
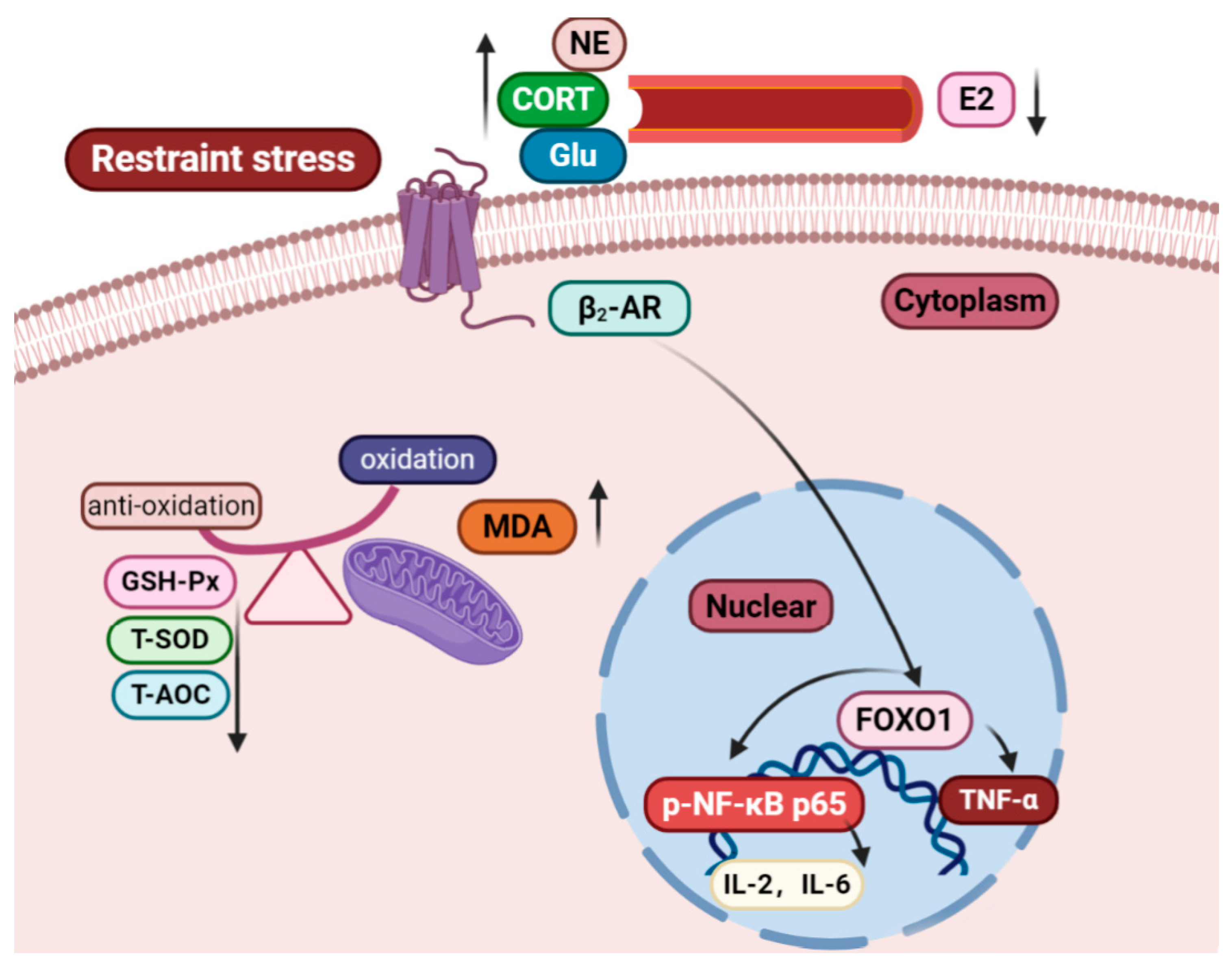
The Role of the FOXO1/β2-AR/p-NF-κB p65 Pathway in the Development of Endometrial Stromal Cells in Pregnant Mice under Restraint Stress


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
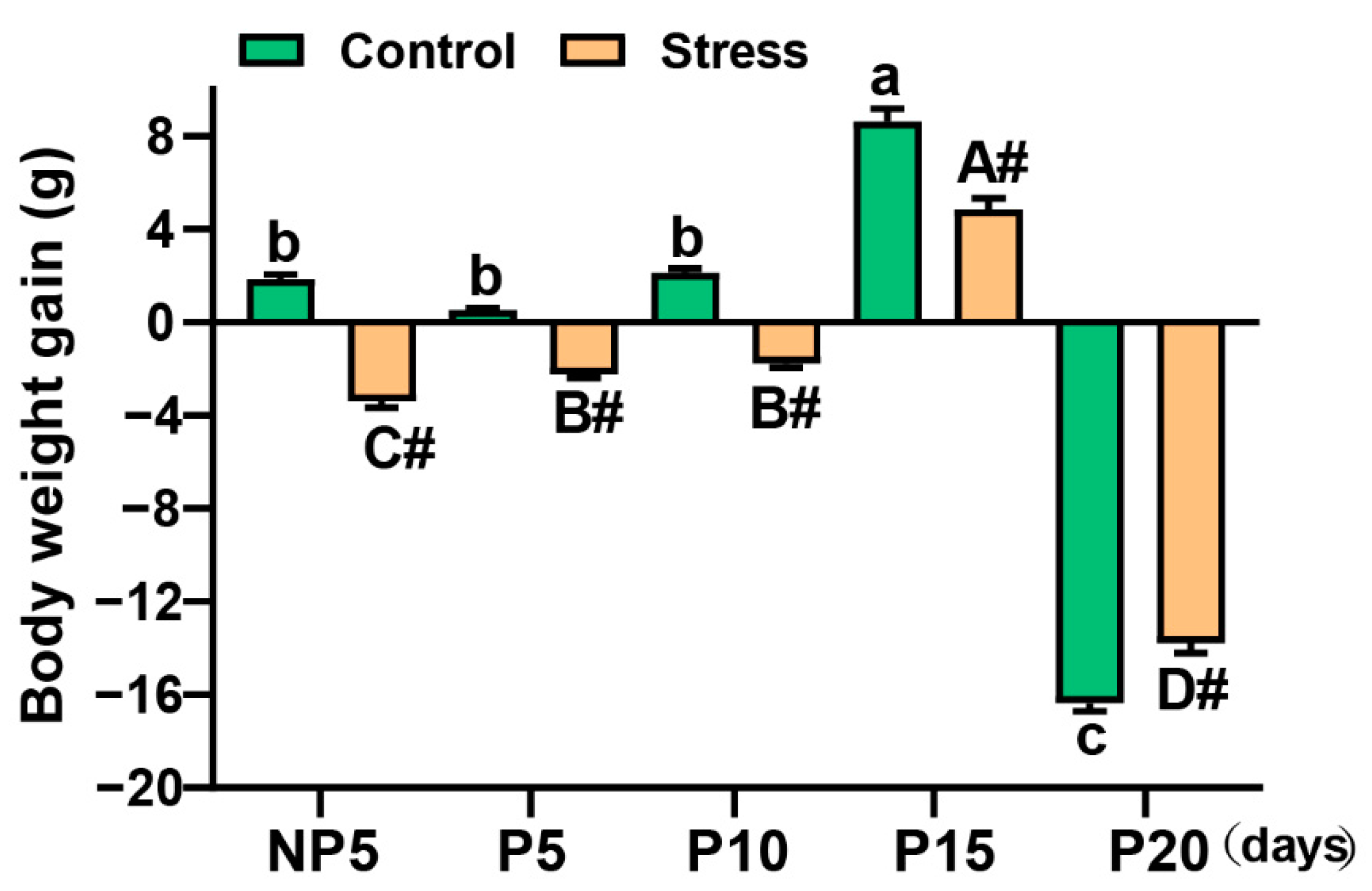
2.1. Effect of Restraint Stress on Body Weight in Pregnant Mice
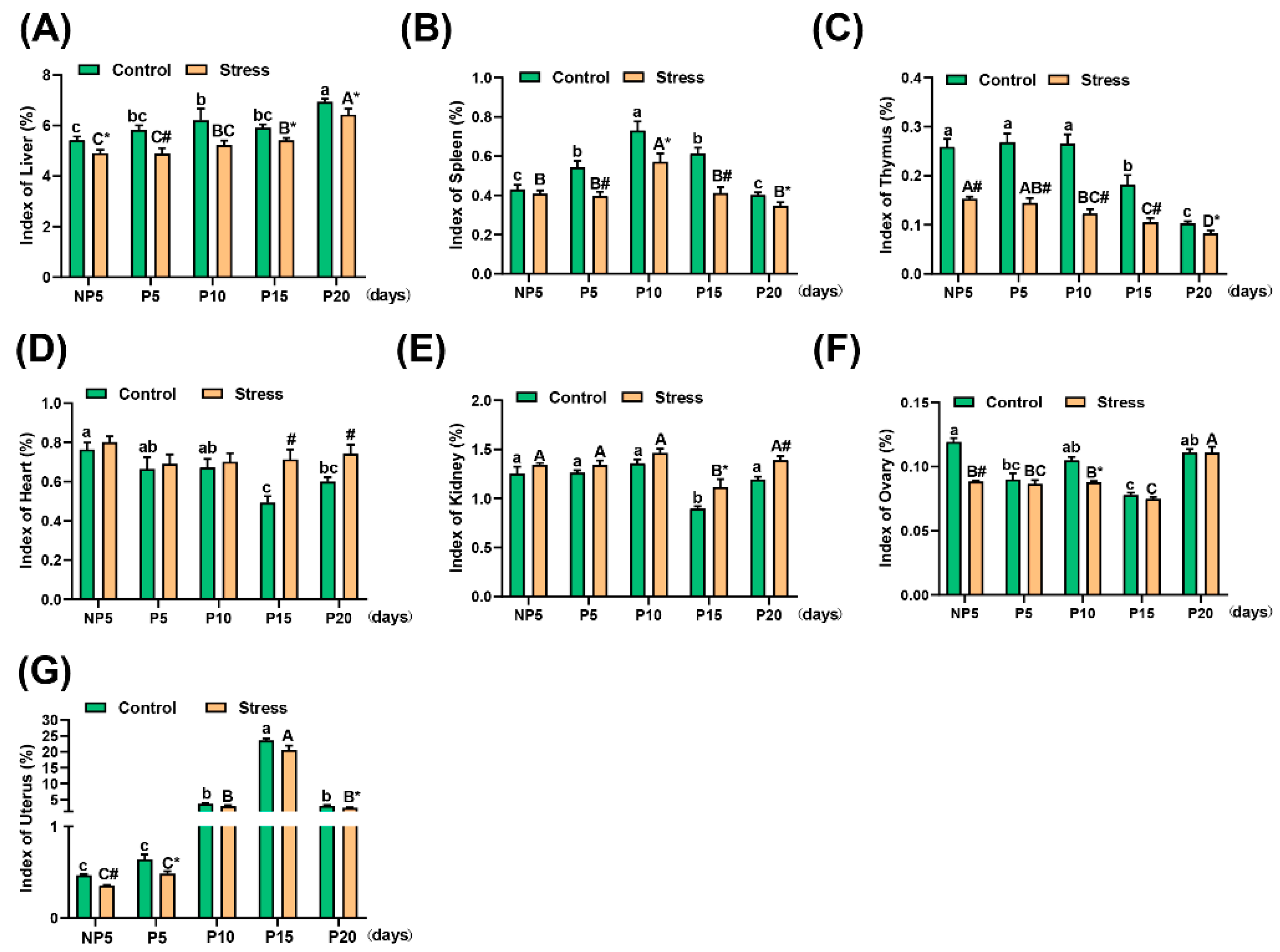
2.2. Effect of Restraint Stress on Organ Index in Pregnant Mice
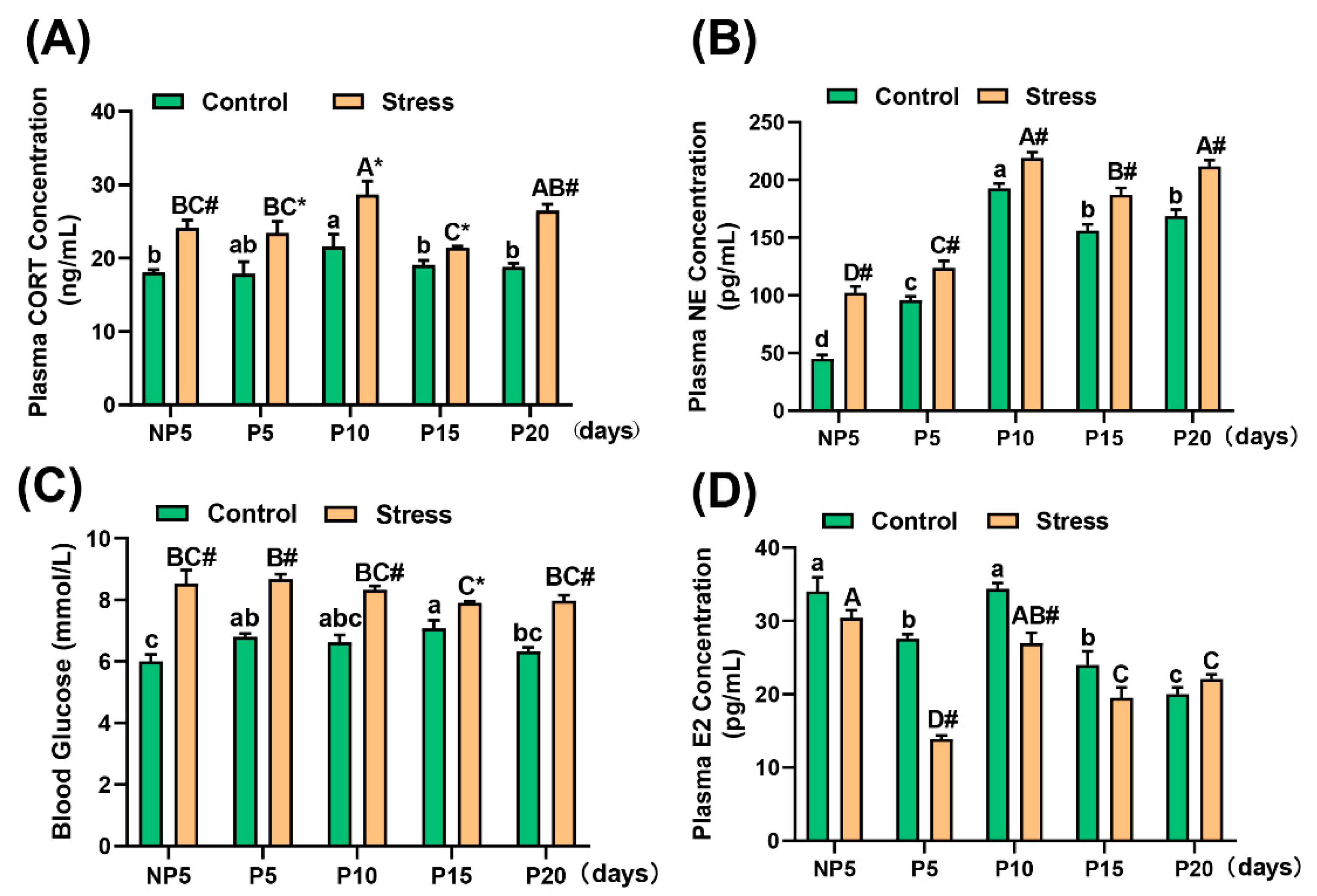
2.3. Effect of Restraint Stress on Plasma Hormone Levels in Pregnant Mice
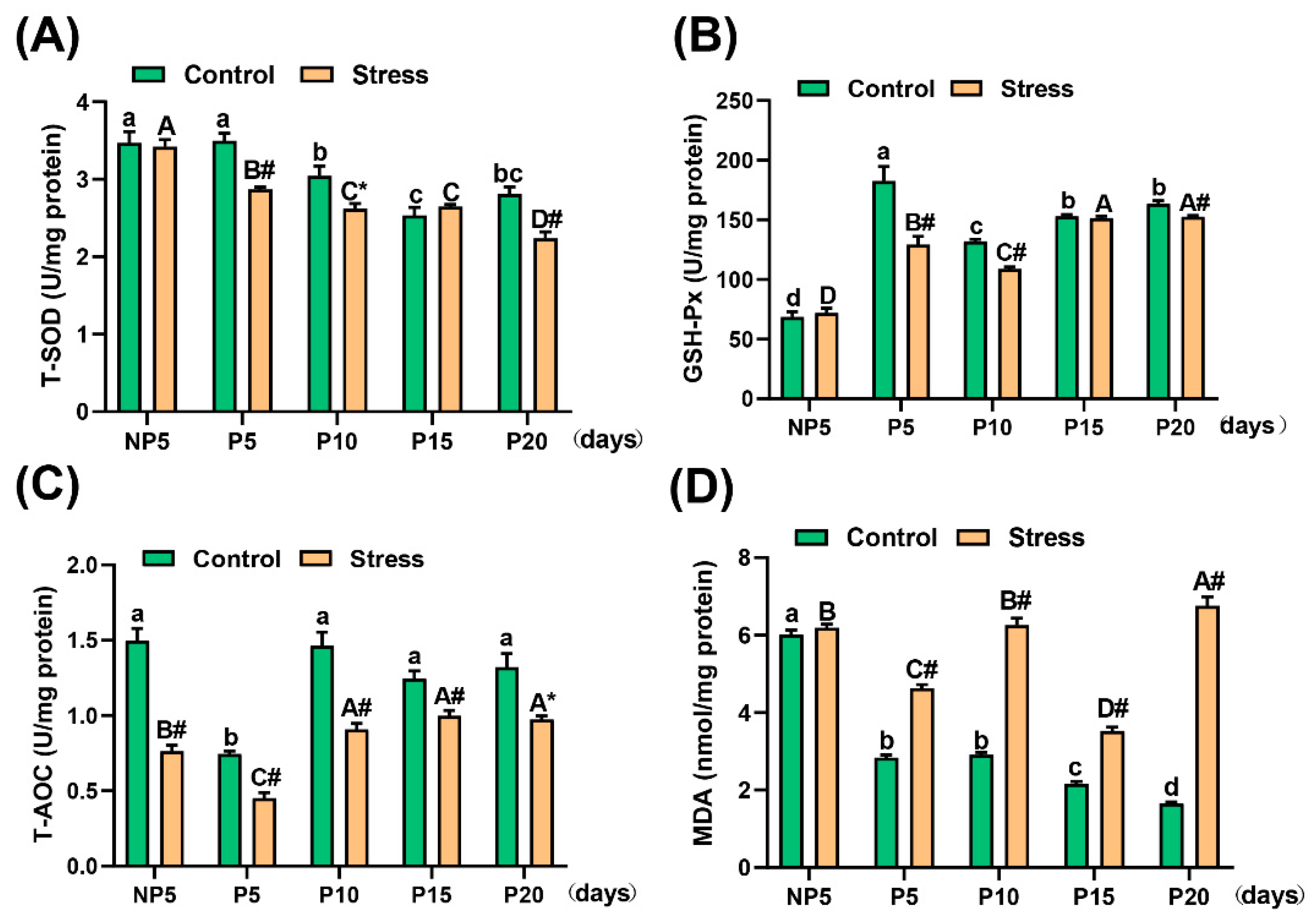
2.4. Effect of Restraint Stress on Antioxidative Ability in the Uterus of Pregnant Mice
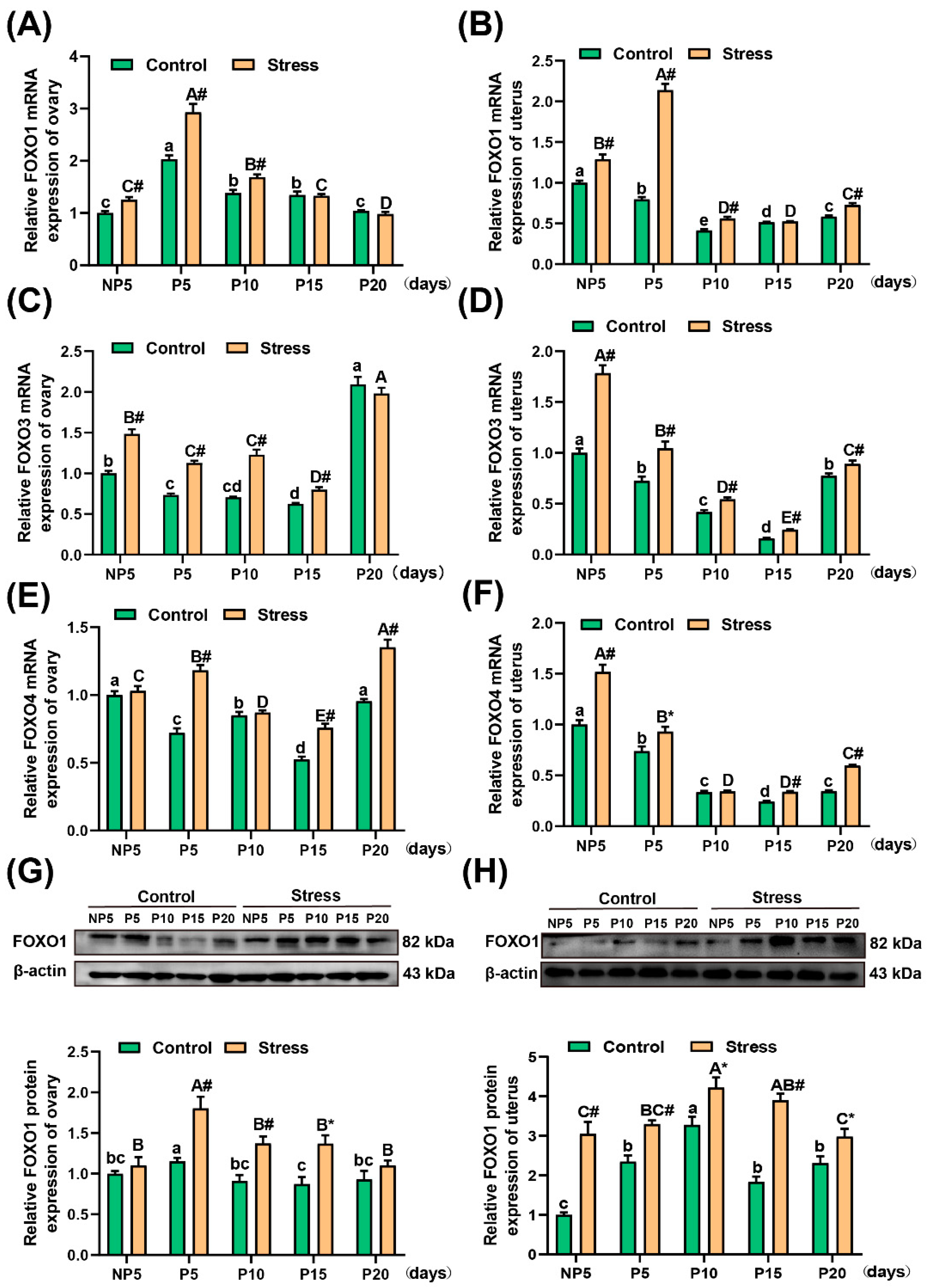
2.5. Effect of Restraint Stress on FOXO1, FOXO3, and FOXO4 mRNA Levels and FOXO1 Protein in the Uterus and Ovary of Pregnant Mice
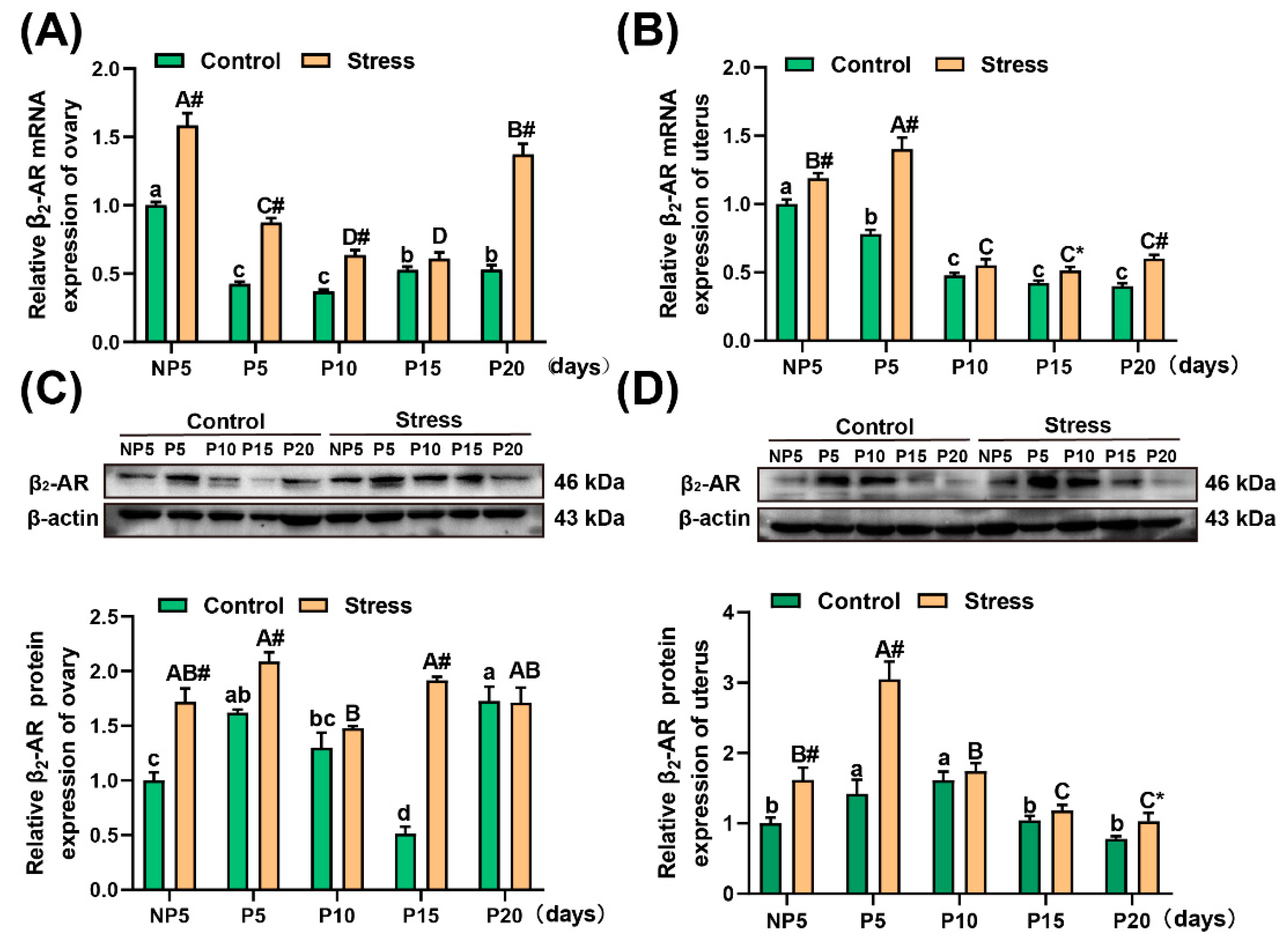
2.6. Effect of Restraint Stress on β2-AR Levels in the Uterus and Ovary of Pregnant Mice
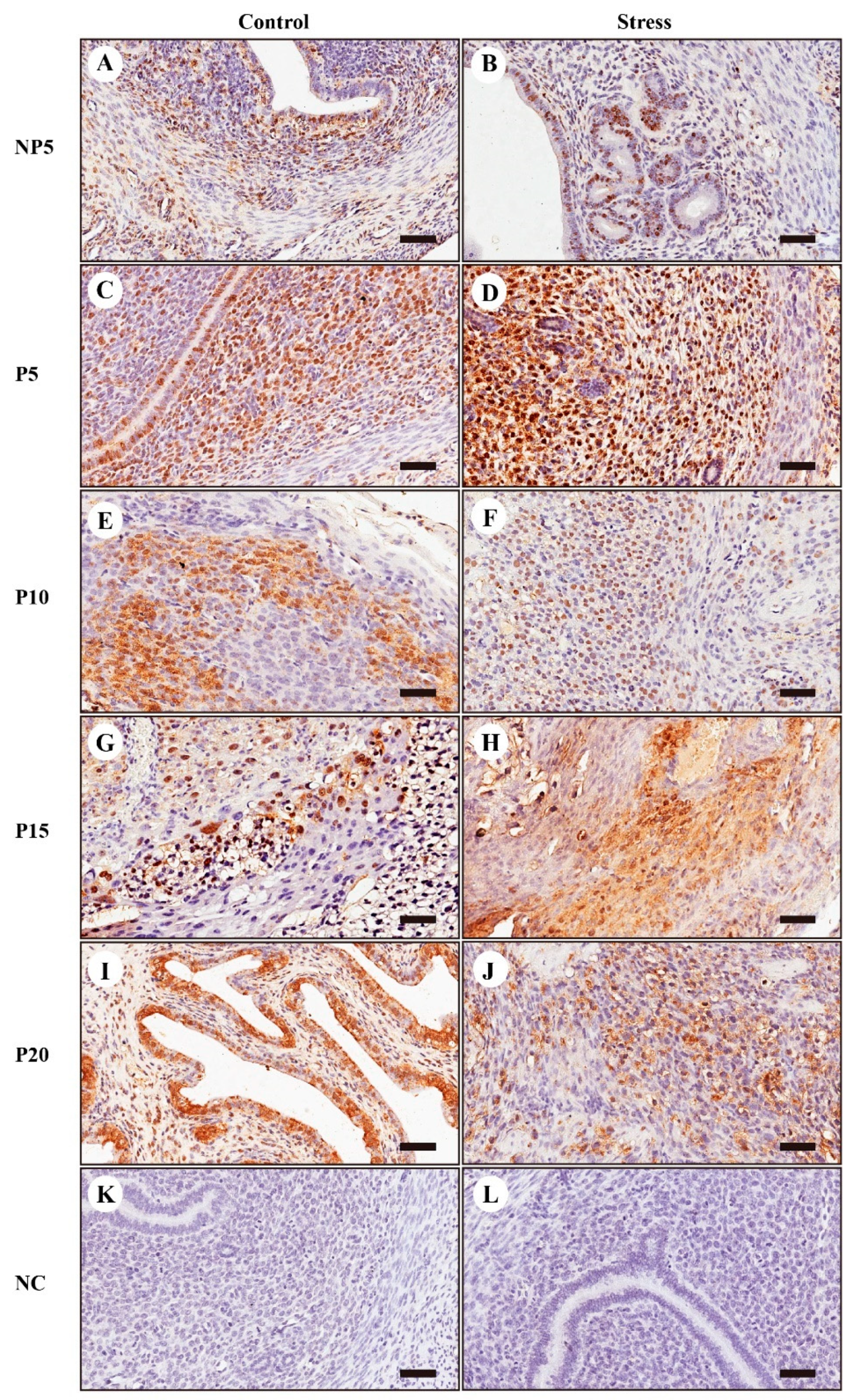
2.7. Immunohistochemical Localization of FOXO1
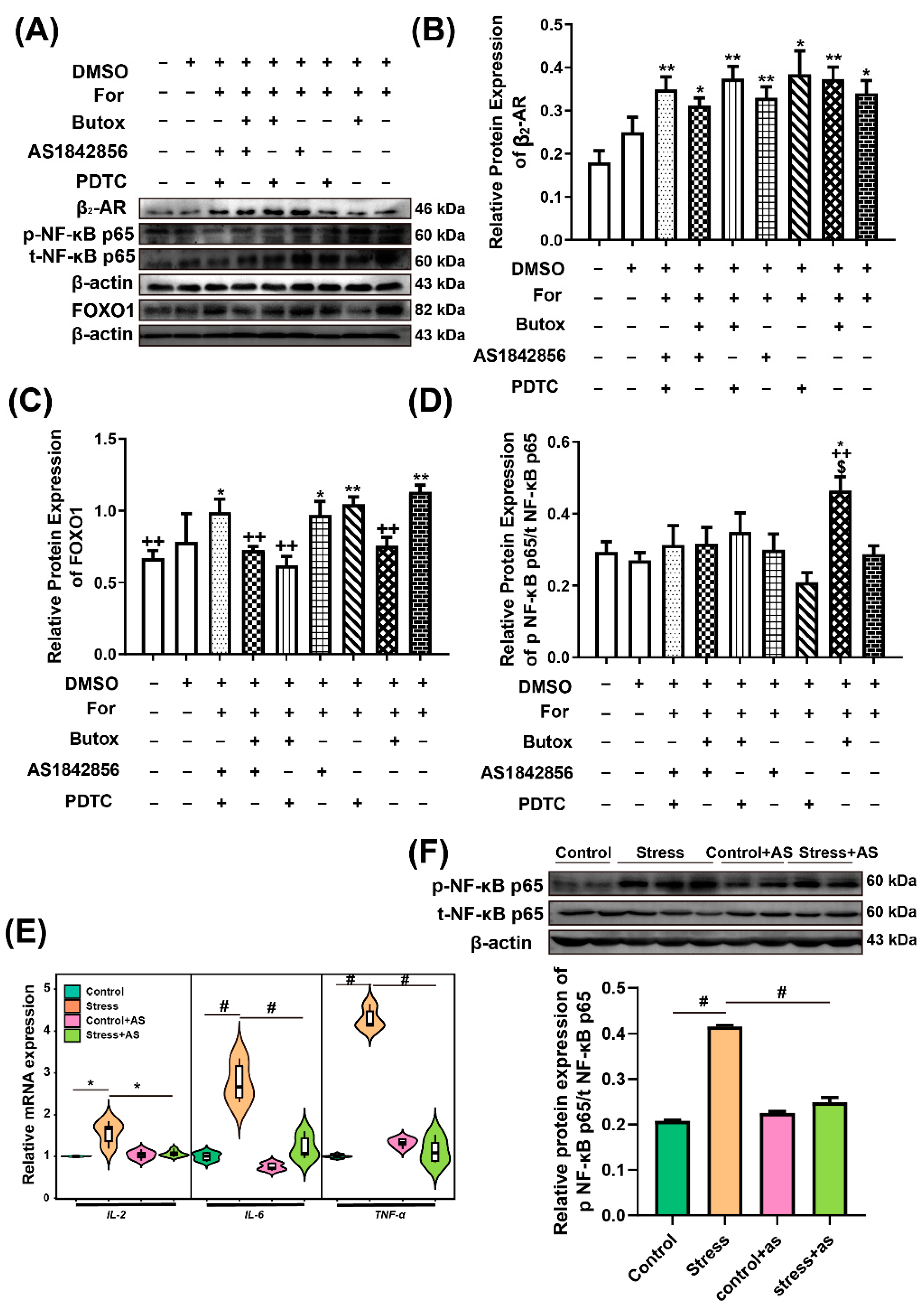
2.8. Relativity Analysis among FOXO1, p-NF-κB p65, and β2-AR
3. Discussion
3.1. Restraint Stress Disturbs Neuroendocrine Signalling in Pregnant Mice
3.2. The FOXO Family is Involved in the Oxidative Stress Induced by Restraint Stress
3.3. Restraint Stress Promotes the β2-AR Pathway
3.4. FOXO1 Mediates the β2-AR and p-NF-κB p65 Signalling Pathways
4. Materials and Methods
4.1. Animal Treatments
4.2. Plasma and Tissue Preparations
4.3. Quantitative Real-Time Polymerase Chain Reaction
4.4. Measurement of Plasma CORT, NE, Blood Glucose, and E2 Concentration
4.5. Measurements of Oxidative Stress-Related Enzymes
4.6. Western Blotting
4.7. Immunohistochemistry
4.8. Cell Culture with Drug Treatment
4.9. Statistical Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BWG | Body weight gain |
| Butox | Butoxamine hydrochloride |
| cAMP | Cyclic adenosine monophosphate |
| CORT | Corticosterone |
| CRH | Corticotropin-releasing hormone |
| HBSS | D’Hanks balanced saline solution |
| DAB | Diaminobenzidine |
| ESCs | Endometrial stromal cells |
| EPI | Epinephrine |
| FOXO | Forkhead transcription factor class O |
| For | Formoterol hemifumarate |
| GSH-Px | Glutathione peroxidase |
| GnRH | Gonadotropin releasing hormone |
| HRP | Horseradish peroxidase |
| HPA | Hypothalamic-pituitary-adrenal |
| HPG | Hypothalamic-pituitary-gonadal |
| IκB | Inhibitor of NF-κB |
| InsRes | Insulin resistance |
| IOD | Integral optical density |
| IL-18 | Interleukin 18 |
| IL-6 | Interleukin-6 |
| IL-8 | Interleukin 8 |
| IL-1β | Interleukin -1 beta |
| IUGR | Intrauterine growth restriction |
| ISO | Isoproterenol |
| MDA | Malondialdehyde |
| NE | Norepinephrin |
| NP1 | Non-pregnancy Day 1 |
| NP5 | Non-pregnancy Day 5 |
| NF-κB | Nuclear factor-kappa B |
| E2 | Oestradiol |
| ANOVA | One-way analysis of variance |
| GSSG | Oxidized glutathione disulphide |
| P1 | Pregnancy Day 1 |
| P10 | Pregnancy Day 10 |
| P15 | Pregnancy Day 15 |
| P20 | Pregnancy Day 20 |
| P11 | Pregnancy Day 11 |
| P16 | Pregnancy Day 16 |
| P5 | Pregnancy Day 5 |
| P6 | Pregnancy Day 6 |
| PI3K-AKT | Phosphatidylinositol 3-kinase-serine/threonine kinase |
| Pomc | Pro-opiomelanocortin |
| PDTC | Pyrrolidine dithiocarbamate ammonium |
| qRT-PCR | Quantitative real-time polymerase chain reaction |
| ROS | Reactive oxygen species |
| RT-PCR | Reverse transcription-polymerase chain reaction |
| STAT3 | Signal transducers and activators of transcription 3 |
| siRNA | Small interfering RNA |
| SAS | Sympathetic-adrenal medullary system |
| SDS-PAGE | Sodium dodecyl sulfate polyacrylamide gel electrophoresis |
| SEM | Standard error of mean |
| TNF-α | Tumor necrosis factor-alpha |
| T-AOC | Total antioxidant capacity |
| T-SOD | Total superoxide dismutase |
| uNK | Uterus natural killer cell |
| β2-AR | β2-adrenergic receptor |
References
- Sandman, C.A.; Davis, E.P.; Buss, C.; Glynn, L.M. Exposure to prenatal psychobiological stress exerts programming influences on the mother and her fetus. Neuroendocrinology 2012, 95, 7–21. [Google Scholar] [CrossRef] [Green Version]
- Veru, F.; Laplante, D.P.; Luheshi, G.; King, S. Prenatal maternal stress exposure and immune function in the offspring. Stress 2014, 17, 133–148. [Google Scholar] [CrossRef]
- Hantsoo, L.; Kornfield, S.; Anguera, M.C.; Epperson, C.N. Inflammation: A Proposed Intermediary between Maternal Stress and Offspring Neuropsychiatric Risk. Biol. Psychiatry 2019, 85, 97–106. [Google Scholar] [CrossRef]
- Murison, R.; Overmier, J.B. Some psychosomatic causal factors of restraint-in-water stress ulcers. Physiol. Behav. 1993, 53, 577–581. [Google Scholar] [CrossRef]
- Liu, G.; Dong, Y.; Wang, Z.; Cao, J.; Chen, Y. Restraint stress alters immune parameters and induces oxidative stress in the mouse uterus during embryo implantation. Stress 2014, 17, 494–503. [Google Scholar] [CrossRef]
- Liu, G.; Dong, Y.; Wang, Z.; Cao, J.; Chen, Y. Restraint stress delays endometrial adaptive remodeling during mouse embryo implantation. Stress 2015, 18, 699–709. [Google Scholar] [CrossRef]
- Chrousos, G.P.; Torpy, D.J.; Gold, P.W. Interactions between the hypothalamic-pituitary-adrenal axis and the female reproductive system: Clinical implications. Ann. Intern. Med. 1998, 129, 229–240. [Google Scholar] [CrossRef]
- de Kloet, E.R.; Joels, M.; Holsboer, F. Stress and the brain: From adaptation to disease. Nat. Rev. Neurosci. 2005, 6, 463–475. [Google Scholar]
- Chen, Q.; Zhang, Y.; Peng, H.; Lei, L.; Kuang, H.; Zhang, L.; Ning, L.; Cao, Y.; Duan, E. Transient {beta}2-adrenoceptor activation confers pregnancy loss by disrupting embryo spacing at implantation. J. Biol. Chem. 2011, 286, 4349–4356. [Google Scholar] [CrossRef] [Green Version]
- Burton, G.J.; Jauniaux, E. Oxidative stress. Best Pract. Res. Clin. Obstet. Gynaecol. 2011, 25, 287–299. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.; Allamaneni, S.S. Role of free radicals in female reproductive diseases and assisted reproduction. Reprod. Biomed. Online 2004, 9, 338–347. [Google Scholar] [CrossRef]
- Gupta, S.; Agarwal, A.; Krajcir, N.; Alvarez, J.G. Role of oxidative stress in endometriosis. Reprod. Biomed. Online 2006, 13, 126–134. [Google Scholar] [CrossRef]
- Abolaji, A.O.; Ojo, M.; Afolabi, T.T.; Arowoogun, M.D.; Nwawolor, D.; Farombi, E.O. Protective properties of 6-gingerol-rich fraction from Zingiber officinale (Ginger) on chlorpyrifos-induced oxidative damage and inflammation in the brain, ovary and uterus of rats. Chem. Biol. Interact. 2017, 270, 15–23. [Google Scholar] [CrossRef]
- Gupta, S.; Agarwal, A.; Banerjee, J.; Alvarez, J.G. The role of oxidative stress in spontaneous abortion and recurrent pregnancy loss: A systematic review. Obstet. Gynecol. Surv. 2007, 62, 335–347, quiz 353–334. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Wang, Z.; Cao, J.; Chen, Y.; Dong, Y. A novel and compact review on the role of oxidative stress in female reproduction. Reprod. Biol. Endocrinol. 2018, 16, 80. [Google Scholar] [CrossRef]
- Hoesel, B.; Schmid, J.A. The complexity of NF-κB signaling in inflammation and cancer. Mol. Cancer 2013, 12, 86. [Google Scholar] [CrossRef] [Green Version]
- Burgering, B.M.; Medema, R.H. Decisions on life and death: FOXO Forkhead transcription factors are in command when PKB/Akt is off duty. J. Leukoc. Biol. 2003, 73, 689–701. [Google Scholar] [CrossRef]
- Curtis, C.; Landis, G.N.; Folk, D.; Wehr, N.B.; Hoe, N.; Waskar, M.; Abdueva, D.; Skvortsov, D.; Ford, D.; Luu, A.; et al. Transcriptional profiling of MnSOD-mediated lifespan extension in Drosophila reveals a species-general network of aging and metabolic genes. Genome Biol. 2007, 8, R262. [Google Scholar] [CrossRef] [Green Version]
- Peternelj, T.T.; Marsh, S.A.; Morais, C.; Small, D.M.; Dalbo, V.J.; Tucker, P.S.; Coombes, J.S. O-GlcNAc protein modification in C2C12 myoblasts exposed to oxidative stress indicates parallels with endogenous antioxidant defense. Biochem. Cell Biol. Biochim. Biol. Cell. 2015, 93, 63–73. [Google Scholar] [CrossRef]
- Kajihara, T.; Brosens, J.J.; Ishihara, O. The role of FOXO1 in the decidual transformation of the endometrium and early pregnancy. Med. Mol. Morphol. 2013, 46, 61–68. [Google Scholar] [CrossRef]
- Pearen, M.A.; Myers, S.A.; Raichur, S.; Ryall, J.G.; Lynch, G.S.; Muscat, G.E. The orphan nuclear receptor, NOR-1, a target of beta-adrenergic signaling, regulates gene expression that controls oxidative metabolism in skeletal muscle. Endocrinology 2008, 149, 2853–2865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolmus, K.; Tavernier, J.; Gerlo, S. β2-Adrenergic receptors in immunity and inflammation: stressing NF-κB. Brain Behav. Immun. 2015, 45, 297–310. [Google Scholar] [CrossRef] [PubMed]
- Fagerholm, V.; Rokka, J.; Nyman, L.; Sallinen, J.; Tiihonen, J.; Tupala, E.; Haaparanta, M.; Hietala, J. Autoradiographic characterization of alpha(2C)-adrenoceptors in the human striatum. Synapse 2008, 62, 508–515. [Google Scholar] [CrossRef] [PubMed]
- Xia, B.; Chen, C.; Zhang, H.; Xue, W.; Tang, J.; Tao, W.; Wu, R.; Ren, L.; Wang, W.; Chen, G. Chronic stress prior to pregnancy potentiated long-lasting postpartum depressive-like behavior, regulated by Akt-mTOR signaling in the hippocampus. Sci. Rep. 2016, 6, 35042. [Google Scholar] [CrossRef] [Green Version]
- Goldstein, J.M.; Holsen, L.; Huang, G.; Hammond, B.D.; James-Todd, T.; Cherkerzian, S.; Hale, T.M.; Handa, R.J. Prenatal stress-immune programming of sex differences in comorbidity of depression and obesity/metabolic syndrome. Dialogues Clin. Neurosci. 2016, 18, 425–436. [Google Scholar]
- Guzel, E.; Arlier, S.; Guzeloglu-Kayisli, O.; Tabak, M.S.; Ekiz, T.; Semerci, N.; Larsen, K.; Schatz, F.; Lockwood, C.J.; Kayisli, U.A. Endoplasmic Reticulum Stress and Homeostasis in Reproductive Physiology and Pathology. Int. J. Mol. Sci. 2017, 18, 792. [Google Scholar] [CrossRef] [Green Version]
- Zafir, A.; Banu, N. Modulation of in vivo oxidative status by exogenous corticosterone and restraint stress in rats. Stress 2009, 12, 167–177. [Google Scholar] [CrossRef]
- Joseph, D.N.; Whirledge, S. Stress and the HPA Axis: Balancing Homeostasis and Fertility. Int. J. Mol. Sci. 2017, 18, 2224. [Google Scholar] [CrossRef]
- Cort, A.; Ozben, T.; Saso, L.; De Luca, C.; Korkina, L. Redox Control of Multidrug Resistance and Its Possible Modulation by Antioxidants. Oxid. Med. Cell. Longev. 2016, 2016, 4251912. [Google Scholar] [CrossRef] [Green Version]
- de Kloet, E.R.; Otte, C.; Kumsta, R.; Kok, L.; Hillegers, M.H.; Hasselmann, H.; Kliegel, D.; Joëls, M. Stress and Depression: A Crucial Role of the Mineralocorticoid Receptor. J. Neuroendocrinol. 2016, 28. [Google Scholar] [CrossRef]
- Schmidt, K.L.; Macdougall-Shackleton, E.A.; Soma, K.K.; Macdougall-Shackleton, S.A. Developmental programming of the HPA and HPG axes by early-life stress in male and female song sparrows. Gen. Comp. Endocrinol. 2014, 196, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Leeners, B.; Geary, N.; Tobler, P.N.; Asarian, L. Ovarian hormones and obesity. Hum. Reprod. Update 2017, 23, 300–321. [Google Scholar] [CrossRef] [PubMed]
- Morris, A. Anxiety-induced weight loss. Nat. Rev. Endocrinol. 2019, 15, 130. [Google Scholar] [PubMed]
- Li, Y.; Jiang, W.; Li, Z.Z.; Zhang, C.; Huang, C.; Yang, J.; Kong, G.Y.; Li, Z.F. Repetitive restraint stress changes spleen immune cell subsets through glucocorticoid receptor or beta-adrenergic receptor in a stage dependent manner. Biochem. Biophys. Res. Commun. 2018, 495, 1108–1114. [Google Scholar] [CrossRef]
- Kodogo, V.; Azibani, F.; Sliwa, K. Role of pregnancy hormones and hormonal interaction on the maternal cardiovascular system: A literature review. Clin. Res. Cardiol. 2019, 108, 831–846. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.Y.; Lv, Z.D.; Wang, K.; Qian, L.; Song, X.X.; Li, X.F.; Shen, H.X. Estradiol Alleviates Intervertebral Disc Degeneration through Modulating the Antioxidant Enzymes and Inhibiting Autophagy in the Model of Menopause Rats. Oxid. Med. Cell. Longev. 2018, 2018, 7890291. [Google Scholar] [CrossRef] [Green Version]
- Lambrinoudaki, I.V.; Augoulea, A.; Christodoulakos, G.E.; Economou, E.V.; Kaparos, G.; Kontoravdis, A.; Papadias, C.; Creatsas, G. Measurable serum markers of oxidative stress response in women with endometriosis. Fertil. Steril. 2009, 91, 46–50. [Google Scholar] [CrossRef]
- Prieto, L.; Quesada, J.F.; Cambero, O.; Pacheco, A.; Pellicer, A.; Codoceo, R.; Garcia-Velasco, J.A. Analysis of follicular fluid and serum markers of oxidative stress in women with infertility related to endometriosis. Fertil. Steril. 2012, 98, 126–130. [Google Scholar] [CrossRef]
- Londero, A.P.; Orsaria, M.; Marzinotto, S.; Grassi, T.; Fruscalzo, A.; Calcagno, A.; Bertozzi, S.; Nardini, N.; Stella, E.; Lellé, R.J.; et al. Placental aging and oxidation damage in a tissue micro-array model: An immunohistochemistry study. Histochem. Cell Biol. 2016, 146, 191–204. [Google Scholar] [CrossRef] [Green Version]
- Kwak-Kim, J.; Bao, S.; Lee, S.K.; Kim, J.W.; Gilman-Sachs, A. Immunological modes of pregnancy loss: Inflammation, immune effectors, and stress. Am. J. Reprod. Immunol. 2014, 72, 129–140. [Google Scholar] [CrossRef]
- Doukali, H.; Ben Salah, G.; Hamdaoui, L.; Hajjaji, M.; Tabebi, M.; Ammar-Keskes, L.; Masmoudi, M.E.; Kamoun, H. Oxidative stress and glutathione S-transferase genetic polymorphisms in medical staff professionally exposed to ionizing radiation. Int. J. Radiat. Biol. 2017, 93, 697–704. [Google Scholar] [CrossRef]
- Sies, H.; Berndt, C.; Jones, D.P. Oxidative Stress. Annu. Rev. Biochem. 2017, 86, 715–748. [Google Scholar] [CrossRef]
- Kaestner, K.H.; Knochel, W.; Martinez, D.E. Unified nomenclature for the winged helix/forkhead transcription factors. Genes Dev. 2000, 14, 142–146. [Google Scholar]
- Brunet, A.; Bonni, A.; Zigmond, M.J.; Lin, M.Z.; Juo, P.; Hu, L.S.; Anderson, M.J.; Arden, K.C.; Blenis, J.; Greenberg, M.E. Akt promotes cell survival by phosphorylating and inhibiting a Forkhead transcription factor. Cell 1999, 96, 857–868. [Google Scholar] [CrossRef] [Green Version]
- Tsuchiya, K.; Westerterp, M.; Murphy, A.J.; Subramanian, V.; Ferrante, A.W., Jr.; Tall, A.R.; Accili, D. Expanded granulocyte/monocyte compartment in myeloid-specific triple FoxO knockout increases oxidative stress and accelerates atherosclerosis in mice. Circ. Res. 2013, 112, 992–1003. [Google Scholar] [CrossRef] [Green Version]
- He, W.; Zhang, A.; Qi, L.; Na, C.; Jiang, R.; Fan, Z.; Chen, J. FOXO1, a Potential Therapeutic Target, Regulates Autophagic Flux, Oxidative Stress, Mitochondrial Dysfunction, and Apoptosis in Human Cholangiocarcinoma QBC939 Cells. Cell. Physiol. Biochem. 2018, 45, 1506–1514. [Google Scholar] [CrossRef]
- Grinius, L.; Kessler, C.; Schroeder, J.; Handwerger, S. Forkhead transcription factor FOXO1A is critical for induction of human decidualization. J. Endocrinol. 2006, 189, 179–187. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.P.; Chen, C.Y.; Wu, Y.H.; Chen, C.Y. Oxidative stress reduces trophoblast FOXO1 and integrin beta3 expression that inhibits cell motility. Free Radic. Biol. Med. 2018, 124, 189–198. [Google Scholar] [CrossRef]
- Kuscu, N.; Celik-Ozenci, C. FOXO1, FOXO3, AND FOXO4 are differently expressed during mouse oocyte maturation and preimplantation embryo development. Gene Expr. Patterns 2015, 18, 16–20. [Google Scholar] [CrossRef]
- Martin, B.; Mattson, M.P.; Maudsley, S. Caloric restriction and intermittent fasting: Two potential diets for successful brain aging. Ageing Res. Rev. 2006, 5, 332–353. [Google Scholar] [CrossRef] [Green Version]
- Manolopoulos, K.N.; Klotz, L.O.; Korsten, P.; Bornstein, S.R.; Barthel, A. Linking Alzheimer’s disease to insulin resistance: The FoxO response to oxidative stress. Mol. Psychiatry 2010, 15, 1046–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rached, M.T.; Kode, A.; Xu, L.; Yoshikawa, Y.; Paik, J.H.; Depinho, R.A.; Kousteni, S. FoxO1 is a positive regulator of bone formation by favoring protein synthesis and resistance to oxidative stress in osteoblasts. Cell Metab. 2010, 11, 147–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grabiec, A.M.; Angiolilli, C.; Hartkamp, L.M.; van Baarsen, L.G.; Tak, P.P.; Reedquist, K.A. JNK-dependent downregulation of FoxO1 is required to promote the survival of fibroblast-like synoviocytes in rheumatoid arthritis. Ann. Rheum. Dis. 2015, 74, 1763–1771. [Google Scholar] [CrossRef]
- Walsh, D.A.; Perkins, J.P.; Krebs, E.G. An adenosine 3’,5’-monophosphate-dependant protein kinase from rabbit skeletal muscle. J. Biol. Chem. 1968, 243, 3763–3765. [Google Scholar] [CrossRef]
- Xiao, R.P.; Ji, X.; Lakatta, E.G. Functional coupling of the beta 2-adrenoceptor to a pertussis toxin-sensitive G protein in cardiac myocytes. Mol. Pharmacol. 1995, 47, 322–329. [Google Scholar]
- Philipp, M.; Hein, L. Adrenergic receptor knockout mice: Distinct functions of 9 receptor subtypes. Pharmacol. Ther. 2004, 101, 65–74. [Google Scholar] [CrossRef]
- Bruzzone, M.E.; Fabres, C.; Benitez, D.A.; Castellon, E.A.; Zegers-Hochschild, F. Influence of embryonic conditioned media upon the endometrial beta-adrenergic receptor. Reprod. Biomed. Online 2005, 11, 58–63. [Google Scholar] [CrossRef]
- Yamazaki, S.; Miyoshi, N.; Kawabata, K.; Yasuda, M.; Shimoi, K. Quercetin-3-O-glucuronide inhibits noradrenaline-promoted invasion of MDA-MB-231 human breast cancer cells by blocking beta(2)-adrenergic signaling. Arch. Biochem. Biophys. 2014, 557, 18–27. [Google Scholar] [CrossRef]
- Madden, K.S. Catecholamines, sympathetic innervation, and immunity. Brain Behav. Immun. 2003, 17 (Suppl. S1), S5–S10. [Google Scholar] [CrossRef]
- McReynolds, J.R.; Vranjkovic, O.; Thao, M.; Baker, D.A.; Makky, K.; Lim, Y.; Mantsch, J.R. Beta-2 adrenergic receptors mediate stress-evoked reinstatement of cocaine-induced conditioned place preference and increases in CRF mRNA in the bed nucleus of the stria terminalis in mice. Psychopharmacology 2014, 231, 3953–3963. [Google Scholar] [CrossRef]
- Mills, P.J.; Adler, K.A.; Dimsdale, J.E.; Perez, C.J.; Ziegler, M.G.; Ancoli-Israel, S.; Patterson, T.L.; Grant, I. Vulnerable caregivers of Alzheimer disease patients have a deficit in beta 2-adrenergic receptor sensitivity and density. Am. J. Geriatr. Psychiatry 2004, 12, 281–286. [Google Scholar] [PubMed]
- Sivamani, R.K.; Pullar, C.E.; Manabat-Hidalgo, C.G.; Rocke, D.M.; Carlsen, R.C.; Greenhalgh, D.G.; Isseroff, R.R. Stress-mediated increases in systemic and local epinephrine impair skin wound healing: Potential new indication for beta blockers. PLoS Med. 2009, 6, e12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, Q.; Liu, X.; Qi, Q.; Guo, S.W. Chronic stress accelerates the development of endometriosis in mouse through adrenergic receptor beta2. Hum. Reprod. 2016, 31, 2506–2519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, M.B.; Jin, D.D.; Jiao, Y.J.; Ni, W.K.; Liu, J.X.; Qu, L.S.; Lu, C.H.; Ni, R.Z.; Jiang, F.; Chen, W.C. beta2-AR regulates the expression of AKR1B1 in human pancreatic cancer cells and promotes their proliferation via the ERK1/2 pathway. Mol. Biol. Rep. 2018, 1, 1863–1871. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M. Molecular mechanisms of beta(2)-adrenergic receptor function, response, and regulation. J. Allergy Clin. Immunol. 2006, 117, 18–24, quiz 25. [Google Scholar] [CrossRef]
- Mantyh, P.W.; Rogers, S.D.; Allen, C.J.; Catton, M.D.; Ghilardi, J.R.; Levin, L.A.; Maggio, J.E.; Vigna, S.R. Beta 2-adrenergic receptors are expressed by glia in vivo in the normal and injured central nervous system in the rat, rabbit, and human. J. Neurosci. 1995, 15, 152–164. [Google Scholar]
- Pullar, C.E.; Rizzo, A.; Isseroff, R.R. beta-Adrenergic receptor antagonists accelerate skin wound healing: Evidence for a catecholamine synthesis network in the epidermis. J. Biol. Chem. 2006, 281, 21225–21235. [Google Scholar] [CrossRef] [Green Version]
- Farmer, P.; Pugin, J. beta-adrenergic agonists exert their “anti-inflammatory” effects in monocytic cells through the IκB/NF-κB pathway. Am. J. Physiol. 2000, 279, L675–L682. [Google Scholar]
- Gavrilyuk, V.; Dello Russo, C.; Heneka, M.T.; Pelligrino, D.; Weinberg, G.; Feinstein, D.L. Norepinephrine increases I κ B alpha expression in astrocytes. J. Biol. Chem. 2002, 277, 29662–29668. [Google Scholar] [CrossRef] [Green Version]
- Sharma, M.; Flood, P.M. β-arrestin2 regulates the anti-inflammatory effects of Salmeterol in lipopolysaccharide-stimulated BV2 cells. J. Neuroimmunol. 2018, 325, 10–19. [Google Scholar] [CrossRef]
- Gao, H.; Sun, Y.; Wu, Y.; Luan, B.; Wang, Y.; Qu, B.; Pei, G. Identification of beta-arrestin2 as a G protein-coupled receptor-stimulated regulator of NF-κB pathways. Mol. Cell 2004, 14, 303–317. [Google Scholar] [CrossRef]
- Chandrasekar, B.; Marelli-Berg, F.M.; Tone, M.; Bysani, S.; Prabhu, S.D.; Murray, D.R. Beta-adrenergic stimulation induces interleukin-18 expression via beta2-AR, PI3K, Akt, IKK, and NF-κB. Biochem. Biophys. Res. Commun. 2004, 319, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.; Ruan, L.; Qian, L.; Liu, X.; Le, Y. Norepinephrine promotes microglia to uptake and degrade amyloid beta peptide through upregulation of mouse formyl peptide receptor 2 and induction of insulin-degrading enzyme. J. Neurosci. 2010, 30, 11848–11857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freddolino, P.L.; Kalani, M.Y.; Vaidehi, N.; Floriano, W.B.; Hall, S.E.; Trabanino, R.J.; Kam, V.W.; Goddard, W.A. 3rd. Predicted 3D structure for the human beta 2 adrenergic receptor and its binding site for agonists and antagonists. Proc. Natl. Acad. Sci. USA 2004, 101, 2736–2741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Yano, N.; Deng, M.; Mao, Q.; Shaw, S.K.; Tseng, Y.T. beta-Adrenergic receptor-PI3K signaling crosstalk in mouse heart: Elucidation of immediate downstream signaling cascades. PLoS ONE 2011, 6, e26581. [Google Scholar]
- Siqueira, M.F.; Li, J.; Chehab, L.; Desta, T.; Chino, T.; Krothpali, N.; Behl, Y.; Alikhani, M.; Yang, J.; Braasch, C.; et al. Impaired wound healing in mouse models of diabetes is mediated by TNF-alpha dysregulation and associated with enhanced activation of forkhead box O1 (FOXO1). Diabetologia 2010, 53, 378–388. [Google Scholar]
- Shi, X.; Wang, X.; Li, Q.; Su, M.; Chew, E.; Wong, E.T.; Lacza, Z.; Radda, G.K.; Tergaonkar, V.; Han, W. Nuclear factor κB (NF-κB) suppresses food intake and energy expenditure in mice by directly activating the Pomc promoter. Diabetologia 2013, 56, 925–936. [Google Scholar] [CrossRef] [Green Version]
- Perry, B.D.; Caldow, M.K.; Brennan-Speranza, T.C.; Sbaraglia, M.; Jerums, G.; Garnham, A.; Wong, C.; Levinger, P.; Asrar Ul Haq, M.; Hare, D.L.; et al. Muscle atrophy in patients with Type 2 Diabetes Mellitus: Roles of inflammatory pathways, physical activity and exercise. Exerc. Immunol. Rev. 2016, 22, 94–109. [Google Scholar]
- De Clercq, K.; Hennes, A.; Vriens, J. Isolation of Mouse Endometrial Epithelial and Stromal Cells for In Vitro Decidualization. J. Vis. Exp. 2017. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Liu, G.; Wang, Z.; Cao, J.; Chen, Y.; Dong, Y. Restraint stress induces uterine microenvironment disorder in mice during early pregnancy through the β2-AR/cAMP/PKA pathway. Stress 2020, 1–15. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, J.; Chen, Y.; Wang, Z.; Cao, J.; Dong, Y. The Role of the FOXO1/β2-AR/p-NF-κB p65 Pathway in the Development of Endometrial Stromal Cells in Pregnant Mice under Restraint Stress. Int. J. Mol. Sci. 2021, 22, 1478. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031478
Lu J, Chen Y, Wang Z, Cao J, Dong Y. The Role of the FOXO1/β2-AR/p-NF-κB p65 Pathway in the Development of Endometrial Stromal Cells in Pregnant Mice under Restraint Stress. International Journal of Molecular Sciences. 2021; 22(3):1478. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031478
Chicago/Turabian StyleLu, Jiayin, Yaoxing Chen, Zixu Wang, Jing Cao, and Yulan Dong. 2021. "The Role of the FOXO1/β2-AR/p-NF-κB p65 Pathway in the Development of Endometrial Stromal Cells in Pregnant Mice under Restraint Stress" International Journal of Molecular Sciences 22, no. 3: 1478. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031478