
Relative Contribution of Proprioceptive and Vestibular Sensory Systems to Locomotion: Opportunities for Discovery in the Age of Molecular Science

{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Intra-Leg Coordination during Locomotion
1.2. Inter-Leg Coordination during Locomotion
2. Role of Segmental Somatosensory Sensory Feedback in Locomotion
2.1. Overview on Segmental Sensory Feedback on Locomotion
2.2. Exteroceptive Sensory Feedback and Locomotion
2.3. Segmental Sensory Feedback and Posture
2.4. Proprioceptive Sensory Feedback and Locomotion
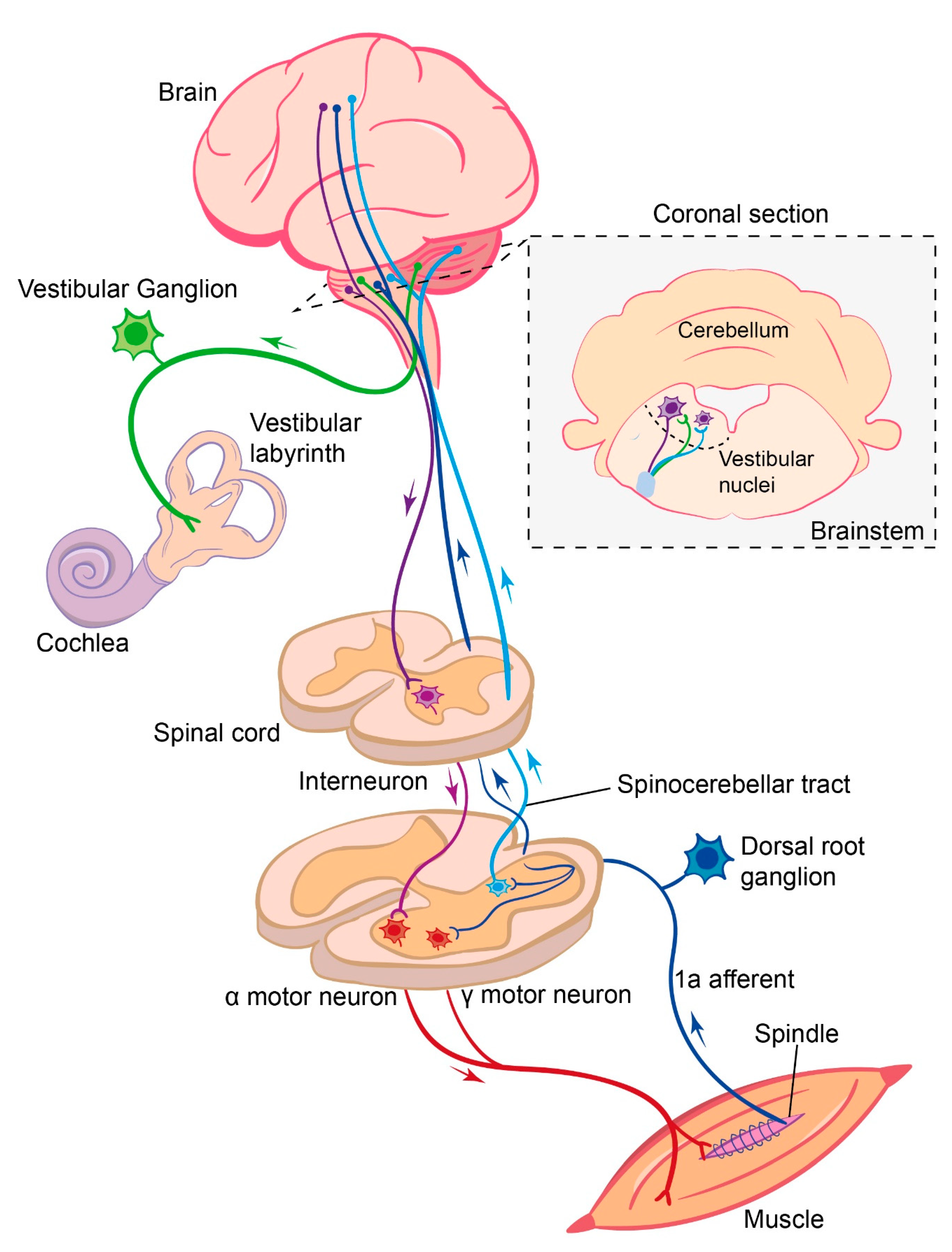
3. Role of Vestibular Sensory Feedback in Locomotion
3.1. Vestibular Sensory Feedback
3.2. Maintaining Vision during Locomotion
3.3. Maintaining Stability and Balance
3.4. Vestibular Damage and Gait
3.5. Locomotor Speed and Vestibular Influence
4. Perspective
4.1. Effect of Segmental Somatosensory and Vestibular Feedback on the Generation of Locomotion
4.2. State-Dependent Modulation of Segmental Somatosensory and Vestibular Feedback
4.3. Influence of Segmental Somatosensory and Vestibular Feedback at Different Locomotor Speeds
4.4. Current Working Hypothesis
- (i).
- Slower locomotor speeds are more variable and unstable than higher speeds. As slower walking speeds are more likely to be associated with exploratory activities, they require frequent changes in the heading direction and body position. Here, the nervous system must consider how each movement or variation in foot placement could affect the equilibrium. Therefore, a direct link to the head movement, and overall position of the body, is imperative.
- (ii).
- Due to the slower speed of movement, there is ample time for the brain to influence the spinal networks to control foot placement and posture. Multiple descending pathways influence locomotion, with the vestibular system “in charge” of making sure that these movements do not cause the animal to lose balance.
- (iii).
- During walking, the features of the step cycle, such as overall cycle duration or length of the support phase, are variable at different walking speeds, whereas the timing of these phases during running is consistent across speeds [130]. High-speed locomotion could therefore be more stereotyped and perhaps dominated by local spinal networks where segmental somatosensory feedback is the main source of sensory feedback, allowing minor adaptations to the musculature as locomotion continues.
- (iv).
- The segmental proprioceptive feedback seems to be important for all speeds, it is necessary for higher speeds, as, without them, animals do not locomote at higher speeds.
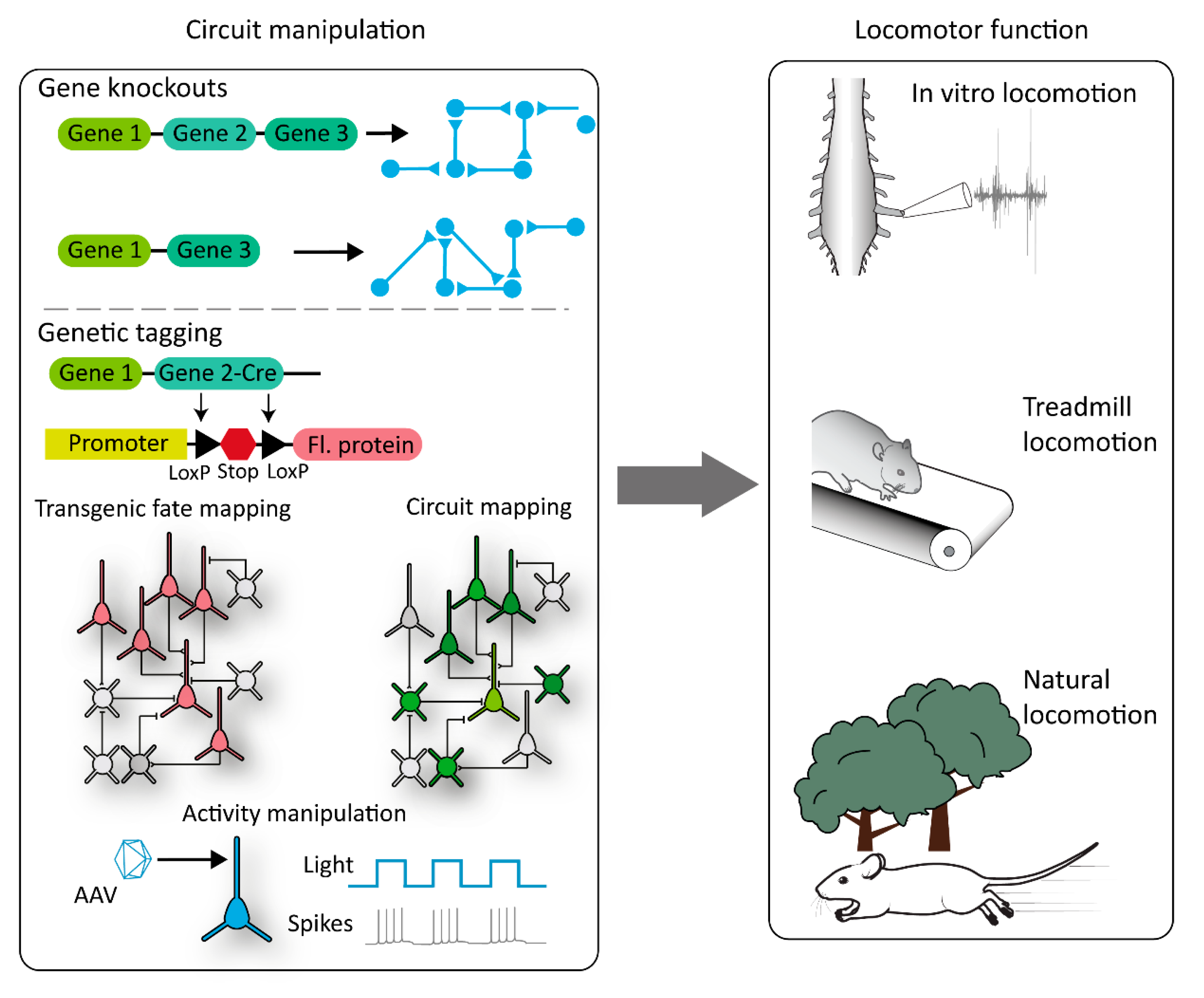
5. Future Perspective in the Age of Molecular Sciences
Funding
Acknowledgments
Conflicts of Interest
References
- Grillner, S. Control of locomotion in bipeds, tetrapods, and fish. In Handbook of Physiology: The Nervous System, 2, Motor Control; Brooks, V., Ed.; Wiley: New York, NY, USA, 1981; pp. 1176–1236. [Google Scholar]
- Akay, T. Sensory Feedback Control of Locomotor Pattern Generation in Cats and Mice. Neuroscience 2020, 450, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Grillner, S.; Jessell, T.M. Measured motion: Searching for simplicity in spinal locomotor networks. Curr. Opin. Neurobiol. 2009, 19, 572–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grillner, S.; El Manira, A. Current Principles of Motor Control, with Special Reference to Vertebrate Locomotion. Physiol. Rev. 2020, 100, 271–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goulding, M. Circuits controlling vertebrate locomotion: Moving in a new direction. Nat. Rev. Neurosci. 2009, 10, 507–518. [Google Scholar] [CrossRef]
- Kiehn, O. Decoding the organization of spinal circuits that control locomotion. Nat. Rev. Neurosci. 2016, 17, 24–238. [Google Scholar] [CrossRef]
- Guertin, P.A. Central Pattern Generator for Locomotion: Anatomical, Physiological, and Pathophysiological Considerations. Front. Neurol. 2013, 3, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Santuz, A.; Akay, T.; Mayer, W.P.; Wells, T.L.; Schroll, A.; Arampatzis, A. Modular organization of murine locomotor pattern in the presence and absence of sensory feedback from muscle spindles. J. Physiol. 2019, 597, 3147–3165. [Google Scholar] [CrossRef]
- McVea, D.A.; Pearson, K.G. Object Avoidance During Locomotion. Adv. Exp. Med. Biol. 2009, 629, 293–315. [Google Scholar] [CrossRef]
- Witts, E.C.; Murray, A.J. Vestibulospinal contributions to mammalian locomotion. Curr. Opin. Physiol. 2019, 8, 56–62. [Google Scholar] [CrossRef]
- Mayer, W.P.; Murray, A.J.; Brenner-Morton, S.; Jessell, T.M.; Tourtellotte, W.G.; Akay, T. Role of muscle spindle feedback in regulating muscle activity strength during walking at different speed in mice. J. Neurophysiol. 2018, 120, 2484–2497. [Google Scholar] [CrossRef]
- Juvin, L.; Simmers, J.; Morin, D. Propriospinal circuitry underlying interlimb coordination in mammalian quadrupedal locomotion. J. Neurosci. 2005, 25, 6025–6035. [Google Scholar] [CrossRef] [PubMed]
- Akay, T.; McVea, D.A.; Tachibana, A.; Pearson, K.G. Coordination of fore and hind leg stepping in cats on a transversely-split treadmill. Exp. Brain Res. 2006, 175. [Google Scholar] [CrossRef] [PubMed]
- Frigon, A. The neural control of interlimb coordination during mammalian locomotion. J. Neurophysiol. 2017, 117, 2224–2241. [Google Scholar] [CrossRef] [PubMed]
- Winter, D.A. A.B.C (Anatomy, Biomechanics and Control) of Balance During Standing and Walking; Waterloo Biomechanics: Waterloo, CA, USA, 1995. [Google Scholar]
- Hof, A.L.; Gazendam, M.G.J.; Sinke, W.E. The condition for dynamic stability. J. Biomech. 2005, 38, 1–8. [Google Scholar] [CrossRef]
- Lemieux, M.; Josset, N.; Roussel, M.; Couraud, S.; Bretzner, F. Speed-dependent modulation of the locomotor behavior in adult mice reveals attractor and transitional gaits. Front. Neurosci. 2016, 10, 42. [Google Scholar] [CrossRef] [Green Version]
- Novacheck, T.F. The biomechanics of running. Gait Posture 1998, 7, 77–95. [Google Scholar] [CrossRef]
- Mendes, C.S.; Bartos, I.; Akay, T.; Márka, S.; Mann, R.S. Quantification of gait parameters in freely walking wild type and sensory deprived Drosophila melanogaster. eLife 2013, 2013, 1–11. [Google Scholar] [CrossRef]
- Wosnitza, A.; Bockemühl, T.; Dübbert, M.; Scholz, H.; Büschges, A. Inter-leg coordination in the control of walking speed in Drosophila. J. Exp. Biol. 2013, 216, 480–491. [Google Scholar] [CrossRef] [Green Version]
- Hildebrand, M. The Quadrupedal Gaits of Vertebrates. Bioscience 1989, 39, 766–775. [Google Scholar] [CrossRef]
- Cavagna, G.A.; Thys, H.; Zamboni, A. The sources of external work in level walking and running. J. Physiol. 1976, 262, 639–657. [Google Scholar] [CrossRef]
- Cavagna, G.A.; Heglund, N.C.; Taylor, C.R. Mechanical work basic mechanisms in terrestrial locomotion: Two for minimizing energy expenditure. Am. J. Physiol. 1977, 233, R243–R261. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.R.; Farley, C.T. Determinants of the center of mass trajectory in human walking and running. J. Exp. Biol. 1998, 201, 2935–2944. [Google Scholar]
- Bidaye, S.S.; Bockemühl, X.T.; Büschges, A. Six-legged walking in insects: How CPGs, peripheral feedback, and descending signals generate coordinated and adaptive motor rhythms. J. Neurophysiol. 2018, 119, 459–475. [Google Scholar] [CrossRef] [PubMed]
- Hurteau, M.-F.; Thibaudier, Y.; Dambreville, C.; Danner, S.M.; Rybak, I.A.; Frigon, A. Intralimb and Interlimb Cutaneous Reflexes during Locomotion in the Intact Cat. J. Neurosci. 2018, 38, 4104–4122. [Google Scholar] [CrossRef] [PubMed]
- Bent, L.R.; McFadyen, B.J.; Merkley, V.F.; Kennedy, P.M.; Inglis, J.T. Magnitude effects of galvanic vestibular stimulation on the trajectory of human gait, Neurosci. Lett. Lett. 2000, 279, 157–160. [Google Scholar]
- Bigley, G.K. Sensation. In Clinical Methods History Physical Laboratory Examinations, 3rd ed.; Walker, H., Hall, W., Hurst, J., Eds.; Butterworths: Boston, UK, 1990; pp. 343–350. [Google Scholar]
- Jack, J.J.B. Some methods for selective activation of muscle afferent fibres. In Studies in Neurophysiology: Presented to A. K. McIntryre; Porter, R., Ed.; Cambridge University Press: London, UK; Melbourne: New York, NY, USA, 1978; pp. 155–176. [Google Scholar]
- Bewick, G.S.; Banks, R.W. Mechanotransduction in the muscle spindle. Eur. J. Physiol. 2015, 467, 175–190. [Google Scholar] [CrossRef] [Green Version]
- Collins, D.F.; Refshauge, K.M.; Todd, G.; Gandevia, S.C. Cutaneous Receptors Contribute to Kinesthesia at the Index Finger, Elbow, and Knee. J. Neurophysiol. 2005, 94, 1699–1706. [Google Scholar] [CrossRef] [Green Version]
- Collins, D.F.; Prochazka, A. Movement illusions evoked by ensemble cutaneous input from the dorsum of the human hand. J. Physiol. 1996, 496, 857–871. [Google Scholar] [CrossRef]
- Hulliger, M.; Nordh, E.; Thelin, A.-E.; Vallbo, A.B. The responses of afferent fibers from the glaborous skin of the hand during voluntary finger movements in man. J. Physiol. 1979, 291, 233–249. [Google Scholar] [CrossRef]
- Mildren, R.L.; Hare, C.M.; Bent, L.R. Cutaneous afferent feedback from the posterior ankle contributes to proprioception. Neurosci. Lett. 2017, 636, 145–150. [Google Scholar] [CrossRef]
- Abraira, V.E.; Ginty, D.D. The Sensory Neurons of Touch. Neuron 2013, 79, 618–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatto, G.; Smith, K.M.; Ross, S.E.; Goulding, M. Neuronal diversity in the somatosensory system: Bridging the gap between cell type and function. Curr. Opin. Neurobiol. 2019, 56, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Lynn, B. Somatosensory receptors and their CNS connections. Annu. Rev. Physiol. 1975, 37, 105–127. [Google Scholar] [CrossRef] [PubMed]
- Wall, P.D.; Dubner, R. Somatosensory pathways. Annu. Rev. Physiol. 1972, 34, 315–336. [Google Scholar] [CrossRef] [PubMed]
- William, D.W., Jr. The somatosensory system, with emphasis on structures important for pain. Brain Res. Rev. 2007, 55, 297–313. [Google Scholar] [CrossRef]
- Jankowska, E. Spinal interneuronal networks in the cat: Elementary components. Brain Res. Rev. 2008, 57, 46–55. [Google Scholar] [CrossRef] [Green Version]
- Jankowska, E. Spinal interneurons. In Neuroscience 21st Century: From Basic to Clinical; Springer: New York, NY, USA, 2013; pp. 1–3111. [Google Scholar] [CrossRef]
- Koch, S.C. ScienceDirect Motor task-selective spinal sensorimotor interneurons in mammalian circuits. Curr. Opin. Physiol. 2019, 8, 129–135. [Google Scholar] [CrossRef]
- Jankowska, E.; Hammar, I. Interactions between spinal interneurons and ventral spinocerebellar tract neurons. J. Physiol. 2013, 591, 5445–5451. [Google Scholar] [CrossRef] [Green Version]
- Stecina, K.; Fedirchuk, B.; Hultborn, H. Information to cerebellum on spinal motor networks mediated by the dorsal spinocerebellar tract. J. Physiol. 2013, 591, 5433–5443. [Google Scholar] [CrossRef] [Green Version]
- Diaz, E.; Morales, H. Spinal Cord Anatomy and Clinical Syndromes. Semin. Ultrasound CT MRI 2016, 37, 360–371. [Google Scholar] [CrossRef]
- Eccles, J.C.; Eccles, R.M.; Lundberg, A. The Convergence of Monosynaptic Excitatory Afferents on to Many Different Species of Alpha Motoneurons. J. Physiol. 1957, 137, 22–50. [Google Scholar] [CrossRef] [PubMed]
- Frank, E.; Mendelson, B. Specification of synaptic connectins between sensory and motor neurons in the developing spinal cord. J. Neurobiol. 1990, 21, 33–50. [Google Scholar] [CrossRef] [PubMed]
- Sherrington, C.S. Flexion-reflex of the limb, crossed extension-reflex, and reflex stepping and standing. J. Physiol. 1910, 40, 28–121. [Google Scholar] [CrossRef] [PubMed]
- Bouyer, L.J.G.; Rossignol, S. Contribution of Cutaneous Inputs From the Hindpaw to the Control of Locomotion. I. Intact Cats. J. Neurophysiol. 2003, 90, 3625–3639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouyer, L.J.G.; Rossignol, S. Contribution of Cutaneous Inputs From the Hindpaw to the Control of Locomotion. II. Spinal Cats. J. Neurophysiol. 2003, 90, 3640–3653. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Latash, E.M.; Molkov, Y.I.; Klishko, A.N.; Frigon, A.; Deweerth, S.P.; Prilutsky, B.I. Cutaneous sensory feedback from paw pads affects lateral balance control during split-belt locomotion in the cat. J. Exp. Biol. 2019, 222, jeb198648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duysens, J.; Pearson, K.G. The role of cutaneous afferents from the distal hindlimb in the regulation of the step cycle of thalamic cats. Exp. Brain Res. 1976, 24, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Forssberg, H.; Grillner, S.; Rossignol, S. Phase dependent reflex reversal during walking in chronic spinal cats. Brain Res. 1975, 85, 103–107. [Google Scholar] [CrossRef]
- Forssberg, H.; Grillner, S.; Rossignol, S. Phasic gain control of reflexes from the dorsum of the paw during spinal locomotion. Brain Res. 1977, 132, 121–139. [Google Scholar] [CrossRef]
- Forssberg, H. Stumbling corrective reaction: A phase-dependent compensatory reaction during locomotion. J. Neurophysiol. 1979, 42, 936–953. [Google Scholar] [CrossRef]
- Bolton, D.A.E.; Misiaszek, J.E. Contribution of Hindpaw Cutaneous Inputs to the Control of Lateral Stability During Walking in the Cat. J. Neurophysiol. 2009, 102, 1711–1724. [Google Scholar] [CrossRef] [PubMed]
- Honeycutt, C.F.; Nichols, T.R. Disruption of cutaneous feedback alters magnitude but not direction of muscle responses to postural perturbations in the decerebrate cat. Exp. Brain Res. 2010, 203, 765–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurteau, M.-F.; Frigon, A. A Spinal Mechanism Related to Left–Right Symmetry Reduces Cutaneous Reflex Modulation Independently of Speed During Split-Belt Locomotion. J. Neurosci. 2018, 38, 10314–10328. [Google Scholar] [CrossRef] [PubMed]
- Bent, L.R.; Lowrey, C.R. Single low-threshold afferents innervating the skin of the human foot modulate ongoing muscle activity in the upper limbs. J. Neurophysiol. 2013, 109, 1614–1625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honeycutt, C.F.; Nardelli, P.; Cope, T.C.; Nichols, T.R. Muscle spindle responses to horizontal support surface perturbation in the anesthetized cat: Insights into the role of autogenic feedback in whole body postural control. J. Neurophysiol. 2012, 108, 1253–1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mauritz, K.H.; Dietz, V.; Haller, M. Balancing as a clinical test in the differential diagnosis of sensory-motor disorders. J. Neurol. Neurosurg. Psychiatry 1980, 43, 407–412. [Google Scholar] [CrossRef] [Green Version]
- Horak, F.B.; Nashner, L.M.; Diener, H.C. Postural strategies associated with somatosensory and vestibular loss. Exp. Brain Res. 1990, 82, 167–177. [Google Scholar] [CrossRef]
- Roden-Reynolds, D.C.; Walker, M.H.; Wasserman, C.R.; Dean, J.C. Hip proprioceptive feedback influences the control of mediolateral stability during human walking. J. Neurophysiol. 2015, 114, 2220–2229. [Google Scholar] [CrossRef] [Green Version]
- Clarac, F. The History of Reflexes Part 1: From Descartes to Pavlov. IBRO Hist. Neurosci. 2005. [Google Scholar]
- Aggelopoulos, N.C.; Edgley, S.A. Segmental localisation of the relays mediating crossed inhibition of hindlimb motoneurones from group II afferents in the anaesthetized cat spinal cord. Neurosci. Lett. 1995, 185, 60–64. [Google Scholar] [CrossRef]
- Krutki, P.; Jankowska, E.; Edgley, S.A. Are Crossed Actions of Reticulospinal and Vestibulospinal Neurons on Feline Motoneurons Mediated by the Same or Separate Commissural Neurons? J. Neurosci. 2003, 23, 8041–8050. [Google Scholar] [CrossRef] [PubMed]
- Jankowska, E.; Edgley, S.a.; Krutki, P.; Hammar, I. Functional differentiation and organization of feline midlumbar commissural interneurones. J. Physiol. 2005, 565, 645–658. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Carlin, K.P.; Brownstone, R.M. An in vitro functionally mature mouse spinal cord preparation for the study of spinal motor networks. Brain Res. 1999, 816, 493–499. [Google Scholar] [CrossRef] [Green Version]
- Nakanishi, S.T.; Whelan, P.J. A decerebrate adult mouse model for examining the sensorimotor control of locomotion. J. Neurophysiol. 2012, 107, 500–515. [Google Scholar] [CrossRef] [PubMed]
- Laflamme, O.D.; Akay, T. Excitatory and inhibitory crossed reflex pathways in mice. J. Neurophysiol. 2018, 120, 2897–2907. [Google Scholar] [CrossRef] [PubMed]
- Gervasio, S.; Farina, D.; Sinkjær, T.; Mrachacz-Kersting, N. Crossed reflex reversal during human locomotion. J. Neurophysiol. 2013, 109, 2335–2344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gervasio, S.; Kersting, U.G.; Farina, D.; Mrachacz-Kersting, N. The effect of crossed reflex responses on dynamic stability during locomotion. J. Neurophysiol. 2015, 114, 1034–1040. [Google Scholar] [CrossRef] [Green Version]
- Jankowska, E.; Edgley, S.A. Functional subdivision of feline spinal interneurons in reflex pathways from group Ib and II muscle afferents; an update. Eur. J. Neurosci. 2010, 32, 881–893. [Google Scholar] [CrossRef] [Green Version]
- Akay, T.; Tourtellotte, W.G.; Arber, S.; Jessell, T.M. Degradation of mouse locomotor pattern in the absence of proprioceptive sensory feedback. Proc. Natl. Acad. Sci. USA 2014, 111, 16877–16882. [Google Scholar] [CrossRef] [Green Version]
- Takeoka, A.; Arber, S. Functional Local Proprioceptive Feedback Circuits Initiate and Maintain Locomotor Recovery after Spinal Cord Injury. Cell Rep. 2019, 27, 71–85. [Google Scholar] [CrossRef] [Green Version]
- Lajoie, Y.; Teasdale, N.; Cole, J.D.; Burnett, M.; Bard, C.; Fleury, M.; Forget, R.; Paillard, J.; Lamarre, Y. Gait of a deafferented subject without large myelinated sensory fibers below the neck. Neurology 1996, 47, 109–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grillner, S.; Zangger, P. On the central generation of locomotion in the low spinal cat. Exp. Brain Res. 1979, 34, 241–261. [Google Scholar] [CrossRef] [PubMed]
- Grillner, S.; Zangger, P. The effect of dorsal root transection on the efferent motor pattern in the cat’s hindlimb during locomotion. Acta Physiol. Scand. 1984, 120, 393–405. [Google Scholar] [CrossRef] [PubMed]
- Hiebert, G.W.; Pearson, K.G. Contribution of Sensory Feedback to the Generation of Extensor Activity During Walking in the Decerebrate Cat. J. Neurophysiol. 1999, 81, 758–770. [Google Scholar] [CrossRef] [PubMed]
- Jordan, L.M.; Pratt, C.A.; Menzies, J.E. Locomotion evoked by brain stem stimulation: Occurrence without phasic segmental afferent input. Brain Res. 1979, 177, 204–207. [Google Scholar] [CrossRef]
- Massion, J. Postural control system. Curr. Opin. Neurobiol. 1994, 4, 877–887. [Google Scholar] [CrossRef]
- Gurfinkel, V.S.; Pal’tsev, E.I. Effect of segmentary apparatus of the spinal cord on the execution of simple movement reactions. Biofizika 1965, 10, 855–860. [Google Scholar]
- Palmieri, R.M.; Ingersoll, C.D.; Hoffman, M.A. The Hoffmann Reflex: Methodologic Considerations and Applications for Use in Sports Medicine and Athletic Training Research. J. Athl. Train. 2004, 39, 268–277. [Google Scholar]
- Capaday, C.; Stein, R.B. Difference in the amplitude of the human soleus H reflex during walking and running. J. Physiol. 1987, 392, 513–522. [Google Scholar] [CrossRef]
- Lomeli, J.; Quevedo, J.; Linares, P.; Rudomin, P. Local control of information flow in segmental and ascending collaterals of single afferents. Nature 1998, 395, 600–604. [Google Scholar] [CrossRef]
- Hoffman, M.A.; Koceja, D.M. The effects of vision and task complexity on Hoffmann reflex gain. Brain Res. 1995, 700, 303–307. [Google Scholar] [CrossRef]
- Pinar, S.; Kitano, K.; Koceja, D.M. Role of vision and task complexity on soleus H-reflex gain. J. Electromyogr. Kinesiol. 2010, 20, 354–358. [Google Scholar] [CrossRef] [PubMed]
- Brandt, T.; Strupp, M.; Benson, J. You are better off running than walking with acute vestibulopathy. Lancet 1999, 354, 746. [Google Scholar] [CrossRef]
- Schniepp, R.; Wuehr, M.; Neuhaeusser, M.; Kamenova, M.; Dimitriadis, K.; Klopstock, T.; Strupp, M.; Brandt, T.; Jahn, K. Locomotion Speed Determines Gait Variability in Cerebellar Ataxia and Vestibular Failure. Mov. Disord. 2012, 27, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Angelaki, D.E.; Cullen, K.E. Vestibular System: The Many Facets of a Multimodal Sense. Annu. Rev. Neurosci. 2008, 31, 125–150. [Google Scholar] [CrossRef] [Green Version]
- Goldberg, J.M. Afferent diversity and the organization of central vestibular pathways. Exp. Brain Res. 2000, 130, 277–297. [Google Scholar] [CrossRef]
- Goldberg, J.M.; Wolson, V.J.; Angelki, D.E.; Cullen, K.E.; Fukushima, K.; Buttner-Ennever, J. The Vestibular System: A Sixth Sense; Oxford University Press: Oxford, UK, 2012. [Google Scholar]
- Vidal, P.P.; Cullen, K.; Curthoys, I.S.; Lac, S.D.; Holstein, G.; Idoux, E. Chapter 28—The Vestibular System. In Rat Nervous System Fourth Edi; Elsevier: Amsterdam, The Netherlands, 2015; pp. 805–864. [Google Scholar] [CrossRef]
- Carleton, S.C.; Carpenter, M.B. Distribution of primary vestibular fibers in the brainstem and cerebellum of the monkey. Brain Res. 1984, 294, 281–298. [Google Scholar] [CrossRef]
- Gdowski, G.T.; McCrea, R.A. Neck proprioceptive inputs to primate vestibular nucleus neurons. Exp. Brain Res. 2000, 135, 511–526. [Google Scholar] [CrossRef]
- Wilson, V.J.; Peterson, B.W. Vestibulospinal and reticulospinal systems. In Handbook of Physiology—The Nervous System, Motor Control; Brooks, V.B., Ed.; American Physiological Society: Bethesda, MD, USA, 2011; pp. 667–702. [Google Scholar]
- Voogd, J. Deiters’ Nucleus. Its Role in Cerebellar Ideogenesis. Cerebellum 2016, 15, 54–66. [Google Scholar] [CrossRef] [Green Version]
- Peterson, B.W.; Boyle, R.D. Vestibulocollic Reflexes. In Vestibular System; Highstein, S.M., Fay, R.R., Popper, A.N., Eds.; Springer: New York, NY, USA, 2004; pp. 343–374. [Google Scholar]
- Highstein, S.M.; Partsalis, A.; Arikan, R. Role of the Y-Group of the vestibular nuclei and flocculus of the cerebellum in motor learning of the vertical vestibulo-ocular reflex. Prog. Brain Res. 1997, 114, 383–397. [Google Scholar]
- Grossman, G.E.; Leigh, R.J.; Bruce, E.N.; Huebner, W.P.; Lanska, D.J. Performance of the Human During Locomotion Vestibuloocular Reflex. J. Neuophysiol. 1989, 62, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Lambert, F.M.; Combes, D.; Simmers, J.; Straka, H. Gaze Stabilization by Efference Copy Signaling without Sensory Feedback during Vertebrate Locomotion Franc. Curr. Biol. 2012, 22, 1649–1658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietrich, H.; Wuehr, M. Selective suppression of the vestibulo—Ocular reflex during human locomotion. J. Neurol. 2019, 266, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Allum, J.H.J.; Tang, K.-S.; Carpenter, M.G.; Nijhuis, L.B.O.; Bloem, B.R. Review of first trial responses in balance control: Influence of vestibular loss and Parkinson’s disease. Hum. Mov. Sci. 2011, 30, 279–295. [Google Scholar] [CrossRef]
- Black, F.O.; Shupert, C.L.; Horak, F.B.; Nashner, L.M. Abnormal postural control associated with peripheral vestibular disorders. Prog. Brain Res. 1988, 76, 263–275. [Google Scholar] [PubMed]
- von Holst, E.; Mittelstaedt, H. Das Reafferenzprinzip. (Wechlselwirkungen zwischen Zentralnervensystem und Peripherie). Naturwissenschaften 1950, 20, 464–476. [Google Scholar] [CrossRef]
- Bent, L.R.; Inglis, J.T.; McFadyen, B.J. Vestibular contributions across the execution of a voluntary forward step. Exp. Brain Res. 2002, 143, 100–105. [Google Scholar] [CrossRef]
- Tisserand, R.; Dakin, C.J.; van der Loos, M.H.F.; Croft, E.A.; Inglis, T.J.; Blouin, J.-S. Down regulation of vestibular balance stabilizing mechanisms to enable transition between motor states. eLife 2018, 7, e36123. [Google Scholar] [CrossRef]
- Porciuncula, F.; Johnson, C.C.; Glickman, L.B. The effect of vestibular rehabilitation on adults with bilateral vestibular hypofunction: A systematic review. J. Vestib. Res. 2012, 22, 283–298. [Google Scholar] [CrossRef]
- Schniepp, R.; Möhwald, K.; Wuehr, M. Gait ataxia in humans: Vestibular and cerebellar control of dynamic stability. J. Neurol. 2017, 264, S87–S92. [Google Scholar] [CrossRef]
- McCrum, C.; Lucieer, F.; van de Berg, R.; Willems, P.; Fornos, A.P.; Guinand, N.; Karamanidis, K.; Kingma, H.; Meijer, K. The walking speed-dependency of gait variability in bilateral vestibulopathy and its association with clinical tests of vestibular function. Sci. Rep. 2019, 9, 18392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strupp, M.; Feil, K.; Dieterich, M.; Brandt, T. Bilateral vestibulopathy. In Handbook of Clinical Neurology, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 235–240. [Google Scholar] [CrossRef]
- Jahn, K.; Strupp, M.; Schneider, E.; Dietrich, M.; Brandt, T. Visually induced gait deviations during different locomotion speeds. Exp. Brain Res. 2001, 141, 370–374. [Google Scholar] [CrossRef] [PubMed]
- Bent, L.R.; Inglis, J.T.; McFadyen, B.J. When is Vestibular Information Important During Walking? J. Neurosci. 2004, 92, 1269–1275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bent, L.R.; McFadyen, B.J.; Inglis, J.T. Vestibular Contributions during Human Locomotor Tasks. Exerc. Sport Sci. Rev. 2005, 33, 107–113. [Google Scholar] [CrossRef]
- Jahn, K.; Strupp, M.; Schneider, E.; Dieterich, M.; Brandt, T. Differential effects of vestibular stimulation on walking and running. Neuroreport 2000, 11, 1745–1748. [Google Scholar] [CrossRef]
- Pfeiffer, C.; Serino, A.; Blanke, O. The vestibular system: A spatial reference for bodily self-consciousness. Front. Integr. Neurosci. 2014, 8, 31. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.C.; Kim, J.Y.; Lee, H.N.; Lee, H.H.; Kwon, J.H.; Kim, N.B.; Kim, M.J.; Hwang, J.H.; Han, G.C. A quantitative analysis of gait patterns in vestibular neuritis patients using gyroscope sensor and a continuous walking protocol. J. Neuroeng. Rehabil. 2014, 11, 58. [Google Scholar]
- Anson, E.; Pineault, K.; Bair, W.; Studenski, S.; Agrawal, Y. Reduced vestibular function is associated with longer, slower steps in healthy adults during normal speed walking. Gait Posture 2019, 68, 340–345. [Google Scholar] [CrossRef]
- Perring, S.; Summers, T. Laboratory-free measurement of gait rhythmicity in the assessment of the degree of impairment and the effectiveness of rehabilitation in patients with vertigo resulting from vestibular hypofunction. Physiol. Meas. 2007, 28, 697–705. [Google Scholar] [CrossRef]
- Iwasaki, S.; Fujimoto, C.; Egami, N.; Kinoshita, M.; Togo, F.; Yamamoto, Y.; Yamasoba, T. Noisy vestibular stimulation increases gait speed in normals and in bilateral vestibulopathy. Brain Stimul. 2018, 11, 709–715. [Google Scholar] [CrossRef]
- Brandt, T. Vestibular cortex: Its locations, functions, and disorders. In Vertigo; Springer: New York, NY, USA, 2003; pp. 219–231. [Google Scholar]
- Hashimoto, T.; Taoka, M.; Obayashi, S.; Hara, Y.; Tanaka, M.; Iriki, A. Modulation of cortical vestibular processing by somatosensory inputs in the posterior insula. Brain Inj. 2013, 27, 1685–1691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearson, K.G.; Collins, D.F. Reversal of the influence of group Ib afferents from plantaris on activity in medial gastrocnemius muscle during locomotor activity. J. Neurophysiol. 1993, 70, 1009–1017. [Google Scholar] [CrossRef] [PubMed]
- Gossard, J.-P.; Brownstone, R.M.; Barajon, I.; Hultborn, H. Transmission in a locomotor-related group Ib pathway from hindlimb extensor muscles in the cat. Exp. Brain Res. 1994, 98, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Takeoka, A.; Vollenweider, I.; Courtine, G.; Arber, S. Muscle Spindle Feedback Directs Locomotor Recovery and Circuit Reorganization after Spinal Cord Injury. Cell 2014, 159, 1626–1639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, W.P.; Akay, T. Stumbling corrective reaction elicited by mechanical and electrical stimulation of the saphenous nerve in walking mice. J. Exp. Biol. 2018, 221, jeb.178095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, A.J.; Croce, K.; Belton, T.; Akay, T.; Jessell, T.M. Balance Control Mediated by Vestibular Circuits Directing Limb Extension or Antagonist Muscle Co- activation. Cell Rep. 2018, 22, 1325–1338. [Google Scholar] [CrossRef] [Green Version]
- Prochazka, A. Proprioceptive Feedback and Movement Regulation. In Comprehensive Physiology; American Physiological Society: Bethesda, MD, USA, 1996; pp. 89–127. [Google Scholar] [CrossRef]
- Pisotta, I.; Molinari, M. Cerebellar contribution to feedforward control of locomotion. Front. Hum. Neurosci. 2014, 8, 1–5. [Google Scholar] [CrossRef]
- Shapiro, D.C.; Zernicke, R.F.; Gregor, R.J.; Diestel, J.D. Evidence for Generalized Motor Programs Using Gait Pattern Analysis. J. Mot. Behav. 1981, 13, 33–47. [Google Scholar] [CrossRef]
- Orlovsky, G.N. Activity of vestibulospinal neurons during locomotion. Brain Res. 1972, 46, 85–98. [Google Scholar] [CrossRef]
- Brownstone, R.M.; Bui, T.V. Spinal interneurons providing input to the final common path during locomotion. Prog. Brain Res. 2010, 187, 81–95. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.J.; Zeng, H. Genetic Approaches to Neural Circuits in the Mouse. Annu. Reveiw Neurosci. 2013, 36, 183–215. [Google Scholar] [CrossRef] [PubMed]
- Heidenreich, M.; Zhang, F. Applications of CRISPR-Cas systems in neuroscience. Nat. Rev. Neurosci. 2016, 17, 36–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kullander, K.; Butt, S.J.B.; Lebret, J.M.; Lundfald, L.; Restrepo, C.E.; Rydström, A.; Klein, R.; Kiehn, O. Role of EphA4 and EphrinB3 in local neuronal circuits that control walking. Science 2003, 299, 1889–1892. [Google Scholar] [CrossRef] [PubMed]
- Akay, T.; Acharya, H.J.; Fouad, K.; Pearson, K.G. Behavioral and electromyographic characterization of mice lacking EphA4 receptors. J. Neurophysiol. 2006, 96, 642–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rybak, I.A.; Shevtsova, N.A.; Kiehn, O. Modelling genetic reorganization in the mouse spinal cord affecting left–right coordination during locomotion. J. Physiol. 2013, 591, 5491–5508. [Google Scholar] [CrossRef]
- Jessell, T.M. Neuronal specification in the spinal cord: Inductive signals and transcriptional codes. Nat. Rev. Genet. 2000, 1, 20–29. [Google Scholar] [CrossRef]
- Bikoff, J.B.; Gabitto, M.I.; Rivard, A.F.; Drobac, E.; Machado, T.A.; Miri, A.; Brenner-Morton, S.; Famojure, E.; Diaz, C.; Alvarez, F.J.; et al. Spinal Inhibitory Interneuron Diversity Delineates Variant Motor Microcircuits Article Spinal Inhibitory Interneuron Diversity Delineates Variant Motor Microcircuits. Cell 2016, 165, 207–219. [Google Scholar] [CrossRef] [Green Version]
- Stepien, A.E.; Arber, S. Probing the Locomotor Conundrum: Descending the “V” Interneuron Ladder. Neuron 2008, 60, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Jensen, P.; Dymecki, S.M. Essentials of Recombinase-Based Genetic Fate Mapping in Mice. In Mouse Molecular Embryology; Lewandoski, M., Ed.; Springer: New York, NY, USA, 2014. [Google Scholar] [CrossRef]
- Gosgnach, S.; Lanuza, G.M.; Butt, S.J.B.; Saueressig, H.; Zhang, Y.; Velasquez, T.; Riethmacher, D.; Callaway, E.M.; Kiehn, O.; Goulding, M. V1 spinal neurons regulate the speed of vertebrate locomotor outputs. Nature 2006, 440, 215–219. [Google Scholar] [CrossRef]
- Chopek, J.W.; Nascimento, F.; Beato, M.; Brownstone, R.M.; Zhang, Y. Sub-populations of Spinal V3 Interneurons Form Focal Modules of Layered Pre-motor Microcircuits. Cell Rep. 2018, 25, 146–156.e3. [Google Scholar] [CrossRef] [Green Version]
- Cregg, J.M.; Leiras, R.; Montalant, A.; Wanken, P.; Wickersham, I.R.; Kiehn, O. Brainstem neurons that command mammalian locomotor asymmetries. Nat. Neurosci. 2020, 23. [Google Scholar] [CrossRef] [PubMed]
- Usseglio, G.; Gatier, E.; Heuzé, A.; Hérent, C.; Bouvier, J. Control of Orienting Movements and Locomotion by Projection-Defined Subsets of Brainstem V2a Neurons. Curr. Biol. 2020, 30, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Zampieri, N.; Jessell, T.M.; Murray, A.J. Mapping sensory circuits by anterograde transsynaptic transfer of recombinant rabies virus. Neuron 2014, 81, 766–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petitpré, C.; Bourien, J.; Wu, H.; Diuba, A.; Petitpre, C.; Puel, J.-L.; Lallemend, F. Genetic and functional diversity of primary auditory afferents. Curr. Opin. Physiol. 2020, 18, 85–94. [Google Scholar] [CrossRef]
- de Nooij, J.C.; Doobar, S.; Jessell, T.M. Etv1 Inactivation Reveals Proprioceptor Subclasses that Reflect the Level of NT3 Expression in Muscle Targets. Neuron 2013, 77, 1055–1068. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.; Schieren, I.; Qian, Y.; Zhang, C.; Jessell, T.M.; de Nooij, J.C. A Role for Sensory end Organ-Derived Signals in Regulating Muscle Spindle Proprioceptor Phenotype. J. Neurosci. 2019, 39, 4252–4267. [Google Scholar] [CrossRef] [Green Version]
- Lunde, A.; Okaty, B.W.; Dymecki, S.M.; Glover, J.C. Molecular Profiling Defines Evolutionarily Conserved Transcription Factor Signatures of Major Vestibulospinal Neuron Groups. eNeuro 2019, 6. [Google Scholar] [CrossRef]
- Chagnaud, B.P.; Engelmann, J.; Fritzsch, B.; Glover, J.C. Sensing External and Self-Motion with Hair Cells: A Comparison of the Lateral Line and Vestibular Systems from a Developmental and Evolutionary Perspective. Brain. Behav. Evol. 2017, 90, 98–116. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akay, T.; Murray, A.J. Relative Contribution of Proprioceptive and Vestibular Sensory Systems to Locomotion: Opportunities for Discovery in the Age of Molecular Science. Int. J. Mol. Sci. 2021, 22, 1467. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031467
Akay T, Murray AJ. Relative Contribution of Proprioceptive and Vestibular Sensory Systems to Locomotion: Opportunities for Discovery in the Age of Molecular Science. International Journal of Molecular Sciences. 2021; 22(3):1467. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031467
Chicago/Turabian StyleAkay, Turgay, and Andrew J. Murray. 2021. "Relative Contribution of Proprioceptive and Vestibular Sensory Systems to Locomotion: Opportunities for Discovery in the Age of Molecular Science" International Journal of Molecular Sciences 22, no. 3: 1467. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031467