
Platelet Lysate Nebulization Protocol for the Treatment of COVID-19 and Its Sequels: Proof of Concept and Scientific Rationale

 , ,
, ,
Abstract
:1. Introduction
2. Results
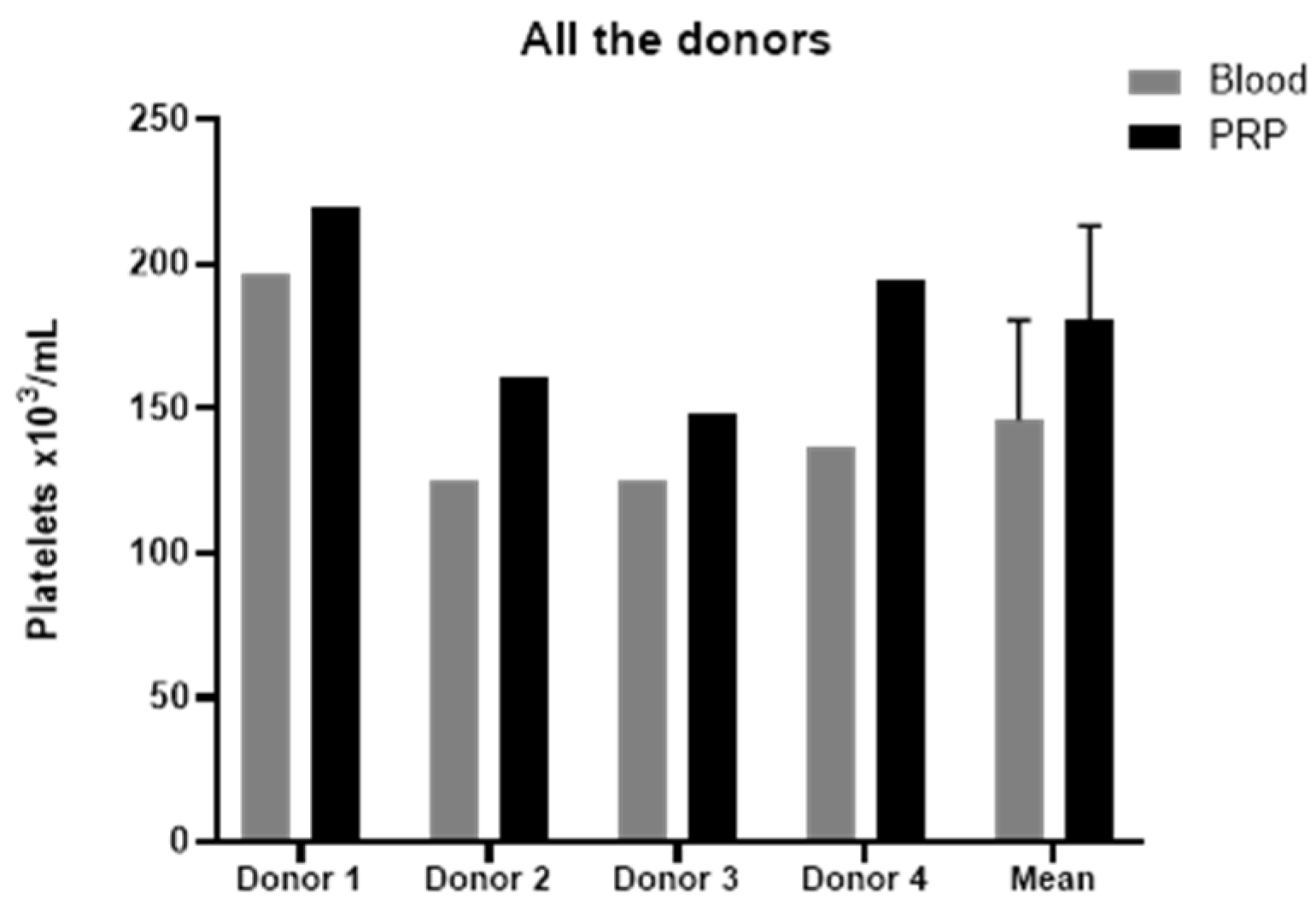
2.1. Platelet-Rich Plasma Characterization
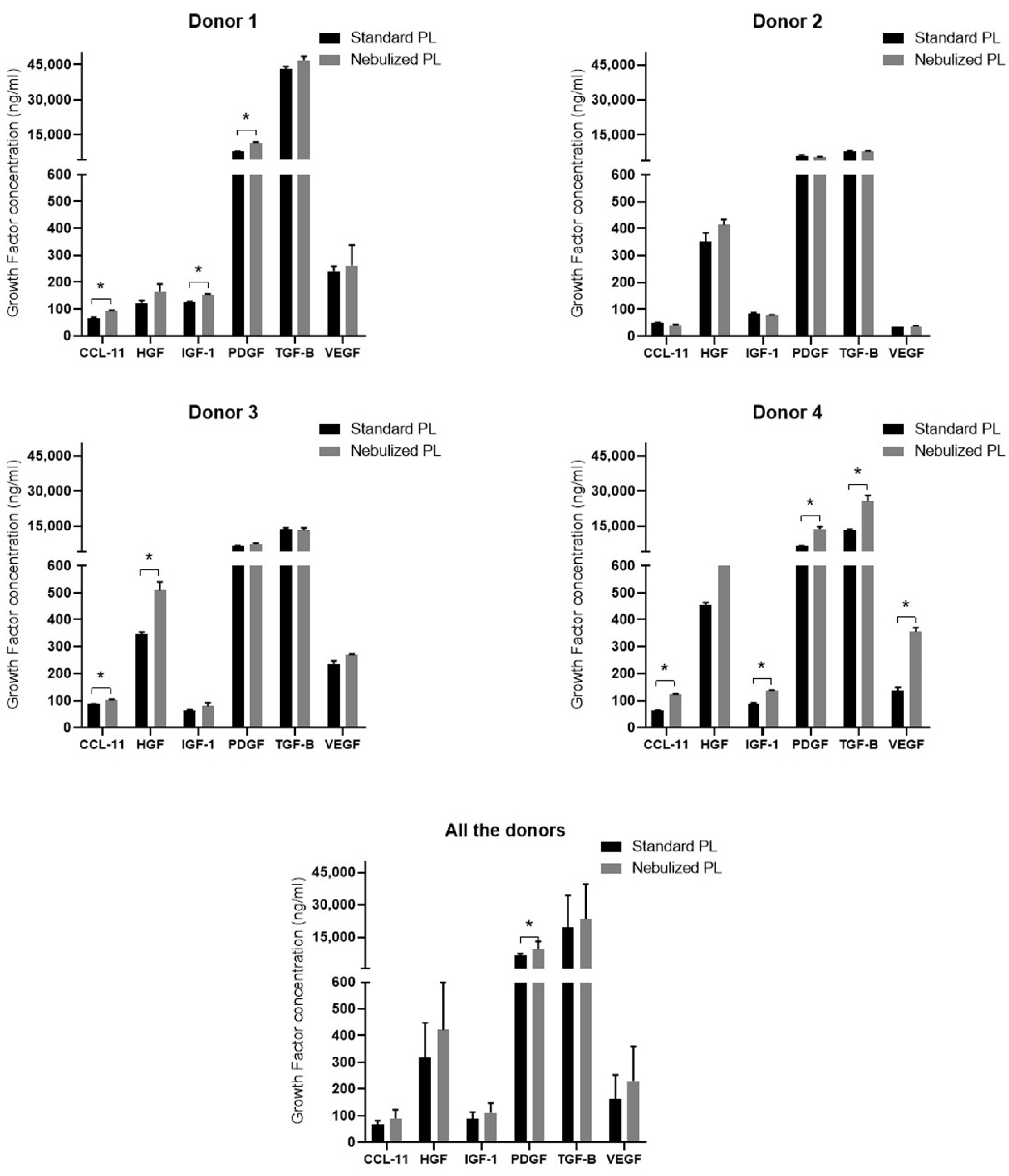
2.2. Growth Factor Profile in Standard and Nebulized Platelet-Rich Plasma
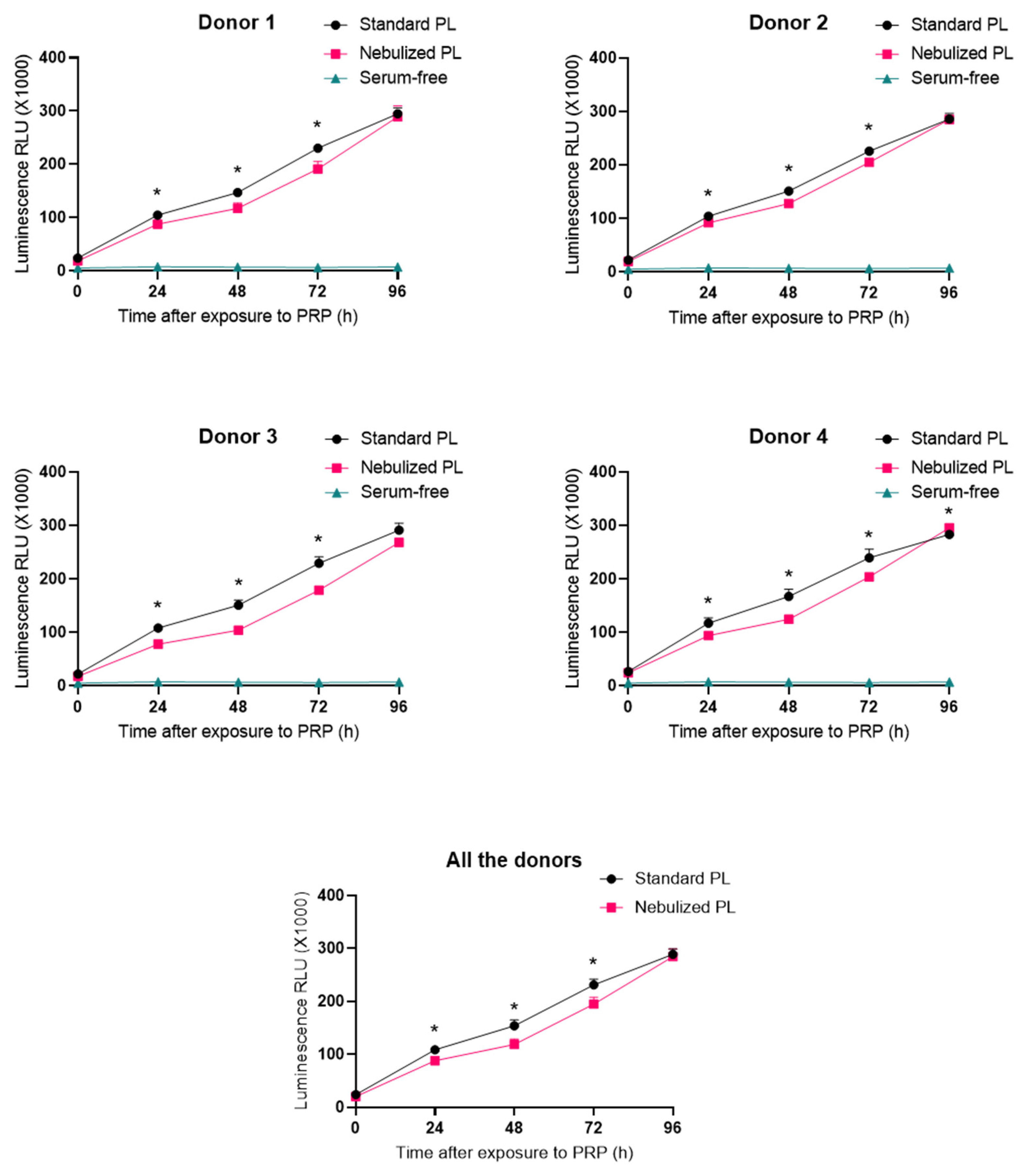
2.3. Impact of Platelet-Rich Plasma Nebulization on Its Bioactivity
3. Discussion
3.1. Role of Platelets against Viral Infections and SARS-CoV-2
3.2. The Importance of Platelets in Lung Biology
3.3. Therapeutic Potential of Platelet-Rich Plasma against COVID-19
4. Materials and Methods
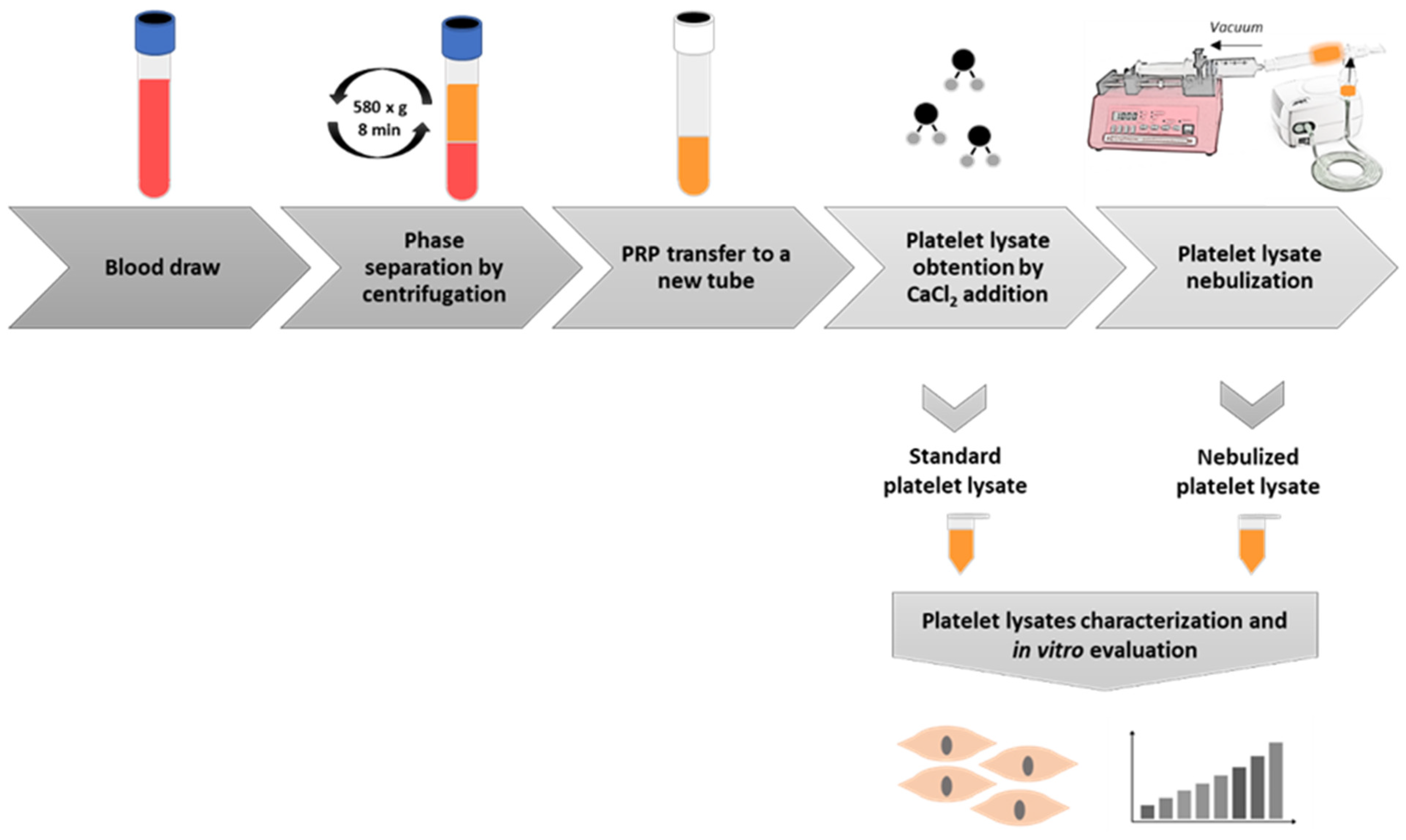
4.1. Platelet-Rich Plasma Preparation
4.2. Platelet-Rich Plasma Lysate Nebulization
4.3. Enzyme-Linked Immunosorbent Assay (ELISA)
4.4. Cell Cultures and Culture Media
4.5. Cell Viability Assay
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| SARS-CoV-2 | Severe Acute Respiratory Syndrome Coronavirus 2 |
| COVID-19 | Coronavirus Disease 2019 |
| ARDS | Acute Respiratory Distress Syndrome |
| PRP | Platelet-Rich Plasma |
| PL | Platelet Lysate |
| CCL-11 | C-C Motif Chemokine Ligand 11 (Eotaxin) |
| CTGF | Connective Tissue Growth Factor |
| HGF | Hepatocyte Growth Factor |
| IGF-1 | Insulin-Like Growth Factor 1 |
| PDGF | Platelet-Derived Growth Factor |
| TGF-β | Transforming Growth Factor Beta |
| VEGF | Vascular Endothelial Growth Factor |
| GPiB | Glycoprotein Ib |
| GPVI | Glycoprotein VI |
| PRR | Pattern Recognition Receptors |
| PAMP | Pathogen-Associated Molecular Pattern |
| DAMP | Damage-Associated Molecular Pattern |
| TLR | Tool-Like Receptor |
| CLR | C-type Lectin Receptor |
| DC-SIGN | Dendritic Cell-Specific ICAM3-Grabbing Non-Integrin |
| ACE-2 | Angiotensin Converting Enzyme 2 |
| NET | Neutrophil Extracellular Trap |
| TMPRSS2 | Transmembrane protease, serine 2 |
| ITGA2B | Integrin alpha-IIB |
| CXCL4 | Platelet Factor 4 |
| IL-1B | Interleukin 1-β |
| MSC | Mesenchymal Stem Cells |
| GM-CSF | Granulocyte-Macrophage Colony-Stimulating Factor |
| ELISA | Enzyme-Linked Immunoabsorbent Assay |
| MCP-1 | Monocyte Chemoattractant Protein-1 |
| RNA | Ribonucleic Acid |
| MPV | Mean Platelet Volume |
References
- Munster, V.J.; Koopmans, M.; van Doremalen, N.; van Riel, D.; de Wit, E. A Novel Coronavirus Emerging in China—Key Questions for Impact Assessment. N. Engl. J. Med. 2020, 382, 692–694. [Google Scholar] [CrossRef]
- Bahl, P.; Doolan, C.; de Silva, C.; Chughtai, A.A.; Bourouiba, L.; MacIntyre, C.R. Airborne or Droplet Precautions for Health Workers Treating Coronavirus Disease 2019? J. Infect. Dis. 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.-Y.; Ma, Y.-T.; Zhang, J.-Y.; Xie, X. COVID-19 and the Cardiovascular System. Nat. Rev. Cardiol. 2020, 17, 259–260. [Google Scholar] [CrossRef] [Green Version]
- Rothan, H.A.; Byrareddy, S.N. The Epidemiology and Pathogenesis of Coronavirus Disease (COVID-19) Outbreak. J. Autoimmun. 2020, 109, 102433. [Google Scholar] [CrossRef]
- Dietz, W.; Santos-Burgoa, C. Obesity and Its Implications for COVID-19 Mortality. Obesity 2020, 28, 1005. [Google Scholar] [CrossRef] [Green Version]
- Guan, W.-J.; Ni, Z.-Y.; Hu, Y.; Liang, W.-H.; Ou, C.-Q.; He, J.-X.; Liu, L.; Shan, H.; Lei, C.-L.; Hui, D.S.C.; et al. Clinical Characteristics of Coronavirus Disease 2019 in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Chen, X.; Cai, Y.; Xia, J.; Zhou, X.; Xu, S.; Huang, H.; Zhang, L.; Zhou, X.; Du, C.; et al. Risk Factors Associated With Acute Respiratory Distress Syndrome and Death in Patients With Coronavirus Disease 2019 Pneumonia in Wuhan, China. JAMA Intern. Med. 2020, 180, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- George, P.M.; Wells, A.U.; Jenkins, R.G. Pulmonary Fibrosis and COVID-19: The Potential Role for Antifibrotic Therapy. Lancet Respir. Med. 2020, 8, 807–815. [Google Scholar] [CrossRef]
- Spagnolo, P.; Balestro, E.; Aliberti, S.; Cocconcelli, E.; Biondini, D.; Casa, G.D.; Sverzellati, N.; Maher, T.M. Pulmonary Fibrosis Secondary to COVID-19: A Call to Arms? Lancet Respir. Med. 2020, 8, 750–752. [Google Scholar] [CrossRef]
- Yang, Y.-H.; Huang, Y.-H.; Chuang, Y.-H.; Peng, C.-M.; Wang, L.-C.; Lin, Y.-T.; Chiang, B.-L. Autoantibodies against Human Epithelial Cells and Endothelial Cells after Severe Acute Respiratory Syndrome (SARS)-Associated Coronavirus Infection. J. Med. Virol. 2005, 77, 1–7. [Google Scholar] [CrossRef]
- Everts, P.; Onishi, K.; Jayaram, P.; Lana, J.F.; Mautner, K. Platelet-Rich Plasma: New Performance Understandings and Therapeutic Considerations in 2020. Int. J. Mol. Sci. 2020, 21, 7794. [Google Scholar] [CrossRef]
- Machlus, K.R.; Italiano, J.E. The Incredible Journey: From Megakaryocyte Development to Platelet Formation. J. Cell Biol. 2013, 201, 785–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefrançais, E.; Ortiz-Muñoz, G.; Caudrillier, A.; Mallavia, B.; Liu, F.; Sayah, D.M.; Thornton, E.E.; Headley, M.B.; David, T.; Coughlin, S.R.; et al. The Lung Is a Site of Platelet Biogenesis and a Reservoir for Haematopoietic Progenitors. Nature 2017, 544, 105–109. [Google Scholar] [CrossRef]
- Padilla, S.; Sánchez, M.; Orive, G.; Anitua, E. Human-Based Biological and Biomimetic Autologous Therapies for Musculoskeletal Tissue Regeneration. Trends Biotechnol. 2017, 35, 192–202. [Google Scholar] [CrossRef]
- Middleton, E.A.; Weyrich, A.S.; Zimmerman, G.A. Platelets in Pulmonary Immune Responses and Inflammatory Lung Diseases. Physiol. Rev. 2016, 96, 1211–1259. [Google Scholar] [CrossRef]
- Middleton, E.A.; Rowley, J.W.; Campbell, R.A.; Grissom, C.K.; Brown, S.M.; Beesley, S.J.; Schwertz, H.; Kosaka, Y.; Manne, B.K.; Krauel, K.; et al. Sepsis Alters the Transcriptional and Translational Landscape of Human and Murine Platelets. Blood 2019, 134, 911–923. [Google Scholar] [CrossRef]
- Kon, E.; Di Matteo, B.; Delgado, D.; Cole, B.J.; Dorotei, A.; Dragoo, J.L.; Filardo, G.; Fortier, L.A.; Giuffrida, A.; Jo, C.H.; et al. Platelet-Rich Plasma for the Treatment of Knee Osteoarthritis: An Expert Opinion and Proposal for a Novel Classification and Coding System. Expert Opin. Biol. Ther. 2020, 20, 1447–1460. [Google Scholar] [CrossRef]
- Wiersinga, W.J.; Rhodes, A.; Cheng, A.C.; Peacock, S.J.; Prescott, H.C. Pathophysiology, Transmission, Diagnosis, and Treatment of Coronavirus Disease 2019 (COVID-19): A Review. JAMA 2020, 324, 782–793. [Google Scholar] [CrossRef]
- Gremmel, T.; Frelinger, A.L.; Michelson, A.D. Platelet Physiology. Semin. Thromb. Hemost. 2016, 42, 191–204. [Google Scholar] [CrossRef] [Green Version]
- Terada, H.; Baldini, M.; Ebbe, S.; Madoff, M.A. Interaction of Influenza Virus with Blood Platelets. Blood 1966, 28, 213–228. [Google Scholar] [CrossRef] [Green Version]
- Seyoum, M.; Enawgaw, B.; Melku, M. Human Blood Platelets and Viruses: Defense Mechanism and Role in the Removal of Viral Pathogens. Thromb. J. 2018, 16, 16. [Google Scholar] [CrossRef]
- Hottz, E.D.; Bozza, F.A.; Bozza, P.T. Platelets in Immune Response to Virus and Immunopathology of Viral Infections. Front. Med. 2018, 5, 121. [Google Scholar] [CrossRef]
- Denis, M.M.; Tolley, N.D.; Bunting, M.; Schwertz, H.; Jiang, H.; Lindemann, S.; Yost, C.C.; Rubner, F.J.; Albertine, K.H.; Swoboda, K.J.; et al. Escaping the Nuclear Confines: Signal-Dependent Pre-MRNA Splicing in Anucleate Platelets. Cell 2005, 122, 379–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manne, B.K.; Denorme, F.; Middleton, E.A.; Portier, I.; Rowley, J.W.; Stubben, C.; Petrey, A.C.; Tolley, N.D.; Guo, L.; Cody, M.; et al. Platelet Gene Expression and Function in Patients with COVID-19. Blood 2020, 136, 1317–1329. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, Y.; Wang, X.; Yang, L.; Li, H.; Wang, Y.; Liu, M.; Zhao, X.; Xie, Y.; Yang, Y.; et al. SARS-CoV-2 Binds Platelet ACE2 to Enhance Thrombosis in COVID-19. J. Hematol. Oncol. 2020, 13, 120. [Google Scholar] [CrossRef]
- Zaid, Y.; Puhm, F.; Allaeys, I.; Naya, A.; Oudghiri, M.; Khalki, L.; Limami, Y.; Zaid, N.; Sadki, K.; Haj, R.B.E.; et al. Platelets Can Contain SARS-CoV-2 RNA and Are Hyperactivated in COVID-19. Circ Res 2020, 127, 1404–1418. [Google Scholar] [CrossRef]
- Yeaman, M.R. Platelets: At the Nexus of Antimicrobial Defence. Nat. Rev. Microbiol. 2014, 12, 426–437. [Google Scholar] [CrossRef] [PubMed]
- Nagasawa, T.; Nakayasu, C.; Rieger, A.M.; Barreda, D.R.; Somamoto, T.; Nakao, M. Phagocytosis by Thrombocytes Is a Conserved Innate Immune Mechanism in Lower Vertebrates. Front. Immunol. 2014, 5, 445. [Google Scholar] [CrossRef] [Green Version]
- Auerbach, D.J.; Lin, Y.; Miao, H.; Cimbro, R.; Difiore, M.J.; Gianolini, M.E.; Furci, L.; Biswas, P.; Fauci, A.S.; Lusso, P. Identification of the Platelet-Derived Chemokine CXCL4/PF-4 as a Broad-Spectrum HIV-1 Inhibitor. Proc. Natl. Acad. Sci. USA 2012, 109, 9569–9574. [Google Scholar] [CrossRef] [Green Version]
- Morrell, C.N.; Aggrey, A.A.; Chapman, L.M.; Modjeski, K.L. Emerging Roles for Platelets as Immune and Inflammatory Cells. Blood 2014, 123, 2759–2767. [Google Scholar] [CrossRef] [Green Version]
- Maouia, A.; Rebetz, J.; Kapur, R.; Semple, J.W. The Immune Nature of Platelets Revisited. Transfus. Med. Rev. 2020, 34, 209–220. [Google Scholar] [CrossRef]
- Jenne, C.N.; Urrutia, R.; Kubes, P. Platelets: Bridging Hemostasis, Inflammation, and Immunity. Int. J. Lab. Hematol. 2013, 35, 254–261. [Google Scholar] [CrossRef]
- Gautam, I.; Storad, Z.; Filipiak, L.; Huss, C.; Meikle, C.K.; Worth, R.G.; Wuescher, L.M. From Classical to Unconventional: The Immune Receptors Facilitating Platelet Responses to Infection and Inflammation. Biology 2020, 9, 343. [Google Scholar] [CrossRef]
- Weyrich, A.S.; Lindemann, S.; Zimmerman, G.A. The Evolving Role of Platelets in Inflammation. J. Thromb. Haemost. 2003, 1, 1897–1905. [Google Scholar] [CrossRef] [Green Version]
- Henn, V.; Slupsky, J.R.; Gräfe, M.; Anagnostopoulos, I.; Förster, R.; Müller-Berghaus, G.; Kroczek, R.A. CD40 Ligand on Activated Platelets Triggers an Inflammatory Reaction of Endothelial Cells. Nature 1998, 391, 591–594. [Google Scholar] [CrossRef]
- Del Conde, I.; Crúz, M.A.; Zhang, H.; López, J.A.; Afshar-Kharghan, V. Platelet Activation Leads to Activation and Propagation of the Complement System. J. Exp. Med. 2005, 201, 871–879. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, M.; von Ungern-Sternberg, S.N.I.; Seizer, P.; Schlegel, F.; Büttcher, M.; Sindhu, N.A.; Müller, S.; Mack, A.; Gawaz, M. Platelet-Derived CXCL12 Regulates Monocyte Function, Survival, Differentiation into Macrophages and Foam Cells through Differential Involvement of CXCR4-CXCR7. Cell Death Dis. 2015, 6, e1989. [Google Scholar] [CrossRef] [Green Version]
- Middleton, E.A.; Rondina, M.T.; Schwertz, H.; Zimmerman, G.A. Amicus or Adversary Revisited: Platelets in Acute Lung Injury and Acute Respiratory Distress Syndrome. Am. J. Respir. Cell Mol. Biol. 2018, 59, 18–35. [Google Scholar] [CrossRef] [PubMed]
- Middleton, E.A.; He, X.-Y.; Denorme, F.; Campbell, R.A.; Ng, D.; Salvatore, S.P.; Mostyka, M.; Baxter-Stoltzfus, A.; Borczuk, A.C.; Loda, M.; et al. Neutrophil Extracellular Traps Contribute to Immunothrombosis in COVID-19 Acute Respiratory Distress Syndrome. Blood 2020, 136, 1169–1179. [Google Scholar] [CrossRef]
- Barnes, B.J.; Adrover, J.M.; Baxter-Stoltzfus, A.; Borczuk, A.; Cools-Lartigue, J.; Crawford, J.M.; Daßler-Plenker, J.; Guerci, P.; Huynh, C.; Knight, J.S.; et al. Targeting Potential Drivers of COVID-19: Neutrophil Extracellular Traps. J. Exp. Med. 2020, 217. [Google Scholar] [CrossRef]
- Zuo, Y.; Yalavarthi, S.; Shi, H.; Gockman, K.; Zuo, M.; Madison, J.A.; Blair, C.; Weber, A.; Barnes, B.J.; Egeblad, M.; et al. Neutrophil Extracellular Traps in COVID-19. JCI Insight 2020, 5. [Google Scholar] [CrossRef] [Green Version]
- Hottz, E.D.; Azevedo-Quintanilha, I.G.; Palhinha, L.; Teixeira, L.; Barreto, E.A.; Pão, C.R.R.; Righy, C.; Franco, S.; Souza, T.M.L.; Kurtz, P.; et al. Platelet Activation and Platelet-Monocyte Aggregate Formation Trigger Tissue Factor Expression in Patients with Severe COVID-19. Blood 2020, 136, 1330–1341. [Google Scholar] [CrossRef]
- Schaphorst, K.L.; Chiang, E.; Jacobs, K.N.; Zaiman, A.; Natarajan, V.; Wigley, F.; Garcia, J.G.N. Role of Sphingosine-1 Phosphate in the Enhancement of Endothelial Barrier Integrity by Platelet-Released Products. Am. J. Physiol. Lung Cell Mol. Physiol. 2003, 285, L258–L267. [Google Scholar] [CrossRef] [Green Version]
- Bozza, F.A.; Shah, A.M.; Weyrich, A.S.; Zimmerman, G.A. Amicus or Adversary: Platelets in Lung Biology, Acute Injury, and Inflammation. Am. J. Respir. Cell Mol. Biol. 2009, 40, 123–134. [Google Scholar] [CrossRef]
- Batah, S.S.; Fabro, A.T. Pulmonary Pathology of ARDS in COVID-19: A Pathological Review for Clinicians. Respir. Med. 2021, 176, 106239. [Google Scholar] [CrossRef]
- Lippi, G.; Plebani, M.; Henry, B.M. Thrombocytopenia Is Associated with Severe Coronavirus Disease 2019 (COVID-19) Infections: A Meta-Analysis. Clin. Chim. Acta 2020, 506, 145–148. [Google Scholar] [CrossRef]
- Yamane, A.; Nakamura, T.; Suzuki, H.; Ito, M.; Ohnishi, Y.; Ikeda, Y.; Miyakawa, Y. Interferon-Alpha 2b-Induced Thrombocytopenia Is Caused by Inhibition of Platelet Production but Not Proliferation and Endomitosis in Human Megakaryocytes. Blood 2008, 112, 542–550. [Google Scholar] [CrossRef]
- Johnston, I.; Hayes, V.; Poncz, M. Threading an Elephant through the Eye of a Needle: Where Are Platelets Made? Cell Res. 2017, 27, 1079–1080. Available online: https://0-www-nature-com.brum.beds.ac.uk/articles/cr201765 (accessed on 3 December 2020). [CrossRef] [Green Version]
- Weyrich, A.S.; Zimmerman, G.A. Platelets in Lung Biology. Annu. Rev. Physiol. 2013, 75, 569–591. [Google Scholar] [CrossRef] [Green Version]
- Burnham, E.L.; Janssen, W.J.; Riches, D.W.H.; Moss, M.; Downey, G.P. The Fibroproliferative Response in Acute Respiratory Distress Syndrome: Mechanisms and Clinical Significance. Eur. Respir. J. 2014, 43, 276–285. [Google Scholar] [CrossRef] [Green Version]
- Khoury, M.; Cuenca, J.; Cruz, F.F.; Figueroa, F.E.; Rocco, P.R.M.; Weiss, D.J. Current Status of Cell-Based Therapies for Respiratory Virus Infections: Applicability to COVID-19. Eur. Respir. J. 2020, 55. [Google Scholar] [CrossRef] [Green Version]
- Meng, F.; Xu, R.; Wang, S.; Xu, Z.; Zhang, C.; Li, Y.; Yang, T.; Shi, L.; Fu, J.; Jiang, T.; et al. Human Umbilical Cord-Derived Mesenchymal Stem Cell Therapy in Patients with COVID-19: A Phase 1 Clinical Trial. Signal. Transduct. Target. Ther. 2020, 5, 172. [Google Scholar] [CrossRef]
- Sengupta, V.; Sengupta, S.; Lazo, A.; Woods, P.; Nolan, A.; Bremer, N. Exosomes Derived from Bone Marrow Mesenchymal Stem Cells as Treatment for Severe COVID-19. Stem Cells Dev. 2020, 29, 747–754. [Google Scholar] [CrossRef]
- Shu, L.; Niu, C.; Li, R.; Huang, T.; Wang, Y.; Huang, M.; Ji, N.; Zheng, Y.; Chen, X.; Shi, L.; et al. Treatment of Severe COVID-19 with Human Umbilical Cord Mesenchymal Stem Cells. Stem Cell Res. Ther. 2020, 11, 361. [Google Scholar] [CrossRef]
- Bendinelli, P.; Matteucci, E.; Dogliotti, G.; Corsi, M.M.; Banfi, G.; Maroni, P.; Desiderio, M.A. Molecular Basis of Anti-Inflammatory Action of Platelet-Rich Plasma on Human Chondrocytes: Mechanisms of NF-ΚB Inhibition via HGF. J. Cell Physiol. 2010, 225, 757–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasina, E.M.; Cauwenberghs, S.; Feijge, M.A.H.; Heemskerk, J.W.M.; Weber, C.; Koenen, R.R. Microparticles from Apoptotic Platelets Promote Resident Macrophage Differentiation. Cell Death Dis. 2011, 2, e211. [Google Scholar] [CrossRef] [Green Version]
- Tohidnezhad, M.; Wruck, C.-J.; Slowik, A.; Kweider, N.; Beckmann, R.; Bayer, A.; Houben, A.; Brandenburg, L.-O.; Varoga, D.; Sönmez, T.-T.; et al. Role of Platelet-Released Growth Factors in Detoxification of Reactive Oxygen Species in Osteoblasts. Bone 2014, 65, 9–17. [Google Scholar] [CrossRef]
- Haselton, F.R.; Alexander, J.S. Platelets and a Platelet-Released Factor Enhance Endothelial Barrier. Am. J. Physiol. 1992, 263, L670–L678. [Google Scholar] [CrossRef]
- Lo, S.K.; Burhop, K.E.; Kaplan, J.E.; Malik, A.B. Role of Platelets in Maintenance of Pulmonary Vascular Permeability to Protein. Am. J. Physiol. 1988, 254, H763–H771. [Google Scholar] [CrossRef]
- Straum, O.K. The Optimal Platelet Concentration in Platelet-Rich Plasma for Proliferation of Human Cells in Vitro-Diversity, Biases, and Possible Basic Experimental Principles for Further Research in the Field: A Review. PeerJ 2020, 8, e10303. [Google Scholar] [CrossRef]
- Giusti, I.; D’Ascenzo, S.; Macchiarelli, G.; Dolo, V. In Vitro Evidence Supporting Applications of Platelet Derivatives in Regenerative Medicine. Blood Transfus. 2020, 18, 117–129. [Google Scholar] [CrossRef]
- Guillamat-Prats, R.; Camprubí-Rimblas, M.; Puig, F.; Herrero, R.; Tantinyà, N.; Serrano-Mollar, A.; Artigas, A. Alveolar Type II Cells or Mesenchymal Stem Cells: Comparison of Two Different Cell Therapies for the Treatment of Acute Lung Injury in Rats. Cells 2020, 9, 1816. [Google Scholar] [CrossRef]
- Rubio-Azpeitia, E.; Andia, I. Partnership between Platelet-Rich Plasma and Mesenchymal Stem Cells: In Vitro Experience. Muscles Ligaments Tendons J. 2014, 4, 52–62. [Google Scholar] [CrossRef]
- Andia, I.; Sánchez, M.; Maffulli, N. Basic Science: Molecular and Biological Aspects of Platelet-Rich Plasma Therapies. Oper. Tech. Orthop. 2012, 22, 3–9. [Google Scholar] [CrossRef]
- Chellini, F.; Tani, A.; Vallone, L.; Nosi, D.; Pavan, P.; Bambi, F.; Zecchi Orlandini, S.; Sassoli, C. Platelet-Rich Plasma Prevents In Vitro Transforming Growth Factor-Β1-Induced Fibroblast to Myofibroblast Transition: Involvement of Vascular Endothelial Growth Factor (VEGF)-A/VEGF Receptor-1-Mediated Signaling. Cells 2018, 7, 142. [Google Scholar] [CrossRef] [Green Version]
- Squecco, R.; Chellini, F.; Idrizaj, E.; Tani, A.; Garella, R.; Pancani, S.; Pavan, P.; Bambi, F.; Zecchi-Orlandini, S.; Sassoli, C. Platelet-Rich Plasma Modulates Gap Junction Functionality and Connexin 43 and 26 Expression During TGF-Β1-Induced Fibroblast to Myofibroblast Transition: Clues for Counteracting Fibrosis. Cells 2020, 9, 1199. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, M.; Anitua, E.; Azofra, J.; Andía, I.; Padilla, S.; Mujika, I. Comparison of Surgically Repaired Achilles Tendon Tears Using Platelet-Rich Fibrin Matrices. Am. J. Sports Med. 2007, 35, 245–251. [Google Scholar] [CrossRef]
- Everts, P.A.M.; Devilee, R.J.J.; Oosterbos, C.J.M.; Mahoney, C.B.; Schattenkerk, M.E.; Knape, J.T.A.; van Zundert, A. Autologous Platelet Gel and Fibrin Sealant Enhance the Efficacy of Total Knee Arthroplasty: Improved Range of Motion, Decreased Length of Stay and a Reduced Incidence of Arthrofibrosis. Knee Surg. Sports Traumatol. Arthrosc. 2007, 15, 888–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, M.; Sugita, K.; Inukai, T.; Goi, K.; Kagami, K.; Kawasaki, K.; Nakazawa, S. Hepatocyte Growth Factor Protects Small Airway Epithelial Cells from Apoptosis Induced by Tumor Necrosis Factor-Alpha or Oxidative Stress. Pediatr. Res. 2004, 56, 336–344. [Google Scholar] [CrossRef] [Green Version]
- Mammoto, T.; Chen, Z.; Jiang, A.; Jiang, E.; Ingber, D.E.; Mammoto, A. Acceleration of Lung Regeneration by Platelet-Rich Plasma Extract through the Low-Density Lipoprotein Receptor-Related Protein 5-Tie2 Pathway. Am. J. Respir. Cell Mol. Biol. 2016, 54, 103–113. [Google Scholar] [CrossRef] [Green Version]
- Mammoto, T.; Jiang, A.; Jiang, E.; Mammoto, A. Platelet-Rich Plasma Extract Prevents Pulmonary Edema through Angiopoietin-Tie2 Signaling. Am. J. Respir. Cell Mol. Biol. 2015, 52, 56–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dzyekanski, B.; Rocha, D.C.C.; de Macedo Lopes, A.C.A.; Kunz, J.R.; Rebelatto, C.L.K.; Pimpão, C.T.; Michelotto, P.V., Jr. Intrabronchial Instillation of Platelet-Rich Plasma in Equines with Inflammatory Airway Disease—Preliminary Report. Estudos de Biologia 2012, 34. [Google Scholar] [CrossRef]
- Salama, S.M.; Kamel, I.H.; Ghanim, M.; Elsherif, A.F. The Efficacy of Autologous Nebulized Platelet Rich Plasma (PRP) As an Early Adjuvant Therapeutic and Prognostic Treatment Modality in the Management of Inhalation Lung Injury. Egypt. J. Plast. Reconstr. Surg. 2019, 43, 203–208. [Google Scholar] [CrossRef] [Green Version]
- Felthaus, O.; Prantl, L.; Skaff-Schwarze, M.; Klein, S.; Anker, A.; Ranieri, M.; Kuehlmann, B. Effects of Different Concentrations of Platelet-Rich Plasma and Platelet-Poor Plasma on Vitality and Differentiation of Autologous Adipose Tissue-Derived Stem Cells. Clin. Hemorheol. Microcirc. 2017, 66, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Giusti, I.; D’Ascenzo, S.; Mancò, A.; Di Stefano, G.; Di Francesco, M.; Rughetti, A.; Dal Mas, A.; Properzi, G.; Calvisi, V.; Dolo, V. Platelet Concentration in Platelet-Rich Plasma Affects Tenocyte Behavior in Vitro. Biomed. Res. Int. 2014, 2014, 630870. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1. Platelet-Rich Plasma Preparation | |
| Initial blood volume | 10 mL |
| Anticoagulant | Sodium citrate 3.8% (wt/V) |
| System | Close |
| Centrifugation | Yes |
| number | 1 |
| speed | 580× g—8 min |
| Final PRP volume | 4 mL per subject |
| 2. Platelet-Rich Plasma Characteristics | |
| PRP Type | 11-00-11 |
| Mean Platelet Volume (MPV) | 9.6 ± 0.6 fL |
| Red Blood Cells | <0.01 × 106/µL |
| White Blood Cells | <0.05 × 106/µL |
| Neutrophils | --- |
| Lymphocytes | --- |
| Monocytes | --- |
| Eosinophils | --- |
| Basophils | --- |
| Activation | CaCl2 (10% wt/vol) |
| 3. Application Characteristics | |
| Dose | 2% |
| Direct/Indirect | Direct |
| Cell line | Lung fibroblast |
| 4. Other Remarkable Platelet-Rich Plasma and Study Features | |
| The product added to the cell cultures was the platelet lysate obtained after activation of PRP with calcium chloride (10%) | |
| Effectors | Target | Action | References |
|---|---|---|---|
| HGF, IGF-1, PDGF, TGF-β | NF-kB cellular pathway | Anti-inflammatory effect | [14,59,65] |
| Platelet microparticles | Macrophages | Favoring the reparative phenotype M2 | [55] |
| VEGF | Nrf2 cellular pathway | Preventing oxidative damage | [56] |
| Sphingosine-1 | endothelial cells | Preserving alveolocapillary barrier | [43] |
| PDGF, TGF-β, EGF, FGF-2, CTGF and other growth factors | Lung cell populations: fibroblast, endothelial and epithelial cells, alveolar Type II Cells | Cell migration and proliferation, and tissue repair | [57,62] |
| VEGF-A/TGF-β1 | Fibroblast | Antifibrotic effect | [58,66] |
| Type of Study | Target Cell/Tissue | Route of Administration | Effect | Reference |
|---|---|---|---|---|
| In vitro study | Airway epithelial cells | HGF at different concentrations in cell culture | Anti-apoptotic effect against pro-inflammatory stimuli | [69] |
| In vivo study | Lung tissue in mice subjected to pneumonectomy | Platelet lysate administered intraperitoneally | Stimulation of adult mouse lung vascular and alveolar regeneration | [70] |
| In vivo study | Lung tissue in mice treated with LPS to generate edema and ARDS | Platelet lysate administered intraperitoneally | Prevention of LPS-induced vascular leakage in lungs | [71] |
| In vivo study | Horses with inflammatory airway disease | Intrabronchial instillation | Significative improvement of horses, with a reduction of the mucus grade and neutrophil amount | [72] |
| Clinical study | Patients with inhalation lung injury | Aerosolized PRP | Lower extubation time, hospital stay and mortality rate | [73] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beitia, M.; Delgado, D.; Sánchez, P.; Vallejo de la Cueva, A.; Cugat, J.R.; Sánchez, M. Platelet Lysate Nebulization Protocol for the Treatment of COVID-19 and Its Sequels: Proof of Concept and Scientific Rationale. Int. J. Mol. Sci. 2021, 22, 1856. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041856
Beitia M, Delgado D, Sánchez P, Vallejo de la Cueva A, Cugat JR, Sánchez M. Platelet Lysate Nebulization Protocol for the Treatment of COVID-19 and Its Sequels: Proof of Concept and Scientific Rationale. International Journal of Molecular Sciences. 2021; 22(4):1856. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041856
Chicago/Turabian StyleBeitia, Maider, Diego Delgado, Pello Sánchez, Ana Vallejo de la Cueva, José Ramón Cugat, and Mikel Sánchez. 2021. "Platelet Lysate Nebulization Protocol for the Treatment of COVID-19 and Its Sequels: Proof of Concept and Scientific Rationale" International Journal of Molecular Sciences 22, no. 4: 1856. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041856