
Atorvastatin Inhibits Endothelial PAI-1-Mediated Monocyte Migration and Alleviates Radiation-Induced Enteropathy

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Atorvastatin Mitigates Radiation-Induced Endothelial Cell Dysfunction
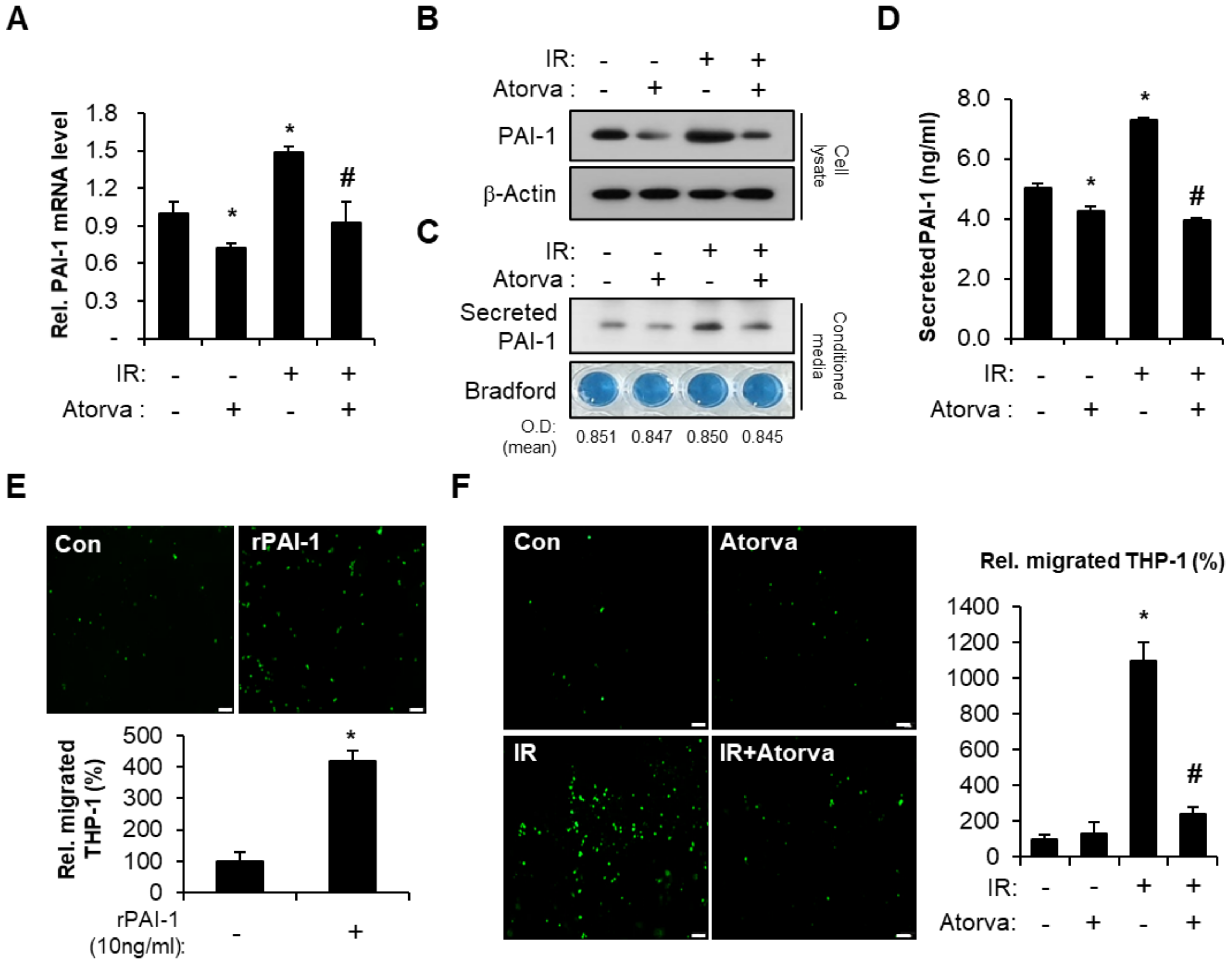
2.2. Atorvastatin Inhibits PAI-1 Expression in Irradiated HUVECs and Suppresses Transendothelial Migration of THP-1 Cells
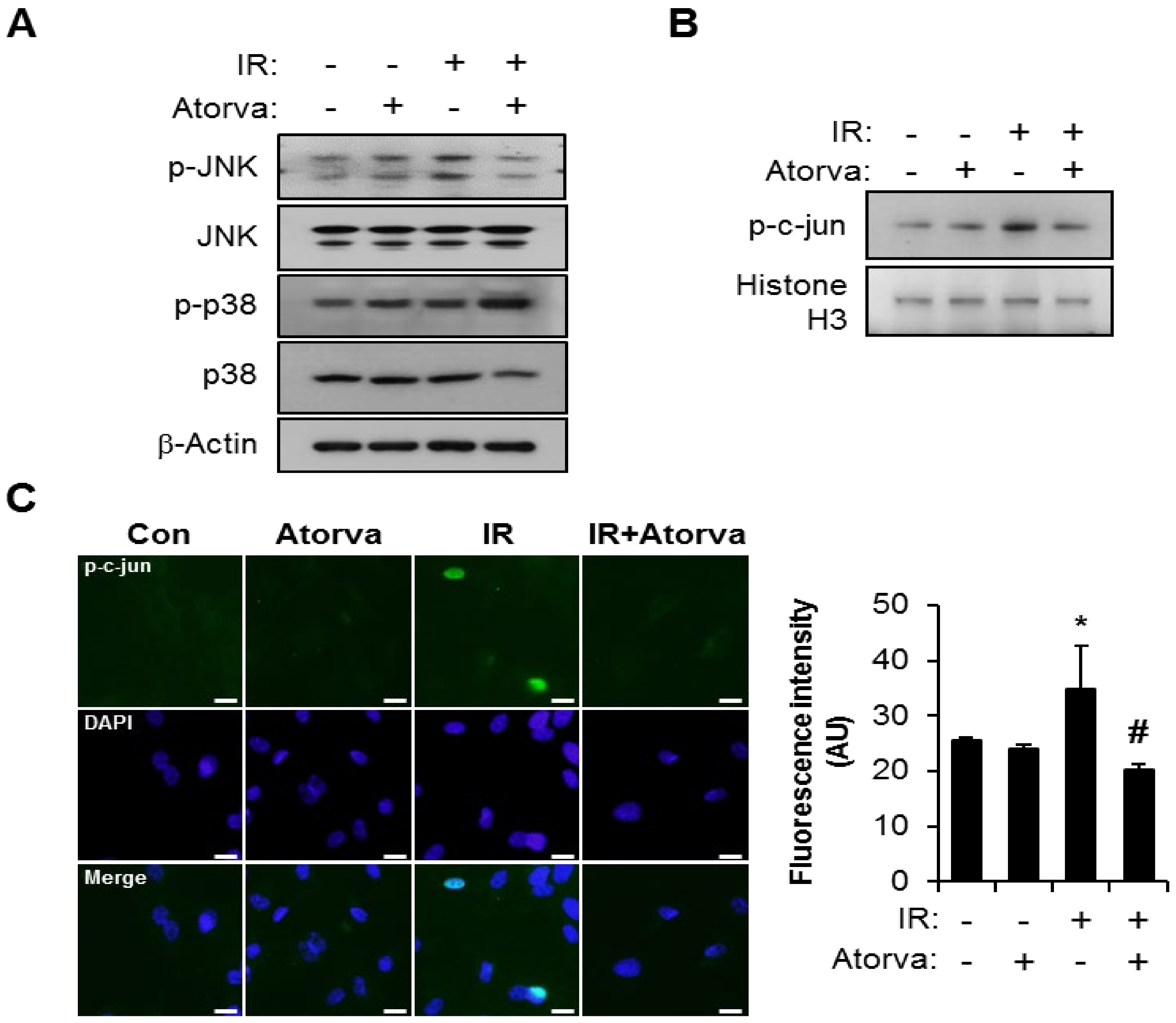
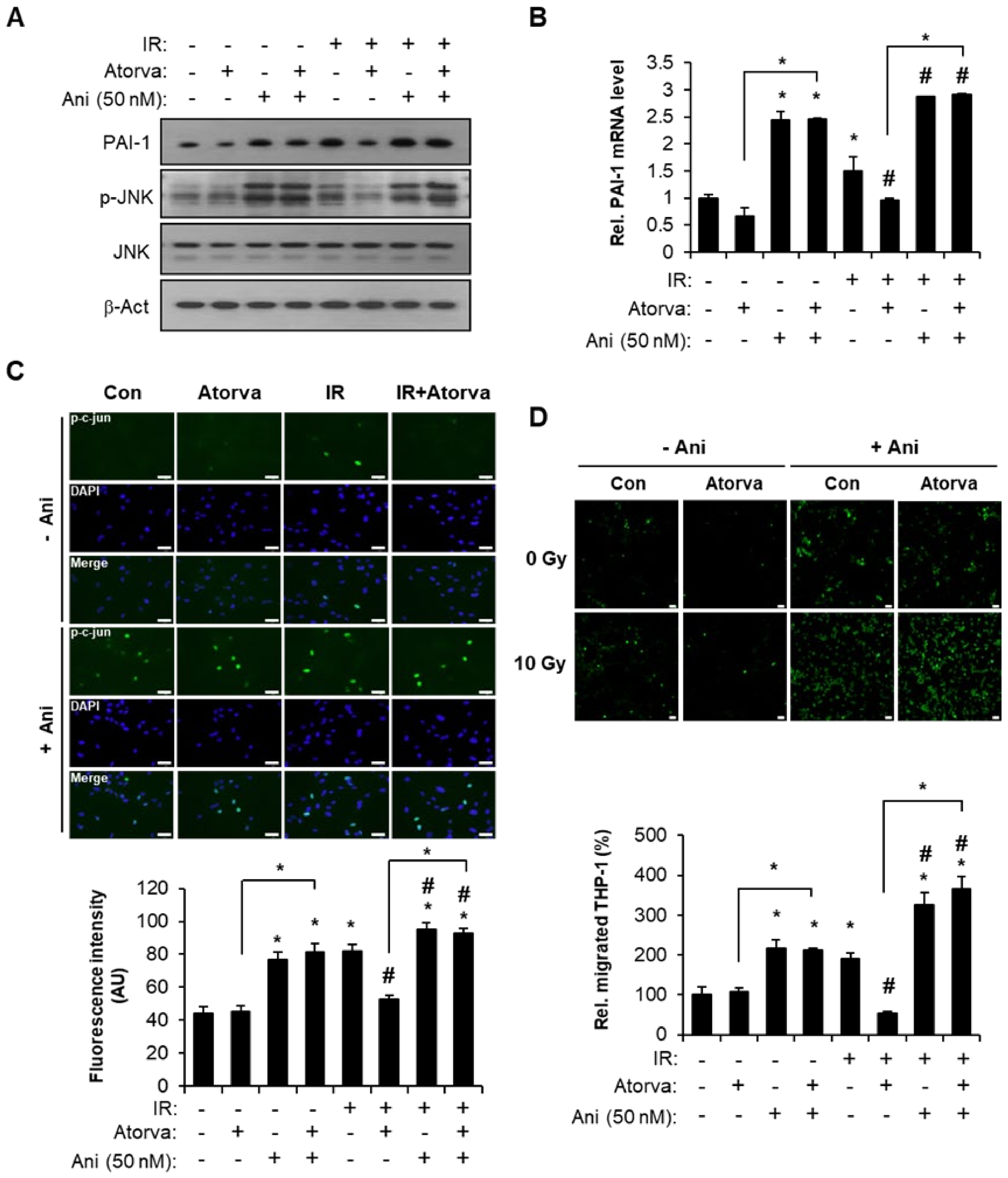
2.3. Atorvastatin Suppress PAI-1 Expression Through JNK–c-Jun Signaling in Irradiated Endothelial Cells
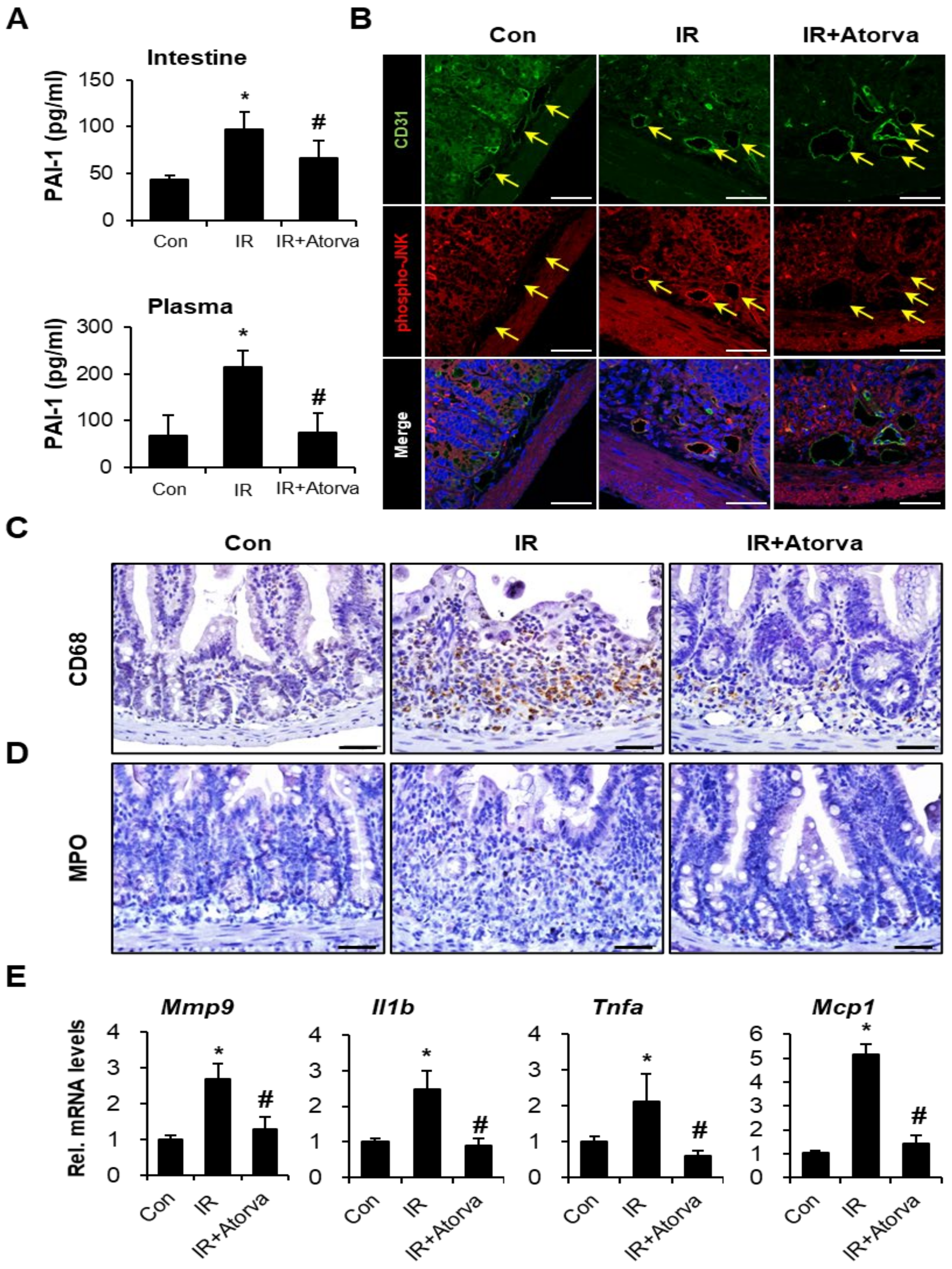
2.4. Atorvastatin Alleviates Radiation-Induced Intestinal Damage In Vivo
2.5. Atorvastatin Inhibits PAI-1 Expression and Attenuates Inflammation Response in Radiation-Induced Intestinal Injury
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Reagents
4.2. Irradiation and Treatment
4.3. Tube Formation Assay
4.4. Invasion Assay
4.5. Senescence-Associated β-Galactosidase Assay (SA-β-gal Assay)
4.6. RNA Extraction and Real-Time RT-PCR
4.7. Western Blot Analysis
4.8. Enzyme-Linked Immunosorbent Assay (ELISA)
4.9. Trans-Endothelial Migration of Monocytes
4.10. Preparation of Nuclear and Cytosolic Fractions
4.11. Immunofluorescence
4.12. In Vivo Experiment and Irradiation
4.13. Histological Analysis
4.14. Bacterial Translocation Assay
4.15. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Singh, V.K.; Seed, T.M.; Olabisi, A.O. Drug discovery strategies for acute radiation syndrome. Expert Opin. Drug Discov. 2019, 14, 701–715. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Zhong, S. From genomes to societies: A holistic view of determinants of human health. Curr. Opin. Biotechnol. 2014, 28, 134–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J. Intestinal stem cell injury and protection during cancer therapy. Transl. Cancer Res. 2013, 2, 384–396. [Google Scholar]
- Slezak, J.; Kura, B.; Ravingerová, T.; Tribulova, N.; Okruhlicova, L.; Barancik, M. Mechanisms of cardiac radiation injury and potential preventive approaches. Can. J. Physiol. Pharmacol. 2015, 93, 737–753. [Google Scholar] [CrossRef] [PubMed]
- Gaugler, M.H.; Neunlist, M.; Bonnaud, S.; Aubert, P.; Benderitter, M.; Paris, F. Intestinal epithelial cell dysfunction is mediated by an endothelial-specific radiation-induced bystander effect. Radiat. Res. 2007, 167, 185–193. [Google Scholar] [CrossRef]
- Belov, O.V.; Batmunkh, M.; Incerti, S.; Lkhagva, O. Radiation damage to neuronal cells: Simulating the energy deposition and water radiolysis in a small neural network. Phys. Med. 2016, 32, 1510–1520. [Google Scholar] [CrossRef] [PubMed]
- Stacey, R.; Green, J.T. Radiation-induced small bowel disease: Latest developments and clinical guidance. Ther. Adv. Chronic Dis. 2014, 5, 15–29. [Google Scholar] [CrossRef] [Green Version]
- Fuccio, L.; Guido, A.; Andreyev, H.J. Management of intestinal complications in patients with pelvic radiation disease. Clin. Gastroenterol. Hepatol. 2012, 10, 1326–1334.e4. [Google Scholar] [CrossRef] [PubMed]
- Hauer-Jensen, M.; Denham, J.W.; Andreyev, H.J. Radiation enteropathy—Pathogenesis, treatment and prevention. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 470–479. [Google Scholar] [CrossRef]
- Cho, C.H.; Kammerer, R.A.; Lee, H.J.; Yasunaga, K.; Kim, K.T.; Choi, H.H.; Kim, W.; Kim, S.H.; Park, S.K.; Lee, G.M.; et al. Designed angiopoietin-1 variant, COMP-Ang1, protects against radiation-induced endothelial cell apoptosis. Proc. Natl. Acad. Sci. USA 2004, 101, 5553–5558. [Google Scholar] [CrossRef] [Green Version]
- Paris, F.; Fuks, Z.; Kang, A.; Capodieci, P.; Juan, G.; Ehleiter, D.; Haimovitz-Friedman, A.; Cordon-Cardo, C.; Kolesnick, R. Endothelial apoptosis as the primary lesion initiating intestinal radiation damage in mice. Science 2001, 293, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Rotolo, J.; Stancevic, B.; Zhang, J.; Hua, G.; Fuller, J.; Yin, X.; Haimovitz-Friedman, A.; Kim, K.; Qian, M.; Cardó-Vila, M.; et al. Anti-ceramide antibody prevents the radiation gastrointestinal syndrome in mice. J. Clin. Invest. 2012, 122, 1786–1790. [Google Scholar] [CrossRef]
- Venkatesulu, B.P.; Mahadevan, L.S.; Aliru, M.L.; Yang, X.; Bodd, M.H.; Singh, P.K.; Yusuf, S.W.; Abe, J.I.; Krishnan, S. Radiation-Induced Endothelial Vascular Injury: A Review of Possible Mechanisms. JACC Basic Transl. Sci. 2018, 3, 563–572. [Google Scholar] [CrossRef]
- Cesari, M.; Pahor, M.; Incalzi, R.A. Plasminogen activator inhibitor-1 (PAI-1): A key factor linking fibrinolysis and age-related subclinical and clinical conditions. Cardiovascular 2010, 28, e72–e91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levi, M.; Poll, T. Coagulation in patients with severe sepsis. Semin. Thromb. Hemost. 2015, 41, 9–15. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.M. Oxidative stress, plasminogen activator inhibitor 1, and lung fibrosis. Antioxid. Redox Signal. 2008, 10, 303–319. [Google Scholar] [CrossRef] [Green Version]
- Marudamuthu, A.S.; Shetty, S.K.; Bhandary, Y.P.; Karandashova, S.; Thompson, M.; Sathish, V.; Florova, G.; Hogan, T.B.; Pabelick, C.M.; Prakash, Y.S.; et al. Plasminogen activator inhibitor-1 suppresses profibrotic responses in fibroblasts from fibrotic lungs. J. Biol. Chem. 2015, 290, 9428–9441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shioya, S.; Masuda, T.; Senoo, T.; Horimasu, Y.; Miyamoto, S.; Nakashima, T.; Iwamoto, H.; Fujitaka, K.; Hamada, H.; Hattori, N. Plasminogen activator inhibitor-1 serves an important role in radiation-induced pulmonary fibrosis. Exp. Med. 2018, 16, 3070–3076. [Google Scholar] [CrossRef]
- Kubala, M.H.; Punj, V.; Placencio-Hickok, V.R.; Fang, H.; Fernandez, G.E.; Sposto, R.; DeClerck, Y.A. Plasminogen Activator Inhibitor-1 Promotes the Recruitment and Polarization of Macrophages in Cancer. Cell Rep. 2018, 25, 2177–2191.e7. [Google Scholar] [CrossRef] [Green Version]
- Ichimura, A.; Matsumoto, S.; Suzuki, S.; Dan, T.; Yamaki, S.; Sato, Y.; Kiyomoto, H.; Ishii, N.; Okada, K.; Matsuo, O.; et al. A small molecule inhibitor to plasminogen activator inhibitor 1 inhibits macrophage migration. Arter. Thromb. Vasc. Biol. 2013, 33, 935–942. [Google Scholar] [CrossRef] [Green Version]
- Milliat, F.; Sabourin, J.C.; Tarlet, G.; Holler, V.; Deutsch, E.; Buard, V.; Tamarat, R.; Atfi, A.; Benderitter, M.; François, A. Essential role of plasminogen activator inhibitor type-1 in radiation enteropathy. Am. J. Pathol. 2008, 172, 691–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scharpfenecker, M.; Kruse, J.J.; Sprong, D.; Russell, N.S.; Ten Dijke, P.; Stewart, F.A. Ionizing radiation shifts the PAI-1/ID-1 balance and activates notch signaling in endothelial cells. Int. J. Radiat. Oncol. Biol. Phys. 2009, 73, 506–513. [Google Scholar] [CrossRef]
- Ebrahimian, T.G.; Squiban, C.; Roque, T.; Lugo-Martinez, H.; Hneino, M.; Buard, V.; Gourmelon, P.; Benderitter, M.; Milliat, F.; Tamarat, R. Plasminogen activator inhibitor-1 controls bone marrow-derived cells therapeutic effect through MMP9 signaling: Role in physiological and pathological wound healing. Stem Cells 2012, 30, 1436–1446. [Google Scholar] [CrossRef]
- Abderrahmani, R.; François, A.; Buard, V.; Tarlet, G.; Blirando, K.; Hneino, M.; Vaurijoux, A.; Benderitter, M.; Sabourin, J.C.; Milliat, F. PAI-1-dependent endothelial cell death determines severity of radiation-induced intestinal injury. PLoS ONE 2012, 7, e35740. [Google Scholar] [CrossRef] [PubMed]
- Rosenson, R.S. Non-lipid-lowering effects of statins on atherosclerosis. Curr. Cardiol. Rep. 1999, 1, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.; Kwak, S.Y.; Park, S.; Kim, K.; Kim, Y.H.; Na, J.; Kim, H.; Jang, W.S.; Lee, S.J.; Kim, M.J.; et al. Pravastatin Alleviates Radiation Proctitis by Regulating Thrombomodulin in Irradiated Endothelial Cells. Int. J. Mol. Sci. 2020, 21, 1897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holler, V.; Buard, V.; Gaugler, M.H.; Guipaud, O.; Baudelin, C.; Sache, A.; Mdel, R.P.; Squiban, C.; Tamarat, R.; Milliat, F.; et al. Pravastatin limits radiation-induced vascular dysfunction in the skin. J. Invest. Derm. 2009, 129, 1280–1291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haydont, V.; Bourgier, C.; Pocard, M.; Lusinchi, A.; Aigueperse, J.; Mathé, D.; Bourhis, J.; Vozenin-Brotons, M.C. Pravastatin Inhibits the Rho/CCN2/extracellular matrix cascade in human fibrosis explants and improves radiation-induced intestinal fibrosis in rats. Clin. Cancer Res. 2007, 13, 5331–5340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haydont, V.; Gilliot, O.; Rivera, S.; Bourgier, C.; François, A.; Aigueperse, J.; Bourhis, J.; Vozenin-Brotons, M.C. Successful mitigation of delayed intestinal radiation injury using pravastatin is not associated with acute injury improvement or tumor protection. Int. J. Radiat. Oncol. Biol. Phys. 2007, 68, 1471–1482. [Google Scholar] [CrossRef]
- Jang, H.; Lee, J.; Park, S.; Myung, H.; Kang, J.; Kim, K.; Kim, H.; Jang, W.S.; Lee, S.J.; Shim, S.; et al. Pravastatin Attenuates Acute Radiation-Induced Enteropathy and Improves Epithelial Cell Function. Front. Pharm. 2018, 9, 1215. [Google Scholar] [CrossRef] [PubMed]
- Laumen, H.; Skurk, T.; Hauner, H. The HMG-CoA reductase inhibitor rosuvastatin inhibits plasminogen activator inhibitor-1 expression and secretion in human adipocytes. Atherosclerosis 2008, 196, 565–573. [Google Scholar] [CrossRef]
- Ni, X.Q.; Zhu, J.H.; Yao, N.H.; Qian, J.; Yang, X.J. Statins suppress glucose-induced plasminogen activator inhibitor-1 expression by regulating RhoA and nuclear factor-κB activities in cardiac microvascular endothelial cells. Exp. Biol. Med. 2013, 238, 37–46. [Google Scholar] [CrossRef]
- Dunoyer-Geindre, S.; Fish, R.J.; Kruithof, E.K. Regulation of the endothelial plasminogen activator system by fluvastatin. Role of Rho family proteins, actin polymerisation and p38 MAP kinase. Thromb. Haemost. 2011, 105, 461–472. [Google Scholar] [PubMed] [Green Version]
- Kruithof, E.K. Regulation of plasminogen activator inhibitor type 1 gene expression by inflammatory mediators and statins. Thromb. Haemost. 2008, 100, 969–975. [Google Scholar] [CrossRef] [PubMed]
- Nordt, T.K.; Peter, K.; Ruef, J.; Kübler, W.; Bode, C. Plasminogen activator inhibitor type-1 (PAI-1) and its role in cardiovascular disease. Thromb. Haemost. 1999, 82 (Suppl. 1), 14–18. [Google Scholar]
- Shao, Q.; Shen, L.H.; Hu, L.H.; Pu, J.; Jing, Q.; He, B. Atorvastatin suppresses inflammatory response induced by oxLDL through inhibition of ERK phosphorylation, IκBα degradation, and COX-2 expression in murine macrophages. J. Cell. Biochem. 2012, 113, 611–618. [Google Scholar] [CrossRef]
- Kim, A.; Seong, K.M.; Choi, Y.Y.; Shim, S.; Park, S.; Lee, S.S. Inhibition of EphA2 by Dasatinib Suppresses Radiation-Induced Intestinal Injury. Int. J. Mol. Sci. 2020, 21, 9096. [Google Scholar] [CrossRef]
- Hwang, H.J.; Lee, Y.R.; Kang, D.; Lee, H.C.; Seo, H.R.; Ryu, J.K.; Kim, Y.N.; Ko, Y.G.; Park, H.J.; Lee, J.S. Endothelial cells under therapy-induced senescence secrete CXCL11, which increases aggressiveness of breast cancer cells. Cancer Lett. 2020, 490, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Tominaga, K. The emerging role of senescent cells in tissue homeostasis and pathophysiology. Pathobiol. Aging Age Relat. Dis. 2015, 5, 27743. [Google Scholar] [CrossRef]
- Khan, S.Y.; Awad, E.M.; Oszwald, A.; Mayr, M.; Yin, X.; Waltenberger, B.; Stuppner, H.; Lipovac, M.; Uhrin, P.; Breuss, J.M. Premature senescence of endothelial cells upon chronic exposure to TNFα can be prevented by N-acetyl cysteine and plumericin. Sci. Rep. 2017, 7, 39501. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Montero, P.; Londoño-Vallejo, A.; Vernot, J.P. Senescence-associated IL-6 and IL-8 cytokines induce a self- and cross-reinforced senescence/inflammatory milieu strengthening tumorigenic capabilities in the MCF-7 breast cancer cell line. Cell Commun. Signal. 2017, 15, 17. [Google Scholar] [CrossRef] [Green Version]
- Abderrahmani, R.; François, A.; Buard, V.; Benderitter, M.; Sabourin, J.C.; Crandall, D.L.; Milliat, F. Effects of pharmacological inhibition and genetic deficiency of plasminogen activator inhibitor-1 in radiation-induced intestinal injury. Int. J. Radiat. Oncol. Biol. Phys. 2009, 74, 942–948. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, M.; Shi, T.; Shen, L.; Zhu, J.; Sun, M.; Deng, Y.; Liang, L.; Li, G.; Wu, Y.; et al. Genetic polymorphisms of PAI-1 and PAR-1 are associated with acute normal tissue toxicity in Chinese rectal cancer patients treated with pelvic radiotherapy. Onco Targets Ther. 2015, 8, 2291–2301. [Google Scholar] [CrossRef] [Green Version]
- Osterholzer, J.J.; Christensen, P.J.; Lama, V.; Horowitz, J.C.; Hattori, N.; Subbotina, N.; Cunningham, A.; Lin, Y.; Murdock, B.J.; Morey, R.E.; et al. PAI-1 promotes the accumulation of exudate macrophages and worsens pulmonary fibrosis following type II alveolar epithelial cell injury. J. Pathol. 2012, 228, 170–180. [Google Scholar] [CrossRef]
- Arndt, P.G.; Young, S.K.; Worthen, G.S. Regulation of lipopolysaccharide-induced lung inflammation by plasminogen activator Inhibitor-1 through a JNK-mediated pathway. J. Immunol. 2005, 175, 4049–4059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.C.; Lin, Y.L.; Hsu, C.C.; Chao, Y.J.; Hou, Y.C.; Chiu, T.J.; Huang, P.H.; Tang, M.J.; Chen, L.T.; Shan, Y.S. Pancreatic stellate cells activated by mutant KRAS-mediated PAI-1 upregulation foster pancreatic cancer progression via IL-8. Theranostics 2019, 9, 7168–7183. [Google Scholar] [CrossRef] [PubMed]
- Kwak, S.H.; Wang, X.Q.; He, Q.; Fang, W.F.; Mitra, S.; Bdeir, K.; Ploplis, V.A.; Xu, Z.; Idell, S.; Cines, D.; et al. Plasminogen activator inhibitor-1 potentiates LPS-induced neutrophil activation through a JNK-mediated pathway. Thromb. Haemost. 2006, 95, 829–835. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, K.; Yamamoto, S.; Tohyama, Y.; Kohsaka, S. Close association of p38 and JNK with plasminogen-dependent upregulation of PAI-1 in rat astrocytes in vitro. Neurosci. Lett. 2010, 471, 66–69. [Google Scholar] [CrossRef] [PubMed]
- Vulin, A.I.; Stanley, F.M. Oxidative stress activates the plasminogen activator inhibitor type 1 (PAI-1) promoter through an AP-1 response element and cooperates with insulin for additive effects on PAI-1 transcription. J. Biol. Chem. 2004, 279, 25172–25178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pontrelli, P.; Ranieri, E.; Ursi, M.; Ghosh-Choudhury, G.; Gesualdo, L.; Schena, F.P.; Grandaliano, G. jun-N-terminal kinase regulates thrombin-induced PAI-1 gene expression in proximal tubular epithelial cells. Kidney Int. 2004, 65, 2249–2261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- François, A.; Milliat, F.; Guipaud, O.; Benderitter, M. Inflammation and immunity in radiation damage to the gut mucosa. BioMed Res. Int. 2013, 2013, 123241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, J.K. Clinical implications for statin pleiotropy. Curr. Opin. Lipidol. 2005, 16, 624–629. [Google Scholar] [CrossRef] [PubMed]
- Sahebkar, A.; Catena, C.; Ray, K.K.; Vallejo-Vaz, A.J.; Reiner, Ž.; Sechi, L.A.; Colussi, G. Impact of statin therapy on plasma levels of plasminogen activator inhibitor-1. A systematic review and meta-analysis of randomised controlled trials. Thromb. Haemost. 2016, 116, 162–171. [Google Scholar]
- Guipaud, O.; Jaillet, C.; Clément-Colmou, K.; François, A.; Supiot, S.; Milliat, F. The importance of the vascular endothelial barrier in the immune-inflammatory response induced by radiotherapy. Br. J. Radiol. 2018, 91, 20170762. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zheng, H.; Ou, X.; Albertson, C.M.; Fink, L.M.; Herbert, J.M.; Hauer-Jensen, M. Hirudin ameliorates intestinal radiation toxicity in the rat: Support for thrombin inhibition as strategy to minimize side-effects after radiation therapy and as countermeasure against radiation exposure. J. Thromb. Haemost. 2004, 2, 2027–2035. [Google Scholar] [CrossRef]
- Maj, J.G.; Paris, F.; Haimovitz-Friedman, A.; Venkatraman, E.; Kolesnick, R.; Fuks, Z. Microvascular function regulates intestinal crypt response to radiation. Cancer Res. 2003, 63, 4338–4341. [Google Scholar] [PubMed]
- Freeberg, M.A.T.; Easa, A.; Lillis, J.A.; Benoit, D.S.W.; van Wijnen, A.J.; Awad, H.A. Transcriptomic Analysis of Cellular Pathways in Healing Flexor Tendons of Plasminogen Activator Inhibitor 1 (PAI-1/Serpine1) Null Mice. J. Orthop. Res. 2020, 38, 43–58. [Google Scholar] [CrossRef]
- Guo, B.; Inoki, K.; Isono, M.; Mori, H.; Kanasaki, K.; Sugimoto, T.; Akiba, S.; Sato, T.; Yang, B.; Kikkawa, R.; et al. MAPK/AP-1-dependent regulation of PAI-1 gene expression by TGF-beta in rat mesangial cells. Kidney Int. 2005, 68, 972–984. [Google Scholar] [CrossRef] [Green Version]
- Rannou, E.; François, A.; Toullec, A.; Guipaud, O.; Buard, V.; Tarlet, G.; Mintet, E.; Jaillet, C.; Iruela-Arispe, M.L.; Benderitter, M.; et al. In vivo evidence for an endothelium-dependent mechanism in radiation-induced normal tissue injury. Sci. Rep. 2015, 5, 15738. [Google Scholar] [CrossRef] [Green Version]
- Eddy, A.A.; Fogo, A.B. Plasminogen activator inhibitor-1 in chronic kidney disease: Evidence and mechanisms of action. J. Am. Soc. Nephrol. 2006, 17, 2999–3012. [Google Scholar] [CrossRef] [Green Version]
- Oda, T.; Jung, Y.O.; Kim, H.S.; Cai, X.; López-Guisa, J.M.; Ikeda, Y.; Eddy, A.A. PAI-1 deficiency attenuates the fibrogenic response to ureteral obstruction. Kidney Int. 2001, 60, 587–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwak, S.Y.; Park, S.; Kim, H.; Lee, S.-J.; Jang, W.-S.; Kim, M.-J.; Lee, S.; Jang, W.I.; Kim, A.R.; Kim, E.H.; et al. Atorvastatin Inhibits Endothelial PAI-1-Mediated Monocyte Migration and Alleviates Radiation-Induced Enteropathy. Int. J. Mol. Sci. 2021, 22, 1828. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041828
Kwak SY, Park S, Kim H, Lee S-J, Jang W-S, Kim M-J, Lee S, Jang WI, Kim AR, Kim EH, et al. Atorvastatin Inhibits Endothelial PAI-1-Mediated Monocyte Migration and Alleviates Radiation-Induced Enteropathy. International Journal of Molecular Sciences. 2021; 22(4):1828. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041828
Chicago/Turabian StyleKwak, Seo Young, Sunhoo Park, Hyewon Kim, Sun-Joo Lee, Won-Suk Jang, Min-Jung Kim, SeungBum Lee, Won Il Jang, Ah Ra Kim, Eun Hye Kim, and et al. 2021. "Atorvastatin Inhibits Endothelial PAI-1-Mediated Monocyte Migration and Alleviates Radiation-Induced Enteropathy" International Journal of Molecular Sciences 22, no. 4: 1828. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041828