
Biosensor-Based Directed Evolution of Methanol Dehydrogenase from Lysinibacillus xylanilyticus

 , , and
, , and
Abstract
:1. Introduction
2. Results and Discussion
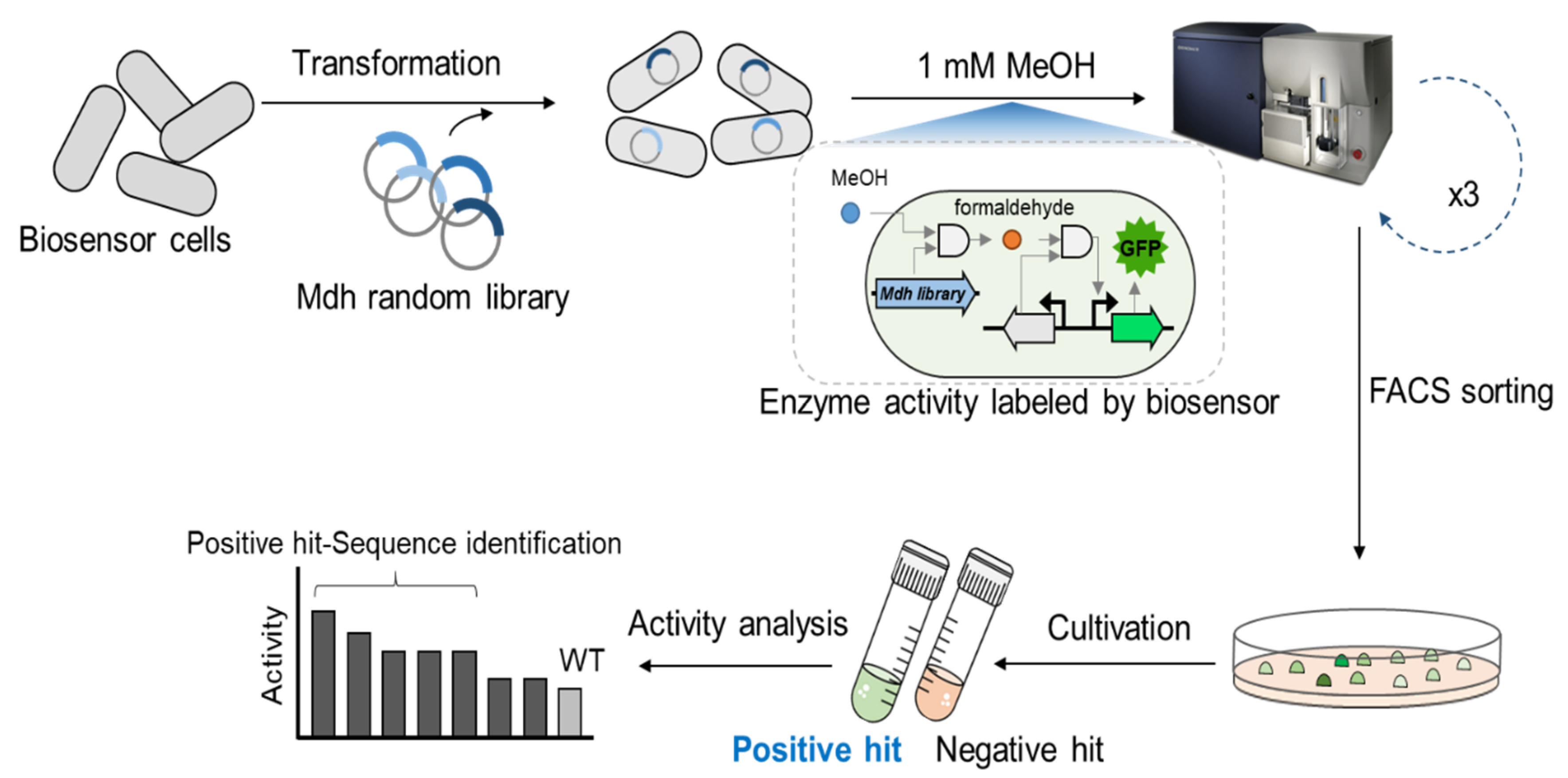
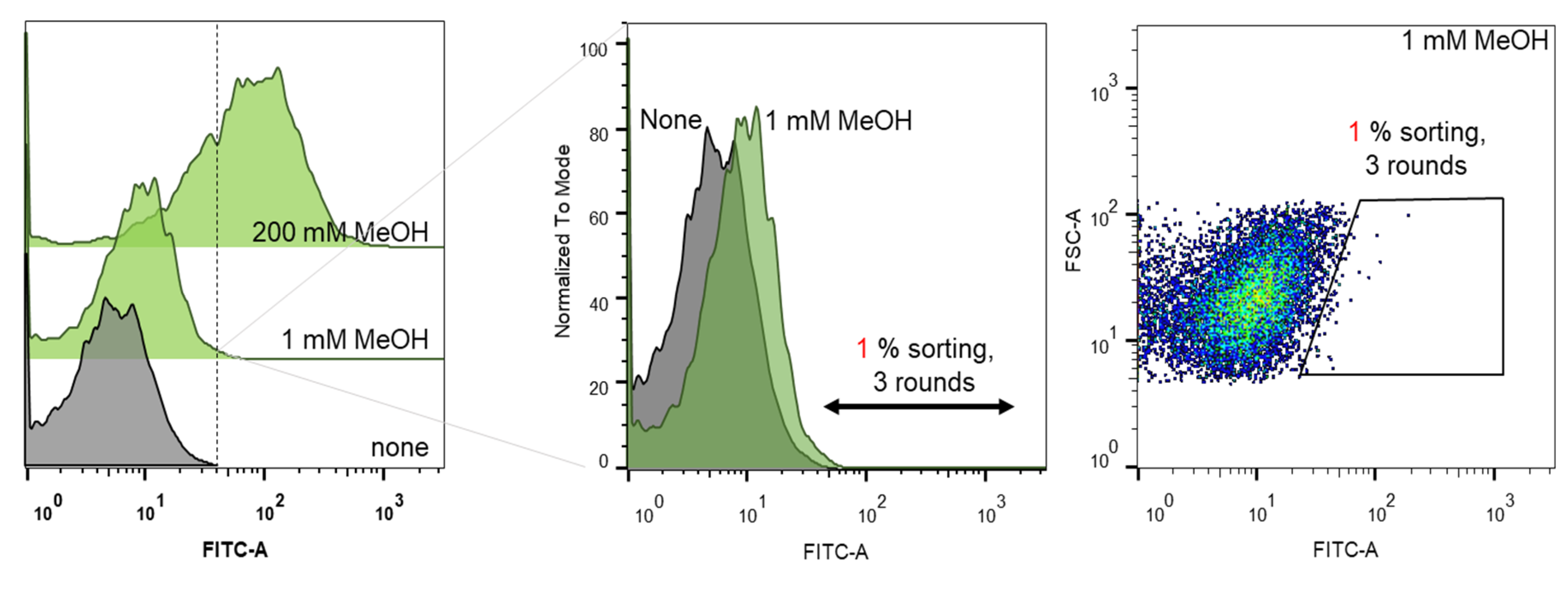
2.1. Biosensor-Based Screening of the Lxmdh Library
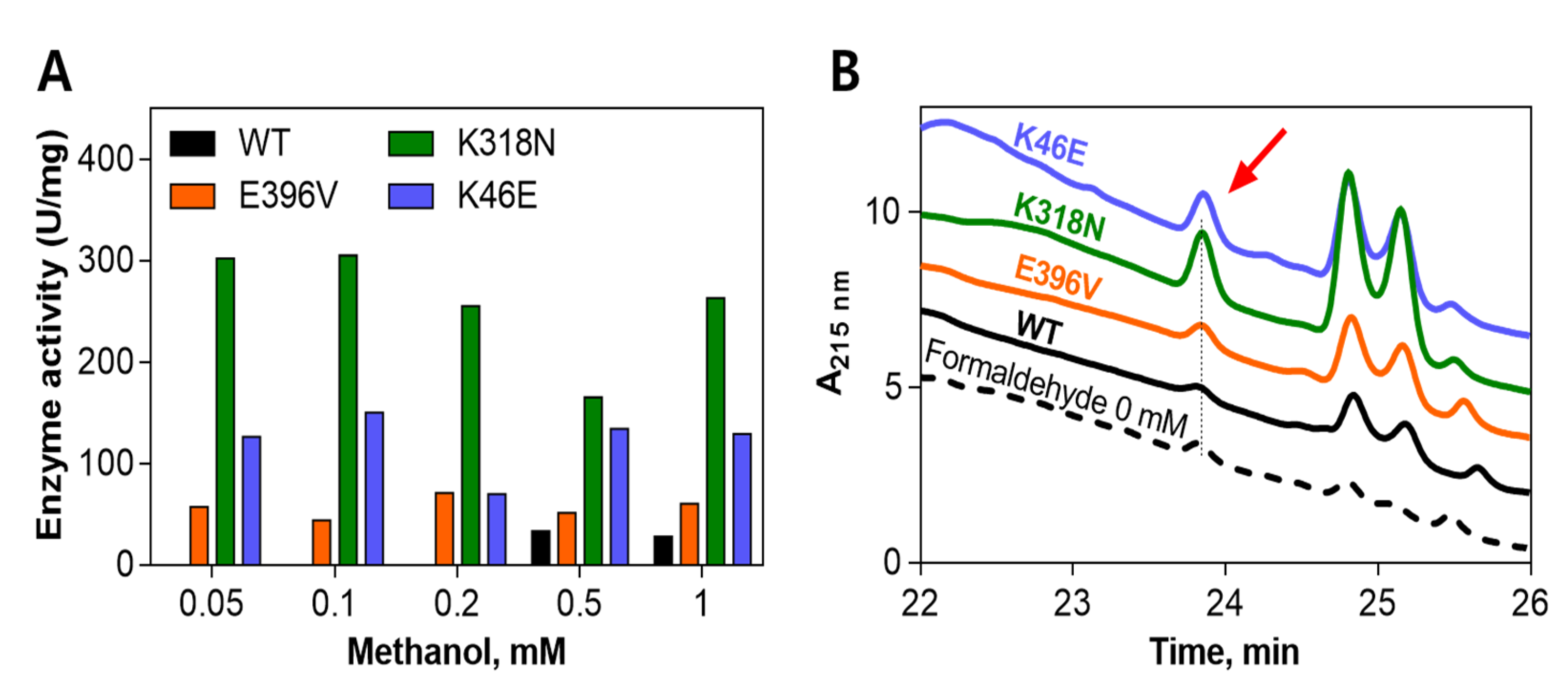
2.2. Comparison of Enzyme Activity between the Wild-Type and Its Mutants
2.3. Enzyme Activity by Concentration of Generated Formaldehyde with Respect to Methanol Concentration
2.4. Kinetic Parameters of the Wild-Type and Mutant Enzymes
3. Materials and Methods
3.1. Chemicals and Materials
3.2. Strains and Plasmids
3.3. Construction of Lxmdh Mutant Library
3.4. Library Screening Using Flow Cytometry
3.5. Screening the Lxmdh Library by Liquid-Phase Fluorescence Analyses
3.6. Purification of Lxmdh
3.7. Comparison of Enzyme Activity between the Wild-Type and Its Mutants
3.8. Enzyme Activity by the Concentration of Formaldehyde Generated with Respect to Methanol Concentration
3.9. Determination of Kinetic Parameters of Wild-Type and Mutant Enzymes
3.10. Analytical Methods
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Quayle, J.R.; Ferenci, T. Evolutionary aspects of autotrophy. Microbiol. Rev. 1978, 42, 251–273. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhang, T.; Wu, S.; Wu, M.; Xin, F.; Dong, W.; Ma, J.; Zhang, M.; Jiang, M. Guidance for engineering of synthetic methylotrophy based on methanol metabolism in methylotrophy. RSC Adv. 2017, 7, 4083–4091. [Google Scholar] [CrossRef] [Green Version]
- Müller, J.E.; Meyer, F.; Litsanov, B.; Kiefer, P.; Potthoff, E.; Heux, S.; Quax, W.J.; Wendisch, V.F.; Brautaset, T.; Portais, J.-C.; et al. Engineering Escherichia coli for methanol conversion. Metab. Eng. 2015, 28, 190–201. [Google Scholar] [CrossRef]
- Wu, T.-Y.; Chen, C.-T.; Liu, J.T.-J.; Bogorad, I.W.; Damoiseaux, R.; Liao, J.C. Characterization and evolution of an activator-independent methanol dehydrogenase from Cupriavidus necator N-1. Appl. Microbiol. Biotechnol. 2016, 100, 4969–4983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitaker, W.B.; Jones, J.A.; Bennett, R.K.; Gonzalez, J.E.; Vernacchio, V.R.; Collins, S.M.; Palmer, M.A.; Schmidt, S.; Antoniewicz, M.R.; Koffas, M.A.G.; et al. Engineering the biological conversion of methanol to specialty chemicals in Escherichia coli. Metab. Eng. 2017, 39, 49–59. [Google Scholar] [CrossRef]
- Hektor, H.J.; Kloosterman, H.; Dijkhuizen, L. Identification of a Magnesium-dependent NAD(P)(H)-binding Domain in the Nicotinoprotein Methanol Dehydrogenase from Bacillus methanolicus. J. Biol. Chem. 2002, 277, 46966–46973. [Google Scholar] [CrossRef] [Green Version]
- Krog, A.; Heggeset, T.M.B.; Müller, J.E.N.; Kupper, C.E.; Schneider, O.; Vorholt, J.A.; Ellingsen, T.E.; Brautaset, T. Methylotrophic Bacillus methanolicus Encodes Two Chromosomal and One Plasmid Born NAD+ Dependent Methanol Dehydrogenase Paralogs with Different Catalytic and Biochemical Properties. PLoS ONE 2013, 8, e59188. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-Y.; Park, S.-H.; Oh, S.-H.; Kwon, K.K.; Kim, S.-J.; Choi, M.; Rha, E.; Lee, H.; Lee, D.-H.; Sung, B.H.; et al. Discovery and Biochemical Characterization of a Methanol Dehydrogenase from Lysinibacillus xylanilyticus. Front. Bioeng. Biotechnol. 2020, 8, 67. [Google Scholar] [CrossRef]
- Kwon, K.K.; Lee, D.-H.; Kim, S.J.; Choi, S.-L.; Rha, E.; Yeom, S.-J.; Subhadra, B.; Lee, J.; Jeong, K.J.; Lee, S.-G. Evolution of enzymes with new specificity by high-throughput screening using DmpR-based genetic circuits and multiple flow cytometry rounds. Sci. Rep. 2018, 8, 2659. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.-L.; Rha, E.; Lee, S.J.; Kim, H.; Kwon, K.; Jeong, Y.-S.; Rhee, Y.H.; Song, J.J.; Kim, H.-S.; Lee, S.-G. Toward a Generalized and High-throughput Enzyme Screening System Based on Artificial Genetic Circuits. ACS Synth. Biol. 2014, 3, 163–171. [Google Scholar] [CrossRef]
- Kim, H.; Rha, E.; Seong, W.; Yeom, S.-J.; Lee, D.-H.; Lee, S.-G. A Cell–Cell Communication-Based Screening System for Novel Microbes with Target Enzyme Activities. ACS Synth. Biol. 2016, 5, 1231–1238. [Google Scholar] [CrossRef] [PubMed]
- Woolston, B.M.; Roth, T.; Kohale, I.; Liu, D.R.; Stephanopoulos, G. Development of a formaldehyde biosensor with application to synthetic methylotrophy. Biotechnol. Bioeng. 2018, 115, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Yeom, S.-J.; Kim, M.; Kwon, K.K.; Fu, Y.; Rha, E.; Park, S.-H.; Lee, H.; Kim, H.; Lee, D.-H.; Kim, D.-M.; et al. A synthetic microbial biosensor for high-throughput screening of lactam biocatalysts. Nat. Commun. 2018, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.; Yeom, S.-J.; Kwon, K.K.; Hwang, J.; Kim, H.; Woo, E.-J.; Lee, D.-H.; Lee, S.-G. Structural and functional analyses of the cellulase transcription regulator CelR. FEBS Lett. 2018, 592, 2776–2785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, K.K.; Yeom, S.-J.; Lee, D.-H.; Jeong, K.J.; Lee, S.-G. Development of a novel cellulase biosensor that detects crystalline cellulose hydrolysis using a transcriptional regulator. Biochem. Biophys. Res. Commun. 2018, 495, 1328–1334. [Google Scholar] [CrossRef]
- Yang, X.A.; Zweifach, A. Temperature-Dependent Expression of a CFP-YFP FRET Diacylglycerol Sensor Enables Multiple-Read Screening for Compounds That Affect C1 Domains. SLAS Discov. Adv. Life Sci. RD 2019, 24, 682–692. [Google Scholar] [CrossRef] [PubMed]
- Dai, B.; Wang, L.; Wang, Y.; Yu, G.; Huang, X. Single-Cell Nanometric Coating Towards Whole-Cell-Based Biodevices and Biosensors. Chem. Sel. 2018, 3, 7208–7221. [Google Scholar] [CrossRef]
- Ganesh, I.; Vidhya, S.; Eom, G.T.; Hong, S.H. Construction of Methanol-Sensing Escherichia coli by the Introduction of a Paracoccus denitrificans MxaY-Based Chimeric Two-Component System. J. Microbiol. Biotechnol. 2017, 27, 1106–1111. [Google Scholar] [CrossRef]
- Lee, J.-Y.; Sung, B.H.; Oh, S.-H.; Kwon, K.K.; Lee, H.; Kim, H.; Lee, D.-H.; Yeom, S.-J.; Lee, S.-G. C1 Compound Biosensors: Design, Functional Study, and Applications. Int. J. Mol. Sci. 2019, 20, 2253. [Google Scholar] [CrossRef] [Green Version]
- Ochsner, A.M.; Müller, J.E.; Mora, C.A.; Vorholt, J.A. In vitro activation of NAD-dependent alcohol dehydrogenases by Nudix hydrolases is more widespread than assumed. FEBS Lett. 2014, 588, 2993–2999. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.G.; Han, G.H.; Kim, D.; Choi, J.S.; Kim, S.W. Comparative analysis of two types of methanol dehydrogenase from Methylophaga aminisulfidivorans MPT grown on methanol. J. Basic Microbiol. 2012, 52, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, K.; Bujons, J.; Joglar, J.; Charnock, S.J.; De María, P.D.; Fessner, W.-D.; Clapés, P. Combining Aldolases and Transaminases for the Synthesis of 2-Amino-4-hydroxybutanoic Acid. ACS Catal. 2017, 7, 1707–1711. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | kcat (min−1) | KM (mM) | kcat/KM (min−1.mM−1) (Relative Catalytic Efficiency) |
|---|---|---|---|
| Wild-type | 2.0 ± 0.2 | 1.311 ± 0.406 | 1.526 (1) |
| E396V | 1.2 ± 0.1 | 0.010 ± 0.003 | 120 (79) |
| E396V+K318N | 1.3 ± 0.1 | 0.233 ± 0.107 | 5.579 (4) |
| K318N | 1.6 ± 0.3 | 0.046 ± 0.072 | 34.78 (23) |
| K46E | 1.7 ± 0.2 | 0.372 ± 0.231 | 4.570 (3) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Le, T.-K.; Ju, S.-B.; Lee, H.; Lee, J.-Y.; Oh, S.-H.; Kwon, K.-K.; Sung, B.-H.; Lee, S.-G.; Yeom, S.-J. Biosensor-Based Directed Evolution of Methanol Dehydrogenase from Lysinibacillus xylanilyticus. Int. J. Mol. Sci. 2021, 22, 1471. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031471
Le T-K, Ju S-B, Lee H, Lee J-Y, Oh S-H, Kwon K-K, Sung B-H, Lee S-G, Yeom S-J. Biosensor-Based Directed Evolution of Methanol Dehydrogenase from Lysinibacillus xylanilyticus. International Journal of Molecular Sciences. 2021; 22(3):1471. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031471
Chicago/Turabian StyleLe, Thien-Kim, Su-Bin Ju, Hyewon Lee, Jin-Young Lee, So-Hyung Oh, Kil-Koang Kwon, Bong-Hyun Sung, Seung-Goo Lee, and Soo-Jin Yeom. 2021. "Biosensor-Based Directed Evolution of Methanol Dehydrogenase from Lysinibacillus xylanilyticus" International Journal of Molecular Sciences 22, no. 3: 1471. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031471