Structural Insight into the Two-Step Mechanism of PAI-1 Inhibition by Small Molecule TM5484

 , and
, and
Abstract
:1. Introduction
2. Results and Discussion
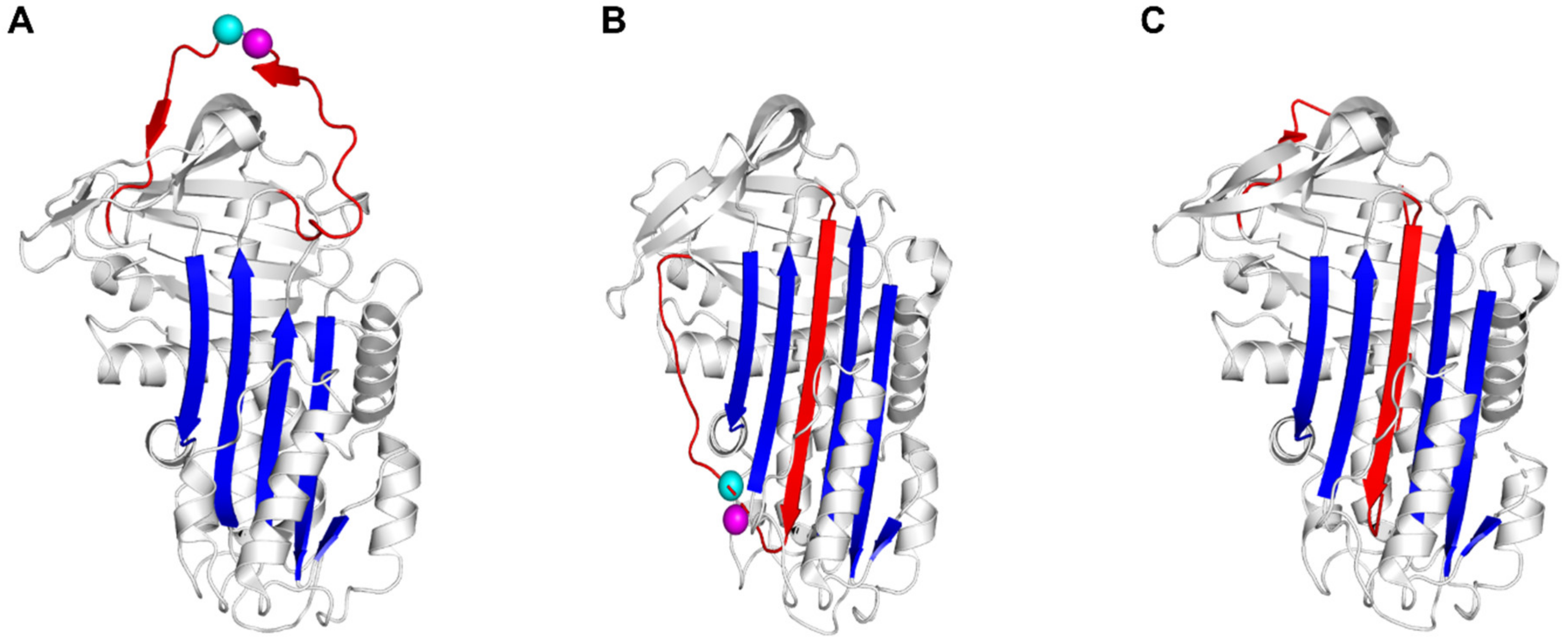
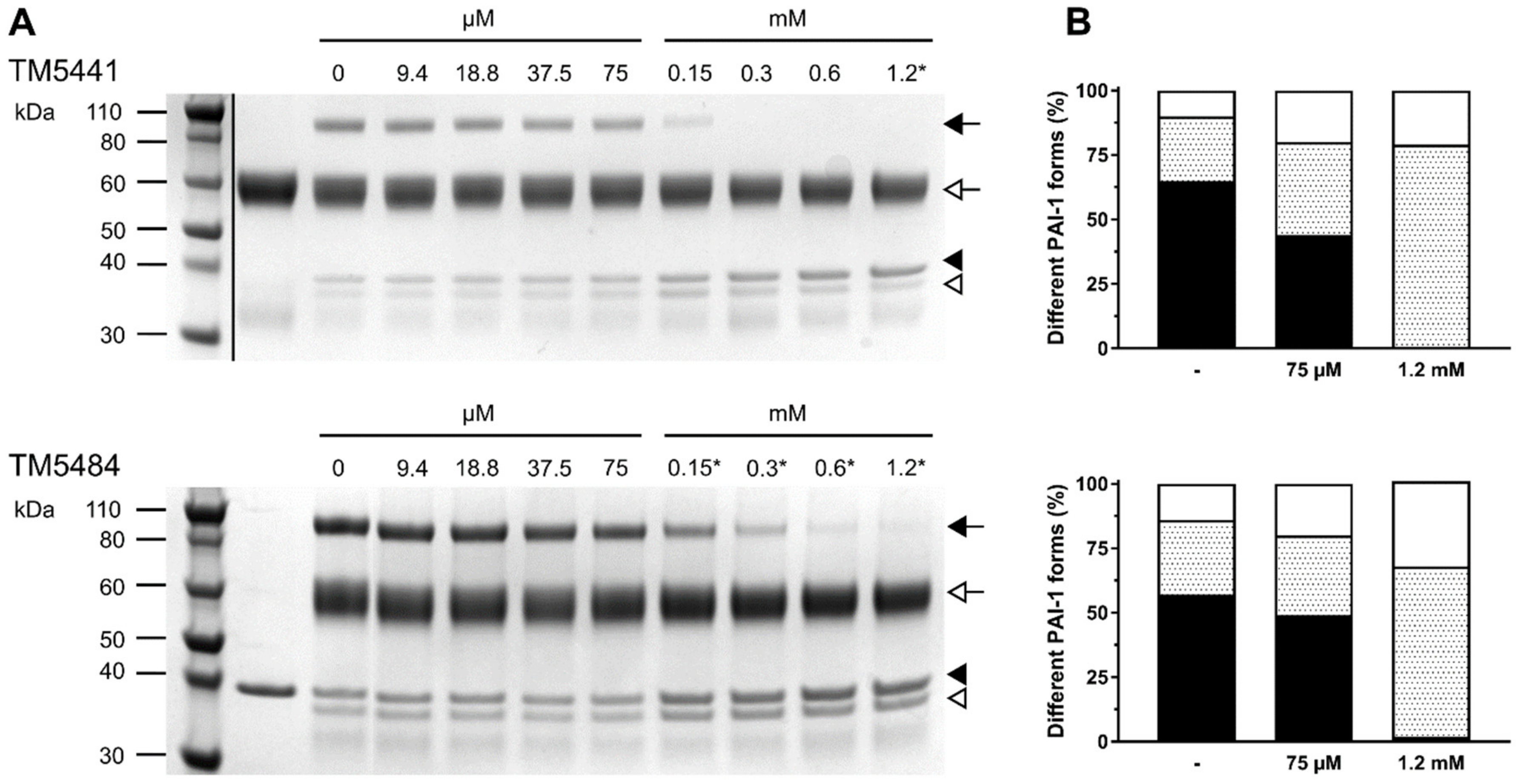
2.1. Mechanism of Compound-Induced PAI-1 Inhibition
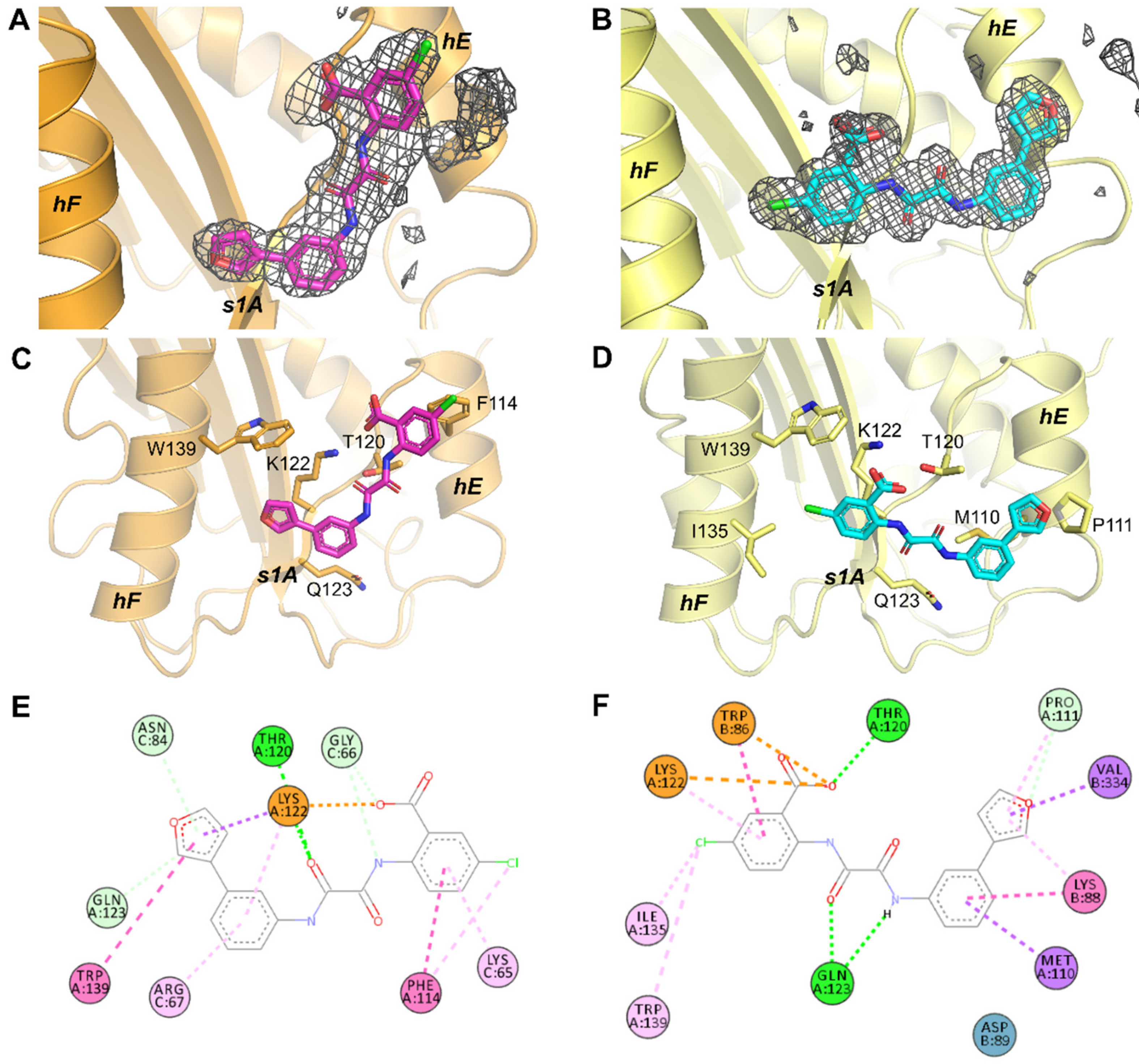
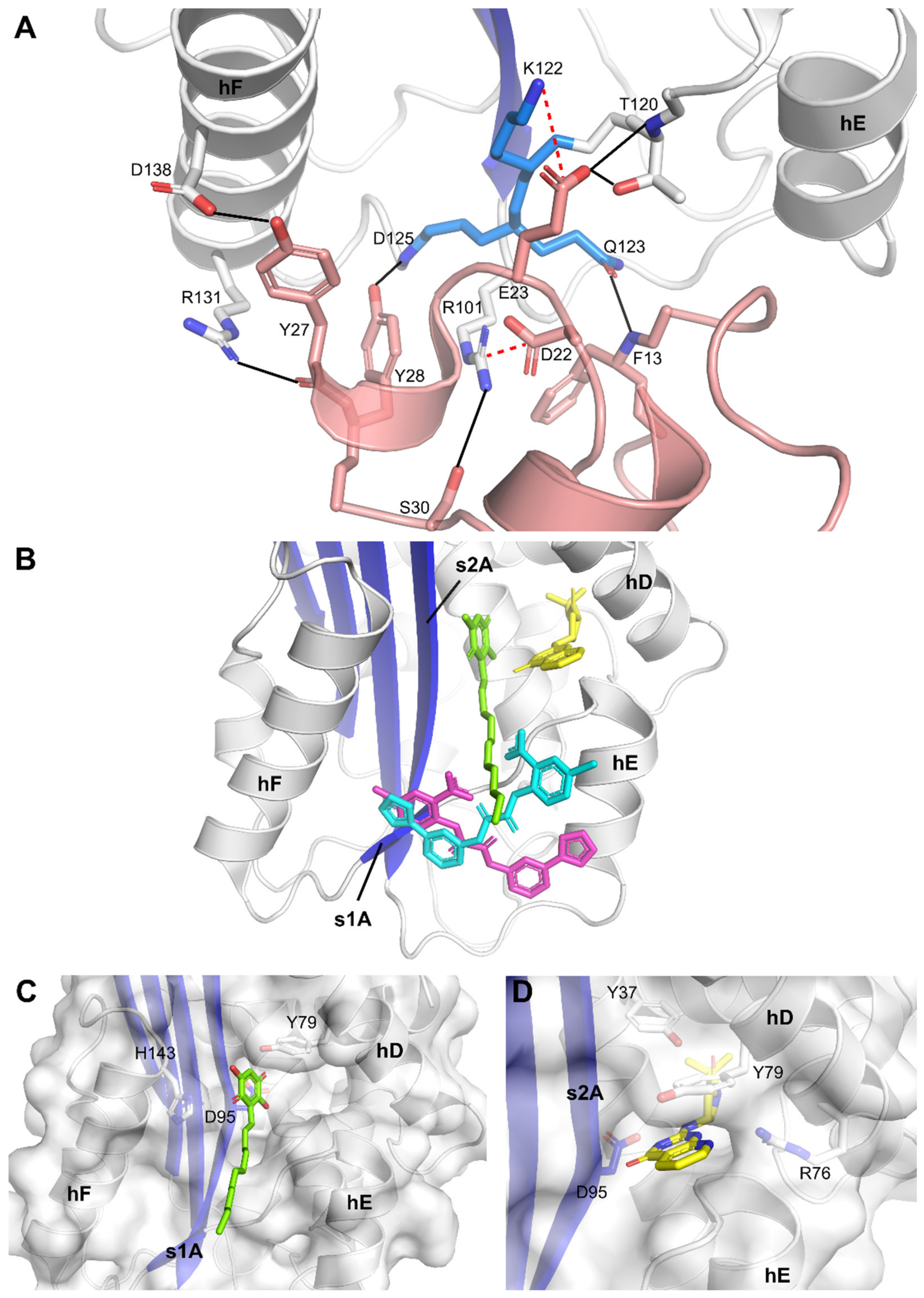
2.2. Crystallographic Analysis of Compound-Bound PAI-1 Complexes
3. Materials and Methods
3.1. Cloning, Expression, and Purification of PAI-1 and Nbs
3.2. Determination of the Conformational Distribution of PAI-1 upon Compound-Induced Inhibition
3.3. Crystallization and Data Collection
3.4. Structure Determination, Refinement, and Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ASU | asymmetric unit |
| Cα RMSD | root-mean-squared deviations for all Cα atoms |
| h[X] | α-helix X |
| PA | plasminogen activator |
| PAI-1 | plasminogen activator inhibitor-1 |
| PAI-1-stab | PAI-1-N150H-K154T-Q301P-Q319L-M354I |
| PDB | Protein Data Bank |
| RCL | reactive center loop |
| s[x]A | strand number [x] of β-sheet A |
| SDS-PAGE | sodium dodecyl sulfate-polyacrylamide gel electrophoresis |
| tPA | tissue-type plasminogen activator |
| uPA | urokinase-type plasminogen activator |
| Vn | vitronectin |
References
- Gong, L.; Liu, M.; Zeng, T.; Shi, X.; Yuan, C.; Andreasen, P.A.; Huang, M. Crystal structure of the Michaelis complex between tissue-type plasminogen activator and plasminogen activators inhibitor-1. J. Biol. Chem. 2015, 290, 25795–25804. [Google Scholar] [CrossRef] [Green Version]
- Li, S.H.; Reinke, A.A.; Sanders, K.L.; Emal, C.D.; Whisstock, J.C.; Stuckey, J.A.; Lawrence, D.A. Mechanistic characterization and crystal structure of a small molecule inactivator bound to plasminogen activator inhibitor-1. Proc. Natl. Acad. Sci. USA 2013, 110, E4941–E4949. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, D.A.; Ginsburg, D.; Day, D.E.; Berkenpas, M.B.; Verhamme, I.M.; Kvassman, J.O.; Shore, J.D. Serpin-protease complexes are trapped as stable acyl-enzyme intermediates. J. Biol. Chem. 1995, 270, 25309–25312. [Google Scholar] [CrossRef] [Green Version]
- Shore, J.D.; Day, D.E.; Francis-Chmura, A.M.; Verhamme, I.; Kvassman, J.; Lawrence, D.A.; Ginsburg, D. A fluorescent probe study of plasminogen activator inhibitor-1. Evidence for reactive center loop insertion and its role in the inhibitory mechanism. J. Biol. Chem. 1995, 270, 5395–5398. [Google Scholar] [CrossRef] [Green Version]
- Huntington, J.A.; Read, R.J.; Carrell, R.W. Structure of a serpin-protease complex shows inhibition by deformation. Nature 2000, 407, 923–926. [Google Scholar] [CrossRef]
- Perron, M.J.; Blouse, G.E.; Shore, J.D. Distortion of the catalytic domain of tissue-type plasminogen activator by plasminogen activator inhibitor-1 coincides with the formation of stable serpin-proteinase complexes. J. Biol. Chem. 2003, 278, 48197–48203. [Google Scholar] [CrossRef] [Green Version]
- Van Meijer, M.; Smilde, A.; Tans, G.; Nesheim, M.E.; Pannekoek, H.; Horrevoets, A.J. The suicide substrate reaction between plasminogen activator inhibitor 1 and thrombin is regulated by the cofactors vitronectin and heparin. Blood 1997, 90, 1874–1882. [Google Scholar] [CrossRef] [Green Version]
- Zhou, A.; Huntington, J.A.; Pannu, N.S.; Carrell, R.W.; Read, R.J. How vitronectin binds PAI-1 to modulate fibrinolysis and cell migration. Nat. Struct. Biol. 2003, 10, 541–544. [Google Scholar] [CrossRef]
- Declerck, P.J.; De Mol, M.; Alessi, M.C.; Baudner, S.; Pâques, E.P.; Preissner, K.T.; Müller-Berghaus, G.; Collen, D. Purification and characterization of a plasminogen activator inhibitor 1 binding protein from human plasma. Identification as a multimeric form of S protein (vitronectin). J. Biol. Chem. 1988, 263, 15454–15461. [Google Scholar] [CrossRef]
- Lindahl, T.L.; Sigurdardottir, O.; Wiman, B. Stability of plasminogen activator inhibitor 1 (PAI-1). Thromb. Haemost. 1989, 62, 748–751. [Google Scholar] [CrossRef]
- Nar, H.; Bauer, M.; Stassen, J.M.; Lang, D.; Gils, A.; Declerck, P.J. Plasminogen activator inhibitor 1. Structure of the native serpin, comparison to its other conformers and implications for serpin inactivation. J. Mol. Biol. 2000, 297, 683–695. [Google Scholar] [CrossRef] [PubMed]
- Stout, T.J.; Graham, H.; Buckley, D.I.; Matthews, D.J. Structures of active and latent PAI-1: A possible stabilizing role for chloride ions. Biochemistry 2000, 39, 8460–8469. [Google Scholar] [CrossRef] [PubMed]
- Dewilde, M.; Strelkov, S.V.; Rabijns, A.; Declerck, P.J. High quality structure of cleaved PAI-1-stab. J. Struct. Biol. 2009, 165, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Jung, R.G.; Motazedian, P.; Ramirez, F.D.; Simard, T.; Di Santo, P.; Visintini, S.; Faraz, M.A.; Labinaz, A.; Jung, Y.; Hibbert, B. Association between plasminogen activator inhibitor-1 and cardiovascular events: A systematic review and meta-analysis. Thromb. J. 2018, 16, 12. [Google Scholar] [CrossRef] [Green Version]
- Flevaris, P.; Vaughan, D. The role of plasminogen activator inhibitor type-1 in fibrosis. Semin. Thromb. Hemost. 2017, 43, 169–177. [Google Scholar] [CrossRef]
- Kubala, M.H.; DeClerck, Y.A. The plasminogen activator inhibitor-1 paradox in cancer: A mechanistic understanding. Cancer Metastasis Rev. 2019, 38, 483–492. [Google Scholar] [CrossRef]
- Vaughan, D.E.; Rai, R.; Khan, S.S.; Eren, M.; Ghosh, A.K. Plasminogen activator inhibitor-1 is a marker and a mediator of senescence. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 1446–1452. [Google Scholar] [CrossRef] [Green Version]
- Boe, A.E.; Eren, M.; Murphy, S.B.; Kamide, C.E.; Ichimura, A.; Terry, D.; McAnally, D.; Smith, L.H.; Miyata, T.; Vaughan, D.E. Plasminogen activator inhibitor-1 antagonist TM5441 attenuates Nω-nitro-L-arginine methyl ester-induced hypertension and vascular senescence. Circulation 2013, 128, 2318–2324. [Google Scholar] [CrossRef] [Green Version]
- Pelisch, N.; Dan, T.; Ichimura, A.; Sekiguchi, H.; Vaughan, D.E.; van Ypersele de Strihou, C.; Miyata, T. Plasminogen activator inhibitor-1 antagonist TM5484 attenuates demyelination and axonal degeneration in a mice model of multiple sclerosis. PLoS ONE 2015, 10, e0124510. [Google Scholar] [CrossRef]
- Izuhara, Y.; Takahashi, S.; Nangaku, M.; Takizawa, S.; Ishida, H.; Kurokawa, K.; van Ypersele de Strihou, C.; Hirayama, N.; Miyata, T. Inhibition of plasminogen activator inhibitor-1: Its mechanism and effectiveness on coagulation and fibrosis. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 672–677. [Google Scholar] [CrossRef] [Green Version]
- Izuhara, Y.; Yamaoka, N.; Kodama, H.; Dan, T.; Takizawa, S.; Hirayama, N.; Meguro, K.; van Ypersele de Strihou, C.; Miyata, T. A novel inhibitor of plasminogen activator inhibitor-1 provides antithrombotic benefits devoid of bleeding effect in nonhuman primates. J. Cereb. Blood. Flow Metab. 2010, 30, 904–912. [Google Scholar] [CrossRef] [PubMed]
- Fortenberry, Y.M. Plasminogen activator inhibitor-1 inhibitors: A patent review (2006–present). Expert Opin. Ther. Pat. 2013, 23, 801–815. [Google Scholar] [CrossRef] [PubMed]
- Rouch, A.; Vanucci-Bacqué, C.; Bedos-Belval, F.; Baltas, M. Small molecules inhibitors of plasminogen activator inhibitor-1—An overview. Eur. J. Med. Chem. 2015, 92, 619–636. [Google Scholar] [CrossRef] [PubMed]
- Yamaoka, N.; Kawano, Y.; Izuhara, Y.; Miyata, T.; Meguro, K. Structure-activity relationships of new 2-acylamino-3-thiophenecarboxylic acid dimers as plasminogen activator inhibitor-1 inhibitors. Chem. Pharm. Bull. 2010, 58, 615–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaoka, N.; Murano, K.; Kodama, H.; Maeda, A.; Dan, T.; Nakabayashi, T.; Miyata, T.; Meguro, K. Identification of novel plasminogen activator inhibitor-1 inhibitors with improved oral bioavailability: Structure optimization of N-acylanthranilic acid derivatives. Bioorg. Med. Chem. Lett. 2018, 28, 809–813. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.L.; Bishop, N.; Li, Z.; Cipolla, M.J. Inhibition of PAI (plasminogen activator inhibitor)-1 improves brain collateral perfusion and injury after acute ischemic stroke in aged hypertensive rats. Stroke 2018, 49, 1969–1976. [Google Scholar] [CrossRef] [PubMed]
- Placencio, V.R.; Ichimura, A.; Miyata, T.; DeClerck, Y.A. Small molecule inhibitors of plasminogen activator inhibitor-1 elicit anti-tumorigenic and anti-angiogenic activity. PLoS ONE 2015, 10, e0133786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henkel, A.S.; Khan, S.S.; Olivares, S.; Miyata, T.; Vaughan, D.E. Inhibition of plasminogen activator inhibitor 1 attenuates hepatic steatosis but does not prevent progressive nonalcoholic steatohepatitis in mice. Hepatol. Commun. 2018, 2, 1479–1492. [Google Scholar] [CrossRef] [Green Version]
- Piao, L.; Jung, I.; Huh, J.Y.; Miyata, T.; Ha, H. A novel plasminogen activator inhibitor-1 inhibitor, TM5441, protects against high-fat diet-induced obesity and adipocyte injury in mice. Br. J. Pharmacol. 2016, 173, 2622–2632. [Google Scholar] [CrossRef]
- Ghosh, A.K.; Rai, R.; Park, K.E.; Eren, M.; Miyata, T.; Wilsbacher, L.D.; Vaughan, D.E. A small molecule inhibitor of PAI-1 protects against doxorubicin-induced cellular senescence. Oncotarget 2016, 7, 72443–72457. [Google Scholar] [CrossRef] [Green Version]
- Sun, T.; Ghosh, A.K.; Eren, M.; Miyata, T.; Vaughan, D.E. PAI-1 contributes to homocysteine-induced cellular senescence. Cell Signal. 2019, 64, 109394. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, S.; Martial, J.A.; Struman, I. A peptide mimicking the C-terminal part of the reactive center loop induces the transition to the latent form of plasminogen activator inhibitor type-1. FEBS Lett. 2012, 586, 686–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorlatova, N.V.; Cale, J.M.; Elokdah, H.; Li, D.; Fan, K.; Warnock, M.; Crandall, D.L.; Lawrence, D.A. Mechanism of inactivation of plasminogen activator inhibitor-1 by a small molecule inhibitor. J. Biol. Chem. 2007, 282, 9288–9296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imai, Y.N.; Inoue, Y.; Nakanishi, I.; Kitaura, K. Cl-pi interactions in protein-ligand complexes. Protein Sci. 2008, 17, 1129–1137. [Google Scholar] [CrossRef] [Green Version]
- Trelle, M.B.; Hirschberg, D.; Jansson, A.; Ploug, M.; Roepstorff, P.; Andreasen, P.A.; Jørgensen, T.J. Hydrogen/deuterium exchange mass spectrometry reveals specific changes in the local flexibility of plasminogen activator inhibitor 1 upon binding to the somatomedin B domain of vitronectin. Biochemistry 2012, 51, 8256–8266. [Google Scholar] [CrossRef]
- Hennan, J.K.; Morgan, G.A.; Swillo, R.E.; Antrilli, T.M.; Mugford, C.; Vlasuk, G.P.; Gardell, S.J.; Crandall, D.L. Effect of tiplaxtinin (PAI-039), an orally bioavailable PAI-1 antagonist, in a rat model of thrombosis. J. Thromb. Haemost. 2008, 6, 1558–1564. [Google Scholar] [CrossRef]
- Elokdah, H.; Abou-Gharbia, M.; Hennan, J.K.; McFarlane, G.; Mugford, C.P.; Krishnamurthy, G.; Crandall, D.L. Tiplaxtinin, a novel, orally efficacious inhibitor of plasminogen activator inhibitor-1: Design, synthesis, and preclinical characterization. J. Med. Chem 2004, 47, 3491–3494. [Google Scholar] [CrossRef] [Green Version]
- Lin, Z.; Jensen, J.K.; Hong, Z.; Shi, X.; Hu, L.; Andreasen, P.A.; Huang, M. Structural insight into inactivation of plasminogen activator inhibitor-1 by a small-molecule antagonist. Chem. Biol. 2013, 20, 253–261. [Google Scholar] [CrossRef] [Green Version]
- Fjellström, O.; Deinum, J.; Sjögren, T.; Johansson, C.; Geschwindner, S.; Nerme, V.; Legnehed, A.; McPheat, J.; Olsson, K.; Bodin, C.; et al. Characterization of a small molecule inhibitor of plasminogen activator inhibitor type 1 that accelerates the transition into the latent conformation. J. Biol. Chem. 2013, 288, 873–885. [Google Scholar] [CrossRef] [Green Version]
- Egelund, R.; Einholm, A.P.; Pedersen, K.E.; Nielsen, R.W.; Christensen, A.; Deinum, J.; Andreasen, P.A. A regulatory hydrophobic area in the flexible joint region of plasminogen activator inhibitor-1, defined with fluorescent activity-neutralizing ligands: Ligand-unduced serpin polymerization. J. Biol. Chem. 2001, 276, 13077–13086. [Google Scholar] [CrossRef] [Green Version]
- Sillen, M.; Weeks, S.D.; Zhou, X.; Komissarov, A.A.; Florova, G.; Idell, S.; Strelkov, S.V.; Declerck, P.J. Molecular mechanism of two nanobodies that inhibit PAI-1 activity reveals a modulation at distinct stages of the PAI-1/plasminogen activator interaction. J. Thromb. Haemost. 2020, 18, 681–692. [Google Scholar] [CrossRef] [PubMed]
- Sillen, M.; Weeks, S.D.; Strelkov, S.V.; Declerck, P.J. Structural insights into the mechanism of a nanobody that stabilizes PAI-1 and modulates its activity. Int. J. Mol. Sci. 2020, 21. [Google Scholar] [CrossRef]
- Studier, F.W. Protein production by auto-induction in high density shaking cultures. Protein Expr. Purif. 2005, 41, 207–234. [Google Scholar] [CrossRef] [PubMed]
- Vonrhein, C.; Flensburg, C.; Keller, P.; Sharff, A.; Smart, O.; Paciorek, W.; Womack, T.; Bricogne, G. Data processing and analysis with the autoPROC toolbox. Acta Crystallogr. D Biol. Crystallogr. 2011, 67, 293–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moriarty, N.W.; Grosse-Kunstleve, R.W.; Adams, P.D. electronic Ligand Builder and Optimization Workbench (eLBOW): A tool for ligand coordinate and restraint generation. Acta Crystallogr. Sect. D Biol. Crystallogr. 2009, 65, 1074–1080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of Coot. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 486–501. [Google Scholar] [CrossRef] [Green Version]
- Afonine, P.V.; Grosse-Kunstleve, R.W.; Echols, N.; Headd, J.J.; Moriarty, N.W.; Mustyakimov, M.; Terwilliger, T.C.; Urzhumtsev, A.; Zwart, P.H.; Adams, P.D. Towards automated crystallographic structure refinement with phenix.refine. Acta Crystallogr. D Biol. Crystallogr. 2012, 68, 352–367. [Google Scholar] [CrossRef] [Green Version]
- Liebschner, D.; Afonine, P.V.; Moriarty, N.W.; Poon, B.K.; Sobolev, O.V.; Terwilliger, T.C.; Adams, P.D. Polder maps: Improving OMIT maps by excluding bulk solvent. Acta Crystallogr. D Struct. Biol. 2017, 73, 148–157. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PAI-1-W175F/Nb42/Nb64 + TM5484 (2 h Soaking) | PAI-1-Stab + TM5484 | |
|---|---|---|
| Data collection | ||
| Space group | P 2 | C 2 |
| Cell parameters | ||
| a, b, c (Å) | 45.5, 71.5, 96.2 | 135.3, 64.3, 106.6 |
| α, β, γ (°) | 90, 101.3, 90 | 90, 117, 90 |
| Resolution range (Å) | 36.15–2.27 (2.35–2.27) | 33.44–1.77 (1.83–1.77) |
| Rmerge | 0.069 (0.603) | 0.087 (1.07) |
| I/σ (I) | 9.44 (2.02) | 9.03 (1.99) |
| CC1/2 | 0.997 (0.83) | 0.999 (0.812) |
| Completeness (%) | 99.45 (99.60) | 99.36 (98.78) |
| Redundancy | 3.7 (3.7) | 6.7 (7.0) |
| Refinement | ||
| No. of PAI-1 molecules/ASU | 1 | 2 |
| Reflections used in refinement | 27,958 (2759) | 79,244 (7845) |
| Reflections used for Rfree | 2083 (220) | 3996 (407) |
| Rwork | 0.214 (0.320) | 0.178 (0.276) |
| Rfree | 0.266 (0.361) | 0.213 (0.322) |
| No. of non-hydrogen atoms | 4637 | 6254 |
| Protein | 4561 | 5746 |
| Ligands | 27 | 27 |
| Water | 49 | 481 |
| B factors (Å2) | ||
| Protein | 58.47 | 29.32 |
| Ligands | 56.09 | 33.82 |
| Water | 49.95 | 39.73 |
| R.m.s. deviations | ||
| Bond lengths (Å) | 0.002 | 0.009 |
| Bond angles (°) | 0.51 | 0.95 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sillen, M.; Miyata, T.; Vaughan, D.E.; Strelkov, S.V.; Declerck, P.J. Structural Insight into the Two-Step Mechanism of PAI-1 Inhibition by Small Molecule TM5484. Int. J. Mol. Sci. 2021, 22, 1482. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031482
Sillen M, Miyata T, Vaughan DE, Strelkov SV, Declerck PJ. Structural Insight into the Two-Step Mechanism of PAI-1 Inhibition by Small Molecule TM5484. International Journal of Molecular Sciences. 2021; 22(3):1482. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031482
Chicago/Turabian StyleSillen, Machteld, Toshio Miyata, Douglas E. Vaughan, Sergei V. Strelkov, and Paul J. Declerck. 2021. "Structural Insight into the Two-Step Mechanism of PAI-1 Inhibition by Small Molecule TM5484" International Journal of Molecular Sciences 22, no. 3: 1482. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031482