
Depletion of Cholesteryl Esters Causes Meibomian Gland Dysfunction-Like Symptoms in a Soat1-Null Mouse Model



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. General Characterization of Soat1-Null Mice and Their Ocular Phenotype
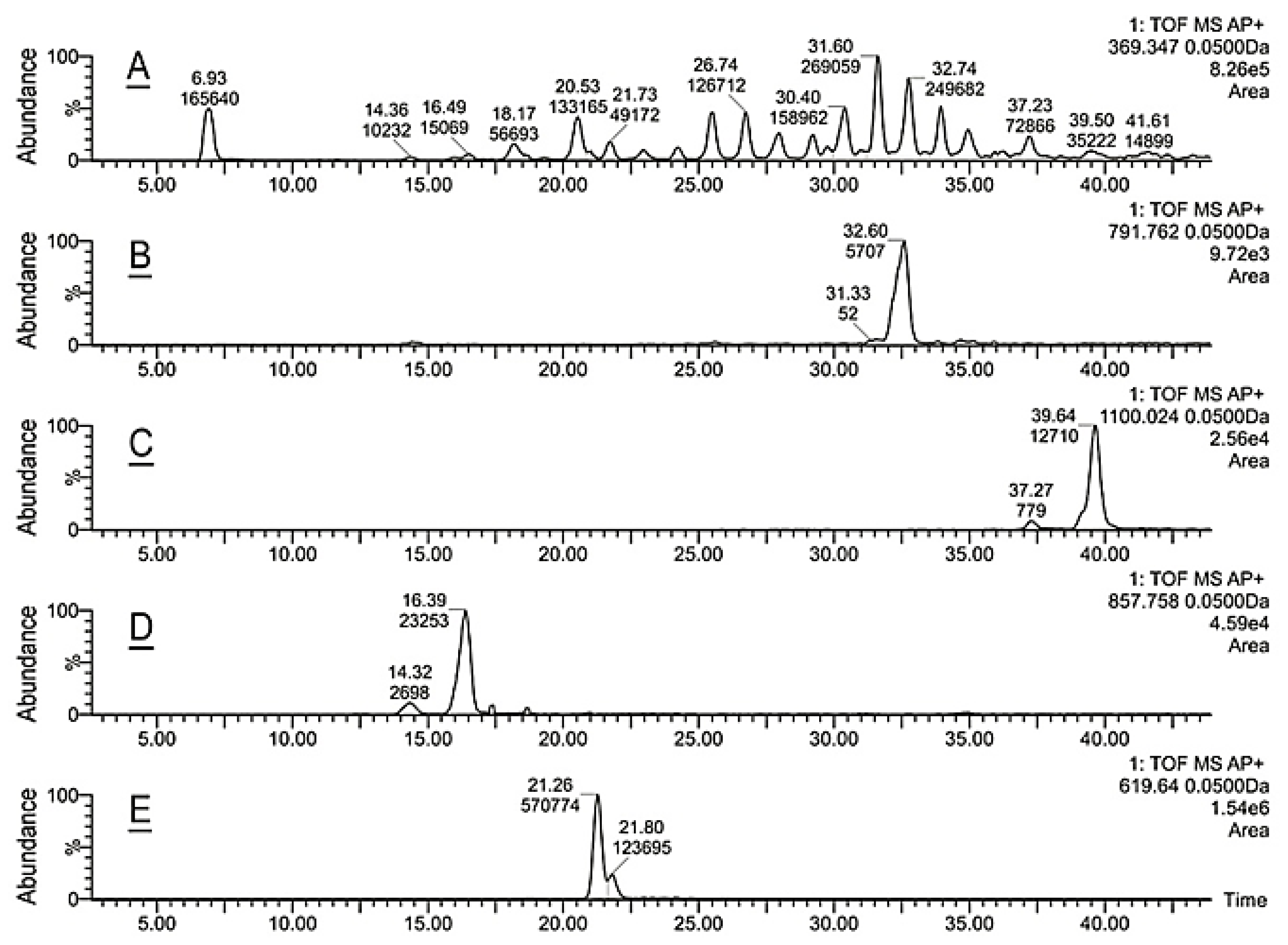
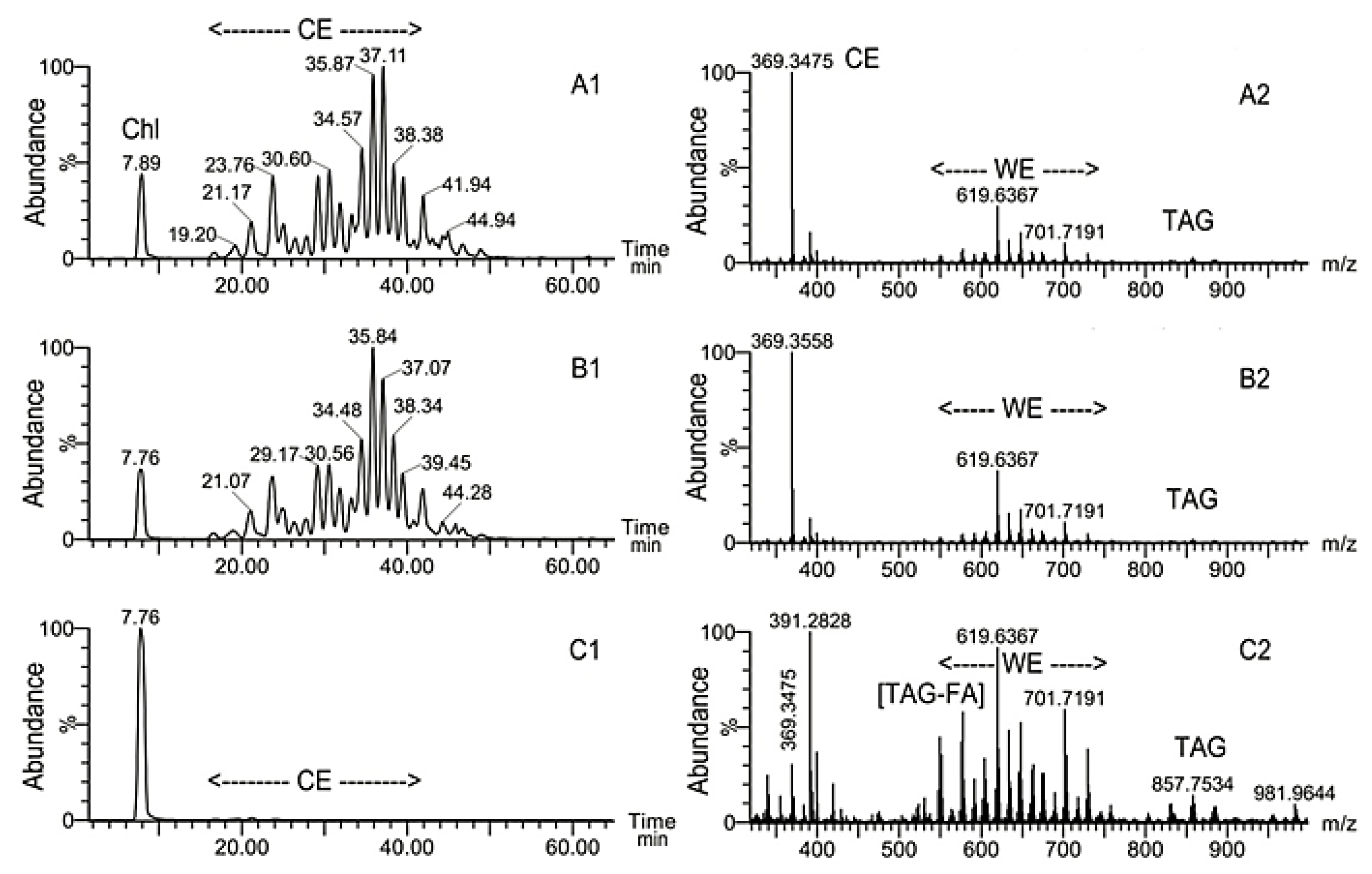
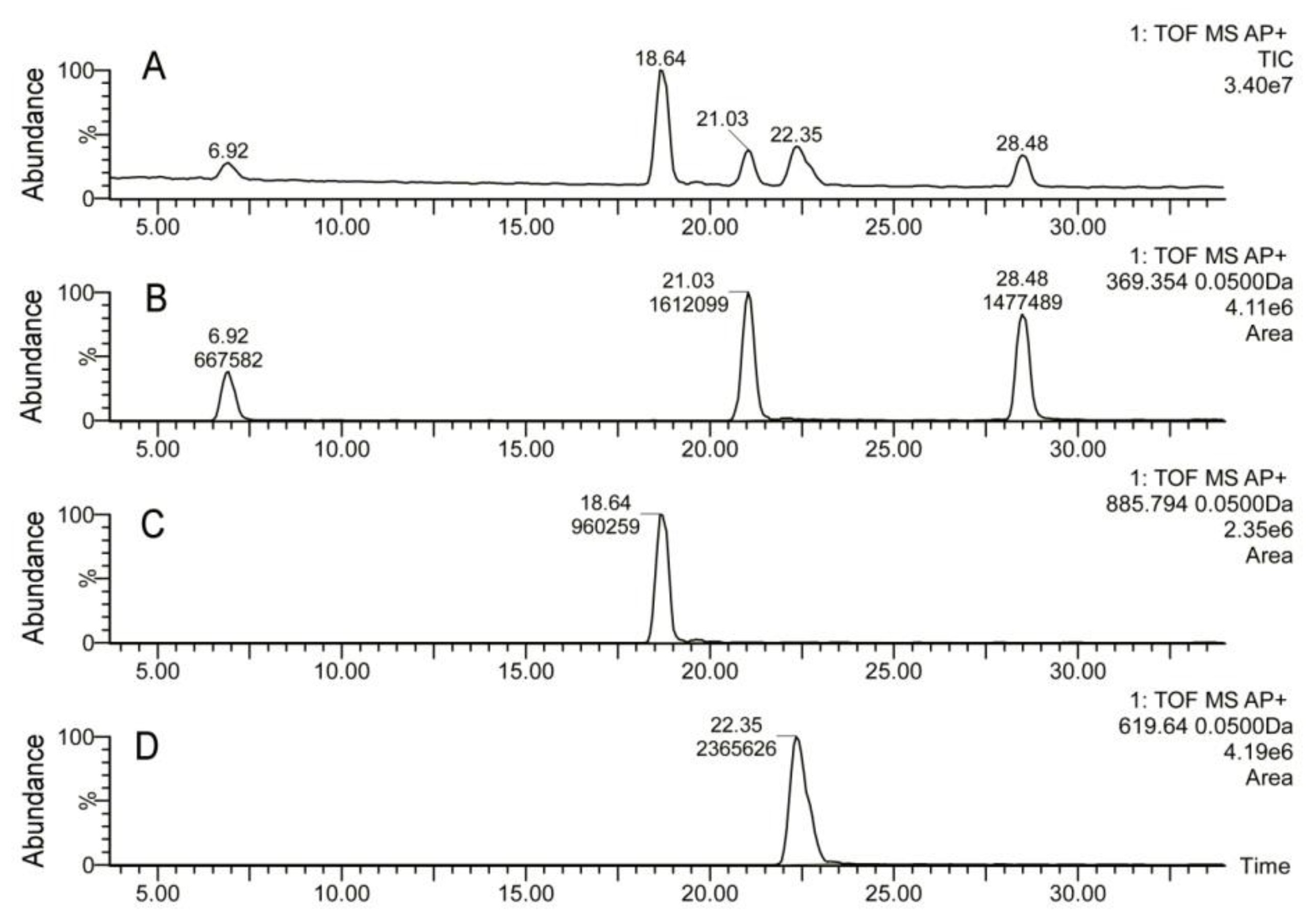
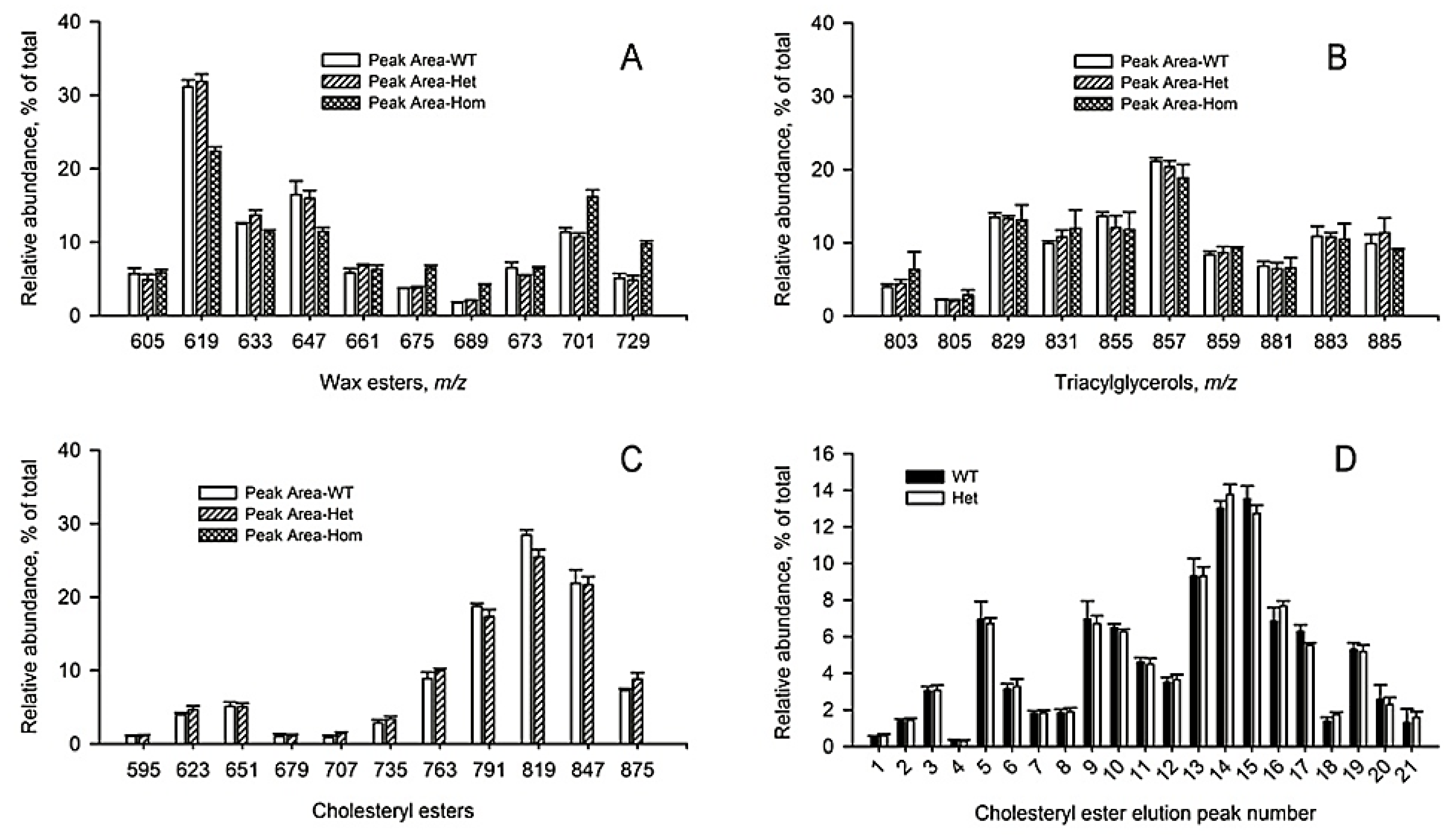
2.2. Inactivation of Soat1 Arrested Biosynthesis of Cholesteryl Esters in MG of Soat1-Null Mice
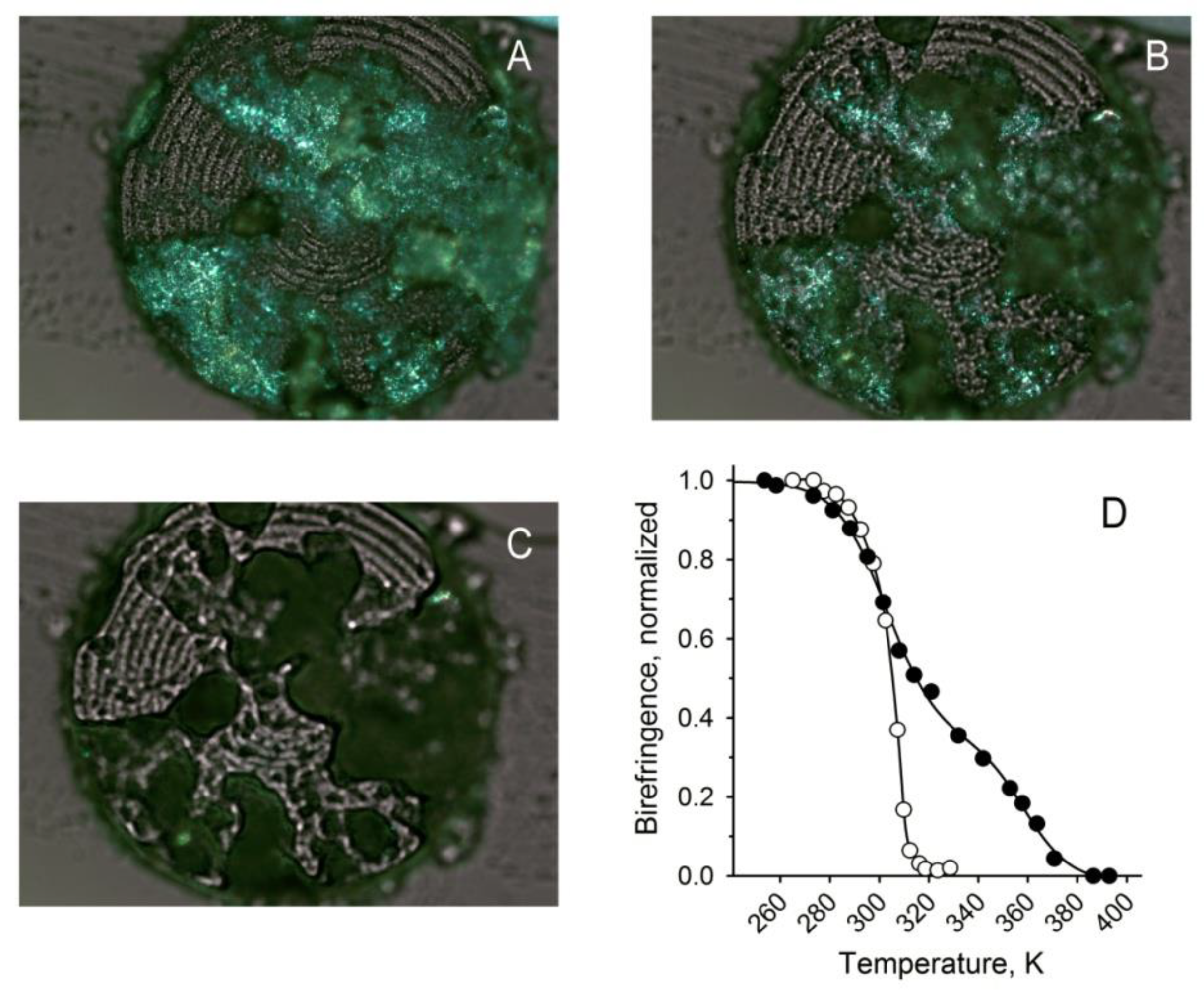
2.3. Inactivation of Soat1 Increased the Melting Temperature of Meibum
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Animals and Animal Procedures
4.3. Analytical Instrumentation and Procedures
| Crystal (solid, birefringent) → |
| Liquid crystal (partially melted, birefringent; |
| possibly, 2 forms—smectic and cholesteric) → |
| Isotropic (fluid, non-birefringent) |
4.4. Data Computation and Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Meibom, H. De Vasis Palpebrarum Novis Epistolae; Henningi Mulleri: Helmstadt, Germany, 1666. [Google Scholar]
- Knop, N.; Knop, E. [Meibomian glands. Part I: Anatomy, embryology and histology of the Meibomian glands]. Ophthalmologe 2009, 106, 872–883. [Google Scholar] [CrossRef]
- Sakaguchi, K. The precorneal tear film. Nihon Ganka Kiyo 1968, 19, 1269–1273. [Google Scholar]
- Iwata, S.; Lemp, M.A.; Holly, F.J.; Dohlman, C.H. Evaporation rate of water from the precorneal tear film and cornea in the rabbit. Investig. Ophthalmol. 1969, 8, 613–619. [Google Scholar]
- Holly, F.J. Formation and rupture of the tear film. Exp. Eye Res. 1973, 15, 515–525. [Google Scholar] [CrossRef]
- Butovich, I.A.; Lu, H.; McMahon, A.; Eule, J.C. Toward an animal model of the human tear film: Biochemical comparison of the mouse, canine, rabbit, and human meibomian lipidomes. Investig. Ophthalmol. Vis. Sci. 2012, 53, 6881–6896. [Google Scholar] [CrossRef]
- Butovich, I.A.; McMahon, A.; Wojtowicz, J.C.; Lin, F.; Mancini, R.; Itani, K. Dissecting lipid metabolism in meibomian glands of humans and mice: An integrative study reveals a network of metabolic reactions not duplicated in other tissues. Biochim. Biophys Acta. 2016, 1861, 538–553. [Google Scholar] [CrossRef] [Green Version]
- Nicolaides, N.; Kaitaranta, J.K.; Rawdah, T.N.; Macy, J.I.; Boswell, F.M., 3rd; Smith, R.E. Meibomian gland studies: Comparison of steer and human lipids. Investig. Ophthalmol. Vis. Sci. 1981, 20, 522–536. [Google Scholar]
- Butovich, I.A. Tear film lipids. Exp. Eye Res. 2013, 117, 4–27. [Google Scholar] [CrossRef] [Green Version]
- Butovich, I.A.; Wojtowicz, J.C.; Molai, M. Human tear film and meibum. Very long chain wax esters and (O-acyl)-omega-hydroxy fatty acids of meibum. J. Lipid Res. 2009, 50, 2471–2485. [Google Scholar] [CrossRef] [Green Version]
- Butovich, I.A. Meibomian glands, meibum, and meibogenesis. Exp. Eye Res. 2017, 163, 2–16. [Google Scholar] [CrossRef]
- Butovich, I.A.; Bhat, N.; Wojtowicz, J.C. Comparative Transcriptomic and Lipidomic Analyses of Human Male and Female Meibomian Glands Reveal Common Signature Genes of Meibogenesis. Int. J. Mol. Sci. 2019, 20, 4539. [Google Scholar] [CrossRef] [Green Version]
- Sassa, T.; Tadaki, M.; Kiyonari, H.; Kihara, A. Very long-chain tear film lipids produced by fatty acid elongase ELOVL1 prevent dry eye disease in mice. FASEB J. 2018, 32, 2966–2978. [Google Scholar] [CrossRef] [Green Version]
- Butovich, I.A.; Wilkerson, A.; Bhat, N.; McMahon, A.; Yuksel, S. On the pivotal role of Elovl3/ELOVL3 in meibogenesis and ocular physiology of mice. FASEB J. 2019, 33, 10034–10048. [Google Scholar] [CrossRef]
- Miyamoto, M.; Sassa, T.; Sawai, M.; Kihara, A. Lipid polarity gradient formed by omega-hydroxy lipids in tear film prevents dry eye disease. eLife 2020, 9. [Google Scholar] [CrossRef]
- Widjaja-Adhi, M.A.K.; Silvaroli, J.A.; Chelstowska, S.; Trischman, T.; Bederman, I.; Sayegh, R.; Golczak, M. Deficiency in Acyl-CoA:Wax Alcohol Acyltransferase 2 causes evaporative dry eye disease by abolishing biosynthesis of wax esters. FASEB J. 2020. [Google Scholar] [CrossRef]
- McMahon, A.; Yuksel, S.; Bhat, N.; Pham, H.; Wilkerson, A.; Butovich, I.A. Inactivation of Awat2 in mice causes loss of wax ester lipids from meibum. Investig. Ophthalmol. Vis. Sci. 2020, 61, 2632. [Google Scholar]
- Anderson, R.A.; Joyce, C.; Davis, M.; Reagan, J.W.; Clark, M.; Shelness, G.S.; Rudel, L.L. Identification of a form of acyl-CoA:cholesterol acyltransferase specific to liver and intestine in nonhuman primates. J. Biol. Chem. 1998, 273, 26747–26754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oelkers, P.; Behari, A.; Cromley, D.; Billheimer, J.T.; Sturley, S.L. Characterization of two human genes encoding acyl coenzyme A:cholesterol acyltransferase-related enzymes. J. Biol. Chem. 1998, 273, 26765–26771. [Google Scholar] [CrossRef] [Green Version]
- Joyce, C.; Skinner, K.; Anderson, R.A.; Rudel, L.L. Acyl-coenzyme A: Cholesteryl acyltransferase 2. Curr. Opin. Lipidol. 1999, 10, 89–95. [Google Scholar] [CrossRef]
- Butovich, I.A.; McMahon, A.; Wojtowicz, J.C.; Bhat, N.; Wilkerson, A. Effects of sex (or lack thereof) on meibogenesis in mice (Mus musculus): Comparative evaluation of lipidomes and transcriptomes of male and female tarsal plates. Ocul. Surf. 2019, 17, 793–808. [Google Scholar] [CrossRef]
- Butovich, I.A.; Lu, H.; McMahon, A.; Ketelson, H.; Senchyna, M.; Meadows, D.; Campbell, E.; Molai, M.; Linsenbardt, E. Biophysical and morphological evaluation of human normal and dry eye meibum using hot stage polarized light microscopy. Investig. Ophthalmol. Vis. Sci. 2014, 55, 87–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkerson, A.; Bhat, N.; Quoc Hai Pham, H.; Yuksel, S.; Butovich, I. Physiological effects of inactivation and the roles of Elovl3/ELOVL3 in maintaining ocular homeostasis. FASEB J. 2021, 35, e21327. [Google Scholar] [CrossRef] [PubMed]
- Yagyu, H.; Kitamine, T.; Osuga, J.; Tozawa, R.; Chen, Z.; Kaji, Y.; Oka, T.; Perrey, S.; Tamura, Y.; Ohashi, K.; et al. Absence of ACAT-1 attenuates atherosclerosis but causes dry eye and cutaneous xanthomatosis in mice with congenital hyperlipidemia. J. Biol. Chem. 2000, 275, 21324–21330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, B.J.; Jee, D.H.; Kim, W.J.; Shin, M.C.; Kim, E.C.; Kim, M.S.; Hwang, H.S. Direct Visualization of Continuous Meibum Secretion From the Orifices of Meibomian Glands to the Tear Film. Cornea 2019, 38, 1245–1252. [Google Scholar] [CrossRef] [PubMed]
- Ginsburg, G.S.; Atkinson, D.; Small, D.M. Physical Properties of Cholesteryl Esters. Prog. Lipid Res. 1984, 23, 135–167. [Google Scholar] [CrossRef]
- Shimobayashi, S.F.; Ohsaki, Y. Universal phase behaviors of intracellular lipid droplets. Proc. Natl. Acad. Sci. USA 2019, 116, 25440–25445. [Google Scholar] [CrossRef]
- Feigenson, G.W. Phase behavior of lipid mixtures. Nat. Chem. Biol. 2006, 2, 560–563. [Google Scholar] [CrossRef]
- Peng, S.; Guo, W.; Morrisett, J.D.; Johnstone, M.T.; Hamilton, J.A. Quantification of cholesteryl esters in human and rabbit atherosclerotic plaques by magic-angle spinning (13)C-NMR. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 2682–2688. [Google Scholar] [CrossRef] [Green Version]
- Small, D.M. George Lyman Duff memorial lecture. Progression and regression of atherosclerotic lesions. Insights from lipid physical biochemistry. Arteriosclerosis 1988, 8, 103–129. [Google Scholar] [CrossRef] [Green Version]
- Renshaw, P.F.; Janoff, A.S.; Miller, K.W. On the nature of dilute aqueous cholesterol suspensions. J. Lipid Res. 1983, 24, 47–51. [Google Scholar] [CrossRef]
- Meiner, V.L.; Cases, S.; Myers, H.M.; Sande, E.R.; Bellosta, S.; Schambelan, M.; Pitas, R.E.; McGuire, J.; Herz, J.; Farese, R.V., Jr. Disruption of the acyl-CoA:cholesterol acyltransferase gene in mice: Evidence suggesting multiple cholesterol esterification enzymes in mammals. Proc. Natl. Acad. Sci. USA 1996, 93, 14041–14046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butovich, I.A.; Suzuki, T.; Wojtowicz, J.; Bhat, N.; Yuksel, S. Comprehensive profiling of Asian and Caucasian meibomian gland secretions reveals similar lipidomic signatures regardless of ethnicity. Sci. Rep. 2020, 10, 14510. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Butovich, I.A.; Wilkerson, A.; Yuksel, S. Depletion of Cholesteryl Esters Causes Meibomian Gland Dysfunction-Like Symptoms in a Soat1-Null Mouse Model. Int. J. Mol. Sci. 2021, 22, 1583. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041583
Butovich IA, Wilkerson A, Yuksel S. Depletion of Cholesteryl Esters Causes Meibomian Gland Dysfunction-Like Symptoms in a Soat1-Null Mouse Model. International Journal of Molecular Sciences. 2021; 22(4):1583. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041583
Chicago/Turabian StyleButovich, Igor A., Amber Wilkerson, and Seher Yuksel. 2021. "Depletion of Cholesteryl Esters Causes Meibomian Gland Dysfunction-Like Symptoms in a Soat1-Null Mouse Model" International Journal of Molecular Sciences 22, no. 4: 1583. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041583