
Estrogen Receptors Alpha and Beta Mediate Synaptic Transmission in the PFC and Hippocampus of Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Slice Preparation and Whole-Cell Recordings
2.2. Kinetics Analysis
2.3. Data Analysis
3. Results
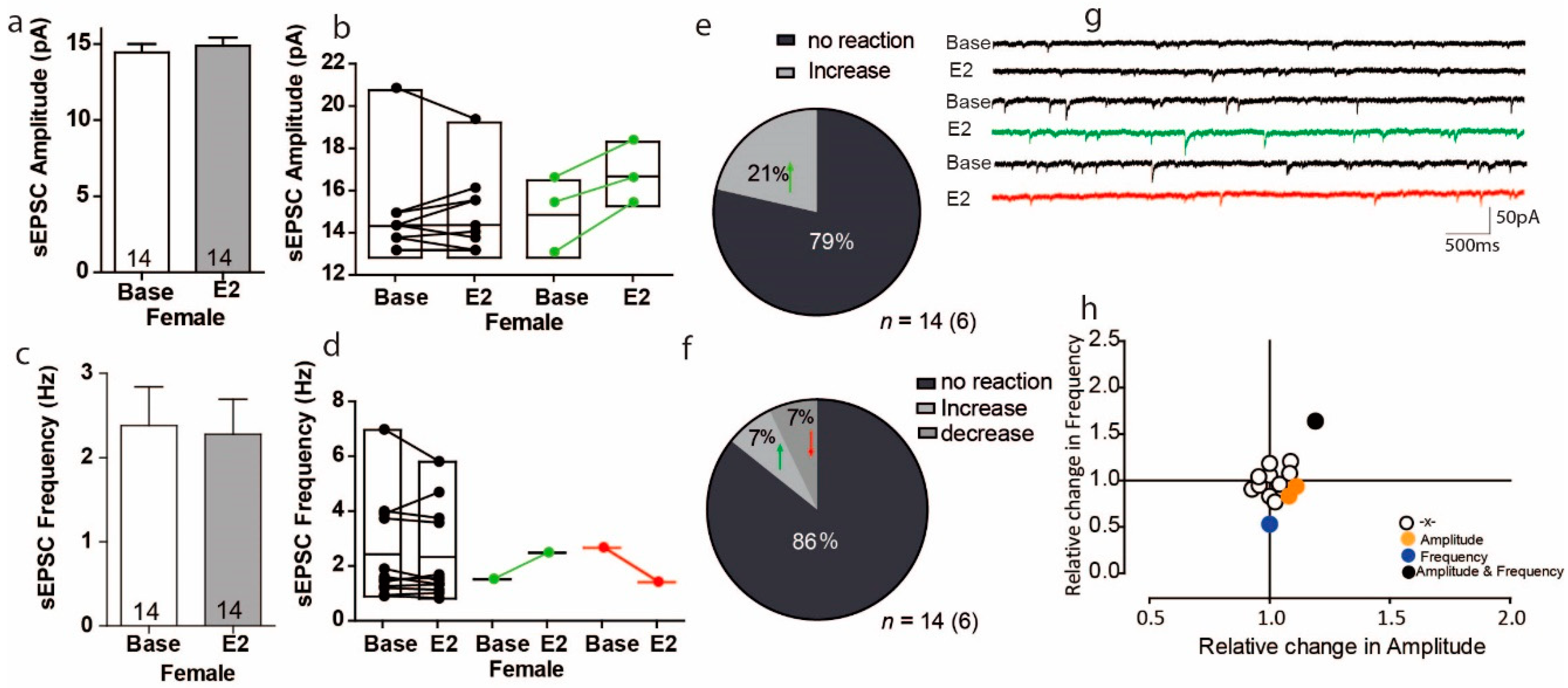
3.1. Estradiol Affects sEPSC Amplitude and Frequency in a Subpopulation of PFC Cells in Female Mice
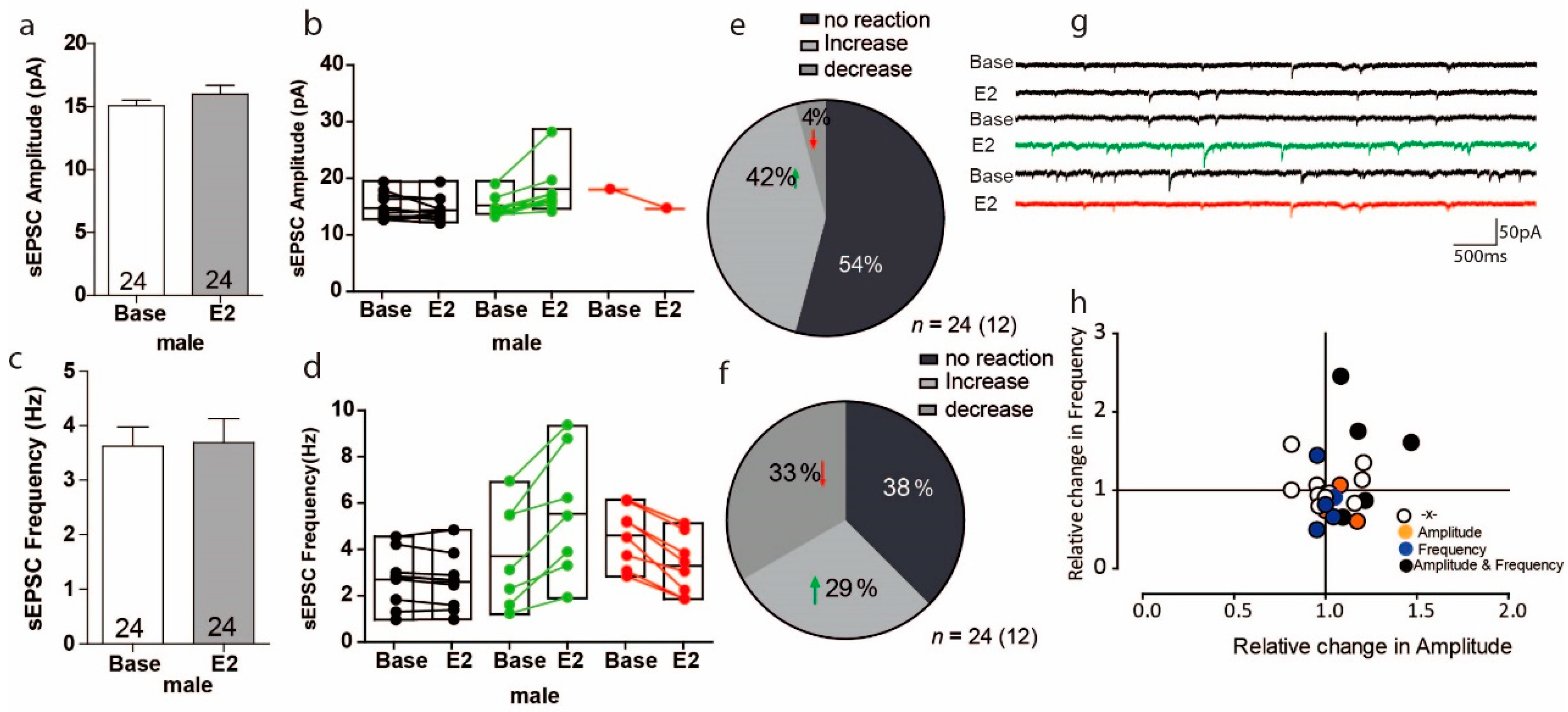
3.2. Estradiol Leads to Opposing Reactions in sEPSC Frequency in a Subset of PFC Cells in Male Mice
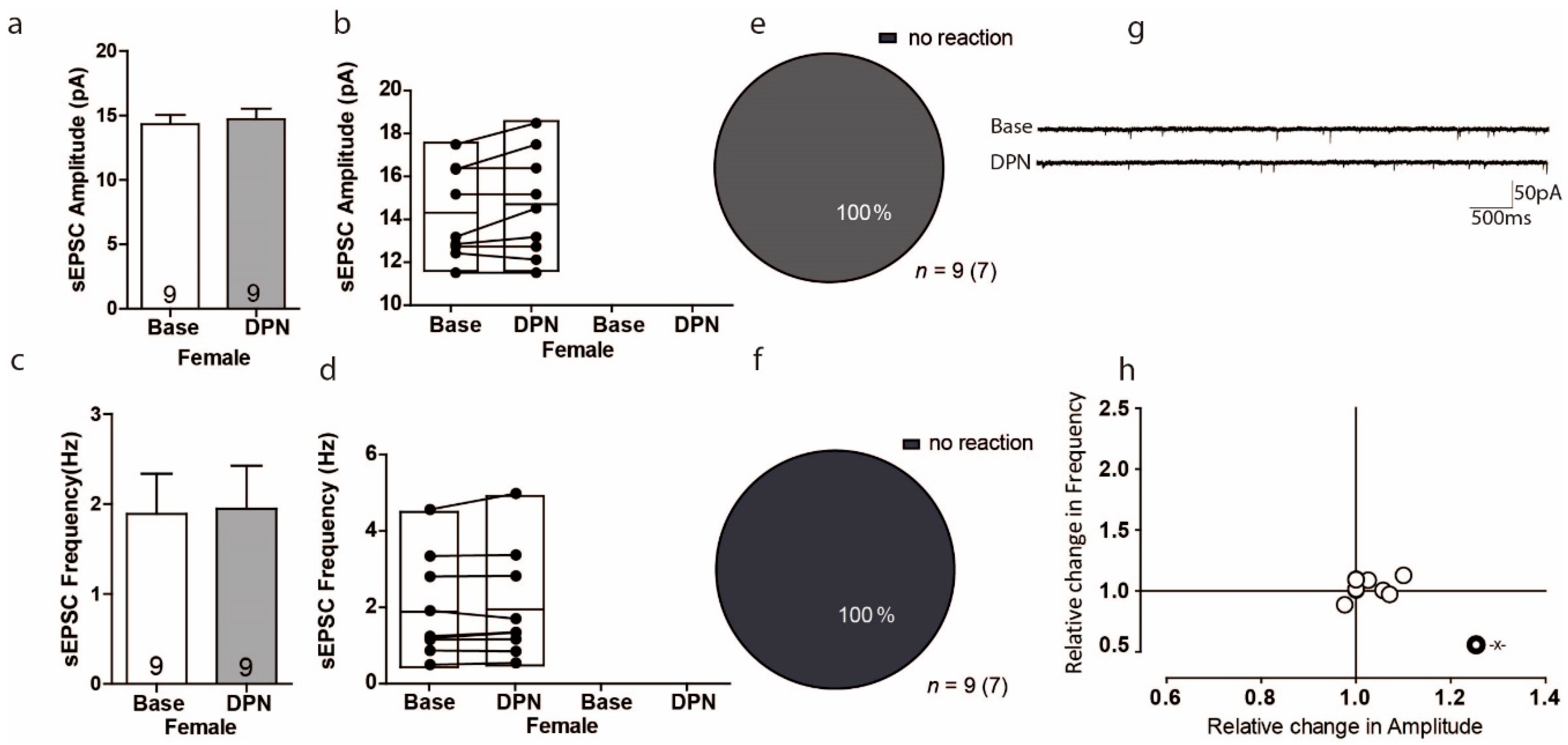
3.3. Activation of ERß Has No Influence on Excitatory Transmission in the PFC of Female Mice
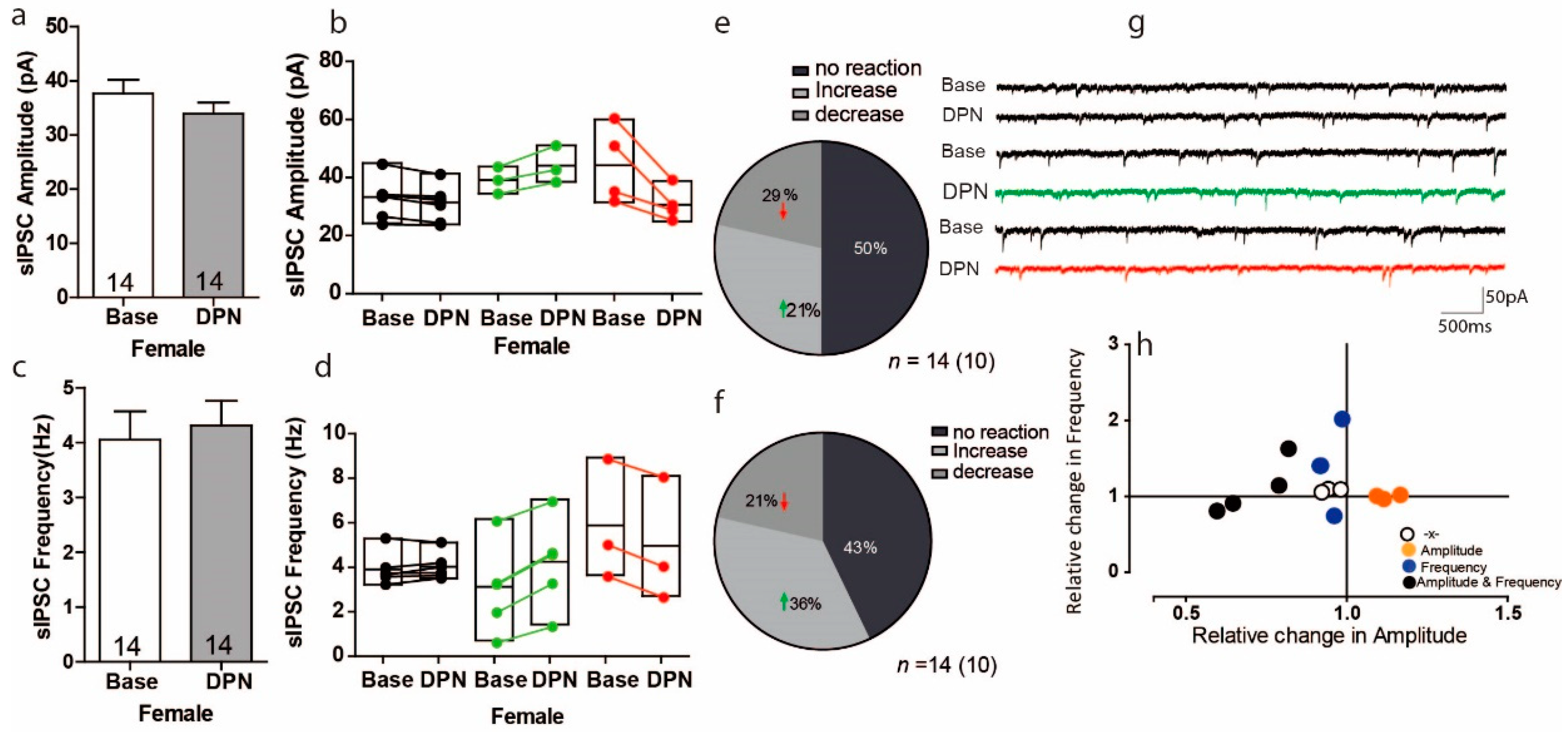
3.4. Activation of ERß on Inhibitory Transmission in the PFC of Female Mice
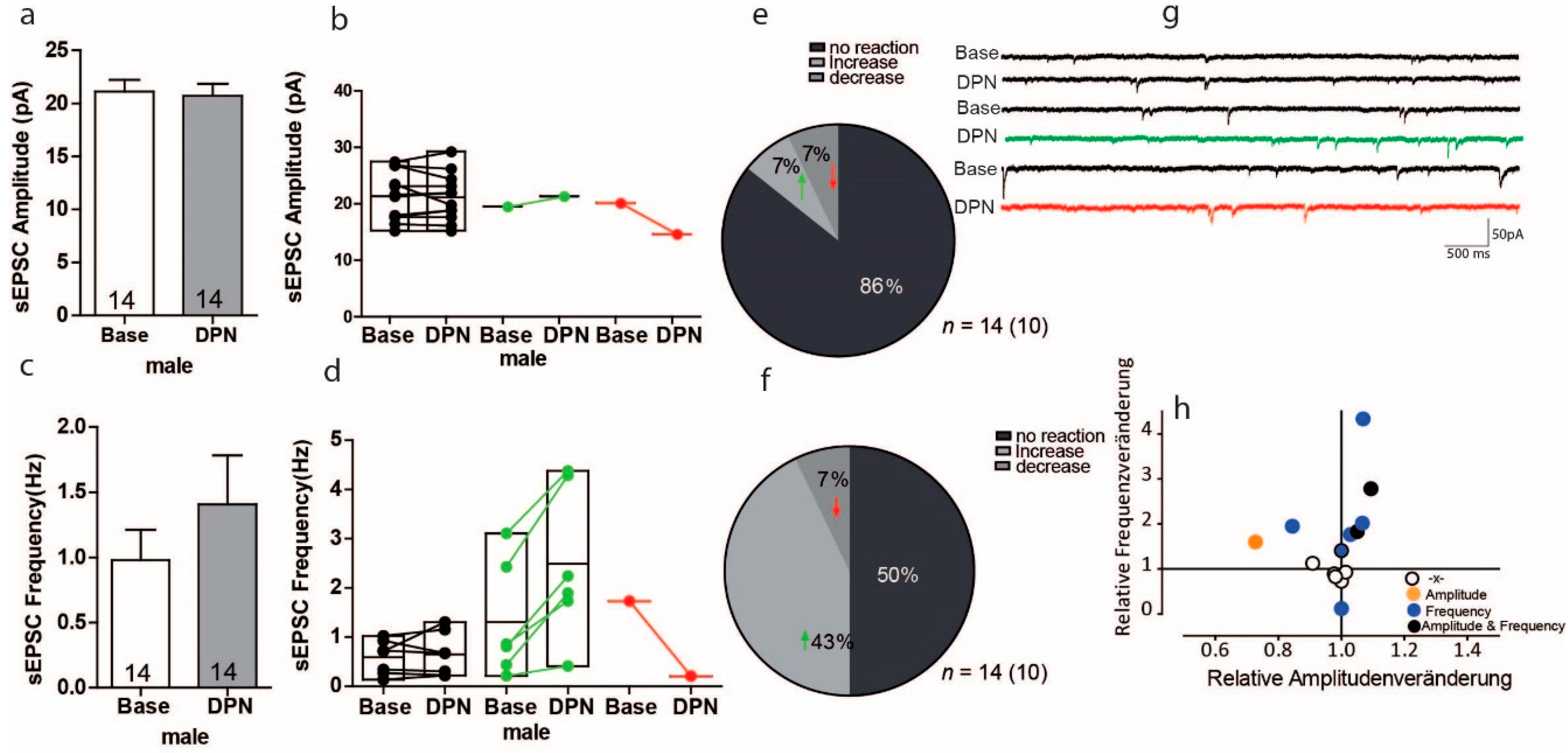
3.5. Activation of ERß on Glutamatergic Transmission in PFC of Male Mice
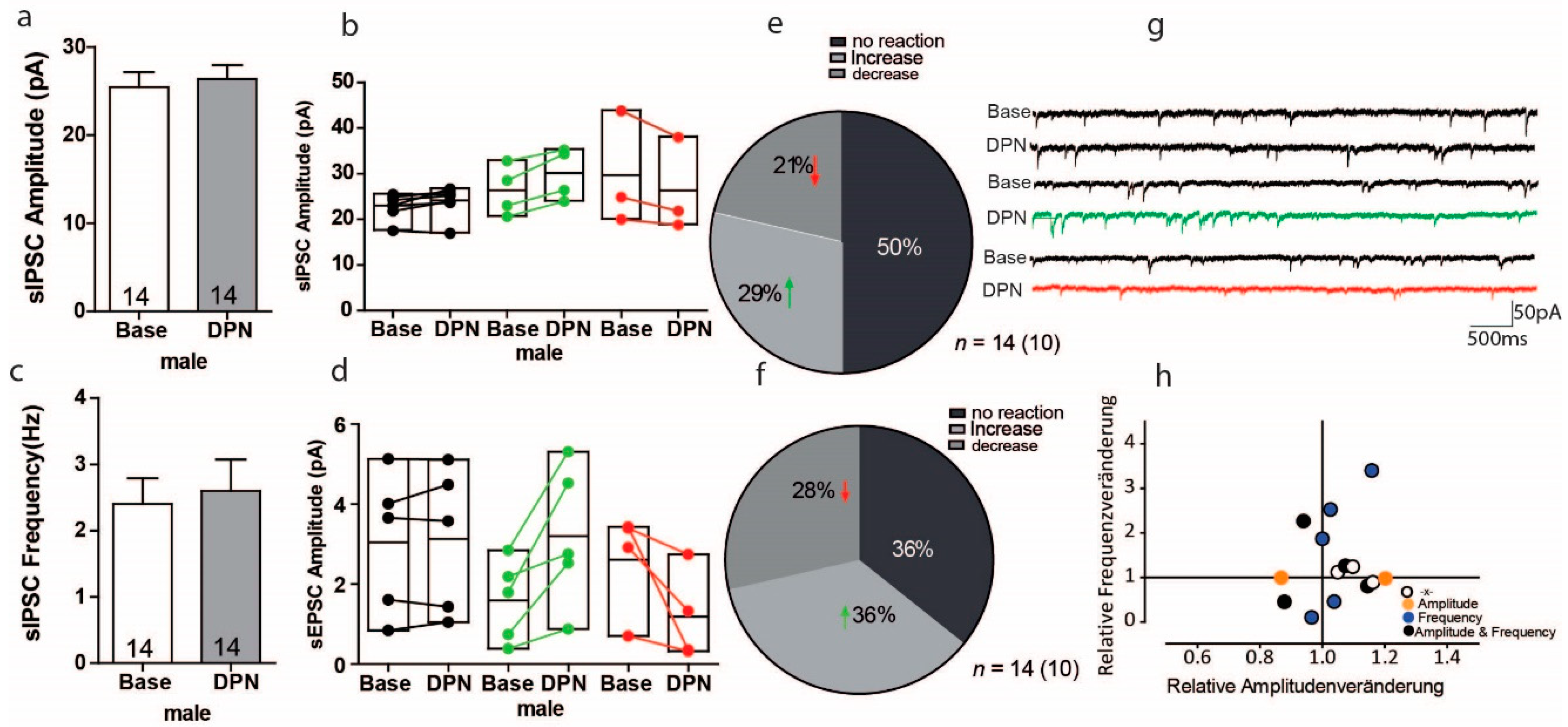
3.6. Activation of ERß on GABAergic Transmission in PFC of Male Mice
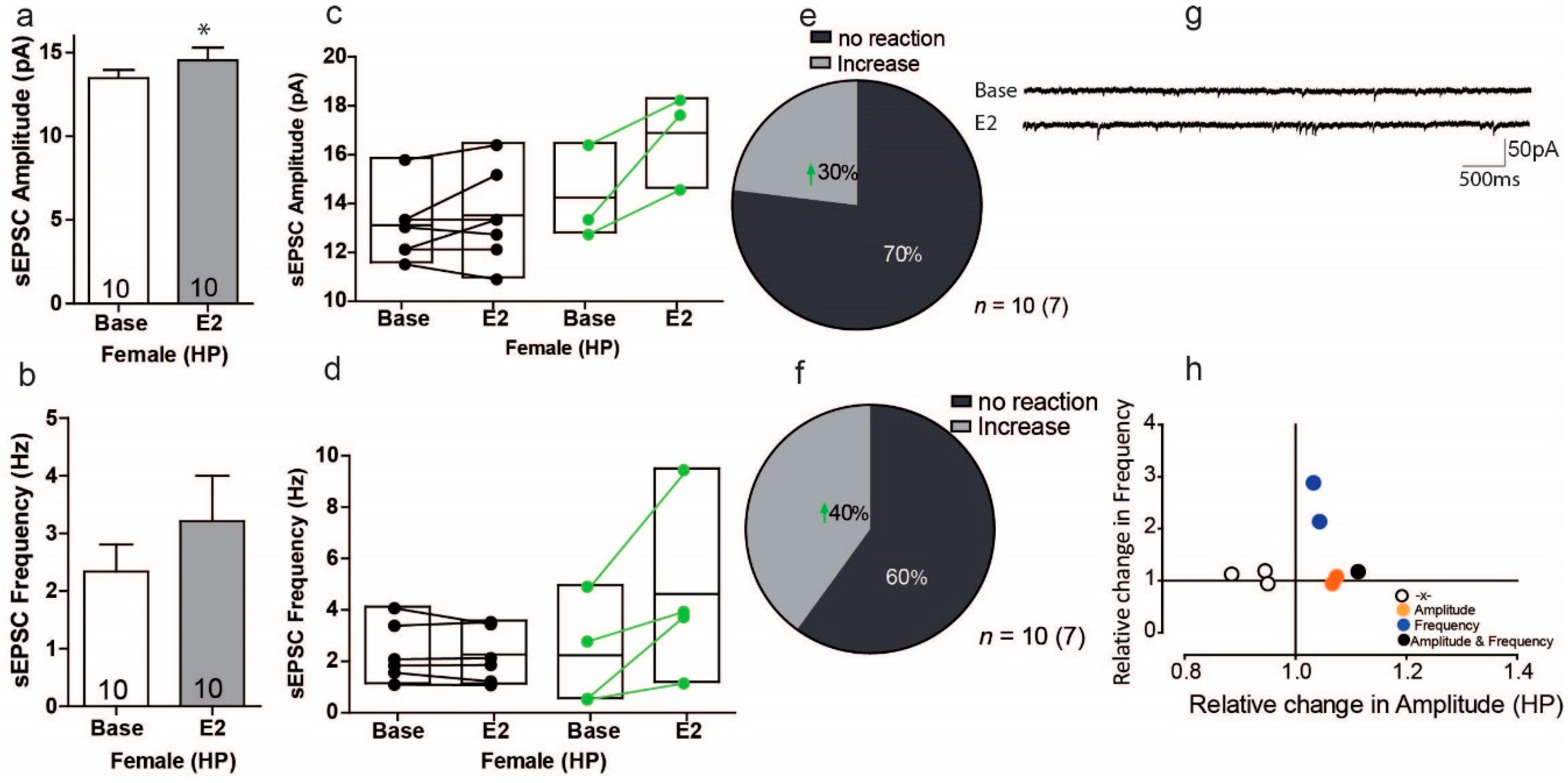
3.7. Estradiol Enhances Excitatory Synaptic Transmission in a Subgroup of Cells in the Hippocampus of Female Mice
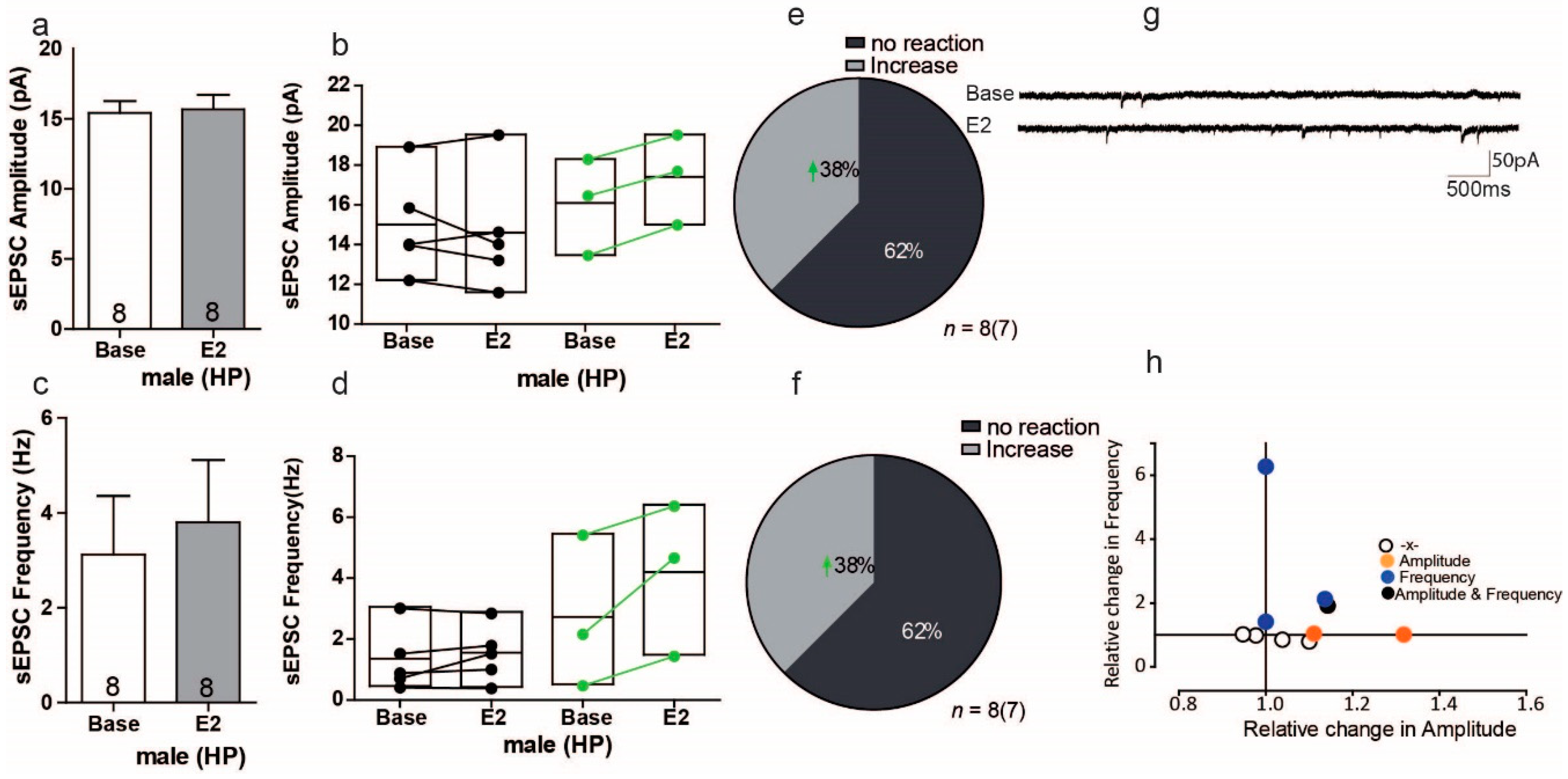
3.8. Estradiol Enhances Excitatory Synaptic Transmission in a Subgroup of Cells in the Hippocampus of Male Mice
4. Discussion
4.1. The Response to E2 in PFC Causes an Different Reaction in Synaptic Transmission in Both Sex
4.2. ERß Activation Especially Modulates Glutamatergic Transmission in the Hippocampus
4.3. E2 and ERß Activation Have Both Potentiating and Suppressive Effects on Excitatory Transmission in the PFC
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Koike, S.; Sakai, M.; Muramatsu, M. Molecular cloning and characterization of the rat estrogen receptor cDNA. Nucleic Acids Res. 1987, 15, 2499–2513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuiper, G.G.; Enmark, E.; Pelto-Huikko, M.; Nilsson, S.; Gustafsson, J.A. Cloning of a novel receptor expressed in rat prostate and ovary. Proc. NatI. Acad. Sci. USA 1996, 93, 5925–5930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulware, M.; Weick, J.P.; Becklund, B.R.; Kuo, S.P.; Groth, R.D.; Mermelstein, P.G.; Smejkalova, T.; Woolley, C.S. Estradiol acutely potentiates hippocampal excitatory synaptic transmission through a presynaptic mechanism. J. Neurosci. 2010, 30, 16137–16148. [Google Scholar]
- Boulware, M.I.; Mermelstein, P.G. The influence of estradiol on nervous system function. Drug News Perspect. 2005, 18, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Sokabe, M. Bidirectional modulatory effect of 17beta-estradiol on NMDA receptors via ERalpha and ERbeta in the dentate gyrus of juvenile male rats. Neuropharmacology 2013, 75, 262–273. [Google Scholar] [CrossRef] [PubMed]
- Chesnoy-Marchais, D.; Meillerais, A. Acute effects of estrogen on neuronal physiology. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 657–680. [Google Scholar]
- Kritzer, M.F. Regional, Laminar, and Cellular Distribution of Immunoreactivity for ER and ERss in the Cerebral Cortex of Hormonally Intact, Adult Male and Female Rats. Cereb. Cortex 2002, 12, 116–128. [Google Scholar] [CrossRef] [Green Version]
- Merchenthaler, I.; Lane, M.V.; Numan, S.; Dellovade, T.L. Distribution of estrogen receptor alpha and beta in the mouse central nervous system: In Vivo autoradiographic and immunocytochemical analyses. J. Comp. Neurol. 2004, 473, 270–291. [Google Scholar] [CrossRef]
- Milner, T.A.; Ayoola, K.; Drake, C.T.; Herrick, S.P.; Tabori, N.E.; Mcewen, B.S. Ultrastructural localization of estrogen receptor β immunoreactivity in the rat hippocampal formation. J. Comp. Neurol. 2005, 491, 81–95. [Google Scholar] [CrossRef]
- Fuster, J.M. The prefrontal cortex—An update: Time is of the essence. Neuron 2001, 30, 319–333. [Google Scholar] [CrossRef] [Green Version]
- Safari, R.; Grotefeld, K.; Kravcenko, M.; Zhang, M.; Zhang, W. Calretinin+-neurons-mediated GABAergic inhibition in mouse prefrontal cortex. Prog. Neuropsychopharmacol. Biol. Psychiatry 2019, 94, 109658. [Google Scholar] [CrossRef] [PubMed]
- Milner, T.A.; Thimpson Li Wang, G.; Kievits, J.A.; Martin, E.; Zhou, P.; McEwen, B.S.; Pfaffb, D.W.; Waters, E.M. Distribution of estrogen receptor β containing cells in the brains of bacterial artificial chromosome transgenic mice. Brain Res. 2010, 1351, 74–96. [Google Scholar] [CrossRef] [Green Version]
- Kelly, M.J.; Moss, R.L.; Dudley, C.A.; Fawcett, C.P. The specificity of the responseof preoptic-septal area neurons to estrogen: 17α-estradiol versus 17β-estradiol and the response of extrahypothalamic neurons. Exp. Brain Res. 1977, 30, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Nabekura, J.; Oomura, Y.; Minami, T.; Mizuno, Y.; Fukuda, A. Mechanism of the rapid effect of 17β–estradiol on medial amygdala neurons. Science 1986, 233, 226–228. [Google Scholar] [CrossRef] [PubMed]
- Wong, M.; Moss, R.L. Electrophysiological evidence for a rapid membrane action of the gonadal steroid, 17beta-estradiol, on CA1 pyramidal neurons of the rat hippocampus. Brain Res. 1991, 543, 148–152. [Google Scholar] [CrossRef]
- Mermelstein, P.G.; Becker, J.B.; Surneier, D.J. Estradiol Reduces Calcium Currents in Rat Neostriatal Neurons via a Membrane Receptor. J. Neurosci. 1996, 15, 595–604. [Google Scholar] [CrossRef]
- Hojo, Y.; Hattori, T.; Enami, A.; Furukawa, K.; Suzuki, K.; Ishii, H.T.; Mukai, H.; Morrison, J.H.; Janssen, W.G.M.; Kominami, S.; et al. Adult male rat hippocampus synthesizes estradiol from pregnenolone by cytochromes P45017α and P450 aromatase localized in neurons. Proc. NatI. Acad. Sci. USA 2004, 101, 865–870. [Google Scholar] [CrossRef] [Green Version]
- Tabatadze, N.; Sato, S.M.; Wooly, C.S. Quantitative analysis of long-form aromatase mRNA in the male and female rat brain. PLoS ONE 2014, 9, e100628. [Google Scholar] [CrossRef] [Green Version]
- Tabatadze, N.; Huang, G.; May, R.M.; Jain, A.; Wooley, C.S. Sex Differences in Molecular Signaling at Inhibitory Synapses in the Hippocampus. J. Neurosci. 2015, 35, 11252–11265. [Google Scholar] [CrossRef]
- McCarthy, M.M.; Woolley, C.S.; Arnold, A.P. Incorporating Sex as a Biological Variable in Neuroscience: What Do We Gain? Nat. Rev. Neurosci. 2017, 18, 707–708. [Google Scholar] [CrossRef]
- Teng, Z.; Zhang, M.; Zhao, M.; Zhang, W. Glucocorticoid Exerts Its Non-Genomic Effect on IPSC by Activation of a Phospholipase C-dependent Pathway in Prefrontal Cortex of Rats. J. Physiol. 2013, 591, 3341–3353. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Zhang, M.; Czéh, B.; Flügge, G.; Zhang, W. Tress impairs GABAergic network function in the hippocampus by activating nongenomic glucocorticoid receptors and affecting the integrity of the parvalbumin-expressing neuronal network. Neuropsychopharmacology 2010, 35, 1693–1707. [Google Scholar] [CrossRef] [PubMed]
- Dunning, D.; Hoover, C.; Soltesz, I.; Smith, M.; O’Dowd, D. GABA(A) receptor–mediated miniature postsynaptic currents and α-subunit expression in developing cortical neurons. J. Neurophysiol. 1999, 82, 3286–3297. [Google Scholar] [CrossRef] [PubMed]
- Hajos, N.; Mody, I. Synaptic communication among hippocampal interneurons: Properties of spontaneous IPSCs in morphologically identified cells. J. Neurosci. 1997, 17, 8427–8442. [Google Scholar] [CrossRef] [PubMed]
- Bálint, F.; Liposits, Z.; Farkas, I. Estrogen Receptor Beta and 2-arachidonoylglycerol Mediate the Suppressive Effects of Estradiol on Frequency of Postsynaptic Currents in Gonadotropin-Releasing Hormone Neurons of Metestrous Mice: An Acute Slice Electrophysiological Study. Front. Cell Neurosci. 2016, 10, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chesnoy-Marchais, D.; Meillerais, A. Oestradiol Rapidly Enhances Spontaneous Glycinergic Synaptic Inhibition of Hypoglossal Motoneurones Neuroendocrinol. J. Neuroendocrinol. 2007, 20, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Dik, A.; Saffari, R.; Zhang, M.; Zhang, W. Contradictory Effects of Erythropoietin on Inhibitory Synaptic Transmission in Left and Right Prelimbic Cortex of Mice. Neurobiol. Stress 2018, 9, 113–123. [Google Scholar] [CrossRef]
- Huang, G.Z.; Wooley, C.S. Estradiol Acutely Suppresses Inhibition in the Hippocampus Through a Sex-Specific Endocannabinoid and mGluR-dependent Mechanism. Neuron 2012, 74, 801–808. [Google Scholar] [CrossRef] [Green Version]
- Hart, S.A.; Snyder, M.A.; Smejkalova, T.; Woolley, C.S. Estrogen Mobilizes a Subset of Estrogen Receptor-α-Immunoreactive Vesicles in Inhibitory Presynaptic Boutons in Hippocampal CA1. J. Neurosci. 2007, 27, 2102–2111. [Google Scholar] [CrossRef]
- McCarthy, M.M.; Kaufman, L.C.; Brooks, P.J.; Pfaff, D.W.; Schwartz-Giblin, S. Estrogen modulation of nRNA levels for the two forms of glutamic acid decarboxylase (GAD) in female rat brain. J. Comp. Neurol. 1995, 360, 685–697. [Google Scholar] [CrossRef]
- Mukherjee, J.; Cardarelli, R.A.; Cantaut-Belarif, Y.; Deeb, T.Z.; Srivastava, D.P.; Tyagarajan, S.T.; Pangalos, M.N.; Triller, A.; Maguire, J.; Brandon, N.J.; et al. Estradiol modulates the efficacy of synaptic inhibition by decreasing the dwell time of GABAA receptors at inhibitory synapses. Proc. Natl. Acad. Sci. USA 2017, 114, 11763–11768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixion, C.; Sah, P.; Lynch, J.W.; Keramidas, A. GABAA receptor α and γ subunits shape synaptic currents via different mechanisms. J. Biol. Chem. 2014, 28, 5399–53411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, Q.; Moss, R.L. 17β-Estradiol Potentiates Kainate-Induced Currents via Activation of the cAMP Cascade. J. Neurosci. 1996, 16, 3620–3629. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.W.; Wang, J.M.; Chen, S.; Brinton, R.D. 1β-estradiol induced Ca2+ influx via L-type calcium channels activates the src/erk/cyclic-AMP response element binding protein signal pathway and bcl-2 expression in rat hippocampal neurons: A potential. initiation mechanism for estrogen-induced neuroprote. Neuroscience 2005, 135, 59–72. [Google Scholar] [CrossRef]
- Thomas, G.M.; Huganir, R.L. MAPK cascade signalling and synaptic plasticity. Nat. Rev. Neurosci. 2004, 5, 173–183. [Google Scholar] [CrossRef]
- Zadran, A.; Qin Qy Bi, X.N.; Zadran, H.; Kim, Y.; Foy, M.R.; Thompson, R.; Baudry, M. 17-Beta-estradiol Increases Neuronal Excitability Through MAP Kinase-Induced Calpain Activation. Proc. Natl. Acad. Sci. USA 2009, 106, 21936–21941. [Google Scholar] [CrossRef] [Green Version]
- Weller, M.; Waltereit, R. Signaling from cAMP/PKA to MAPK and synaptic plasticity. Mol. Neurobiol. 2003, 27, 99–106. [Google Scholar]
- Meitzen, J.; Mermelstein, P.G. Estrogen receptors stimulate brain region specific metabotropic glutamate receptors to rapidly initiate signal transduction pathways. J. Chem. Neuroanat. 2011, 42, 236–241. [Google Scholar] [CrossRef] [Green Version]
- Boulware, M.I.; Mermelstein, P.G. Membrane estrogen receptors activate metabotropic glutamate receptors to influence nervous system physiology. Steroids 2009, 74, 608–613. [Google Scholar] [CrossRef] [Green Version]
- Warwick, H.K.; Nahorski, S.R.; Challiss, R.A.J. Group I Metabotropic Glutamate Receptors, mGlu1a and mGlu5a, Couple to Cyclic AMP Response Element Binding Protein (CREB) Through a Common Ca2+- and Protein Kinase C-dependent Pathway. J. Neurochem. 2005, 93, 232–245. [Google Scholar] [CrossRef]
- Arnold, A.P.; Breedlove, S.M. Rganizational and activational effects of sex steroids on brain and behavior: A reanalysis. Horm. Behav. 1985, 19, 469–498. [Google Scholar] [CrossRef]
- Williams, C.L. Reevaluation of the concept of separable periods of organizational and activational actions of estrogens in development of brain and behavior. Ann. N.Y. Acad. Sci. 1986, 474, 282–292. [Google Scholar] [CrossRef] [PubMed]
- Meitzen, J.; Britson, K.A.; Tuomela, K.; Mermelstein, P.G. The Expression of Select Genes Necessary for Membrane-Associated Estrogen Receptor Signaling Differ by Sex in Adult Rat Hippocampus. Steroids 2019, 142, 21–27. [Google Scholar] [CrossRef]
- Meitzen, J.; Luoma, J.I.; Bouware, M.I.; Hedges, V.L.; Peterson, B.M.; Tuomela, K.; Britson, K.A.; Mermelstein, P.G. Palmitoylation of Estrogen Receptors Is Essential for Neuronal Membrane Signaling. Endocrinology 2013, 154, 4293–4304. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| aIPSC | Deca Time (ms) | Half-Width (ms) | Area (pA × ms) |
|---|---|---|---|
| Base | 42.2 ± 7.2 | 13.3 ± 0.5 | 968 ± 74.8 |
| DPN | 52.5 ± 4.5 * | 12.6 ± 0.6 | 1102 ± 104.7 * |
| sIPSC | Decay Time (ms) | Half-Width (ms) | Area (pA × ms) |
|---|---|---|---|
| Base | 25.0 ± 1.8 | 8.9 ± 0.4 | 724.8 ± 52.5 |
| DPN | 31.9 ± 2.8 * | 9.3 ± 0.5 | 908.7 ± 85.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Weiland, H.; Schöfbänker, M.; Zhang, W. Estrogen Receptors Alpha and Beta Mediate Synaptic Transmission in the PFC and Hippocampus of Mice. Int. J. Mol. Sci. 2021, 22, 1485. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031485
Zhang M, Weiland H, Schöfbänker M, Zhang W. Estrogen Receptors Alpha and Beta Mediate Synaptic Transmission in the PFC and Hippocampus of Mice. International Journal of Molecular Sciences. 2021; 22(3):1485. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031485
Chicago/Turabian StyleZhang, Mingyue, Hannah Weiland, Michael Schöfbänker, and Weiqi Zhang. 2021. "Estrogen Receptors Alpha and Beta Mediate Synaptic Transmission in the PFC and Hippocampus of Mice" International Journal of Molecular Sciences 22, no. 3: 1485. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031485