
C1q/TNF-Related Protein 3 (CTRP-3) Deficiency of Adipocytes Affects White Adipose Tissue Mass but Not Systemic CTRP-3 Concentrations

 , , ,
, , ,
Abstract
:1. Introduction
- -
- overall morphological and physiological development, especially adipose tissue and liver;
- -
- differentiation and gene expression profile of adipocytes in different adipose tissue compartments;
- -
- regulation of immunomodulatory and metabolically active adipokines, cytokines, and chemokines;
- -
- comparative transcriptome analysis of mature adipocytes and total liver by RNA-Seq.
2. Results
2.1. Biometric Analysis of Adipocyte-Specific CTRP-3 KO Mice
General Phenotype and Body Weight
2.2. Adipose Tissue Morphology
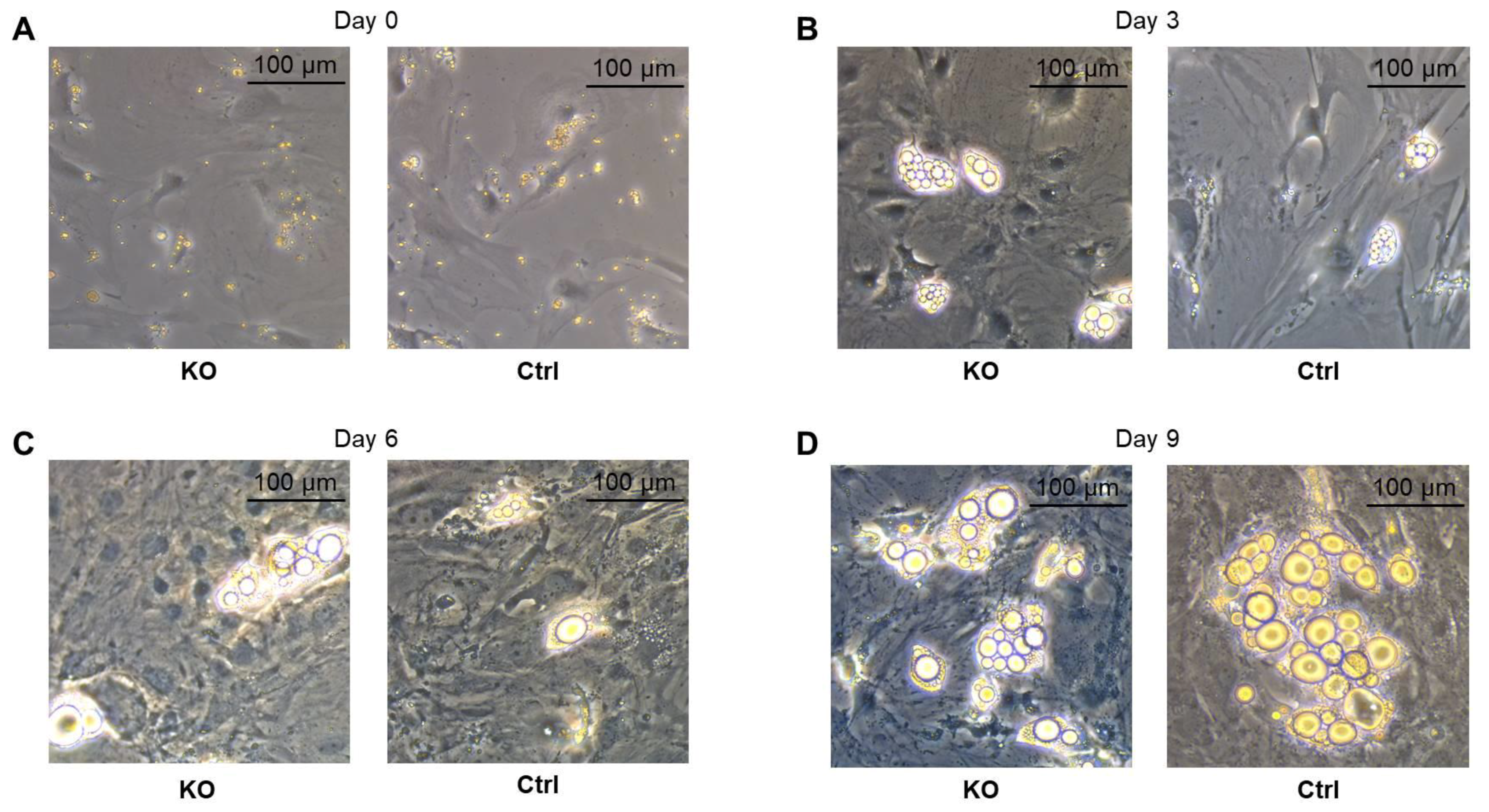
2.3. Differentiation of Primary Preadipocytes into Mature Adipocytes
2.4. Liver Morphology
2.5. Serum Adipokine Concentrations in CTRP-3 KO Mice
2.6. Adipokine and Cytokine Gene Expression Studies in Murine Adipose Tissue
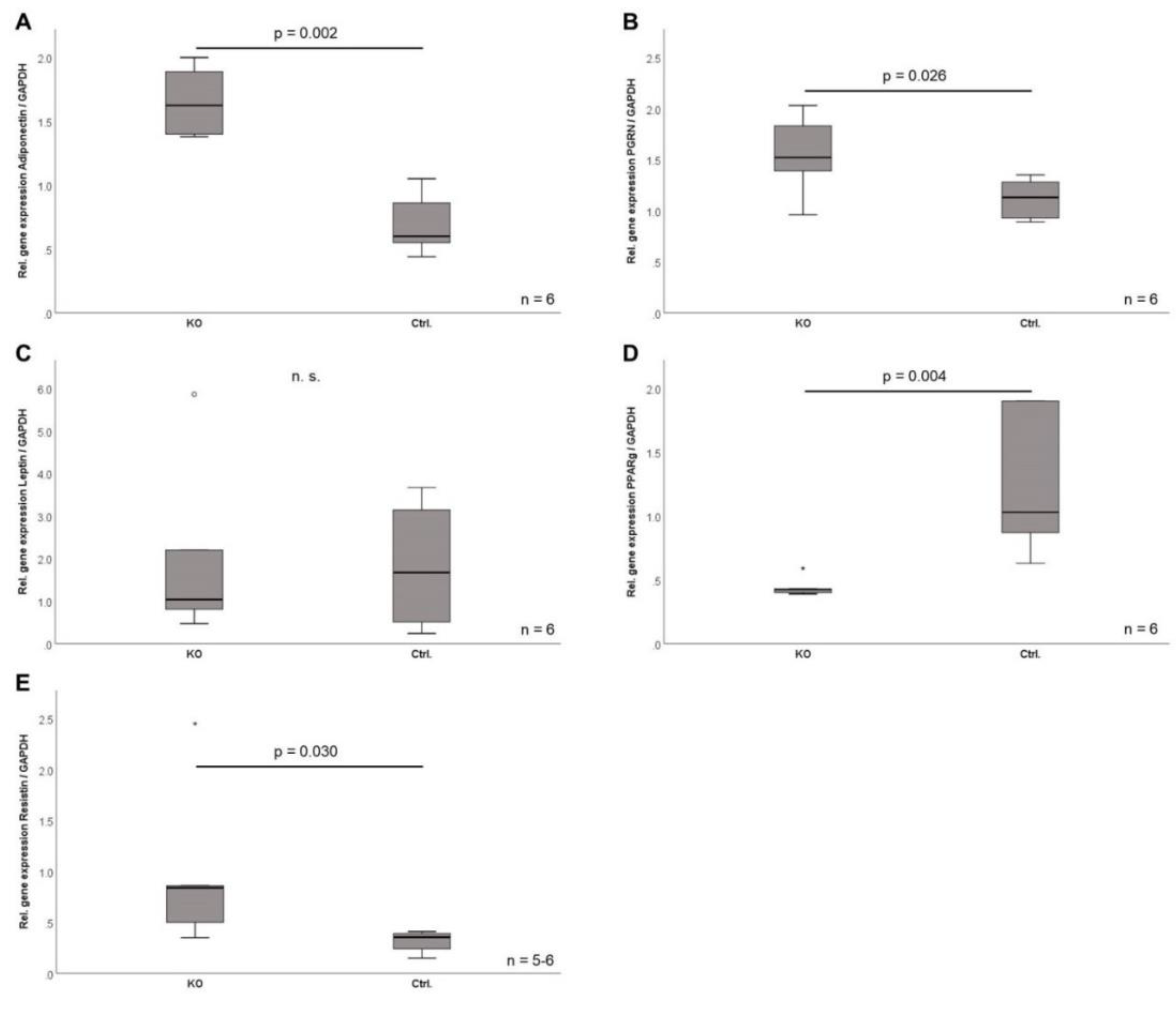
2.7. Adipokine Gene Expression in CTRP-3 Deficient Adipocytes
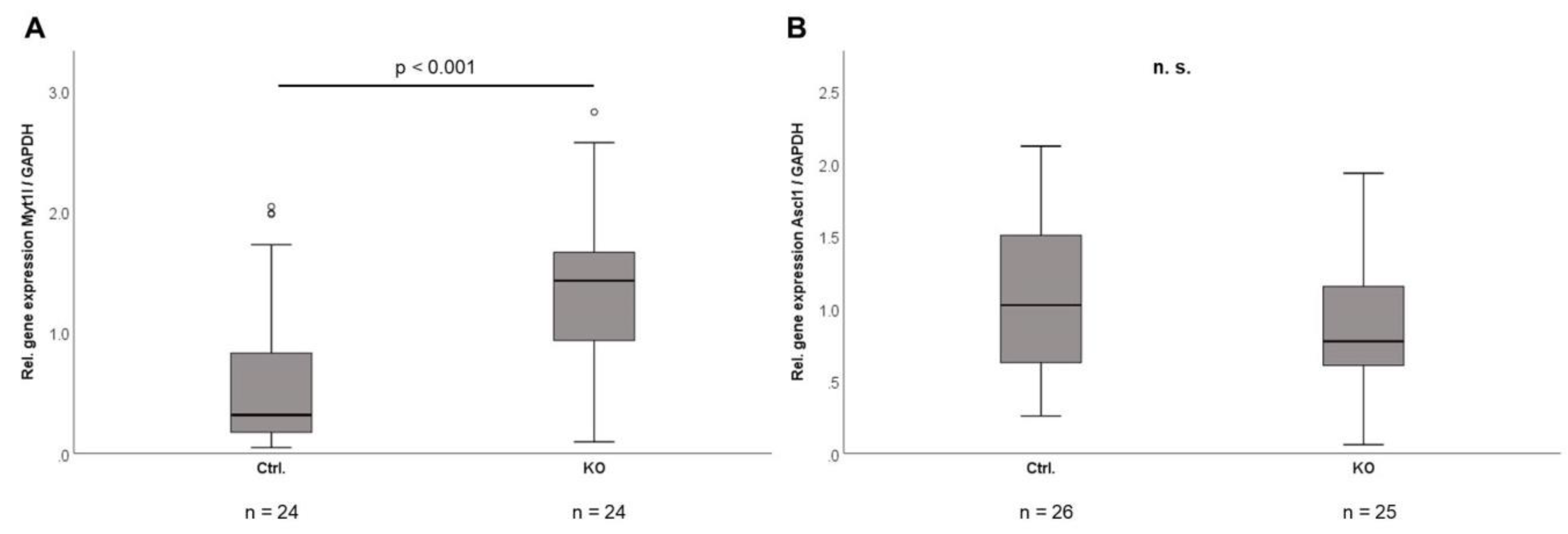
2.8. Transcriptome Sequencing of Isolated Mature Adipocytes in CTRP-3 KO vs. Control Mice
2.9. Transcriptome Sequencing of Total Liver Tissue from CTRP-3 KO vs. Control Mice
3. Discussion
4. Materials and Methods
4.1. Quantification of Adipokine Concentrations in Murine Serum
4.2. Adipose Tissue and Adipocyte mRNA Extraction
4.3. Real-Time PCR Analysis of Adipose Tissue mRNA Expression
4.4. Animals
4.4.1. Adipocyte-Specific CTRP-3 Knockout
4.4.2. Analysis of Hepatic and Adipose Tissue Integrity
4.5. Glycogen Analysis
4.6. Isolation and Cell Culture of Primary Murine Adipocytes
4.7. Comparative Transcriptome Analysis of Single-Cell Adipocytes and Liver Tissue
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Trayhurn, P.; Beattie, J.H. Physiological role of adipose tissue: White adipose tissue as an endocrine and secretory organ. Proc. Nutr. Soc. 2001, 60, 329–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kershaw, E.E.; Flier, J.S. Adipose tissue as an endocrine organ. J. Clin. Endocrinol. Metab. 2004, 89, 2548–2556. [Google Scholar] [CrossRef] [PubMed]
- Abella, V.; Scotece, M.; Conde, J.; Pino, J.; Gonzalez-Gay, M.A.; Gomez-Reino, J.J.; Mera, A.; Lago, F.; Gomez, R.; Gualillo, O. Leptin in the interplay of inflammation, metabolism and immune system disorders. Nat. Rev. Rheumatol. 2017, 13, 100–109. [Google Scholar] [CrossRef]
- Wang, Z.V.; Scherer, P.E. Adiponectin, the past two decades. J. Mol. Cell Biol. 2016, 8, 93–100. [Google Scholar] [CrossRef]
- Fruhbeck, G.; Catalan, V.; Rodriguez, A.; Ramirez, B.; Becerril, S.; Salvador, J.; Portincasa, P.; Colina, I.; Gomez-Ambrosi, J. Involvement of the leptin-adiponectin axis in inflammation and oxidative stress in the metabolic syndrome. Sci. Rep. 2017, 7, 6619. [Google Scholar] [CrossRef]
- Hotamisligil, G.S. Inflammation, metaflammation and immunometabolic disorders. Nature 2017, 542, 177–185. [Google Scholar] [CrossRef]
- Schaffler, A.; Scholmerich, J. Innate immunity and adipose tissue biology. Trends Immunol. 2010, 31, 228–235. [Google Scholar] [CrossRef]
- Mastrocola, R.; Aragno, M.; Alloatti, G.; Collino, M.; Penna, C.; Pagliaro, P. Metaflammation: Tissue-Specific Alterations of the NLRP3 Inflammasome Platform in Metabolic Syndrome. Curr. Med. Chem. 2018, 25, 1294–1310. [Google Scholar] [CrossRef]
- Schaffler, A.; Scholmerich, J.; Salzberger, B. Adipose tissue as an immunological organ: Toll-like receptors, C1q/TNFs and CTRPs. Trends Immunol. 2007, 28, 393–399. [Google Scholar] [CrossRef]
- Seldin, M.M.; Tan, S.Y.; Wong, G.W. Metabolic function of the CTRP family of hormones. Rev. Endocr. Metab. Disord. 2014, 15, 111–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, G.W.; Krawczyk, S.A.; Kitidis-Mitrokostas, C.; Revett, T.; Gimeno, R.; Lodish, H.F. Molecular, biochemical and functional characterizations of C1q/TNF family members: Adipose-tissue-selective expression patterns, regulation by PPAR-gamma agonist, cysteine-mediated oligomerizations, combinatorial associations and metabolic functions. Biochem. J. 2008, 416, 161–177. [Google Scholar] [CrossRef] [Green Version]
- Akiyama, H.; Furukawa, S.; Wakisaka, S.; Maeda, T. CTRP3/cartducin promotes proliferation and migration of endothelial cells. Mol. Cell Biochem. 2007, 304, 243–248. [Google Scholar] [CrossRef]
- Schmid, A.; Kopp, A.; Aslanidis, C.; Wabitsch, M.; Muller, M.; Schaffler, A. Regulation and function of C1Q/TNF-related protein-5 (CTRP-5) in the context of adipocyte biology. Exp. Clin. Endocrinol. Diabetes 2013, 121, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Schmid, A.; Kopp, A.; Hanses, F.; Bala, M.; Muller, M.; Schaffler, A. The novel adipokine C1q/TNF-related protein-3 is expressed in human adipocytes and regulated by metabolic and infection-related parameters. Exp. Clin. Endocrinol. Diabetes 2012, 120, 611–617. [Google Scholar] [CrossRef] [PubMed]
- Schmid, A.; Kopp, A.; Hanses, F.; Karrasch, T.; Schaffler, A. C1q/TNF-related protein-3 (CTRP-3) attenuates lipopolysaccharide (LPS)-induced systemic inflammation and adipose tissue Erk-1/-2 phosphorylation in mice in vivo. Biochem. Biophys. Res. Commun. 2014, 452, 8–13. [Google Scholar] [CrossRef]
- Peterson, J.M.; Seldin, M.M.; Wei, Z.; Aja, S.; Wong, G.W. CTRP3 attenuates diet-induced hepatic steatosis by regulating triglyceride metabolism. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 305, G214–G224. [Google Scholar] [CrossRef] [Green Version]
- Peterson, J.M.; Wei, Z.; Wong, G.W. C1q/TNF-related protein-3 (CTRP3), a novel adipokine that regulates hepatic glucose output. J. Biol. Chem. 2010, 285, 39691–39701. [Google Scholar] [CrossRef] [Green Version]
- Compton, S.A.; Cheatham, B. CTRP-3: Blocking a toll booth to obesity-related inflammation. Endocrinology 2010, 151, 5095–5097. [Google Scholar] [CrossRef] [Green Version]
- Petersen, P.S.; Wolf, R.M.; Lei, X.; Peterson, J.M.; Wong, G.W. Immunomodulatory roles of CTRP3 in endotoxemia and metabolic stress. Physiol. Rep. 2016, 4. [Google Scholar] [CrossRef]
- Murayama, M.A.; Kakuta, S.; Maruhashi, T.; Shimizu, K.; Seno, A.; Kubo, S.; Sato, N.; Saijo, S.; Hattori, M.; Iwakura, Y. CTRP3 plays an important role in the development of collagen-induced arthritis in mice. Biochem. Biophys. Res. Commun. 2014, 443, 42–48. [Google Scholar] [CrossRef] [Green Version]
- Kopp, A.; Bala, M.; Buechler, C.; Falk, W.; Gross, P.; Neumeier, M.; Scholmerich, J.; Schaffler, A. C1q/TNF-related protein-3 represents a novel and endogenous lipopolysaccharide antagonist of the adipose tissue. Endocrinology 2010, 151, 5267–5278. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Wang, D.M.; Luo, L.L. CTRP3 acts as a novel regulator in depressive-like behavior associated inflammation and apoptosis by meditating p38 and JNK MAPK signaling. Biomed. Pharmacother. 2019, 120, 109489. [Google Scholar] [CrossRef] [PubMed]
- Wolf, R.M.; Lei, X.; Yang, Z.C.; Nyandjo, M.; Tan, S.Y.; Wong, G.W. CTRP3 deficiency reduces liver size and alters IL-6 and TGFbeta levels in obese mice. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E332–E345. [Google Scholar] [CrossRef] [Green Version]
- Maeda, T.; Wakisaka, S. Deficiency of C1q/TNF-related protein 3 (CTRP3) decreases adipose tissue weight in diet-induced obesity mice. Mol. Biol. Rep. 2020, 10.1007/s11033-020-05905-6. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.G.; Yuan, Y.P.; Zhang, X.; Xu, S.C.; Kong, C.Y.; Song, P.; Li, N.; Tang, Q.Z. C1q-tumour necrosis factor-related protein-3 exacerbates cardiac hypertrophy in mice. Cardiovasc. Res. 2019, 115, 1067–1077. [Google Scholar] [CrossRef]
- Zhang, B.; Zhang, P.; Tan, Y.; Feng, P.; Zhang, Z.; Liang, H.; Duan, W.; Jin, Z.; Wang, X.; Liu, J.; et al. C1q-TNF-related protein-3 attenuates pressure overload-induced cardiac hypertrophy by suppressing the p38/CREB pathway and p38-induced ER stress. Cell Death Dis. 2019, 10, 520. [Google Scholar] [CrossRef] [Green Version]
- Pagano, E.; Calvo, J.C. ErbB2 and EGFR are downmodulated during the differentiation of 3T3-L1 preadipocytes. J. Cell Biochem. 2003, 90, 561–572. [Google Scholar] [CrossRef]
- Huang, C.K.; Tsai, M.Y.; Luo, J.; Kang, H.Y.; Lee, S.O.; Chang, C. Suppression of androgen receptor enhances the self-renewal of mesenchymal stem cells through elevated expression of EGFR. Biochim. Biophys. Acta 2013, 1833, 1222–1234. [Google Scholar] [CrossRef] [Green Version]
- Brock, J.; Schmid, A.; Karrasch, T.; Pfefferle, P.; Schlegel, J.; Busse, I.; Hauenschild, A.; Schmidt, B.; Koukou, M.; Arapogianni, E.; et al. Progranulin serum levels and gene expression in subcutaneous vs visceral adipose tissue of severely obese patients undergoing bariatric surgery. Clin. Endocrinol. 2019, 91, 400–410. [Google Scholar] [CrossRef]
- Petersen, G.F.; Hilbert, B.J.; Trope, G.D.; Kalle, W.H.; Strappe, P.M. Direct Conversion of Equine Adipose-Derived Stem Cells into Induced Neuronal Cells Is Enhanced in Three-Dimensional Culture. Cell Reprogram. 2015, 17, 419–426. [Google Scholar] [CrossRef]
- Yang, Y.; Jiao, J.; Gao, R.; Yao, H.; Sun, X.F.; Gao, S. Direct conversion of adipocyte progenitors into functional neurons. Cell Reprogram. 2013, 15, 484–489. [Google Scholar] [CrossRef]
- Zhang, P. Analysis of mouse liver glycogen content. Bio-Protocol. 2012, 2, e186. [Google Scholar] [CrossRef] [Green Version]
- Zaitsu, H.; Serrero, G. Pedersen fetuin contains three adipogenic factors with distinct biochemical characteristics. J. Cell Physiol. 1990, 144, 485–491. [Google Scholar] [CrossRef]
- Bachmeier, M.; Loffler, G. Adipogenic activities in commercial preparations of fetuin. Horm. Metab. Res. 1994, 26, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Cornelius, P.; MacDougald, O.A.; Lane, M.D. Regulation of adipocyte development. Annu. Rev. Nutr. 1994, 14, 99–129. [Google Scholar] [CrossRef]
- Green, H.; Kehinde, O. Formation of normally differentiated subcutaneous fat pads by an established preadipose cell line. J. Cell Physiol. 1979, 101, 169–171. [Google Scholar] [CrossRef] [PubMed]
- Green, H.; Kehinde, O. An established preadipose cell line and its differentiation in culture. II. Factors affecting the adipose conversion. Cell 1975, 5, 19–27. [Google Scholar] [CrossRef]
- Green, H.; Meuth, M. An established pre-adipose cell line and its differentiation in culture. Cell 1974, 3, 127–133. [Google Scholar] [CrossRef]
- MacDougald, O.A.; Lane, M.D. Transcriptional regulation of gene expression during adipocyte differentiation. Annu. Rev. Biochem. 1995, 64, 345–373. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tissue Weight | Control Mice n | CTRP-3 KO Mice n | p | ||
|---|---|---|---|---|---|
| Total body weight (g) | 33.4 ± 5.9 | 65 | 31.2 ± 5.5 | 62 | 0.034 * |
| Intra-abdominal adipose tissue (mg) | 1361 ± 678 | 61 | 1051 ± 677 | 59 | 0.016 * |
| Subcutaneous adipose tissue (mg) | 1163 ± 561 | 60 | 915 ± 500 | 55 | 0.019 * |
| Perirenal adipose tissue (mg) | 637 ± 315 | 60 | 475 ± 266 | 53 | 0.006 * |
| Brown adipose tissue (mg) | 342 ± 133 | 60 | 284 ± 109 | 56 | 0.017 |
| Liver weight (mg) | 1600 ± 300 | 62 | 1568 ± 314 | 59 | n.s. |
| Adipokine serum concentrations | |||||
| CTRP-3 (pg/mL) | 5485 ± 1034 | 12 | 5835 ± 857 | 14 | n.s. |
| Adiponectin (µg/mL) | 7.35 ± 2.30 | 30 | 6.59 ± 2.18 | 28 | n.s. |
| Leptin (ng/mL) | 47.2 ± 25.0 | 28 | 32.8 ± 22.7 | 26 | 0.048 * |
| Resistin (ng/mL) | 91.0 ± 33.4 | 30 | 100.0 ± 52.9 | 29 | n.s. |
| Progranulin (ng/mL) | 979 ± 382 | 65 | 983 ± 347 | 61 | n.s. |
| Quantitative gene expression in visceral adipose tissue | |||||
| Adipokines/adipocyte differentiation | |||||
| Adiponectin | 29 | unchanged | 28 | n.s. | |
| Leptin | 29 | unchanged | 28 | n.s. | |
| Resistin | 29 | unchanged | 28 | n.s. | |
| PGRN | 33 | unchanged | 33 | n.s. | |
| PPARγ | 29 | unchanged | 28 | n.s. | |
| Lipases | |||||
| ATGL | 27 | unchanged | 29 | n.s. | |
| HSL | 30 | unchanged | 29 | n.s. | |
| M1 polarization marker | |||||
| MCP-1 | 32 | decreased | 31 | 0.013 * | |
| TNFα | 34 | unchanged | 33 | n.s. | |
| IL-6 | 31 | unchanged | 29 | n.s. | |
| RANTES | 32 | unchanged | 32 | n.s. | |
| CD80 | 27 | unchanged | 28 | n.s. | |
| M2 polarization marker | |||||
| CD206 | 26 | increased | 27 | 0.005 * | |
| Arginase-1 | 27 | unchanged | 25 | n.s. | |
| IL-10 | 27 | unchanged | 28 | n.s. | |
| A | |||||
| Upregulated Genes | |||||
| Stable_id | Name | Description | log2FC | p Value | fdr |
| ENSMUSG00000061911 | Myt1l | myelin transcription factor 1-like [Source:MGI Symbol;Acc:MGI:1100511] | 1.307465982 | 3.00303 × 10–11 | 4.6544 × 10–7 |
| ENSMUSG00000076258 | Gm23935 | predicted gene, 23935 [Source:MGI Symbol;Acc:MGI:5453712] | 0.987707951 | 7.74724 × 10–10 | 6.00373 × 10–6 |
| ENSMUSG00000087943 | Gm24245 | predicted gene, 24245 [Source:MGI Symbol;Acc:MGI:5454022] | 0.955497729 | 1.47437 × 10–8 | 7.61711 × 10–5 |
| ENSMUSG00000096887 | Gm20594 | predicted gene, 20594 [Source:MGI Symbol;Acc:MGI:5295700] | 0.895398238 | 1.35443 × 10–5 | 0.029989107 |
| ENSMUSG00000086324 | Gm15564 | predicted gene 15564 [Source:MGI Symbol;Acc:MGI:3783013] | 0.865783185 | 3.09707 × 10–6 | 0.009600304 |
| ENSMUSG00000076281 | Gm24270 | predicted gene, 24270 [Source:MGI Symbol;Acc:MGI:5454047] | 0.813376051 | 4.23787 × 10–6 | 0.010947126 |
| ENSMUSG00000088246 | Gm25911 | predicted gene, 25911 [Source:MGI Symbol;Acc:MGI:5455688] | 0.77894899 | 1.97157 × 10–5 | 0.038196634 |
| Downregulated Genes | |||||
| Stable_id | Name | Description | log2FC | p Value | fdr |
| ENSMUSG00000026107 | Nabp1 | nucleic acid binding protein 1 [Source:MGI Symbol;Acc:MGI:1923258] | −0.691646461 | 2.44321 × 10–6 | 0.009466828 |
| B | |||||
| Upregulated Genes | |||||
| Stable_id | Name | Description | log2FC | p Value | fdr |
| ENSMUSG00000041293 | Adgrf1 | adhesion G protein-coupled receptor F1 [Source:MGI Symbol;Acc:MGI:1924846] | 1.239856554 | 1.13594 × 10–9 | 7.16403 × 10–6 |
| ENSMUSG00000020122 | Egfr | epidermal growth factor receptor [Source:MGI Symbol;Acc:MGI:95294] | 0.829495923 | 3.30804 × 10–5 | 0.029803838 |
| Downregulated Genes | |||||
| Stable_id | Name | Description | log2FC | p Value | fdr |
| ENSMUSG00000022129 | Dct | dopachrome tautomerase [Source:MGI Symbol;Acc:MGI:102563] | −0.822078018 | 5.91212 × 10–7 | 0.001623412 |
| ENSMUSG00000012187 | Mogat1 | monoacylglycerol O-acyltransferase 1 [Source:MGI Symbol;Acc:MGI:1915643] | −0.847754458 | 2.81302 × 10–5 | 0.027588471 |
| ENSMUSG00000073460 | Pnldc1 | poly(A)-specific ribonuclease (PARN)-like domain containing 1 [Source:MGI Symbol;Acc:MGI:2685159] | −0.852074869 | 7.12385 × 10–6 | 0.008985543 |
| ENSMUSG00000061780 | Cfd | complement factor D (adipsin) [Source:MGI Symbol;Acc:MGI:87931] | −0.897079711 | 4.32255 × 10–6 | 0.006290967 |
| ENSMUSG00000022096 | Hr | hairless [Source:MGI Symbol;Acc:MGI:96223] | −0.93125267 | 9.04715 × 10–7 | 0.001623412 |
| ENSMUSG00000089665 | Fcor | Foxo1 corepressor [Source:MGI Symbol;Acc:MGI:1915484] | −0.952767753 | 9.43844 × 10–7 | 0.001623412 |
| ENSMUSG00000012123 | Aim1l | absent in melanoma 1-like [Source:MGI Symbol;Acc:MGI:1334463] | −0.975100738 | 1.54815 × 10–7 | 0.00058582 |
| ENSMUSG00000002831 | Plin4 | perilipin 4 [Source:MGI Symbol;Acc:MGI:1929709] | −0.999361647 | 7.5625 × 10–7 | 0.001623412 |
| ENSMUSG00000034634 | Ly6d | lymphocyte antigen 6 complex, locus D [Source:MGI Symbol;Acc:MGI:96881] | −1.007058408 | 7.44224 × 10–7 | 0.001623412 |
| ENSMUSG00000046415 | B430212C06Rik | RIKEN cDNA B430212C06 gene [Source:MGI Symbol;Acc:MGI:2442134] | −1.108254865 | 7.43126 × 10–9 | 3.51498 × 10–5 |
| ENSMUSG00000039092 | Sptlc3 | serine palmitoyltransferase, long chain base subunit 3 [Source:MGI Symbol;Acc:MGI:2444678] | −1.247137857 | 2.08131 × 10–10 | 1.96892 × 10–6 |
| Target Gene | Forward Primer 5′–3′ | Reverse Primer 5′–3′ |
|---|---|---|
| Adiponectin | AGGGAGAGAAAGGAGATGCAG | CAGACTTGGGCTCCCACCTC |
| Arg1 | TTTTAGGGTTACGGCCGGTG | CCTCGAGGCTGTCCTTTTGA |
| Ascl1 | GGAACAAGAGCTGCTGGACT | GTTTTTCTGCCTCCCCATTT |
| ATGL | GAGGAATGGCCTACTGAACC | AGGCTGCAATTGATCCTCCT |
| CD80 | TCTGTAAGCACAGAAGCTGTTT | GGCTTCACCTAGAGAACCGT |
| CD206 | GGCTGATTACGAGCAGTGGA | CATCACTCCAGGTGAACCCC |
| GAPDH | TGTCCGTCGTGGATCTGAC | AGGGAGATGCTCAGTGTTGG |
| HSL | TGAGATGGTAACTGTGAGCC | ACTGAGATTGAGGTGCTGTC |
| IL6 | AGTTGCCTTCTTGGGACTGA | TCCACGATTTCCCAGAGAAC |
| IL10 | ATTTGAATTCCCTGGGTGAGAAG | CACAGGGGAGAAATCGATGACA |
| Leptin | AGGATCTGAGGGGTGATGTG | AGGTGACCAAGGTGGCATAG |
| MCP-1 | AGGTCCCTGTCATGCTTCTG | TCTGGACCCATTCCTTCTTG |
| Myt1l | CGAGCCAGCAACGGTATAGA | CCATTGTGGTGATCTGAGTTCTG |
| RANTES | CCACTTCTTCTCTGGGTTGG | GTGCCCACGTCAAGGAGTAT |
| Resistin | TGCTAAGTCCTCTGCCACGTA | TCAACTGACCGACATCAGGA |
| PPARγ | TTATAGCTGTCATTATTCTCAGTGGAG | CAGCACCTGCCTTAAGTTGA |
| Progranulin | GTGGCTGGCCTGGAGAAGAT | CCTCACAGCACACAGCATGG |
| TNFα | CCAGACCCTCACACTCAGATCAT | ACTCCAGCTGCTCCTCCACTT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schmid, A.; Roderfeld, M.; Gehl, J.; Roeb, E.; Nist, A.; Chung, H.-R.; Stiewe, T.; Karrasch, T.; Schäffler, A. C1q/TNF-Related Protein 3 (CTRP-3) Deficiency of Adipocytes Affects White Adipose Tissue Mass but Not Systemic CTRP-3 Concentrations. Int. J. Mol. Sci. 2021, 22, 1670. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041670
Schmid A, Roderfeld M, Gehl J, Roeb E, Nist A, Chung H-R, Stiewe T, Karrasch T, Schäffler A. C1q/TNF-Related Protein 3 (CTRP-3) Deficiency of Adipocytes Affects White Adipose Tissue Mass but Not Systemic CTRP-3 Concentrations. International Journal of Molecular Sciences. 2021; 22(4):1670. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041670
Chicago/Turabian StyleSchmid, Andreas, Martin Roderfeld, Jonas Gehl, Elke Roeb, Andrea Nist, Ho-Ryun Chung, Thorsten Stiewe, Thomas Karrasch, and Andreas Schäffler. 2021. "C1q/TNF-Related Protein 3 (CTRP-3) Deficiency of Adipocytes Affects White Adipose Tissue Mass but Not Systemic CTRP-3 Concentrations" International Journal of Molecular Sciences 22, no. 4: 1670. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041670