture
4.3. Treatment with Recombinant Human IGF-1 (rhIGF-1)
4.4. Gene Silencing with siRNA
4.5. Quantitative Real-Time PCR
4.6. Western Blot Analysis
4.7. Human Skin Samples
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| IGF-1 | Insulin-like growth factor-1 |
| ADAMTS5 | a disintegrin and metalloproteinase with thrombospondin motifs 5 |
| PG | proteoglycan |
| GAG | glycosaminoglycan |
| NHDFs | normal human dermal fibroblasts |
| DS | dermatan sulfate |
| MMP | matrix metalloproteinase |
| Act D | actinomycin D |
| CHX | cycloheximide |
| rhIGF-1 | recombinant human IGF-1 |
| siRNA | small interference RNA |
References
- Perrimon, N.; Bernfield, M. Cellular functions of proteoglycans—An overview. Semin. Cell Dev. Biol. 2001, 12, 65–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaefer, L. Complexity of danger: The diverse nature of damage-associated molecular patterns. J. Biol. Chem 2014, 289, 35237–35245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrino, D.A.; Calabro, A.; Darr, A.B.; Dours-Zimmermann, M.T.; Sandy, J.D.; Zimmermann, D.R.; Sorrell, J.M.; Hascall, V.C.; Caplan, A.I. Age-related differences in human skin proteoglycans. Glycobiology 2011, 21, 257–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Liu, Y.; Xia, W.; Lei, D.; Voorhees, J.J.; Fisher, G.J. Age-dependent alterations of decorin glycosaminoglycans in human skin. Sci Rep. 2013, 3, 2422. [Google Scholar] [CrossRef] [Green Version]
- Carrino, D.A.; Onnerfjord, P.; Sandy, J.D.; Cs-Szabo, G.; Scott, P.G.; Sorrell, J.M.; Heinegard, D.; Caplan, A.I. Age-related changes in the proteoglycans of human skin. Specific cleavage of decorin to yield a major catabolic fragment in adult skin. J. Biol. Chem 2003, 278, 17566–17572. [Google Scholar] [CrossRef] [Green Version]
- Jin, C.L.; Oh, J.H.; Han, M.; Shin, M.K.; Yao, C.; Park, C.H.; Jin, Z.H.; Chung, J.H. UV irradiation-induced production of monoglycosylated biglycan through downregulation of xylosyltransferase 1 in cultured human dermal fibroblasts. J. Derm. Sci. 2015, 79, 20–29. [Google Scholar] [CrossRef]
- Lee, D.H.; Oh, J.H.; Chung, J.H. Glycosaminoglycan and proteoglycan in skin aging. J. Derm. Sci. 2016, 83, 174–181. [Google Scholar] [CrossRef]
- Danielson, K.G.; Baribault, H.; Holmes, D.F.; Graham, H.; Kadler, K.E.; Iozzo, R.V. Targeted disruption of decorin leads to abnormal collagen fibril morphology and skin fragility. J. Cell Biol. 1997, 136, 729–743. [Google Scholar] [CrossRef] [Green Version]
- Zeng, J.P.; Bi, B.; Chen, L.; Yang, P.; Guo, Y.; Zhou, Y.Q.; Liu, T.Y. Repeated exposure of mouse dermal fibroblasts at a sub-cytotoxic dose of UVB leads to premature senescence: A robust model of cellular photoaging. J. Derm. Sci. 2014, 73, 49–56. [Google Scholar] [CrossRef]
- Xia, W.; Quan, T.; Hammerberg, C.; Voorhees, J.J.; Fisher, G.J. A mouse model of skin aging: Fragmentation of dermal collagen fibrils and reduced fibroblast spreading due to expression of human matrix metalloproteinase-1. J. Derm. Sci. 2015, 78, 79–82. [Google Scholar] [CrossRef]
- Li, Y.; Xia, W.; Liu, Y.; Remmer, H.A.; Voorhees, J.; Fisher, G.J. Solar ultraviolet irradiation induces decorin degradation in human skin likely via neutrophil elastase. PLoS ONE 2013, 8, e72563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corsi, A.; Xu, T.; Chen, X.D.; Boyde, A.; Liang, J.; Mankani, M.; Sommer, B.; Iozzo, R.V.; Eichstetter, I.; Robey, P.G.; et al. Phenotypic effects of biglycan deficiency are linked to collagen fibril abnormalities, are synergized by decorin deficiency, and mimic Ehlers-Danlos-like changes in bone and other connective tissues. J. Bone Miner. Res. 2002, 17, 1180–1189. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.Y.; Johnson, P.Y.; Braun, K.R.; Hinek, A.; Fischer, J.W.; O’Brien, K.D.; Starcher, B.; Clowes, A.W.; Merrilees, M.J.; Wight, T.N. Retrovirally mediated overexpression of glycosaminoglycan-deficient biglycan in arterial smooth muscle cells induces tropoelastin synthesis and elastic fiber formation in vitro and in neointimae after vascular injury. Am. J. Pathol 2008, 173, 1919–1928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schonherr, E.; Beavan, L.A.; Hausser, H.; Kresse, H.; Culp, L.A. Differences in decorin expression by papillary and reticular fibroblasts in vivo and in vitro. Biochem J. 1993, 290 Pt 3, 893–899. [Google Scholar] [CrossRef] [Green Version]
- Seidler, D.G.; Faiyaz-Ul-Haque, M.; Hansen, U.; Yip, G.W.; Zaidi, S.H.; Teebi, A.S.; Kiesel, L.; Gotte, M. Defective glycosylation of decorin and biglycan, altered collagen structure, and abnormal phenotype of the skin fibroblasts of an Ehlers-Danlos syndrome patient carrying the novel Arg270Cys substitution in galactosyltransferase I (beta4GalT-7). J. Mol. Med. (Berl) 2006, 84, 583–594. [Google Scholar] [CrossRef]
- Tufvesson, E.; Malmstrom, J.; Marko-Varga, G.; Westergren-Thorsson, G. Biglycan isoforms with differences in polysaccharide substitution and core protein in human lung fibroblasts. Eur. J. Biochem./Febs 2002, 269, 3688–3696. [Google Scholar] [CrossRef]
- Wiberg, C.; Heinegard, D.; Wenglen, C.; Timpl, R.; Morgelin, M. Biglycan organizes collagen VI into hexagonal-like networks resembling tissue structures. J. Biol. Chem. 2002, 277, 49120–49126. [Google Scholar] [CrossRef] [Green Version]
- Trask, B.C.; Trask, T.M.; Broekelmann, T.; Mecham, R.P. The microfibrillar proteins MAGP-1 and fibrillin-1 form a ternary complex with the chondroitin sulfate proteoglycan decorin. Mol. Biol Cell 2000, 11, 1499–1507. [Google Scholar] [CrossRef] [Green Version]
- Reinboth, B.; Hanssen, E.; Cleary, E.G.; Gibson, M.A. Molecular interactions of biglycan and decorin with elastic fiber components: Biglycan forms a ternary complex with tropoelastin and microfibril-associated glycoprotein 1. J. Biol. Chem. 2002, 277, 3950–3957. [Google Scholar] [CrossRef] [Green Version]
- Kolb, M.; Margetts, P.J.; Sime, P.J.; Gauldie, J. Proteoglycans decorin and biglycan differentially modulate TGF-beta-mediated fibrotic responses in the lung. Am. J. Physiol. Lung Cell Mol. Physiol. 2001, 280, L1327–L1334. [Google Scholar] [CrossRef]
- Hildebrand, A.; Romaris, M.; Rasmussen, L.M.; Heinegard, D.; Twardzik, D.R.; Border, W.A.; Ruoslahti, E. Interaction of the small interstitial proteoglycans biglycan, decorin and fibromodulin with transforming growth factor beta. Biochem J. 1994, 302 Pt 2, 527–534. [Google Scholar] [CrossRef]
- Santra, M.; Reed, C.C.; Iozzo, R.V. Decorin binds to a narrow region of the epidermal growth factor (EGF) receptor, partially overlapping but distinct from the EGF-binding epitope. J. Biol. Chem. 2002, 277, 35671–35681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nili, N.; Cheema, A.N.; Giordano, F.J.; Barolet, A.W.; Babaei, S.; Hickey, R.; Eskandarian, M.R.; Smeets, M.; Butany, J.; Pasterkamp, G.; et al. Decorin inhibition of PDGF-stimulated vascular smooth muscle cell function: Potential mechanism for inhibition of intimal hyperplasia after balloon angioplasty. Am. J. Pathol. 2003, 163, 869–878. [Google Scholar] [CrossRef]
- Goldoni, S.; Humphries, A.; Nystrom, A.; Sattar, S.; Owens, R.T.; McQuillan, D.J.; Ireton, K.; Iozzo, R.V. Decorin is a novel antagonistic ligand of the Met receptor. J. Cell Biol. 2009, 185, 743–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iozzo, R.V.; Buraschi, S.; Genua, M.; Xu, S.Q.; Solomides, C.C.; Peiper, S.C.; Gomella, L.G.; Owens, R.C.; Morrione, A. Decorin antagonizes IGF receptor I (IGF-IR) function by interfering with IGF-IR activity and attenuating downstream signaling. J. Biol. Chem. 2011, 286, 34712–34721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.D.; Fisher, L.W.; Robey, P.G.; Young, M.F. The small leucine-rich proteoglycan biglycan modulates BMP-4-induced osteoblast differentiation. Faseb J. 2004, 18, 948–958. [Google Scholar] [CrossRef] [PubMed]
- Miguez, P.A.; Terajima, M.; Nagaoka, H.; Mochida, Y.; Yamauchi, M. Role of glycosaminoglycans of biglycan in BMP-2 signaling. Biochem. Biophys. Res. Commun. 2011, 405, 262–266. [Google Scholar] [CrossRef] [Green Version]
- Berendsen, A.D.; Fisher, L.W.; Kilts, T.M.; Owens, R.T.; Robey, P.G.; Gutkind, J.S.; Young, M.F. Modulation of canonical Wnt signaling by the extracellular matrix component biglycan. Proc. Natl. Acad. Sci. USA 2011, 108, 17022–17027. [Google Scholar] [CrossRef] [Green Version]
- Iacob, S.; Cs-Szabo, G. Biglycan regulates the expression of EGF receptors through EGF signaling pathways in human articular chondrocytes. Connect. Tissue Res. 2010, 51, 347–358. [Google Scholar] [CrossRef]
- Babelova, A.; Moreth, K.; Tsalastra-Greul, W.; Zeng-Brouwers, J.; Eickelberg, O.; Young, M.F.; Bruckner, P.; Pfeilschifter, J.; Schaefer, R.M.; Grone, H.J.; et al. Biglycan, a danger signal that activates the NLRP3 inflammasome via toll-like and P2X receptors. J. Biol. Chem. 2009, 284, 24035–24048. [Google Scholar] [CrossRef] [Green Version]
- Oh, J.-H.; Chung, J.H. Do Proteoglycans Mediate Chronic Photoaging? In Cutaneous Photoaging; Watson, R.E.B., Griffiths, C.E.M., Eds.; Royal Society of Chemistry: Manchester, UK, 2019; pp. 231–274. [Google Scholar]
- Christopoulos, P.F.; Msaouel, P.; Koutsilieris, M. The role of the insulin-like growth factor-1 system in breast cancer. Mol. Cancer 2015, 14, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laron, Z. Insulin-like growth factor 1 (IGF-1): A growth hormone. Mol. Pathol. 2001, 54, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Lewis, D.A.; Travers, J.B.; Somani, A.K.; Spandau, D.F. The IGF-1/IGF-1R signaling axis in the skin: A new role for the dermis in aging-associated skin cancer. Oncogene 2010, 29, 1475–1485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gennigens, C.; Menetrier-Caux, C.; Droz, J.P. Insulin-Like Growth Factor (IGF) family and prostate cancer. Crit. Rev. Oncol. /Hematol. 2006, 58, 124–145. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Rohan, T. Role of the insulin-like growth factor family in cancer development and progression. J. Natl Cancer Inst. 2000, 92, 1472–1489. [Google Scholar] [CrossRef] [PubMed]
- Bartke, A.; Chandrashekar, V.; Dominici, F.; Turyn, D.; Kinney, B.; Steger, R.; Kopchick, J.J. Insulin-like growth factor 1 (IGF-1) and aging: Controversies and new insights. Biogerontology 2003, 4, 1–8. [Google Scholar] [CrossRef]
- Oh, J.H.; Kim, Y.K.; Jung, J.Y.; Shin, J.E.; Chung, J.H. Changes in glycosaminoglycans and related proteoglycans in intrinsically aged human skin in vivo. Exp. Derm. 2011, 20, 454–456. [Google Scholar] [CrossRef]
- Zhu, H.; Xu, Y.; Gong, F.; Shan, G.; Yang, H.; Xu, K.; Zhang, D.; Cheng, X.; Zhang, Z.; Chen, S.; et al. Reference ranges for serum insulin-like growth factor I (IGF-I) in healthy Chinese adults. PLoS ONE 2017, 12, e0185561. [Google Scholar] [CrossRef]
- Blackstock, C.D.; Higashi, Y.; Sukhanov, S.; Shai, S.Y.; Stefanovic, B.; Tabony, A.M.; Yoshida, T.; Delafontaine, P. Insulin-like Growth Factor-1 Increases Synthesis of Collagen Type I via Induction of the mRNA-binding Protein LARP6 Expression and Binding to the 5 ’ Stem-loop of COL1a1 and COL1a2 mRNA. J. Biol. Chem. 2014, 289, 7264–7274. [Google Scholar] [CrossRef] [Green Version]
- Verma, P.; Dalal, K. ADAMTS-4 and ADAMTS-5: Key enzymes in osteoarthritis. J. Cell. Biochem. 2011, 112, 3507–3514. [Google Scholar] [CrossRef]
- Longpre, J.M.; McCulloch, D.R.; Koo, B.H.; Alexander, J.P.; Apte, S.S.; Leduc, R. Characterization of proADAMTS5 processing by proprotein convertases. Int. J. Biochem. Cell Biol. 2009, 41, 1116–1126. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Sharghi-Namini, S.; Rao, N.; Ge, R.W. ADAMTS5 Functions as an Anti-Angiogenic and Anti-Tumorigenic Protein Independent of Its Proteoglycanase Activity. Am. J. Pathol. 2012, 181, 1056–1068. [Google Scholar] [CrossRef] [PubMed]
- Fosang, A.J.; Rogerson, F.M.; East, C.J.; Stanton, H. ADAMTS-5: The story so far. Eur. Cells Mater. 2008, 15, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Iranmanesh, A.; Lizarralde, G.; Veldhuis, J.D. Age and relative adiposity are specific negative determinants of the frequency and amplitude of growth hormone (GH) secretory bursts and the half-life of endogenous GH in healthy men. J. Clin. Endocrinol. Metab. 1991, 73, 1081–1088. [Google Scholar] [CrossRef]
- Noordam, R.; Gunn, D.A.; Tomlin, C.C.; Maier, A.B.; Griffiths, T.; Catt, S.D.; Ogden, S.; Slagboom, P.E.; Westendorp, R.G.; Griffiths, C.E.; et al. Serum insulin-like growth factor 1 and facial ageing: High levels associate with reduced skin wrinkling in a cross-sectional study. Br. J. Derm. 2013, 168, 533–538. [Google Scholar] [CrossRef]
- Dufner, A.; Thomas, G. Ribosomal S6 kinase signaling and the control of translation. Exp. Cell Res. 1999, 253, 100–109. [Google Scholar] [CrossRef]
- Gingras, A.C.; Raught, B.; Sonenberg, N. Regulation of translation initiation by FRAP/mTOR. Genes Dev. 2001, 15, 807–826. [Google Scholar] [CrossRef] [Green Version]
- James, M.J.; Zomerdijk, J.C. Phosphatidylinositol 3-kinase and mTOR signaling pathways regulate RNA polymerase I transcription in response to IGF-1 and nutrients. J. Biol. Chem. 2004, 279, 8911–8918. [Google Scholar] [CrossRef] [Green Version]
- Haase, H.R.; Clarkson, R.W.; Waters, M.J.; Bartold, P.M. Growth factor modulation of mitogenic responses and proteoglycan synthesis by human periodontal fibroblasts. J. Cell Physiol. 1998, 174, 353–361. [Google Scholar] [CrossRef]
- Aggelidakis, J.; Berdiaki, A.; Nikitovic, D.; Papoutsidakis, A.; Papachristou, D.J.; Tsatsakis, A.M.; Tzanakakis, G.N. Biglycan Regulates MG63 Osteosarcoma Cell Growth Through a LPR6/beta-Catenin/IGFR-IR Signaling Axis. Front. Oncol. 2018, 8, 470. [Google Scholar] [CrossRef] [Green Version]
- Roughley, P.J.; Melching, L.I.; Recklies, A.D. Changes in the expression of decorin and biglycan in human articular cartilage with age and regulation by TGF-beta. Matrix Biol. 1994, 14, 51–59. [Google Scholar] [CrossRef]
- Gendron, C.; Kashiwagi, M.; Lim, N.H.; Enghild, J.J.; Thogersen, I.B.; Hughes, C.; Caterson, B.; Nagase, H. Proteolytic activities of human ADAMTS-5: Comparative studies with ADAMTS-4. J. Biol. Chem. 2007, 282, 18294–18306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, R.L.; Zhang, X.M.; Jia, L.N.; Song, W.; Sun, Y.L.; Meng, X.Y.; Peng, X.X. (p)NNS-Conjugated Chitosan Mediated IGF-1 and miR-140 Overexpression in Articular Chondrocytes Improves Cartilage Repair. Biomed. Res. Int. 2019, 2019, 2761241. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Qin, J.; Wang, H.; Magdalou, J.; Chen, L. Effects of adenovirus-mediated bFGF, IL-1Ra and IGF-1 gene transfer on human osteoarthritic chondrocytes and osteoarthritis in rabbits. Exp. Mol. Med. 2010, 42, 684–695. [Google Scholar] [CrossRef]
- Fisher, G.J.; Datta, S.C.; Talwar, H.S.; Wang, Z.Q.; Varani, J.; Kang, S.; Voorhees, J.J. Molecular basis of sun-induced premature skin ageing and retinoid antagonism. Nature 1996, 379, 335–339. [Google Scholar] [CrossRef]
- Chung, J.H.; Seo, J.Y.; Lee, M.K.; Eun, H.C.; Lee, J.H.; Kang, S.; Fisher, G.J.; Voorhees, J.J. Ultraviolet modulation of human macrophage metalloelastase in human skin in vivo. J. Investig. Derm. 2002, 119, 507–512. [Google Scholar] [CrossRef] [Green Version]
- Parkinson, L.G.; Toro, A.; Zhao, H.; Brown, K.; Tebbutt, S.J.; Granville, D.J. Granzyme B mediates both direct and indirect cleavage of extracellular matrix in skin after chronic low-dose ultraviolet light irradiation. Aging Cell 2015, 14, 67–77. [Google Scholar] [CrossRef]
- Boivin, W.A.; Shackleford, M.; Vanden Hoek, A.; Zhao, H.; Hackett, T.L.; Knight, D.A.; Granville, D.J. Granzyme B cleaves decorin, biglycan and soluble betaglycan, releasing active transforming growth factor-beta1. PLoS ONE 2012, 7, e33163. [Google Scholar] [CrossRef]
- Zhen, E.Y.; Brittain, I.J.; Laska, D.A.; Mitchell, P.G.; Sumer, E.U.; Karsdal, M.A.; Duffin, K.L. Characterization of metalloprotease cleavage products of human articular cartilage. Arthritis Rheum 2008, 58, 2420–2431. [Google Scholar] [CrossRef]
- Hausmann, C.; Zoschke, C.; Wolff, C.; Darvin, M.E.; Sochorova, M.; Kovacik, A.; Wanjiku, B.; Schumacher, F.; Tigges, J.; Kleuser, B.; et al. Fibroblast origin shapes tissue homeostasis, epidermal differentiation, and drug uptake. Sci. Rep. 2019, 9, 2913. [Google Scholar] [CrossRef]
- Rinn, J.L.; Bondre, C.; Gladstone, H.B.; Brown, P.O.; Chang, H.Y. Anatomic demarcation by positional variation in fibroblast gene expression programs. PLoS Genet. 2006, 2, e119. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.; Lim, J.; Oh, J.-H.; Cho, S.; Chung, J.H. IGF-1 Upregulates Biglycan and Decorin by Increasing Translation and Reducing ADAMTS5 Expression. Int. J. Mol. Sci. 2021, 22, 1403. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031403
Lee H, Lim J, Oh J-H, Cho S, Chung JH. IGF-1 Upregulates Biglycan and Decorin by Increasing Translation and Reducing ADAMTS5 Expression. International Journal of Molecular Sciences. 2021; 22(3):1403. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031403
Chicago/Turabian StyleLee, Hanon, Jiyeong Lim, Jang-Hee Oh, Soyun Cho, and Jin Ho Chung. 2021. "IGF-1 Upregulates Biglycan and Decorin by Increasing Translation and Reducing ADAMTS5 Expression" International Journal of Molecular Sciences 22, no. 3: 1403. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031403