
Guide Cells Support Muscle Regeneration and Affect Neuro-Muscular Junction Organization

 , , , , , , , and
, , , , , , , and 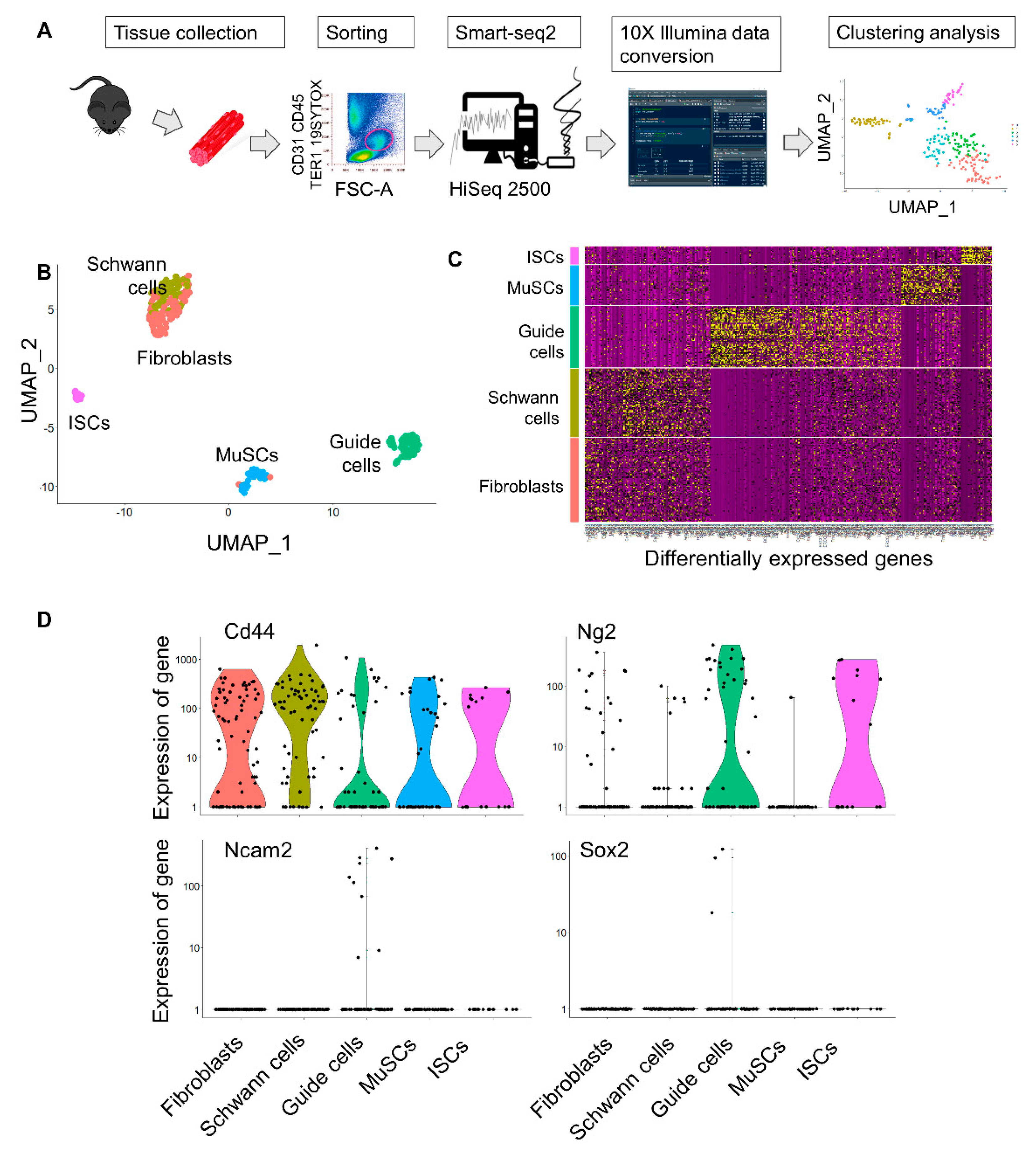{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
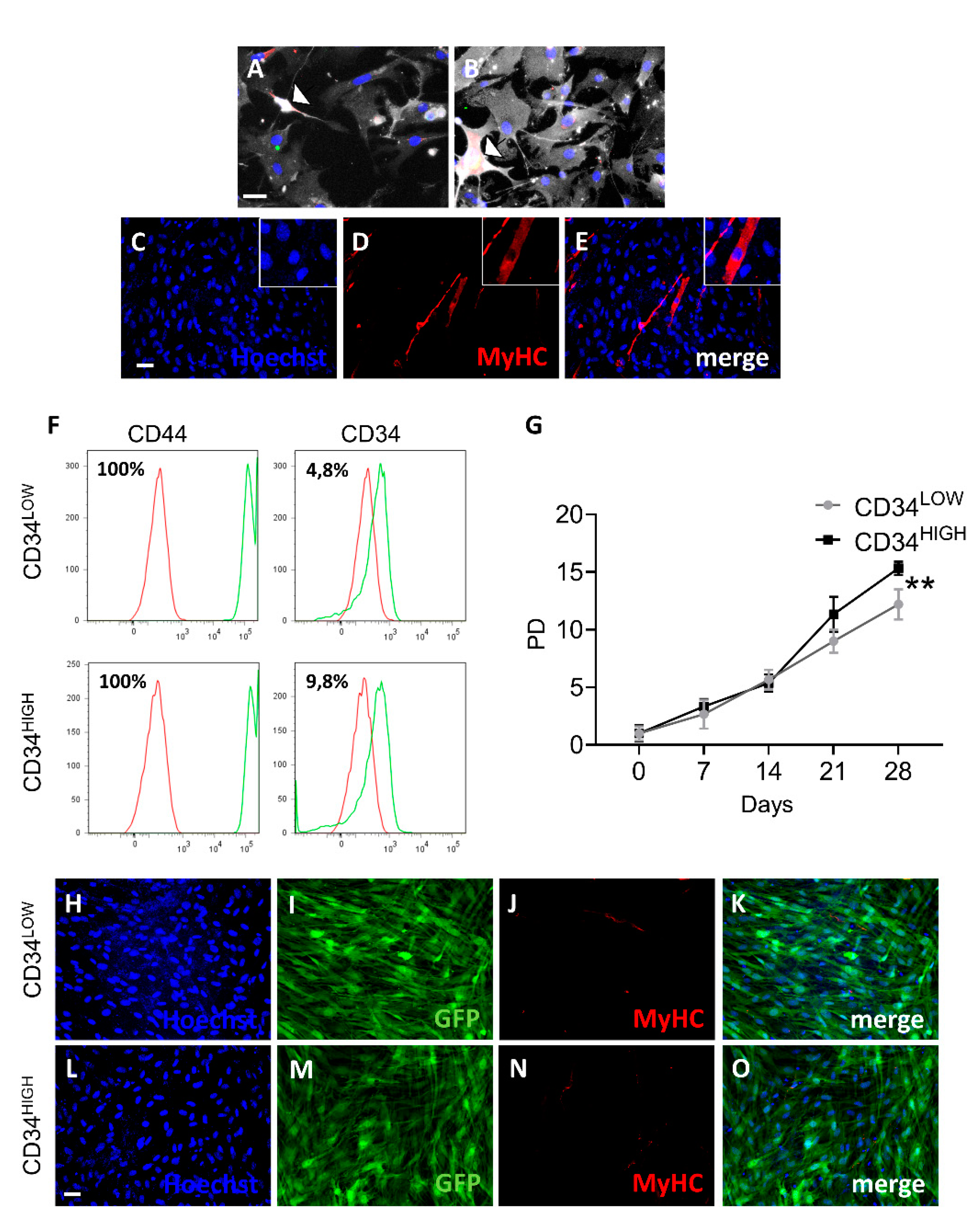
2.1. Isolation of Mesoangioblast Guide Cells
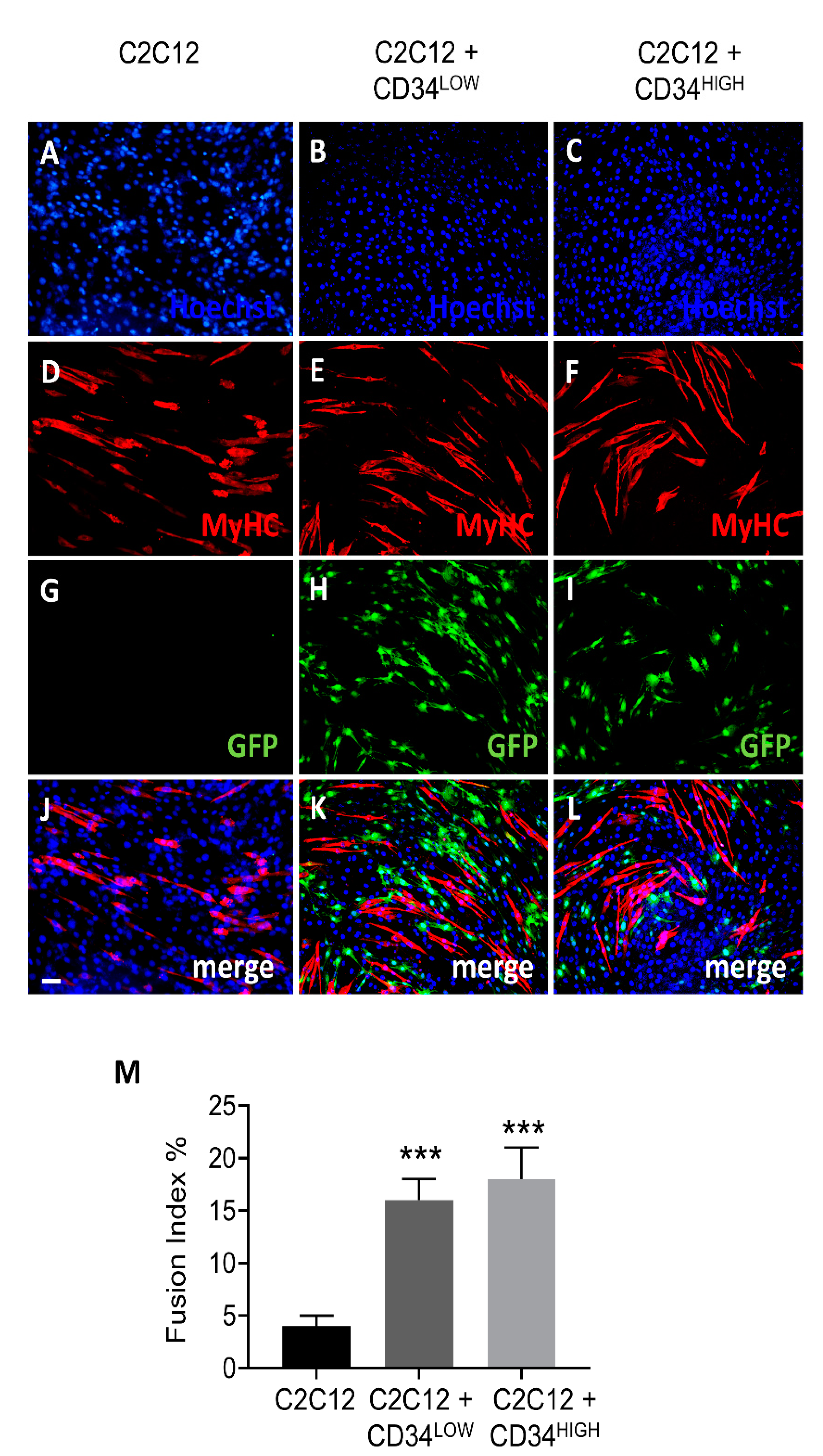
2.2. Guide Cells Increase Fusion Index of C2C12 Myoblasts and H4ven Mesoangioblasts
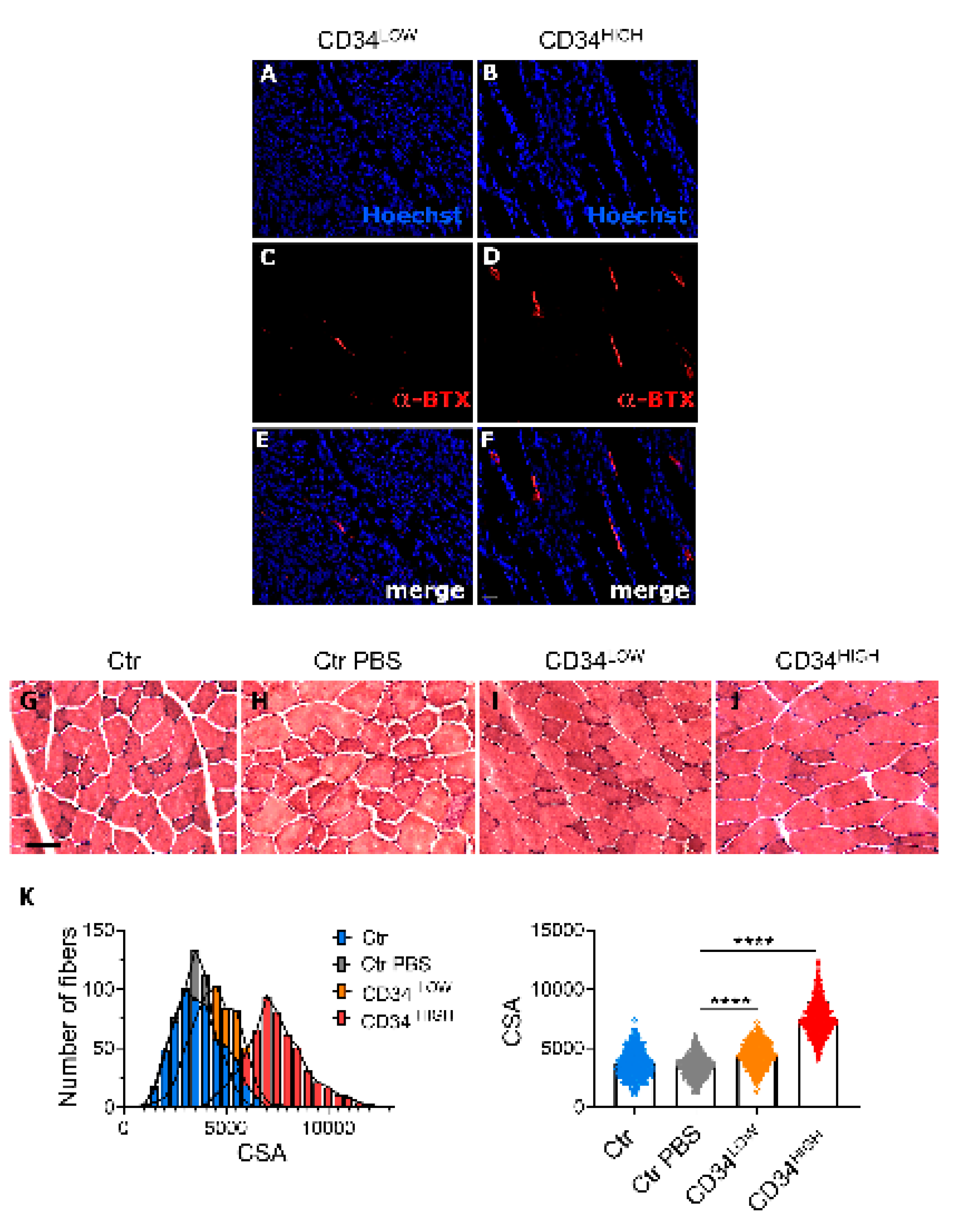
2.3. Guide CD34HIGH Cells Increased α-BTX Positive Junctions In Vivo
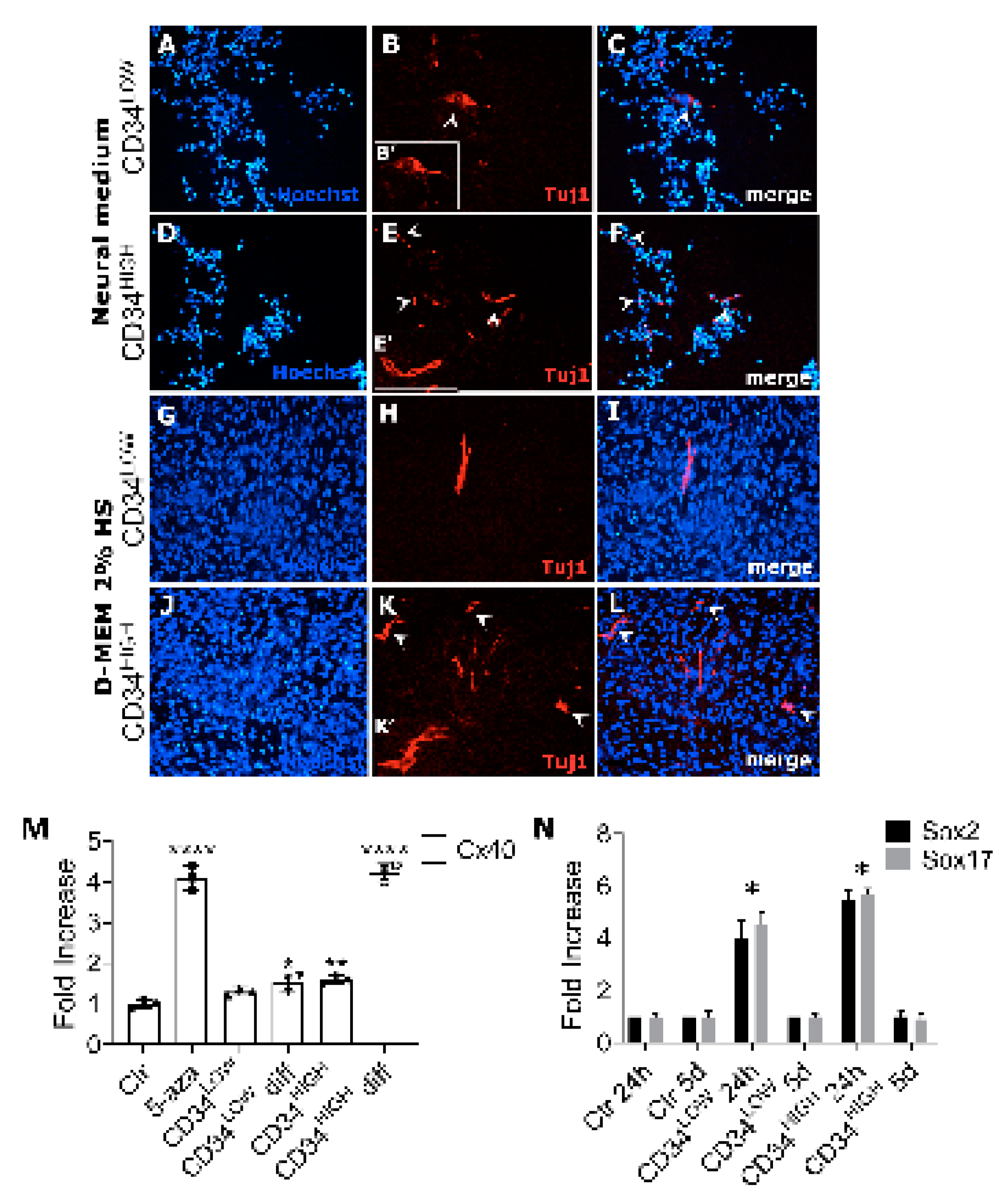
2.4. Guide Cells Differently Express Neural Markers
3. Discussion
4. Materials and Methods
4.1. Single Cell RNAseq Analysis
4.2. Cytofluorimetric Analysis
4.3. Cell Cultures
4.4. Cells and Tissue Immunofluorescence
4.5. Gene Expression Analysis
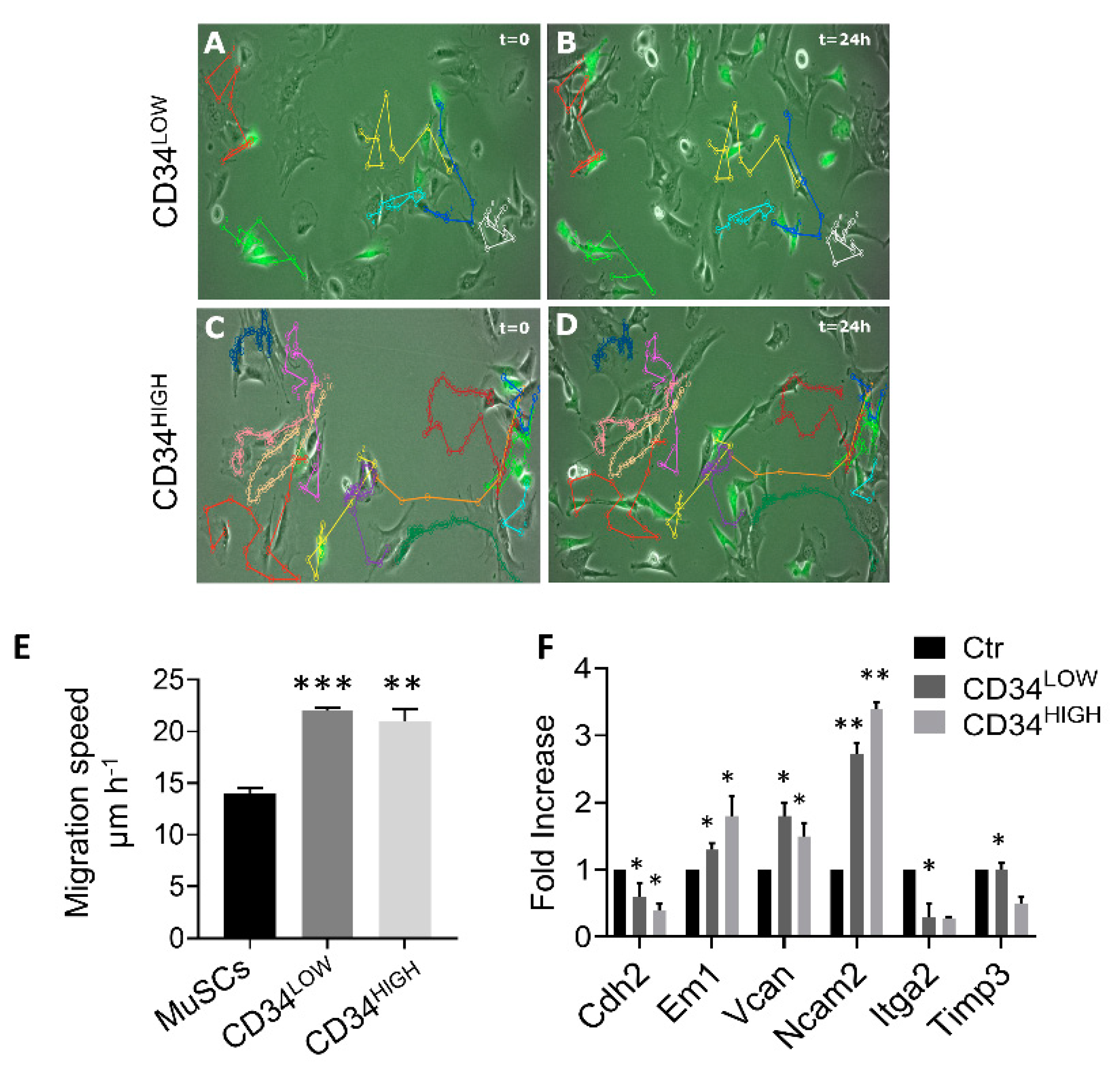
4.6. Motility Assay
4.7. In Vivo Experiments
4.8. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cossu, G.; Previtali, S.C.; Napolitano, S.; Cicalese, M.P.; Tedesco, F.S.; Nicastro, F.; Noviello, M.; Roostalu, U.; Sora, M.G.N.; Scarlato, M.; et al. Intra-arterial transplantation of HLA-matched donor mesoangioblasts in Duchenne muscular dystrophy. EMBO Mol. Med. 2015, 7, 1513–1528. [Google Scholar] [CrossRef] [PubMed]
- Galli, D.; Vitale, M.; Vaccarezza, M. Bone Marrow-Derived Mesenchymal Cell Differentiation toward Myogenic Lineages: Facts and Perspectives. BioMed Res. Int. 2014, 2014, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Shapiro, L.; Flynn, A. The clinical application of mesenchymal stem cells and cardiac stem cells as a therapy for cardiovascular disease. Pharm. Ther. 2015, 151, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Mathiasen, A.B.; Qayyum, A.A.; Jørgensen, E.; Helqvist, S.; Fischer-Nielsen, A.; Kofoed, K.F.; Haack-Sørensen, M.; Ekblond, A.; Kastrup, J. Bone marrow-derived mesenchymal stromal cell treatment in patients with severe ischaemic heart failure: A randomized placebo-controlled trial (MSC-HF trial). Eur. Heart J. 2015, 36, 1744–1753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Périé, S.; Trollet, C.; Mouly, V.; Vanneaux, V.; Mamchaoui, K.; Bouazza, B.; Marolleau, J.P.; Laforêt, P.; Chapon, F.; Eymard, B.; et al. Autologous Myoblast Transplantation for Oculopharyngeal Muscular Dystrophy: A Phase I/Iia Clinical Study. Mol. Ther. 2014, 22, 219–225. [Google Scholar] [CrossRef] [Green Version]
- Quattrocelli, M.; Costamagna, D.; Giacomazzi, G.; Camps, J.; Sampaolesi, M. Notch signaling regulates myogenic regenerative capacity of murine and human mesoangioblasts. Cell Death Dis. 2014, 5, e1448. [Google Scholar] [CrossRef] [Green Version]
- Young, P.P.; Schafer, R. Cell-based therapies for cardiac disease: A cellular therapist’s perspective. Transfusion 2015, 55, 441–451. [Google Scholar] [CrossRef] [Green Version]
- Cassano, M.; Quattrocelli, M.; Crippa, S.; Perini, I.; Ronzoni, F.; Sampaolesi, M. Cellular mechanisms and local progenitor activation to regulate skeletal muscle mass. J. Muscle. Res. Cell Motil. 2009, 30, 243–253. [Google Scholar] [CrossRef] [Green Version]
- Duelen, R.; Sampaolesi, M. Stem Cell Technology in Cardiac Regeneration: A Pluripotent Stem Cell Promise. EBioMedicine 2017, 16, 30–40. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, T.Y.; Iwata, Y.; Sampaolesi, M.; Hanada, H.; Saito, N.; Artman, M.; Coetzee, W.A.; Shigekawa, M. Stretch-activated cation channels in skeletal muscle myotubes from sarcoglycan-deficient hamsters. Am. J. Physiol. Physiol. 2001, 281, C690–C699. [Google Scholar] [CrossRef]
- Seuntjens, E.; Umans, L.; Zwijsen, A.; Sampaolesi, M.; Verfaillie, C.M.; Huylebroeck, D. Transforming Growth Factor type β and Smad family signaling in stem cell function. Cytokine Growth Factor Rev. 2009, 20, 449–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costamagna, D.; Quattrocelli, M.; Duelen, R.; Sahakyan, V.; Perini, I.; Palazzolo, G.; Sampaolesi, M. Fate choice of post-natal mesoderm progenitors: Skeletal versuscardiac muscle plasticity. Cell. Mol. Life Sci. 2014, 71, 615–627. [Google Scholar] [CrossRef] [PubMed]
- Ronzoni, F.L.; Lemeille, S.; Kuzyakiv, R.; Sampaolesi, M.; Jaconi, M.E. Human fetal mesoangioblasts reveal tissue-dependent transcriptional signatures. Stem Cells Transl. Med. 2020, 9, 575–589. [Google Scholar] [CrossRef] [Green Version]
- Lescroart, F.; Hamou, W.; Francou, A.; Théveniau-Ruissy, M.; Kelly, R.G.; Buckingham, M. Clonal analysis reveals a common origin between nonsomite-derived neck muscles and heart myocardium. Proc. Natl. Acad. Sci. USA 2015, 112, 1446–1451. [Google Scholar] [CrossRef] [Green Version]
- Quattrocelli, M.; Sampaolesi, M. The mesmiRizing complexity of microRNAs for striated muscle tissue engineering. Adv. Drug Deliv. Rev. 2015, 88, 37–52. [Google Scholar] [CrossRef] [Green Version]
- Quattrocelli, M.; Swinnen, M.; Giacomazzi, G.; Camps, J.; Barthélemy, I.; Ceccarelli, G.; Caluwé, E.; Grosemans, H.; Thorrez, L.; Pelizzo, G.; et al. Mesodermal iPSC–derived progenitor cells functionally regenerate cardiac and skeletal muscle. J. Clin. Investig. 2015, 125, 4463–4482. [Google Scholar] [CrossRef] [Green Version]
- Hashem, S.I.; Claycomb, W.C. Genetic isolation of stem cell-derived pacemaker-nodal cardiac myocytes. Mol. Cell Biochem. 2013, 383, 161–171. [Google Scholar] [CrossRef]
- Chauveau, S.; Brink, P.R.; Cohen, I.S. Stem cell-based biological pacemakers from proof of principle to therapy: A review. Cytotherapy 2014, 16, 873–880. [Google Scholar] [CrossRef] [Green Version]
- Toma, J.S.; Shettar, B.C.; Chipman, P.H.; Pinto, D.M.; Borowska, J.P.; Ichida, J.K.; Fawcett, J.P.; Zhang, Y.; Eggan, K.; Rafuse, V. Motoneurons Derived from Induced Pluripotent Stem Cells Develop Mature Phenotypes Typical of Endogenous Spinal Motoneurons. J. Neurosci. 2015, 35, 1291–1306. [Google Scholar] [CrossRef] [Green Version]
- Lavasani, M.; Thompson, S.D.; Pollett, J.B.; Usas, A.; Lu, A.; Stolz, N.B.; Clark, K.A.; Sun, B.; Péault, B.; Huard, J. Human muscle-derived stem/progenitor cells promote functional murine peripheral nerve regeneration. J. Clin. Investig. 2014, 124, 1745–1756. [Google Scholar] [CrossRef] [Green Version]
- Kelder, T.P.; Vicente-Steijn, R.; Harryvan, T.J.; Kosmidis, G.; Groot, A.C.G.; Poelmann, R.E.; Schalij, M.J.; DeRuiter, M.C.; Jongbloed, M. The sinus venosus myocardium contributes to the atrioventricular canal: Potential role during atrioventricular node development? J. Cell. Mol. Med. 2015, 19, 1375–1389. [Google Scholar] [CrossRef]
- Galvez, B.G.; Sampaolesi, M.; Barbuti, A.; Crespi, A.; Covarello, D.; Brunelli, S.; Dellavalle, A.; Crippa, S.; Balconi, G.; Cuccovillo, I.; et al. Cardiac mesoangioblasts are committed, self-renewable progenitors, associated with small vessels of juvenile mouse ventricle. Cell Death Differ. 2008, 15, 1417–1428. [Google Scholar] [CrossRef] [Green Version]
- Sampaolesi, M.; Torrente, Y.; Innocenzi, A.; Tonlorenzi, R.; D’Antona, G.; Pellegrino, M.A.; Barresi, R.; Bresolin, N.; De Angelis, M.G.C.; Campbell, K.P.; et al. Cell Therapy of α-Sarcoglycan Null Dystrophic Mice Through Intra-Arterial Delivery of Mesoangioblasts. Science 2003, 301, 487–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sampaolesi, M.; Blot, S.; D’Antona, G.; Granger, N.; Tonlorenzi, R.; Innocenzi, A.; Mognol, P.; Thibaud, J.-L.; Galvez, B.G.; Barthélémy, I.; et al. Mesoangioblast stem cells ameliorate muscle function in dystrophic dogs. Nature 2006, 444, 574–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbuti, A.; Gálvez, B.G.; Crespi, A.; Scavone, A.; Baruscotti, M.; Brioschi, C.; Cossu, G.; DiFrancesco, D. Mesoangioblasts from ventricular vessels can differentiate in vitro into cardiac myocytes with sinoatrial-like properties. J. Mol. Cell. Cardiol. 2010, 48, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Camps, J.; Breuls, N.; Sifrim, A.; Giarratana, N.; Corvelyn, M.; Danti, L.; Grosemans, H.; VanUytven, S.; Thiry, I.; Belicchi, M.; et al. Interstitial Cell Remodeling Promotes Aberrant Adipogenesis in Dystrophic Muscles. Cell Rep. 2020, 31, 107597. [Google Scholar] [CrossRef]
- Tonlorenzi, R.; Dellavalle, A.; Schnapp, E.; Cossu, G.; Sampaolesi, M. Isolation and Characterization of Mesoangioblasts from Mouse, Dog, and Human Tissues. Curr. Protoc. Stem Cell Biol. 2007, 3, 2B.1.1–2B.1.29. [Google Scholar] [CrossRef]
- Kim, D.S.; Lee, D.R.; Kim, H.-S.; Yoo, J.E.; Jung, S.J.; Lim, B.Y.; Jang, J.; Kang, H.C.; You, S.; Hwang, D.-Y.; et al. Highly Pure and Expandable PSA-NCAM-Positive Neural Precursors from Human ESC and iPSC-Derived Neural Rosettes. PLoS ONE 2012, 7, e39715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pevny, L.H.; Nicolis, S.K. Sox2 roles in neural stem cells. Int. J. Biochem. Cell Biol. 2010, 42, 421–424. [Google Scholar] [CrossRef]
- Tanaka, H.; Takafuji, K.; Taguchi, A.; Wiriyasermkul, P.; Ohgaki, R.; Nagamori, S.; Suh, P.-G.; Kanai, Y. Linkage of N-cadherin to multiple cytoskeletal elements revealed by a proteomic approach in hippocampal neurons. Neurochem. Int. 2012, 61, 240–250. [Google Scholar] [CrossRef]
- Litteri, G.; Carnevale, D.; D’Urso, A.; Cifelli, G.; Braghetta, P.; Damato, A.; Bizzotto, D.; Landolfi, A.; Da Ros, F.; Sabatelli, P.; et al. Vascular Smooth Muscle Emilin-1 Is a Regulator of Arteriolar Myogenic Response and Blood Pressure. Arter. Thromb. Vasc. Biol. 2012, 32, 2178–2184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burns, T.A.; Dours-Zimmermann, M.T.; Zimmermann, D.R.; Krug, E.L.; Comte-Walters, S.; Reyes, L.; Davis, M.A.; Schey, K.L.; Schwacke, J.H.; Kern, C.B.; et al. Imbalanced Expression of Vcan mRNA Splice Form Proteins Alters Heart Morphology and Cellular Protein Profiles. PLoS ONE 2014, 9, e89133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paoloni-Giacobino, A.; Chen, H.; Antonarakis, S.E. Cloning of a novel human neural cell adhesion molecule gene (NCAM2) that maps to chromosome region 21q21 and is potentially involved in Down syndrome. Genomics 1997, 43, 43–51. [Google Scholar] [CrossRef]
- Das, A.M.; Seynhaeve, A.L.B.; Rens, J.A.P.; Vermeulen, C.E.; Koning, G.A.; Eggermont, A.M.M.; Hagen, T.T. Differential TIMP3 expression affects tumor progression and angiogenesis in melanomas through regulation of directionally persistent endothelial cell migration. Angiogenesis 2013, 17, 163–177. [Google Scholar] [CrossRef] [PubMed]
- Crippa, S.; Cassano, M.; Messina, G.; Galli, D.; Galvez, B.G.; Curk, T.; Altomare, C.; Ronzoni, F.; Toelen, J.; Gijsbers, R.; et al. miR669a and miR669q prevent skeletal muscle differentiation in postnatal cardiac progenitors. J. Cell Biol. 2011, 193, 1197–1212. [Google Scholar] [CrossRef] [PubMed]
- De Sio, F.L.R.S.; Gritti, A.; Cascio, P.; Neri, M.; Sampaolesi, M.; Galli, C.; Luban, J.; Naldini, L. Lentiviral Vector Gene Transfer Is Limited by the Proteasome at Postentry Steps in Various Types of Stem Cells. Stem Cells 2008, 26, 2142–2152. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.Y. Chemistry and pharmacology of polypeptide toxins in snake venoms. Annu. Rev. Pharmacol. 1972, 12, 265–286. [Google Scholar] [CrossRef]
- Parkington, J.D.; LeBrasseur, N.K.; Siebert, A.P.; Fielding, R.A. Contraction-mediated mTOR, p70S6k, and ERK1/2 phosphorylation in aged skeletal muscle. J. Appl. Physiol. 2004, 97, 243–248. [Google Scholar] [CrossRef] [Green Version]
- Leaf, D.E.; Feig, J.E.; Vasquez, C.; Riva, P.L.; Yu, C.; Lader, J.M.; Kontogeorgis, A.; Baron, E.L.; Peters, N.S.; Fisher, E.A.; et al. Connexin40 Imparts Conduction Heterogeneity to Atrial Tissue. Circ. Res. 2008, 103, 1001–1008. [Google Scholar] [CrossRef] [PubMed]
- Agosti, E.; De Feudis, M.; Angelino, E.; Belli, R.; Teixeira, M.A.; Zaggia, I.; Tamiso, E.; Raiteri, T.; Scircoli, A.; Ronzoni, F.L.; et al. Both ghrelin deletion and unacylated ghrelin overexpression preserve muscles in aging mice. Aging 2020, 12, 13939–13957. [Google Scholar] [CrossRef]
- Perini, I.; Elia, I.; Nigro, A.L.; Ronzoni, F.; Berardi, E.; Grosemans, H.; Fukada, S.-I.; Sampaolesi, M. Myogenic induction of adult and pluripotent stem cells using recombinant proteins. Biochem. Biophys. Res. Commun. 2015, 464, 755–761. [Google Scholar] [CrossRef] [Green Version]
- Ronzoni, F.; Bongio, M.; Conte, S.; Vercesi, L.; Cassano, M.; Tribioli, C.; Galli, D.; Bellazzi, R.; Magenes, G.; De Angelis, M.G.C.; et al. Localization of Magic-F1 Transgene, Involved in Muscular Hypertrophy, during Early Myogenesis. J. Biomed. Biotechnol. 2011, 2011, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Ronzoni, F.; Ceccarelli, G.; Perini, I.; Benedetti, L.; Galli, D.; Mulas, F.; Balli, M.; Magenes, G.; Bellazzi, R.; De Angelis, G.C.; et al. Met-Activating Genetically Improved Chimeric Factor-1 Promotes Angiogenesis and Hypertrophy in Adult Myogenesis. Curr. Pharm. Biotechnol. 2017, 18, 309–317. [Google Scholar] [CrossRef]
- Tirosh-Finkel, L.; Elhanany, H.; Rinon, A.; Tzahor, E. Mesoderm progenitor cells of common origin contribute to the head musculature and the cardiac outflow tract. Development 2006, 133, 1943–1953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Dévédec, S.E.; Yan, K.; De Bont, H.; Ghotra, V.; Truong, H.; Danen, E.H.; Verbeek, F.; Van De Water, B. Systems microscopy approaches to understand cancer cell migration and metastasis. Cell. Mol. Life Sci. 2010, 67, 3219–3240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singer, A.J.; Clark, R.A. Cutaneous wound healing. N. Engl. J. Med. 1999, 341, 738–746. [Google Scholar] [CrossRef]
- Entschladen, F.; Drell, I.V.T.L.; Lang, K.; Masur, K.; Palm, D.; Bastian, P.; Niggemann, B.; Zaenker, K.S. Analysis methods of human cell migration. Exp. Cell Res. 2005, 307, 418–426. [Google Scholar] [CrossRef]
- Mylona, E.; Jones, K.A.; Mills, S.T.; Pavlath, G.K. CD44 regulates myoblast migration and differentiation. J. Cell Physiol. 2006, 209, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Lynch, G.S.; Ryall, J.G. Role of beta-adrenoceptor signaling in skeletal muscle: Implications for muscle wasting and disease. Physiol. Rev. 2008, 88, 729–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picelli, S.; Faridani, O.R.; Björklund, Å.K.; Winberg, G.; Sagasser, S.; Sandberg, R. Full-length RNA-seq from single cells using Smart-seq2. Nat. Protoc. 2014, 9, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Butler, A.; Hoffman, P.; Smibert, P.; Papalexi, E.; Satija, R. Integrating single-cell transcriptomic data across different conditions, technologies, and species. Nat. Biotechnol. 2018, 36, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Galli, D.; Carubbi, C.; Masselli, E.; Corradi, D.; Cas, A.D.; Nouvenne, A.; Bucci, G.; Arcari, M.; Mirandola, P.; Vitale, M.; et al. PKCε is a negative regulator of PVAT-derived vessel formation. Exp. Cell Res. 2015, 330, 277–286. [Google Scholar] [CrossRef]
- Galvez, B.G.; Sampaolesi, M.; Barbuti, A. Complete repair of dystrophic skeletal muscle by mesoangioblasts with enhanced migration ability. J. Cell Biol. 2006, 174, 231–243. [Google Scholar] [CrossRef] [Green Version]
- Martino, S.; Di Girolamo, I.; Cavazzin, C.; Tiribuzi, R.; Galli, R.; Rivaroli, A.; Valsecchi, M.; Sandhoff, K.; Sonnino, S.; Vescovi, A.; et al. Neural precursor cell cultures from GM2 gangliosidosis animal models recapitulate the biochemical and molecular hallmarks of the brain pathology. J. Neurochem. 2009, 109, 135–147. [Google Scholar] [CrossRef] [PubMed]
- Martini, S.; Soliman, T.; Gobbi, G.; Mirandola, P.; Carubbi, C.; Masselli, E.; Pozzi, G.; Parker, P.J.; Vitale, M. PKCε Controls Mitotic Progression by Regulating Centrosome Migration and Mitotic Spindle Assembly. Mol. Cancer Res. 2018, 16, 3–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertolo, A.; Gemperli, A.; Gruber, M.; Gantenbein, B.; Baur, M.; Pötzel, T.; Stoyanov, J.V. In vitro cell motility as a potential mesenchymal stem cell marker for multipotency. Stem Cells Transl. Med. 2014, 4, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Giarratana, N.; Conti, F.; La Rovere, R.; Gijsbers, R.; Carai, P.; Duelen, R.; Vervliet, T.; Bultynck, G.; Ronzoni, F.; Piciotti, R.; et al. MICAL2 is essential for myogenic lineage commitment. Cell Death Dis. 2020, 11, 1–19. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ronzoni, F.L.; Giarratana, N.; Crippa, S.; Quattrocelli, M.; Cassano, M.; Ceccarelli, G.; Benedetti, L.; Van Herck, J.; Cusella De Angelis, M.G.; Vitale, M.; et al. Guide Cells Support Muscle Regeneration and Affect Neuro-Muscular Junction Organization. Int. J. Mol. Sci. 2021, 22, 1939. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041939
Ronzoni FL, Giarratana N, Crippa S, Quattrocelli M, Cassano M, Ceccarelli G, Benedetti L, Van Herck J, Cusella De Angelis MG, Vitale M, et al. Guide Cells Support Muscle Regeneration and Affect Neuro-Muscular Junction Organization. International Journal of Molecular Sciences. 2021; 22(4):1939. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041939
Chicago/Turabian StyleRonzoni, Flavio L., Nefele Giarratana, Stefania Crippa, Mattia Quattrocelli, Marco Cassano, Gabriele Ceccarelli, Laura Benedetti, Jens Van Herck, Maria G. Cusella De Angelis, Marco Vitale, and et al. 2021. "Guide Cells Support Muscle Regeneration and Affect Neuro-Muscular Junction Organization" International Journal of Molecular Sciences 22, no. 4: 1939. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041939