
Visualization of Three Sclerotiniaceae Species Pathogenic on Onion Reveals Distinct Biology and Infection Strategies

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
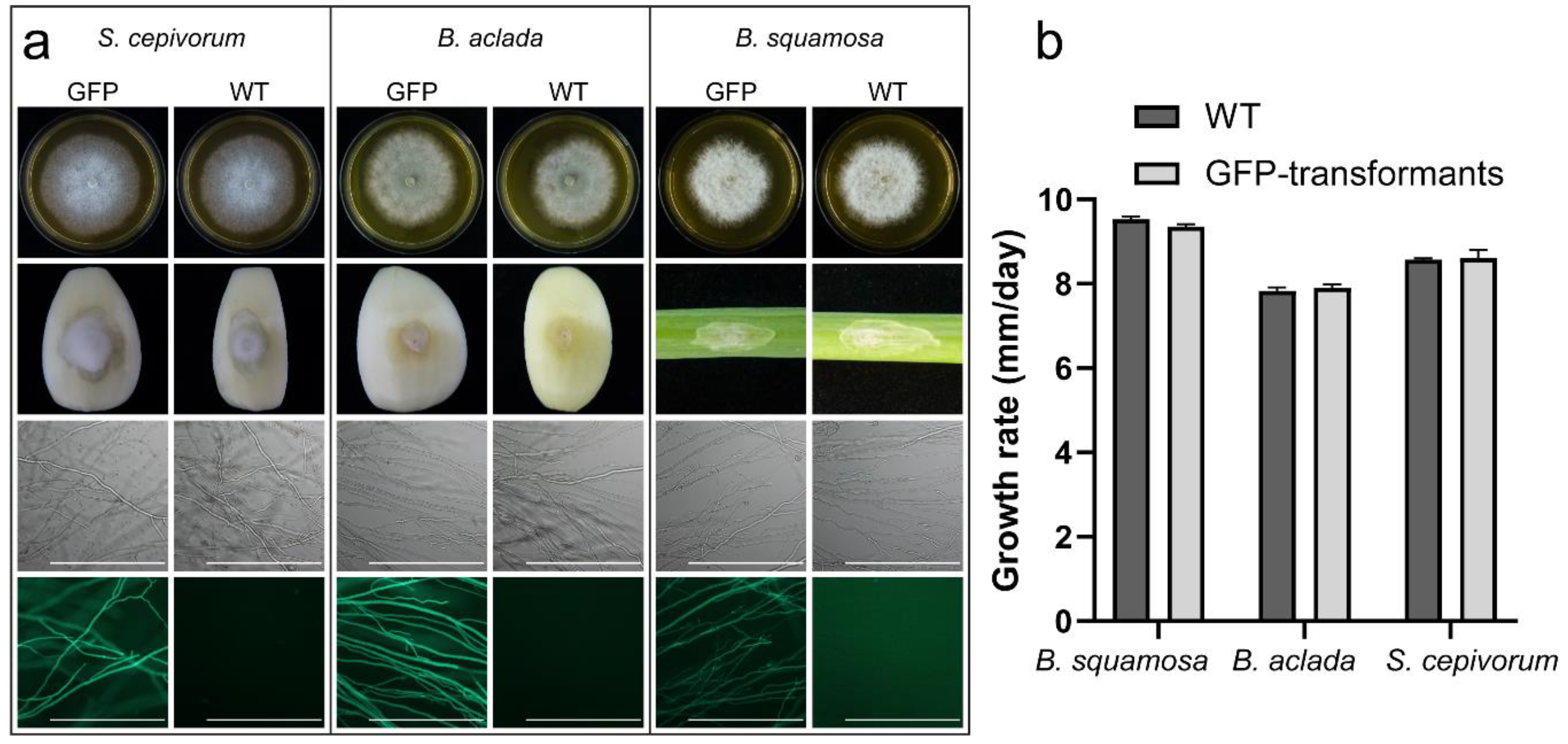
2.1. Transformation of B. squamosa, B. aclada and S. cepivorum with gfp Yields Fluorescently Labeled Fungi
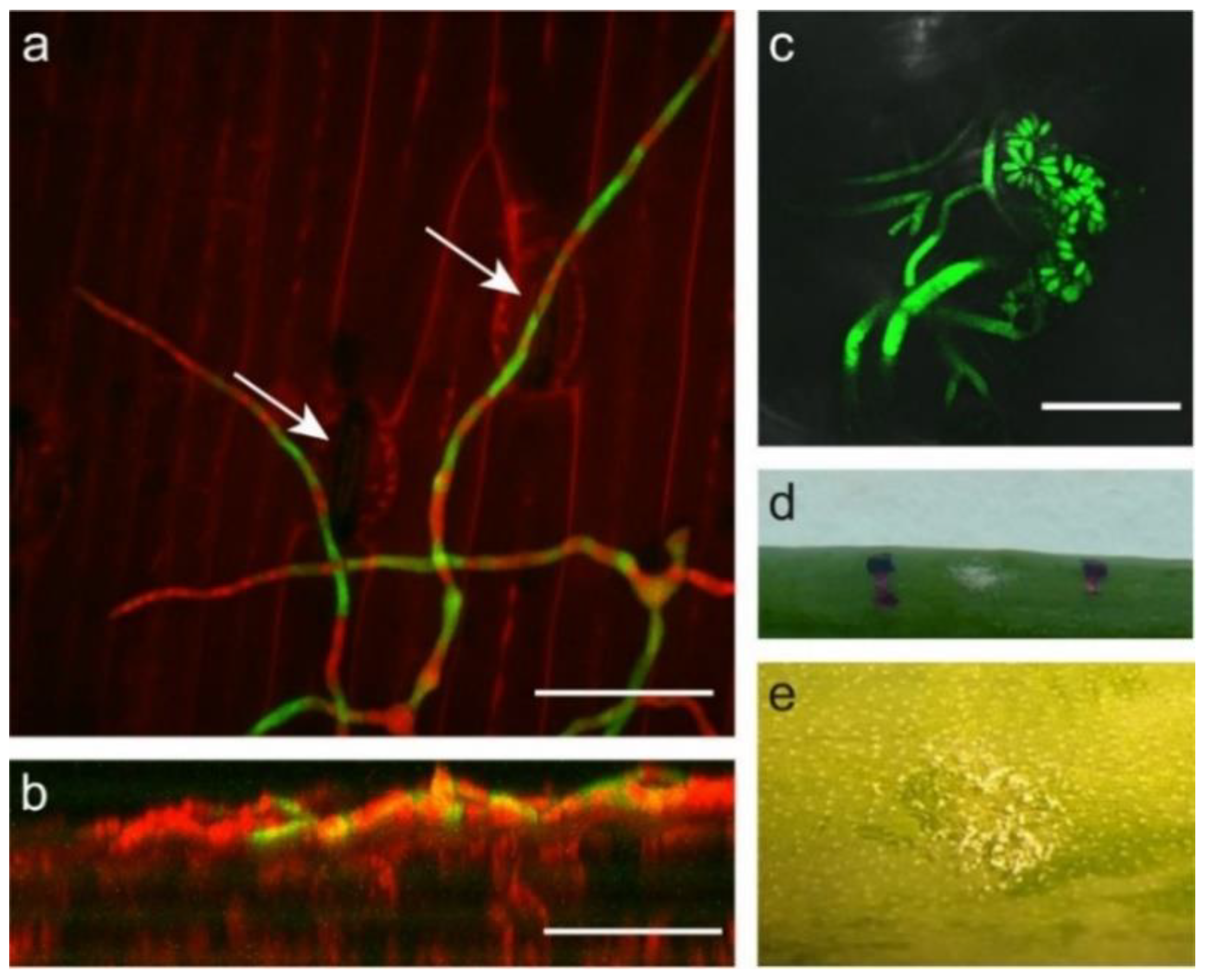
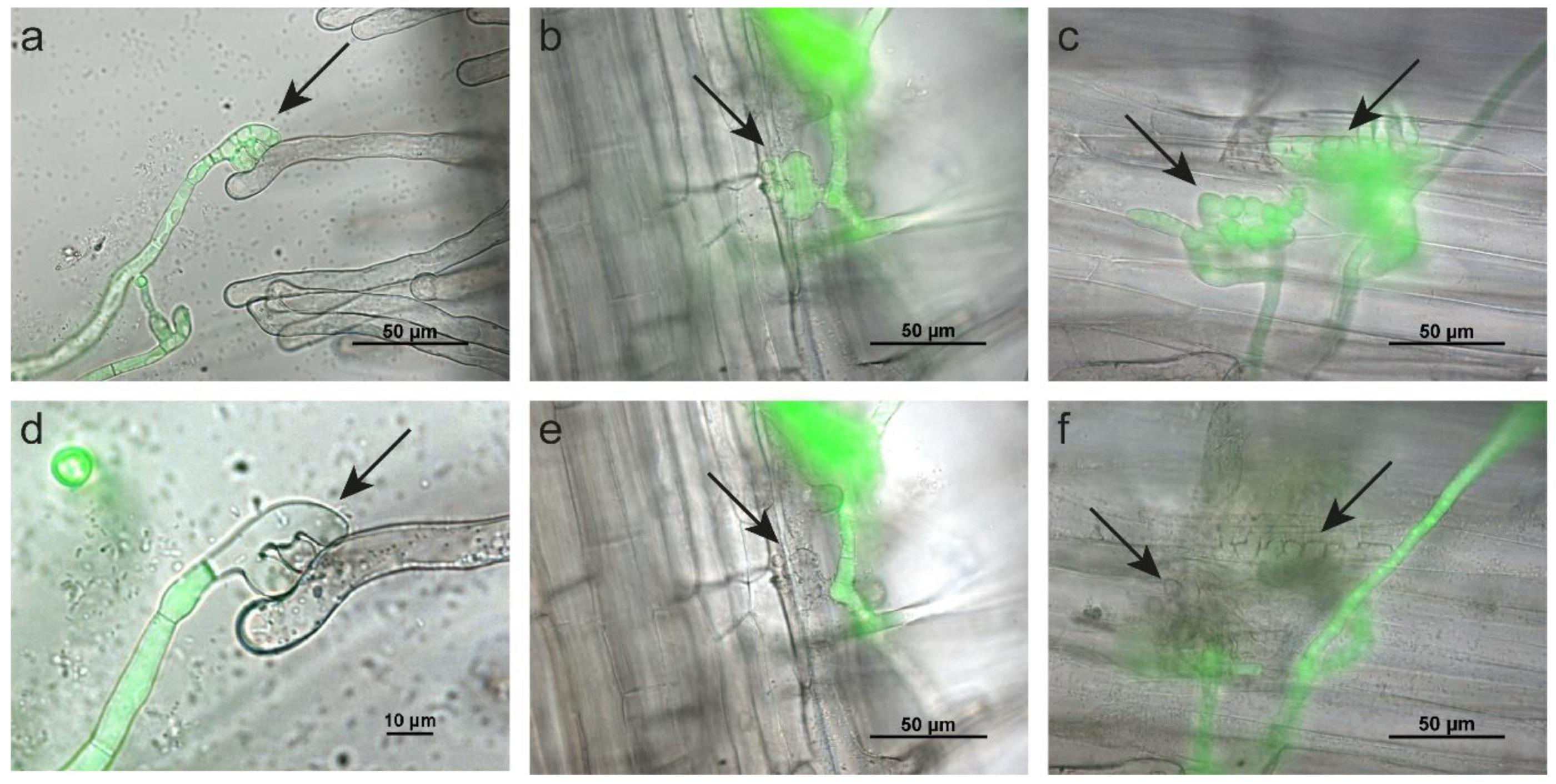
2.2. B. squamosa Enters the Onion Leaf by Growth through Stomata or into Anticlinal Walls
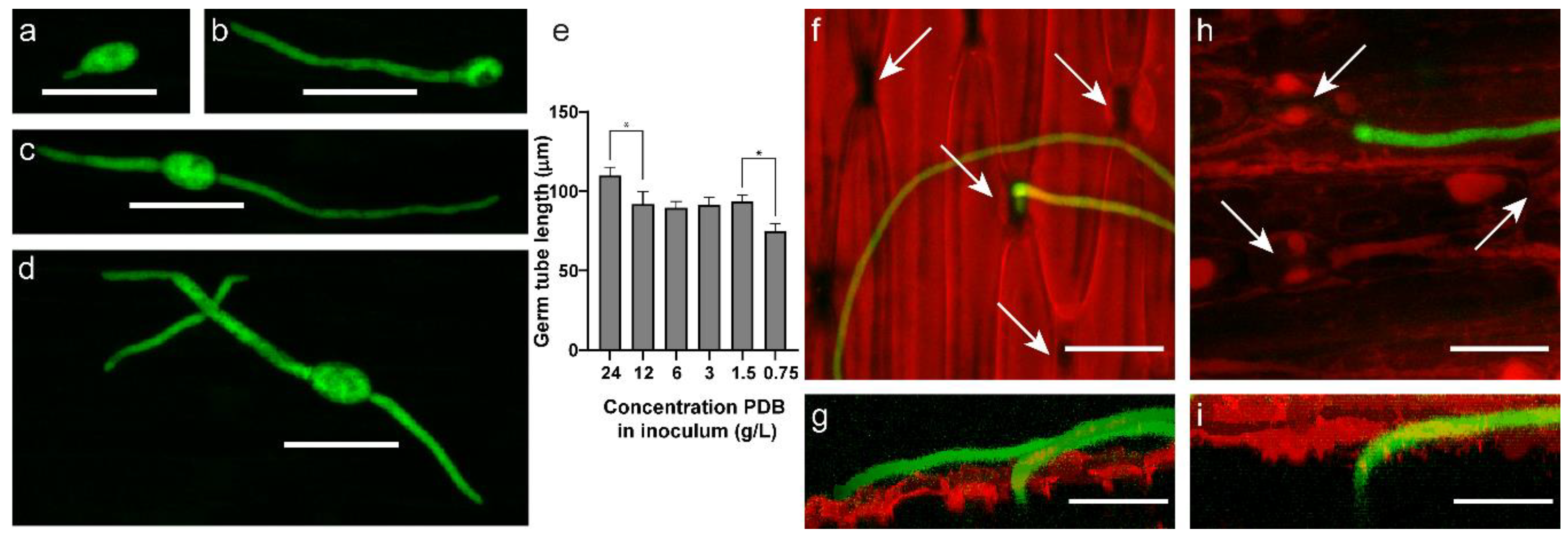
2.3. Botrytis Aclada Reproduces Asexually without Penetrating the Onion Leaf Surface
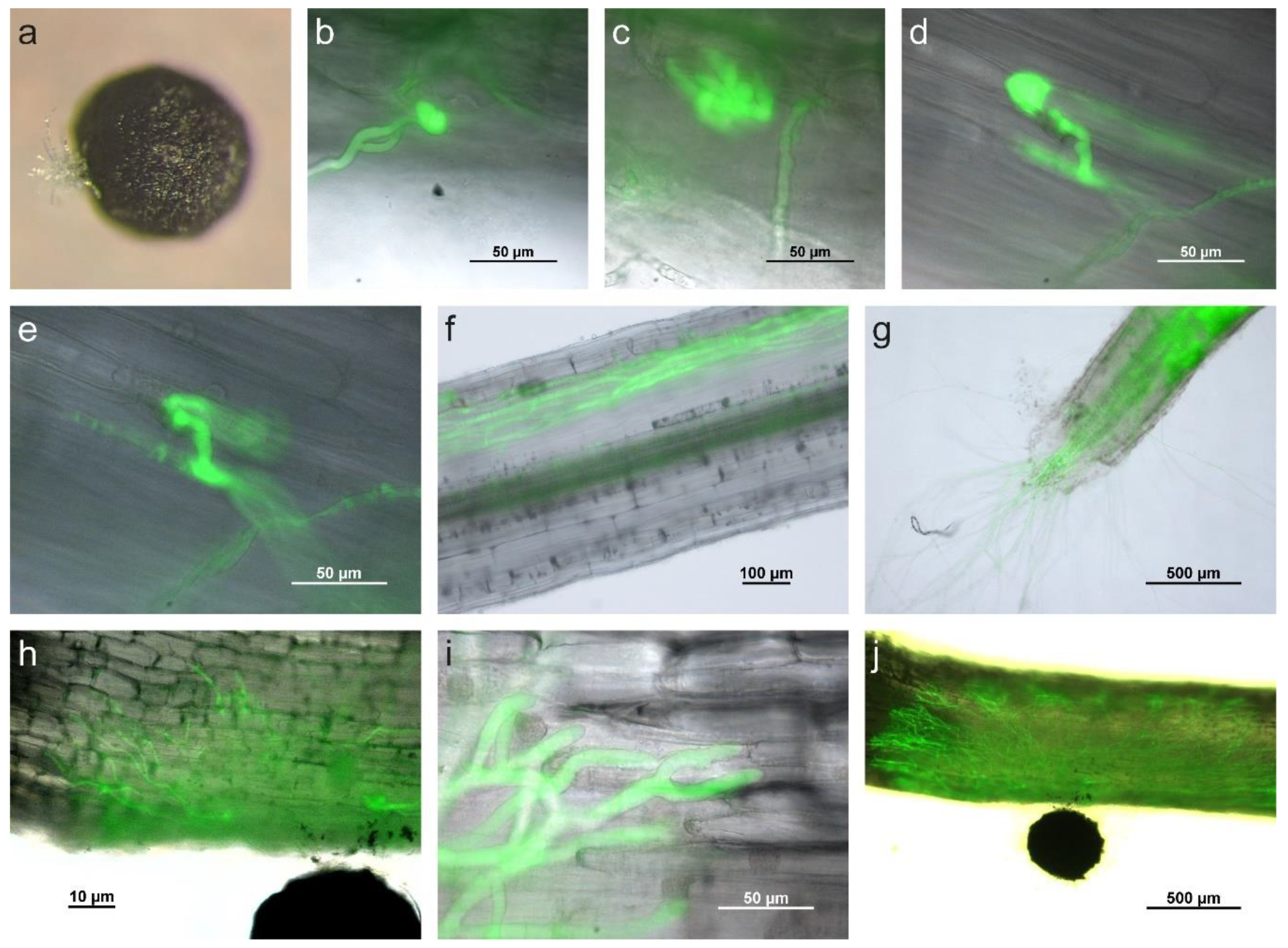
2.4. Sclerotium cepivorum Infects Onion by Formation of Appressorium-Like Structures and Infection Cushions
3. Discussion
4. Materials and Methods
4.1. Fungal Isolates and Culture Conditions
4.2. Fungal Transformation
4.3. Plant Material and Inoculation
4.4. Fluorescence Microscopy and Germ Tube Measurements
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| cv. | Cultivar |
| DPI | days post inoculation |
| GFP | green fluorescent protein |
| HPI | hours post inoculation |
| PI | propidium iodide |
References
- Carisse, O.; Tremblay, D.-M.; McDonald, M.R.; Brodeur, L.; McRoberts, N. Management of Botrytis leaf blight of onion: The Québec experience of 20 years of continual improvement. Plant Dis. 2011, 95, 504–514. [Google Scholar] [CrossRef]
- Araújo, E.R.; Alves, D.P.; Higashikawa, F.S. Cultivar resistance and chemical, biological and fertilizer treatments for controling Botrytis leaf blight of onion. Trop. Plant Pathol. 2018, 43, 160–164. [Google Scholar] [CrossRef]
- Tanović, B.; Koščica, M.; Hrustić, J.; Mihajlović, M.; Trkulja, V.; Delibašić, G. Botrytis squamosa: The causal agent of onion leaf blight in Bosnia and Herzegovina. Pestic. fitomed. 2019, 34, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Lacy, M.; Lorbeer, J. Botrytis leaf blight. In Compendium of Onion and Garlic Diseases, 2nd ed.; Schwartz, H.F., Mohan, S.F., Eds.; Am Phytopath Society: St. Paul, MN, USA, 2008; pp. 26–29. [Google Scholar]
- Steentjes, M.B.F.; Scholten, O.E.; van Kan, J.A.L. Peeling the onion: Towards a better understanding of Botrytis diseases of onion. Phytopathology 2021, in press. [Google Scholar] [CrossRef]
- Lorbeer, J. Botrytis leaf blight of onion. In Plant Diseases of International Importance. Volume II. Diseases of Vegetables and Oil Seed Crops; Chaube, H., Kumar, J., Mukhopadhyay, A., Singh, U., Eds.; Pretice-Hall, Inc.: Englewood Cliffs, NJ, USA, 1992; Volume 2, pp. 186–211. [Google Scholar]
- Lorbeer, J.W.; Seyb, A.M.; de Boer, M.; van den Ende, J.E. Botrytis species on bulb crops. In Botrytis: Biology, Pathology and Control; Elad, Y., Williamson, B., Tudzynski, P., Delen, N., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 273–294. [Google Scholar]
- Lacy, M.; Lorbeer, J. Neck rot. In Compendium of Onion and Garlic Diseases, 2nd ed.; Schwartz, H.F., Mohan, S.F., Eds.; Am Phytopath Society: St. Paul, MN, USA, 2008; pp. 47–49. [Google Scholar]
- Nielsen, K.; Yohalem, D.S. Origin of a polyploid Botrytis pathogen through interspecific hybridization between Botrytis aclada and B. byssoidea. Mycologia 2001, 93, 1064–1071. [Google Scholar] [CrossRef]
- Chilvers, M.I.; du Toit, L.J. Detection and identification of Botrytis species associated with neck rot, scape blight, and umbel blight of onion. Plant Health Prog. 2006, 7, 38. [Google Scholar] [CrossRef] [Green Version]
- Tichelaar, G. Studies on the biology of Botrytis allii on Allium cepa. Neth. J. Plant Pathol. 1967, 73, 157–160. [Google Scholar] [CrossRef]
- Maude, R. The correlation between seed-borne infection by Botrytis allii and neck rot development in store. Seed Sci. Technol. 1983, 11, 829–834. [Google Scholar]
- Maude, R.; Presly, A. Neck rot (Botrytis allii) of bulb onions: II. Seed-borne infection in relationship to the disease in store and the effect of seed treatment. Ann. Appl. Biol. 1977, 86, 181–188. [Google Scholar] [CrossRef]
- Stewart, A.; Franicevic, S. Infected seed as a source of inoculum for Botrytis infection of onion bulbs in store. Australas. Plant Pathol. 1994, 23, 36–40. [Google Scholar] [CrossRef]
- Crowe, F. White rot. In Compendium of Onion and Garlic Diseases, 2nd ed.; Schwartz, H.F., Mohan, S.F., Eds.; Am Phytopath Society: St. Paul, MN, USA, 2008; pp. 22–26. [Google Scholar]
- Sammour, R.H.; Yehia, A.-G.M.; Mustafa, A.; Alhoziem, R. Biology, controlling and genetic variability in Sclerotium cepivorum Berk; the causal agent of Allium white rot disease. Microbiology 2011, 7, 101–111. [Google Scholar]
- Coley-Smith, J. Studies of the biology of Sclerotium cepivorum Berk. IV. Germination of sclerotia. Ann. Appl. Biol. 1960, 48, 8–18. [Google Scholar] [CrossRef]
- Tyson, J.; Fullerton, R.; Elliott, G.; Reynolds, P. Use of diallyl disulphide for the commercial control of Sclerotium cepivorum. N. Z. Plant Prot. 2000, 53, 393–397. [Google Scholar] [CrossRef] [Green Version]
- Davis, R.; Hao, J.; Romberg, M.; Nunez, J.; Smith, R. Efficacy of germination stimulants of sclerotia of Sclerotium cepivorum for management of white rot of garlic. Plant Dis. 2007, 91, 204–208. [Google Scholar] [CrossRef] [Green Version]
- Coley-Smith, J. A comparison of flavour and odour compounds of onion, leek, garlic and Allium fistulosum in relation to germination of sclerotia of Sclerotium cepivorum. Plant pathol. 1986, 35, 370–376. [Google Scholar] [CrossRef]
- Presly, A. Studies on Botrytis spp. occurring on onions (Allium cepa) and leeks (Allium porrum). Plant Pathol. 1985, 34, 422–427. [Google Scholar] [CrossRef]
- Nielsen, K.; Justesen, A.F.; Jensen, D.F.; Yohalem, D.S. Universally primed polymerase chain reaction alleles and internal transcribed spacer restriction fragment length polymorphisms distinguish two subgroups in Botrytis aclada distinct from B. byssoidea. Phytopathology 2001, 91, 527–533. [Google Scholar] [CrossRef] [Green Version]
- Yohalem, D.S.; Nielsen, K.; Nicolaisen, M. Taxonomic and nomenclatural clarification of the onion neck rotting Botrytis species. Mycotaxon 2003, 85, 175–182. [Google Scholar]
- Huang, D.; Bhairi, S.; Staples, R. A transformation procedure for Botryotinia squamosa. Curr. Genet. 1989, 15, 411–414. [Google Scholar] [CrossRef]
- Leisen, T.; Bietz, F.; Werner, J.; Wegner, A.; Schaffrath, U.; Scheuring, D.; Willmund, F.; Mosbach, A.; Scalliet, G.; Hahn, M. CRISPR/Cas with ribonucleoprotein complexes and transiently selected telomere vectors allows highly efficient marker-free and multiple genome editing in Botrytis cinerea. PLoS Pathog. 2020, 16, e1008326. [Google Scholar] [CrossRef]
- Clark, C.; Lorbeer, J. Comparative histopathology of Botrytis squamosa. Phytopathology 1976, 66, 1279–1289. [Google Scholar] [CrossRef]
- Mansfield, J.; Richardson, A. The ultrastructure of interactions between Botrytis species and broad bean leaves. Physiol. Plant Pathol. 1981, 19, 41–48. [Google Scholar] [CrossRef]
- Hancock, J.; Lorbeer, J.; Millar, R. Role of pectolytic and cellulolytic enzymes in Botrytis leaf blight of onion. Phytopathology 1964, 54, 932–935. [Google Scholar]
- Kars, I.; van Kan, J.A. Extracellular enzymes and metabolites involved in pathogenesis of Botrytis. In Botrytis: Biology, Pathology and Control; Elad, Y., Williamson, B., Tudzynski, P., Delen, N., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 99–118. [Google Scholar]
- Ward, M.; Wilson, L.; Carmona, C.; Turner, G. The oliC3 gene of Aspergillus niger: Isolation, sequence and use as a selectable marker for transformation. Curr. Genet. 1988, 14, 37–42. [Google Scholar] [CrossRef]
- De Silva, D.D.; Crous, P.W.; Ades, P.K.; Hyde, K.D.; Taylor, P.W. Life styles of Colletotrichum species and implications for plant biosecurity. Fungal Biol. Rev. 2017, 31, 155–168. [Google Scholar] [CrossRef]
- Carisse, K.X.; Burnett, P.; Tewari, J.; Chen, M.; Turkington, T.; Helm, J. Histopathological study of barley cultivars resistant and susceptible to Rhynchosporium secalis. Phytopathology 2000, 90, 94–102. [Google Scholar] [CrossRef]
- Yeats, T.H.; Rose, J.K. The formation and function of plant cuticles. Plant physiol. 2013, 163, 5–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maude, R.; Presly, A. Neck rot (Botrytis allii) of bulb onions: I. Seed-borne infection and its relationship to the disease in the onion crop. Ann. Appl. Biol. 1977, 86, 163–180. [Google Scholar] [CrossRef]
- Maude, R.; Shipway, M.; Presly, A.; O’connor, D. The effects of direct harvesting and drying systems on the incidence and control of neck rot (Botrytis allii) in onions. Plant Pathol. 1984, 33, 263–268. [Google Scholar] [CrossRef]
- Coley-Smith, J.; King, J. The production by species of Allium of alkyl sulphides and their effect on germination of sclerotia of Sclerotium cepivorum Berk. Ann. Appl. Biol. 1969, 64, 289–301. [Google Scholar] [CrossRef]
- Esler, G.; Coley-Smith, J. Flavor and odour characteristics of species of Allium in relation to their capacity to stimulate germination of sclerotia of Sclerotium cepivorum. Plant Pathol. 1983, 32, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Coley-Smith, J.; Parfitt, D.; Taylor, I.; Reese, R. Studies of dormancy in sclerotia of Sclerotium cepivorum. Plant Pathol. 1987, 36, 594–599. [Google Scholar] [CrossRef]
- Allen, J.; Young, J. Soil fungistasis and Sclerotium cepivorum Berk. Plant Soil 1968, 29, 479–480. [Google Scholar] [CrossRef]
- Abd El-Razik, A.; Shatla, M.; Rushdi, M. Relationship of pectolytic enzyme production by isolates of Sclerotium cepivorum Berk to their pathogenicity. Zentralbl. Bakteriol. Parasitenkd. Infektionskr. Hyg. 1974, 129, 253–258. [Google Scholar] [CrossRef]
- Stewart, A.; Backhouse, D.; Sutherland, P.; Fullerton, R. The development of infection structures of Sclerotium cepivorum on onion. J. Phytopathol. 1989, 126, 22–32. [Google Scholar] [CrossRef]
- Stewart, A.; Fullerton, R.; Sutherland, P. Infection of onion by the white rot pathogen, Sclerotium cepivorum. J. Phytopathol. 1989, 126, 33–42. [Google Scholar] [CrossRef]
- Metcalf, D.; Wilson, C. Histology of Sclerotium cepivorum infection of onion roots and the spatial relationships of pectinases in the infection process. Plant Pathol. 1999, 48, 445–452. [Google Scholar] [CrossRef]
- Podila, G.K.; Rogers, L.M.; Kolattukudy, P.E. Chemical signals from avocado surface wax trigger germination and appressorium formation in Colletotrichum gloeosporioides. Plant Physiol. 1993, 103, 267–272. [Google Scholar] [CrossRef] [Green Version]
- Tsuba, M.; Katagiri, C.; Takeuchi, Y.; Takada, Y.; Yamaoka, N. Chemical factors of the leaf surface involved in the morphogenesis of Blumeria graminis. Physiol. Mol. Plant Pathol. 2002, 60, 51–57. [Google Scholar] [CrossRef]
- El-Samra, I.; El-Faham, Y.; Kamara, A. Selective Induction of Infection Cushions by Rhizoctonia solani in Relation to Host Responses. J. Phytopathol. 1981, 102, 122–126. [Google Scholar] [CrossRef]
- Shlezinger, N.; Minz, A.; Gur, Y.; Hatam, I.; Dagdas, Y.F.; Talbot, N.J.; Sharon, A. Anti-apoptotic machinery protects the necrotrophic fungus Botrytis cinerea from host-induced apoptotic-like cell death during plant infection. PLoS Pathog. 2011, 7, e1002185. [Google Scholar] [CrossRef]
- Ito, S.-i.; Ihara, T.; Tamura, H.; Tanaka, S.; Ikeda, T.; Kajihara, H.; Dissanayake, C.; Abdel-Motaal, F.F.; El-Sayed, M.A. α-Tomatine, the major saponin in tomato, induces programmed cell death mediated by reactive oxygen species in the fungal pathogen Fusarium oxysporum. FEBS Lett. 2007, 581, 3217–3222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valero-Jiménez, C.A.; Steentjes, M.B.; Slot, J.C.; Shi-Kunne, X.; Scholten, O.E.; van Kan, J.A.L. Dynamics in secondary metabolite gene clusters in otherwise highly syntenic and stable genomes in the fungal genus Botrytis. Genome Biol. Evol. 2020, 12, 2491–2507. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, J. Tools for Botrytis cinerea: New expression vectors make the gray mold fungus more accessible to cell biology approaches. Fungal Genet. Biol. 2012, 49, 483–497. [Google Scholar] [CrossRef]
- ten Have, A.; Mulder, W.; Visser, J.; van Kan, J.A.L. The endopolygalacturonase gene Bcpg1 is required for full virulence of Botrytis cinerea. Mol. Plant Microbe Interact. 1998, 11, 1009–1016. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Steentjes, M.B.F.; Tonn, S.; Coolman, H.; Langebeeke, S.; Scholten, O.E.; van Kan, J.A.L. Visualization of Three Sclerotiniaceae Species Pathogenic on Onion Reveals Distinct Biology and Infection Strategies. Int. J. Mol. Sci. 2021, 22, 1865. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041865
Steentjes MBF, Tonn S, Coolman H, Langebeeke S, Scholten OE, van Kan JAL. Visualization of Three Sclerotiniaceae Species Pathogenic on Onion Reveals Distinct Biology and Infection Strategies. International Journal of Molecular Sciences. 2021; 22(4):1865. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041865
Chicago/Turabian StyleSteentjes, Maikel B. F., Sebastian Tonn, Hilde Coolman, Sander Langebeeke, Olga E. Scholten, and Jan A. L. van Kan. 2021. "Visualization of Three Sclerotiniaceae Species Pathogenic on Onion Reveals Distinct Biology and Infection Strategies" International Journal of Molecular Sciences 22, no. 4: 1865. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041865