
The Multi-Targeting Ligand ST-2223 with Histamine H3 Receptor and Dopamine D2/D3 Receptor Antagonist Properties Mitigates Autism-Like Repetitive Behaviors and Brain Oxidative Stress in Mice

 , , and
, , and 
Abstract
:1. Introduction
2. Results
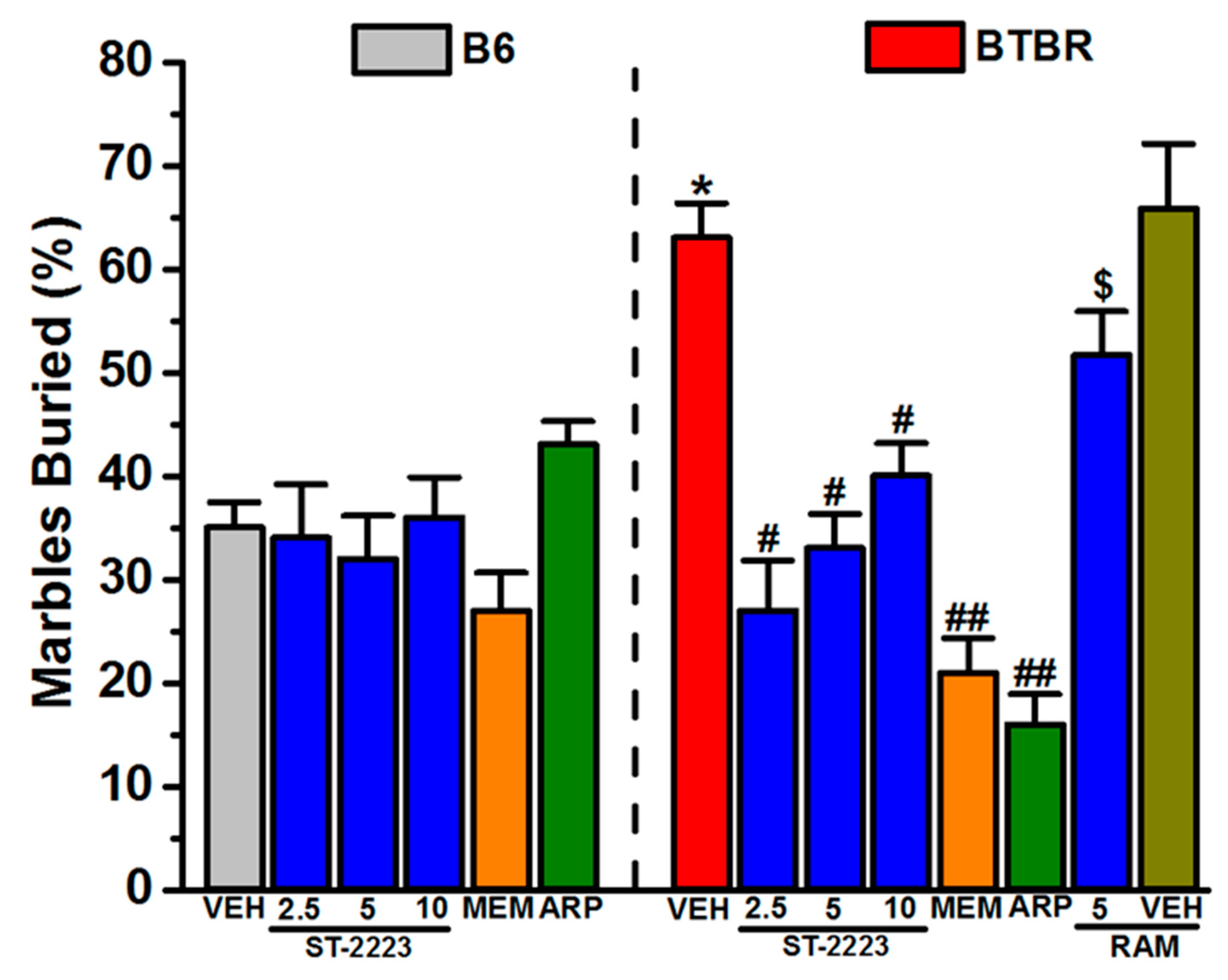
2.1. Marble Burying Behavior
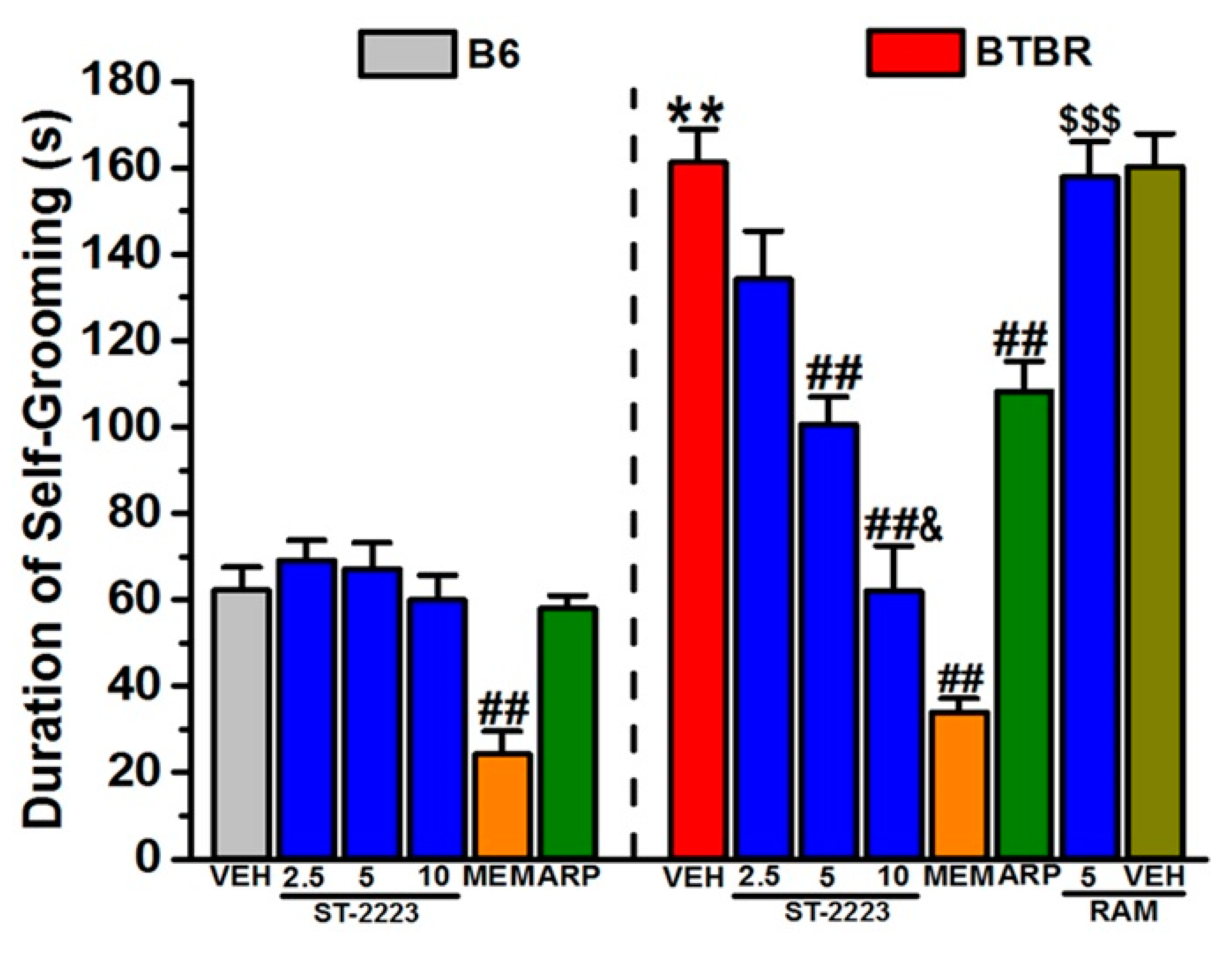
2.2. Spontaneous Self-Grooming Behavior
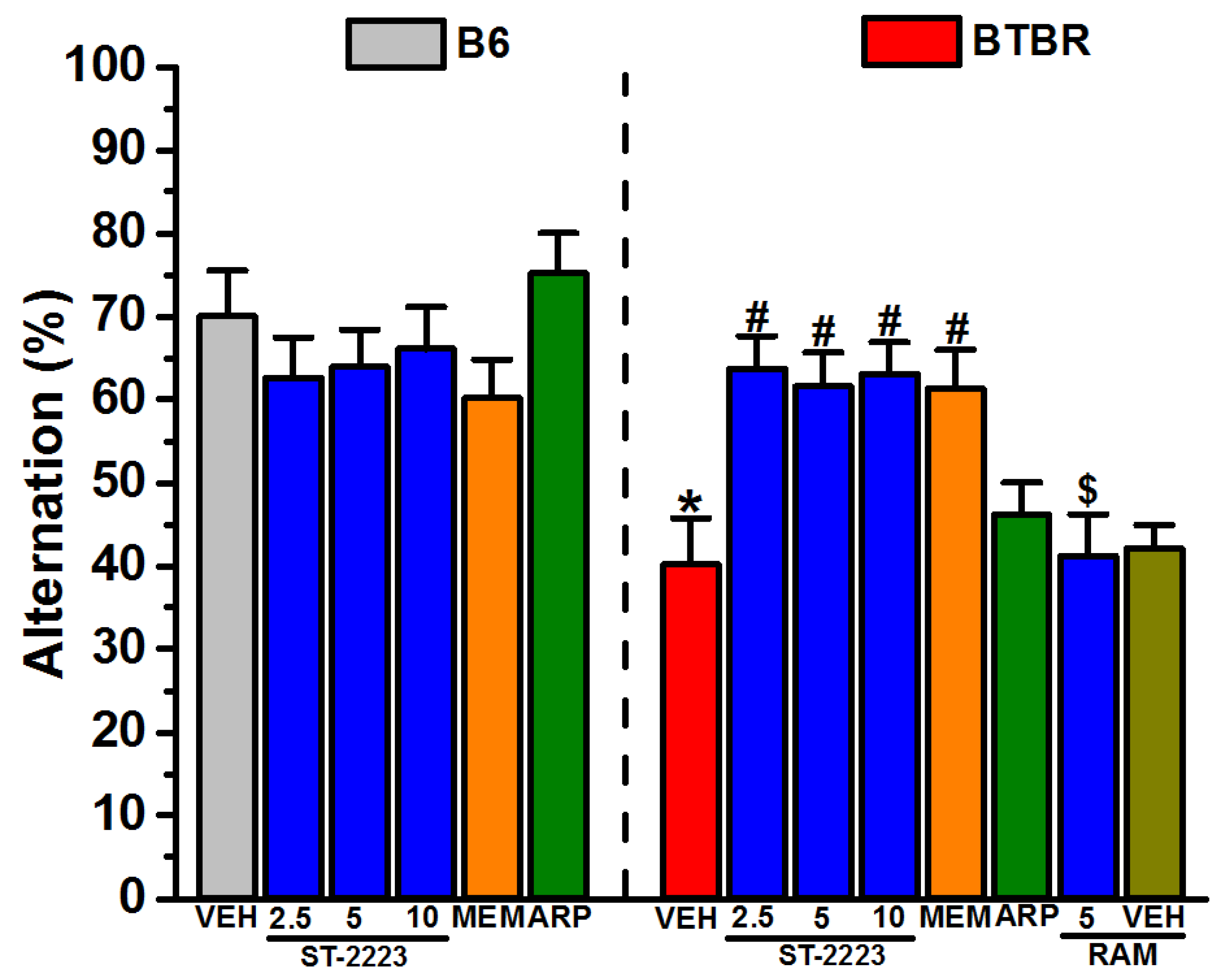
2.3. Spontaneous Alternation Behavior
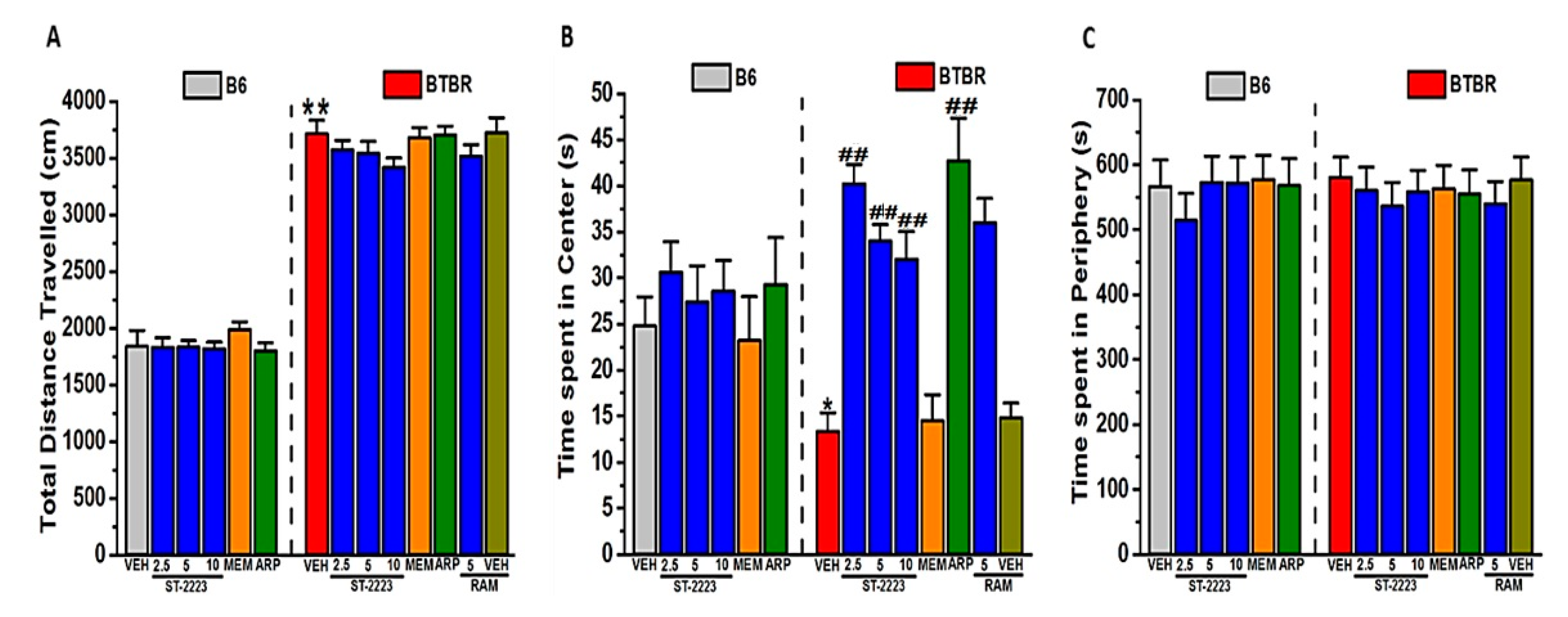
2.4. Locomotor Activity and Anxiety Levels
2.5. Levels of Oxidative Stress Markers in Different Brain Parts of Treated BTBR Mice
3. Discussion
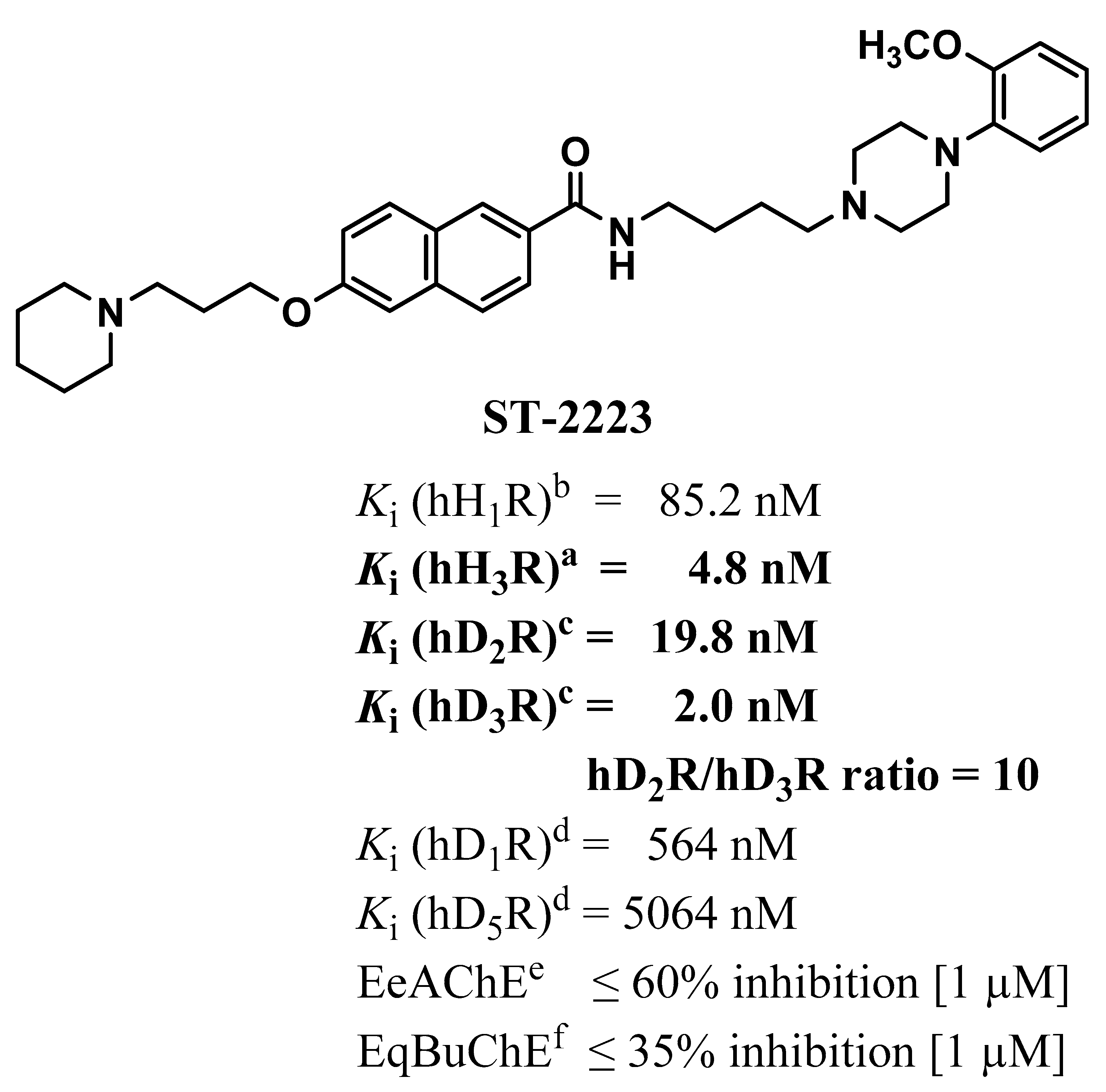
3.1. In Vitro Affinities for hH1Rs, hH3Rs, hD1Rs, hD2Rs, hD3Rs and hD5Rs
3.2. In Vivo Ameliorative Effects of ST-2223 on Behaviors of Treated BTBR Mice
3.3. In Vivo Mitigating Effects of ST-2223 on Oxidative Stress in Different Brain Areas of Treated BTBR Mice
4. Materials and Methods
4.1. Animals
4.2. Drugs
4.3. In Vitro Pharmacological Binding Assays for ST-2223
4.4. In Vivo Behavioral Tests
4.4.1. Marble Burying Task (MBT)
4.4.2. Self-Grooming Paradigm (SGP)
4.4.3. Spontaneous Alternation Behavior (SAB)
4.4.4. Locomotor Activity
4.5. Brain Collection and Tissue Processing for Biochemical Analyses
4.5.1. Oxidative Stress Marker Estimations
Malondialdehyde (MDA) Assay
Quantification of Glutathione (GSH)
4.5.2. Assay of Antioxidant Enzymes Activities
4.6. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Amodeo, D.A.; Rivera, E.; Cook, E.H., Jr.; Sweeney, J.A.; Ragozzino, M.E. 5HT2A receptor blockade in dorsomedial striatum reduces repetitive behaviors in BTBR mice. Genes Brain Behav. 2017, 16, 342–351. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.C.; Cole, T.B.; Costa, L.G. Behavioral Phenotyping for Autism Spectrum Disorders in Mice. Curr. Protoc. Toxicol. 2017, 72, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Fitzpatrick, S.E.; Srivorakiat, L.; Wink, L.K.; Pedapati, E.V.; Erickson, C.A. Aggression in autism spectrum disorder: Presentation and treatment options. Neuropsychiatr. Dis. Treat. 2016, 12, 1525–1538. [Google Scholar]
- Bishop, S.L.; Richler, J.; Cain, A.C.; Lord, C. Predictors of Perceived Negative Impact in Mothers of Children with Autism Spectrum Disorder. Am. J. Ment. Retard. 2007, 112, 450–461. [Google Scholar] [CrossRef]
- Lewis, M.H.; Bodfish, J.W. Repetitive behavior disorders in autism. Ment. Retard. Dev. Disabil. Res. Rev. 1998, 4, 80–89. [Google Scholar] [CrossRef]
- D’Cruz, A.M.; Ragozzino, M.E.; Mosconi, M.W.; Shrestha, S.; Cook, E.H.; Sweeney, J.A. Reduced behavioral flexibility in autism spectrum disorders. Neuropsychology 2013, 27, 152–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabriels, R.L.; Cuccaro, M.L.; Hill, D.E.; Ivers, B.J.; Goldson, E. Repetitive behaviors in autism: Relationships with associated clinical features. Res. Dev. Disabil. 2005, 26, 169–181. [Google Scholar] [CrossRef]
- Miller, H.L.; Ragozzino, M.E.; Cook, E.H.; Sweeney, J.A.; Mosconi, M.W. Cognitive set shifting deficits and their relationship to repetitive behaviors in autism spectrum disorder. J. Autism Dev. Disord. 2015, 45, 805–815. [Google Scholar] [CrossRef] [PubMed]
- Boyd, B.A.; McDonough, S.G.; Bodfish, J.W. Evidence-based behavioral interventions for repetitive behaviors in autism. J. Autism Dev. Disord. 2012, 42, 1236–1248. [Google Scholar] [CrossRef] [PubMed]
- Ravizza, S.M.; Solomon, M.; Ivry, R.B.; Carter, C.S. Restricted and repetitive behaviors in autism spectrum disorders: The relationship of attention and motor deficits. Dev. Psychopathol. 2013, 25, 773–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masi, A.; DeMayo, M.M.; Glozier, N.; Guastella, A.J. An Overview of Autism Spectrum Disorder, Heterogeneity and Treatment Options. Neurosci. Bull. 2017, 33, 183–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wink, L.K.; Plawecki, M.H.; Erickson, C.A.; Stigler, K.A.; McDougle, C.J. Emerging drugs for the treatment of symptoms associated with autism spectrum disorders. Expert Opin. Emerg. Drugs 2010, 15, 481–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyatsanza, S.; Shetty, T.; Gregory, C.; Lough, S.; Dawson, K.; Hodges, J.R. A study of stereotypic behaviours in Alzheimer’s disease and frontal and temporal variant frontotemporal dementia. J. Neurol. Neurosurg. Psychiatry 2003, 74, 1398–1402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frith, C.D.; Done, D.J. Stereotyped behaviour in madness and in health. In Neurobiology of Stereotyped Behaviour; Clarendon Press/Oxford University Press: New York, NY, USA, 1990; pp. 232–259. [Google Scholar]
- Evans, D.W.; Uljarevic, M.; Lusk, L.G.; Loth, E.; Frazier, T. Development of Two Dimensional Measures of Restricted and Repetitive Behavior in Parents and Children. J. Am. Acad. Child Adolesc. Psychiatry 2017, 56, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Darrow, S.M.; Grados, M.; Sandor, P.; Hirschtritt, M.E.; Illmann, C.; Osiecki, L.; Dion, Y.; King, R.; Pauls, D.; Budman, C.L.; et al. Autism Spectrum Symptoms in a Tourette’s Disorder Sample. J. Am. Acad. Child Adolesc. Psychiatry 2017, 56, 610–617. [Google Scholar] [CrossRef] [Green Version]
- Hommer, R.E.; Swedo, S.E. Schizophrenia and autism-related disorders. Schizophr. Bull. 2015, 41, 313–314. [Google Scholar] [CrossRef]
- Guo, W.; Samuels, J.F.; Wang, Y.; Cao, H.; Ritter, M.; Nestadt, P.S.; Krasnow, J.; Greenberg, B.D.; Fyer, A.J.; McCracken, J.T.; et al. Polygenic risk score and heritability estimates reveals a genetic relationship between ASD and OCD. Eur. Neuropsychopharmacol. 2017, 27, 657–666. [Google Scholar] [CrossRef] [PubMed]
- Eissa, N.; Al-Houqani, M.; Sadeq, A.; Ojha, S.K.; Sasse, A.; Sadek, B. Current Enlightenment About Etiology and Pharmacological Treatment of Autism Spectrum Disorder. Front. Neurosci. 2018, 12, 304. [Google Scholar] [CrossRef] [Green Version]
- Santangelo, A.; Passani, M.B.; Casarrubea, M. Brain histamine and behavioral neuroscience. Oncotarget 2017, 8, 16107–16108. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.; Kaminga, A.C.; Wen, S.W.; Wu, X.; Acheampong, K.; Liu, A. Dopamine and Dopamine Receptors in Alzheimer’s Disease: A Systematic Review and Network Meta-Analysis. Front. Aging Neurosci. 2019, 11, 175. [Google Scholar] [CrossRef] [Green Version]
- DiCarlo, G.E.; Aguilar, J.I.; Matthies, H.J.G.; Harrison, F.E.; Bundschuh, K.E.; West, A.; Hashemi, P.; Herborg, F.; Rickhag, M.; Chen, H.; et al. Autism-linked dopamine transporter mutation alters striatal dopamine neurotransmission and dopamine-dependent behaviors. J. Clin. Investig. 2019, 129, 3407–3419. [Google Scholar] [CrossRef] [PubMed]
- Seeman, P. Dopamine D2 receptors as treatment targets in schizophrenia. Clin. Schizophr. Relat. Psychoses 2010, 4, 56–73. [Google Scholar] [CrossRef] [PubMed]
- Baronio, D.; Gonchoroski, T.; Castro, K.; Zanatta, G.; Gottfried, C.; Riesgo, R. Histaminergic system in brain disorders: Lessons from the translational approach and future perspectives. Ann. Gen. Psychiatry 2014, 13, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamilton, P.J.; Campbell, N.G.; Sharma, S.; Erreger, K.; Herborg Hansen, F.; Saunders, C.; Belovich, A.N.; Consortium, N.A.A.S.; Sahai, M.A.; Cook, E.H.; et al. De novo mutation in the dopamine transporter gene associates dopamine dysfunction with autism spectrum disorder. Mol. Psychiatry 2013, 18, 1315–1323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berridge, K.C.; Aldridge, J.W.; Houchard, K.R.; Zhuang, X. Sequential super-stereotypy of an instinctive fixed action pattern in hyper-dopaminergic mutant mice: A model of obsessive compulsive disorder and Tourette’s. BMC Biol. 2005, 3, 4. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Kim, H.; Kim, J.E.; Park, J.Y.; Choi, J.; Lee, J.E.; Lee, E.H.; Han, P.L. Excessive D1 Dopamine Receptor Activation in the Dorsal Striatum Promotes Autistic-Like Behaviors. Mol. Neurobiol. 2018, 55, 5658–5671. [Google Scholar] [CrossRef]
- Wright, C.; Shin, J.H.; Rajpurohit, A.; Deep-Soboslay, A.; Collado-Torres, L.; Brandon, N.J.; Hyde, T.M.; Kleinman, J.E.; Jaffe, A.E.; Cross, A.J.; et al. Altered expression of histamine signaling genes in autism spectrum disorder. Transl. Psychiatry 2017, 7, e1126. [Google Scholar] [CrossRef] [Green Version]
- Gillberg, C.; Billstedt, E. Autism and Asperger syndrome: Coexistence with other clinical disorders. Acta Psychiatr. Scand. 2000, 102, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Clarke, R.A.; Lee, S.; Eapen, V. Pathogenetic model for Tourette syndrome delineates overlap with related neurodevelopmental disorders including Autism. Transl. Psychiatry 2012, 2, e158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez, T.V.; Sanders, S.J.; Yurkiewicz, I.R.; Ercan-Sencicek, A.G.; Kim, Y.S.; Fishman, D.O.; Raubeson, M.J.; Song, Y.; Yasuno, K.; Ho, W.S.; et al. Rare copy number variants in tourette syndrome disrupt genes in histaminergic pathways and overlap with autism. Biol. Psychiatry 2012, 71, 392–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paschou, P.; Fernandez, T.V.; Sharp, F.; Heiman, G.A.; Hoekstra, P.J. Genetic susceptibility and neurotransmitters in Tourette syndrome. Int. Rev. Neurobiol. 2013, 112, 155–177. [Google Scholar]
- Rapanelli, M. The magnificent two: Histamine and the H3 receptor as key modulators of striatal circuitry. Prog. Neuropsychopharmacol. Biol. Psychiatry 2017, 73, 36–40. [Google Scholar] [CrossRef] [PubMed]
- Rapanelli, M.; Frick, L.R.; Horn, K.D.; Schwarcz, R.C.; Pogorelov, V.; Nairn, A.C.; Pittenger, C. The Histamine H3 Receptor Differentially Modulates Mitogen-activated Protein Kinase (MAPK) and Akt Signaling in Striatonigral and Striatopallidal Neurons. J. Biol. Chem. 2016, 291, 21042–21052. [Google Scholar] [CrossRef] [Green Version]
- Rapanelli, M.; Pittenger, C. Histamine and histamine receptors in Tourette syndrome and other neuropsychiatric conditions. Neuropharmacology 2016, 106 (Suppl. C), 85–90. [Google Scholar] [CrossRef] [PubMed]
- Eissa, N.; Sadeq, A.; Sasse, A.; Sadek, B. Role of Neuroinflammation in Autism Spectrum Disorder and the Emergence of Brain Histaminergic System. Lessons Also for BPSD? Front. Pharmacol. 2020, 11, 886. [Google Scholar] [CrossRef]
- Frick, L.; Rapanelli, M.; Abbasi, E.; Ohtsu, H.; Pittenger, C. Histamine regulation of microglia: Gene-environment interaction in the regulation of central nervous system inflammation. Brain Behav. Immun. 2016, 57, 326–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ercan-Sencicek, A.G.; Stillman, A.A.; Ghosh, A.K.; Bilguvar, K.; O’Roak, B.J.; Mason, C.E.; Abbott, T.; Gupta, A.; King, R.A.; Pauls, D.L.; et al. L-histidine decarboxylase and Tourette’s syndrome. N. Engl. J. Med. 2010, 362, 1901–1908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldan, L.C.; Williams, K.A.; Gallezot, J.D.; Pogorelov, V.; Rapanelli, M.; Crowley, M.; Anderson, G.M.; Loring, E.; Gorczyca, R.; Billingslea, E.; et al. Histidine decarboxylase deficiency causes tourette syndrome: Parallel findings in humans and mice. Neuron 2014, 81, 77–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wingen, K.; Stark, H. Scaffold variations in amine warhead of histamine H (3) receptor antagonists. Drug Discov. Today Technol. 2013, 10, e483–e489. [Google Scholar] [CrossRef] [PubMed]
- Ghamari, N.; Zarei, O.; Arias-Montano, J.A.; Reiner, D.; Dastmalchi, S.; Stark, H.; Hamzeh-Mivehroud, M. Histamine H3 receptor antagonists/inverse agonists: Where do they go? Pharmacol. Ther. 2019, 200, 69–84. [Google Scholar] [CrossRef]
- Pilla, M.; Perachon, S.; Sautel, F.; Garrido, F.; Mann, A.; Wermuth, C.G.; Schwartz, J.C.; Everitt, B.J.; Sokoloff, P. Selective inhibition of cocaine-seeking behaviour by a partial dopamine D3 receptor agonist. Nature 1999, 400, 371–375. [Google Scholar] [CrossRef]
- Hsu, A.; Togasaki, D.M.; Bezard, E.; Sokoloff, P.; Langston, J.W.; Di Monte, D.A.; Quik, M. Effect of the D3 dopamine receptor partial agonist BP897 [N-[4-(4-(2-methoxyphenyl) piperazinyl) butyl]-2-naphthamide] on L-3,4-dihydroxyphenylalanine-induced dyskinesias and parkinsonism in squirrel monkeys. J. Pharm. Exp. Ther. 2004, 311, 770–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Coburg, Y.; Kottke, T.; Weizel, L.; Ligneau, X.; Stark, H. Potential utility of histamine H3 receptor antagonist pharmacophore in antipsychotics. Bioorg. Med. Chem. Lett. 2009, 19, 538–542. [Google Scholar] [CrossRef] [PubMed]
- Sander, K.; Kottke, T.; Weizel, L.; Stark, H. Kojic acid derivatives as histamine H (3) receptor ligands. Chem. Pharm. Bull. 2010, 58, 1353–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellman, G.L.; Courtney, K.D.; Andres, V., Jr.; Feather-Stone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharm. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Amodeo, D.A.; Jones, J.H.; Sweeney, J.A.; Ragozzino, M.E. Differences in BTBR T+ tf/J and C57BL/6J mice on probabilistic reversal learning and stereotyped behaviors. Behav. Brain Res. 2012, 227, 64–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McFarlane, H.G.; Kusek, G.K.; Yang, M.; Phoenix, J.L.; Bolivar, V.J.; Crawley, J.N. Autism-like behavioral phenotypes in BTBR T+tf/J mice. Genes Brain Behav. 2008, 7, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Moy, S.S.; Nadler, J.J.; Poe, M.D.; Nonneman, R.J.; Young, N.B.; Koller, B.H.; Crawley, J.N.; Duncan, G.E.; Bodfish, J.W. Development of a mouse test for repetitive, restricted behaviors: Relevance to autism. Behav. Brain Res. 2008, 188, 178–194. [Google Scholar] [CrossRef] [Green Version]
- Sadek, B.; Saad, A.; Subramanian, D.; Shafiullah, M.; Lazewska, D.; Kiec-Kononowiczc, K. Anticonvulsant and procognitive properties of the non-imidazole histamine H3 receptor antagonist DL77 in male adult rats. Neuropharmacology 2016, 106, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Yui, K.; Kawasaki, Y.; Yamada, H.; Ogawa, S. Oxidative Stress and Nitric Oxide in Autism Spectrum Disorder and Other Neuropsychiatric Disorders. CNS Neurol. Disord. Drug Targets 2016, 15, 587–596. [Google Scholar] [CrossRef]
- Nadeem, A.; Ahmad, S.F.; Al-Harbi, N.O.; Attia, S.M.; Alshammari, M.A.; Alzahrani, K.S.; Bakheet, S.A. Increased oxidative stress in the cerebellum and peripheral immune cells leads to exaggerated autism-like repetitive behavior due to deficiency of antioxidant response in BTBR T + tf/J mice. Prog. Neuropsychopharmacol. Biol. Psychiatry 2019, 89, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Squillace, M.; Dodero, L.; Federici, M.; Migliarini, S.; Errico, F.; Napolitano, F.; Krashia, P.; Di Maio, A.; Galbusera, A.; Bifone, A.; et al. Dysfunctional dopaminergic neurotransmission in asocial BTBR mice. Transl. Psychiatry 2014, 4, e427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marotta, R.; Risoleo, M.C.; Messina, G.; Parisi, L.; Carotenuto, M.; Vetri, L.; Roccella, M. The Neurochemistry of Autism. Brain Sci. 2020, 10, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baronio, D.; Castro, K.; Gonchoroski, T.; De Melo, G.M.; Nunes, G.D.; Bambini-Junior, V.; Gottfried, C.; Riesgo, R. Effects of an H3R antagonist on the animal model of autism induced by prenatal exposure to valproic acid. PLoS ONE 2015, 10, e0116363. [Google Scholar] [CrossRef] [Green Version]
- Karagiannidis, I.; Dehning, S.; Sandor, P.; Tarnok, Z.; Rizzo, R.; Wolanczyk, T.; Madruga-Garrido, M.; Hebebrand, J.; Nothen, M.M.; Lehmkuhl, G.; et al. Support of the histaminergic hypothesis in Tourette syndrome: Association of the histamine decarboxylase gene in a large sample of families. J. Med. Genet. 2013, 50, 760–764. [Google Scholar] [CrossRef] [Green Version]
- Rapanelli, M.; Frick, L.; Pogorelov, V.; Ohtsu, H.; Bito, H.; Pittenger, C. Histamine H3R receptor activation in the dorsal striatum triggers stereotypies in a mouse model of tic disorders. Transl. Psychiatry 2017, 7, e1013. [Google Scholar] [CrossRef] [PubMed]
- Schwartzer, J.J.; Careaga, M.; Onore, C.E.; Rushakoff, J.A.; Berman, R.F.; Ashwood, P. Maternal immune activation and strain specific interactions in the development of autism-like behaviors in mice. Transl. Psychiatry 2013, 3, e240. [Google Scholar] [CrossRef] [Green Version]
- Burns, A.; Irvine, M.; Woodcock, K. Self-Focused Attention and Depressive Symptoms in Adults with Autistic Spectrum Disorder (ASD). J. Autism Dev. Disord. 2019, 49, 692–703. [Google Scholar] [CrossRef]
- Berlin, M.; Boyce, C.W.; Ruiz Mde, L. Histamine H3 receptor as a drug discovery target. J. Med. Chem. 2011, 54, 26–53. [Google Scholar] [CrossRef] [PubMed]
- Parmentier, R.; Anaclet, C.; Guhennec, C.; Brousseau, E.; Bricout, D.; Giboulot, T.; Bozyczko-Coyne, D.; Spiegel, K.; Ohtsu, H.; Williams, M.; et al. The brain H3-receptor as a novel therapeutic target for vigilance and sleep-wake disorders. Biochem. Pharm. 2007, 73, 1157–1171. [Google Scholar] [CrossRef] [Green Version]
- Khan, N.; Saad, A.; Nurulain, S.M.; Darras, F.H.; Decker, M.; Sadek, B. The dual-acting H3 receptor antagonist and AChE inhibitor UW-MD-71 dose-dependently enhances memory retrieval and reverses dizocilpine-induced memory impairment in rats. Behav. Brain Res. 2016, 297, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Panula, P.; Chazot, P.L.; Cowart, M.; Gutzmer, R.; Leurs, R.; Liu, W.L.; Stark, H.; Thurmond, R.L.; Haas, H.L. International Union of Basic and Clinical Pharmacology. XCVIII. Histamine Receptors. Pharm. Rev. 2015, 67, 601–655. [Google Scholar] [CrossRef] [Green Version]
- Panula, P.; Rinne, J.; Kuokkanen, K.; Eriksson, K.S.; Sallmen, T.; Kalimo, H.; Relja, M. Neuronal histamine deficit in Alzheimer’s disease. Neuroscience 1998, 82, 993–997. [Google Scholar] [CrossRef]
- Sadek, B.; Saad, A.; Sadeq, A.; Jalal, F.; Stark, H. Histamine H3 receptor as a potential target for cognitive symptoms in neuropsychiatric diseases. Behav. Brain Res. 2016, 312, 415–430. [Google Scholar] [CrossRef] [PubMed]
- Sadek, B.; Stark, H. Cherry-picked ligands at histamine receptor subtypes. Neuropharmacology 2016, 106, 56–73. [Google Scholar] [CrossRef] [PubMed]
- Eissa, N.; Jayaprakash, P.; Azimullah, S.; Ojha, S.K.; Al-Houqani, M.; Jalal, F.Y.; Lazewska, D.; Kiec-Kononowicz, K.; Sadek, B. The histamine H3R antagonist DL77 attenuates autistic behaviors in a prenatal valproic acid-induced mouse model of autism. Sci. Rep. 2018, 8, 13077. [Google Scholar] [CrossRef]
- Vestlund, J.; Kalafateli, A.L.; Studer, E.; Westberg, L.; Jerlhag, E. Neuromedin U induces self-grooming in socially-stimulated mice. Neuropharmacology 2020, 162, 107818. [Google Scholar] [CrossRef]
- Rapanelli, M.; Frick, L.; Bito, H.; Pittenger, C. Histamine modulation of the basal ganglia circuitry in the development of pathological grooming. Proc. Natl. Acad. Sci. USA 2017, 114, 6599–6604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, J.; Kim, E. Suppression of NMDA receptor function in mice prenatally exposed to valproic acid improves social deficits and repetitive behaviors. Front. Mol. Neurosci. 2015, 8, 17. [Google Scholar] [CrossRef] [Green Version]
- Kalueff, A.V.; Stewart, A.M.; Song, C.; Berridge, K.C.; Graybiel, A.M.; Fentress, J.C. Neurobiology of rodent self-grooming and its value for translational neuroscience. Nat. Rev. Neurosci. 2016, 17, 45–59. [Google Scholar] [CrossRef] [Green Version]
- Kas, M.J.; Glennon, J.C.; Buitelaar, J.; Ey, E.; Biemans, B.; Crawley, J.; Ring, R.H.; Lajonchere, C.; Esclassan, F.; Talpos, J.; et al. Assessing behavioural and cognitive domains of autism spectrum disorders in rodents: Current status and future perspectives. Psychopharmacology 2014, 231, 1125–1146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Timberlake, M.A.; Prall, K.; Dwivedi, Y. The recent progress in animal models of depression. Prog. Neuropsychopharmacol. Biol. Psychiatry 2017, 77, 99–109. [Google Scholar] [CrossRef]
- Griebel, G.; Pichat, P.; Pruniaux, M.-P.; Beeské, S.; Lopez-Grancha, M.; Genet, E.; Terranova, J.-P.; Castro, A.; Sánchez, J.A.; Black, M.; et al. SAR110894, a potent histamine H3-receptor antagonist, displays procognitive effects in rodents. Pharmacol. Biochem. Behav. 2012, 102, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Bagewadi, H.G.; Ak, A.K.; Shivaramegowda, R.M. An Experimental Study to Evaluate the Effect of Memantine in Animal Models of Anxiety in Swiss Albino Mice. J. Clin. Diagn. Res. 2015, 9, FF01. [Google Scholar] [CrossRef] [PubMed]
- Orekhova, E.V.; Stroganova, T.A.; Prokofyev, A.O.; Nygren, G.; Gillberg, C.; Elam, M. Sensory gating in young children with autism: Relation to age, IQ, and EEG gamma oscillations. Neurosci. Lett. 2008, 434, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Rubenstein, J.L.; Merzenich, M.M. Model of autism: Increased ratio of excitation/inhibition in key neural systems. Genes Brain Behav. 2003, 2, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Mabunga, D.F.; Gonzales, E.L.; Kim, J.W.; Kim, K.C.; Shin, C.Y. Exploring the Validity of Valproic Acid Animal Model of Autism. Exp. Neurobiol. 2015, 24, 285–300. [Google Scholar] [CrossRef] [Green Version]
- Casanova, M.F.; Buxhoeveden, D.; Gomez, J. Disruption in the inhibitory architecture of the cell minicolumn: Implications for autism. Neuroscientist 2003, 9, 496–507. [Google Scholar] [CrossRef]
- Mohammadi, M.R.; Mohammadzadeh, S.; Akhondzadeh, S. Memantine versus Methylphenidate in Children and Adolescents with Attention Deficit Hyperactivity Disorder: A Double-Blind, Randomized Clinical Trial. Iran. J. Psychiatry 2015, 10, 106–114. [Google Scholar]
- Meyza, K.Z.; Defensor, E.B.; Jensen, A.L.; Corley, M.J.; Pearson, B.L.; Pobbe, R.L.; Bolivar, V.J.; Blanchard, D.C.; Blanchard, R.J. The BTBR T+ tf/J mouse model for autism spectrum disorders-in search of biomarkers. Behav. Brain Res. 2013, 251, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Lucchina, L.; Depino, A.M. Altered Peripheral and Central Inflammatory Responses in a Mouse Model of Autism. Autism Res. 2014, 7, 273–289. [Google Scholar] [CrossRef]
- DeLorey, T.M.; Sahbaie, P.; Hashemi, E.; Homanics, G.E.; Clark, J.D. Gabrb3 gene deficient mice exhibit impaired social and exploratory behaviors, deficits in non-selective attention and hypoplasia of cerebellar vermal lobules: A potential model of autism spectrum disorder. Behav. Brain Res. 2008, 187, 207–220. [Google Scholar] [CrossRef] [Green Version]
- Eissa, N.; Azimullah, S.; Jayaprakash, P.; Jayaraj, R.L.; Reiner, D.; Ojha, S.K.; Beiram, R.; Stark, H.; Lazewska, D.; Kiec-Kononowicz, K.; et al. The dual-active histamine H3 receptor antagonist and acetylcholine esterase inhibitor E100 ameliorates stereotyped repetitive behavior and neuroinflammmation in sodium valproate induced autism in mice. Chem. Biol. Interact. 2019, 312, 108775. [Google Scholar] [CrossRef] [PubMed]
- Eissa, N.; Azimullah, S.; Jayaprakash, P.; Jayaraj, R.L.; Reiner, D.; Ojha, S.K.; Beiram, R.; Stark, H.; Lazewska, D.; Kiec-Kononowicz, K.; et al. The Dual-Active Histamine H3 Receptor Antagonist and Acetylcholine Esterase Inhibitor E100 Alleviates Autistic-Like Behaviors and Oxidative Stress in Valproic Acid Induced Autism in Mice. Int. J. Mol. Sci. 2020, 21, 3996. [Google Scholar] [CrossRef] [PubMed]
- Eissa, N.; Jayaprakash, P.; Stark, H.; Lazewska, D.; Kiec-Kononowicz, K.; Sadek, B. Simultaneous Blockade of Histamine H3 Receptors and Inhibition of Acetylcholine Esterase Alleviate Autistic-Like Behaviors in BTBR T+ tf/J Mouse Model of Autism. Biomolecules 2020, 10, 1251. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, D. Histamine H3 receptors and its antagonism as a novel mechanism for antipsychotic effect: A current preclinical & clinical perspective. Int. J. Health Sci. 2016, 10, 564–575. [Google Scholar]
- Mahmood, D.; Akhtar, M.; Jahan, K.; Goswami, D. Histamine H3 receptor antagonists display antischizophrenic activities in rats treated with MK-801. J. Basic Clin. Physiol. Pharm. 2016, 27, 463–471. [Google Scholar] [CrossRef]
- Mahmood, D.; Khanam, R.; Pillai, K.K.; Akhtar, M. Reversal of oxidative stress by histamine H (3) receptor-ligands in experimental models of schizophrenia. Arzneimittelforschung 2012, 62, 222–229. [Google Scholar] [CrossRef]
- Bastaki, S.M.; Abdulrazzaq, Y.M.; Shafiullah, M.; Wiecek, M.; Kiec-Kononowicz, K.; Sadek, B. Anticonvulsant and reproductive toxicological studies of the imidazole-based histamine H3R antagonist 2-18 in mice. Drug Des. Devel. Ther. 2018, 12, 179–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hackling, A.; Ghosh, R.; Perachon, S.; Mann, A.; Holtje, H.D.; Wermuth, C.G.; Schwartz, J.C.; Sippl, W.; Sokoloff, P.; Stark, H. N-(omega-(4-(2-methoxyphenyl) piperazin-1-yl) alkyl) carboxamides as dopamine D2 and D3 receptor ligands. J. Med. Chem. 2003, 46, 3883–3899. [Google Scholar] [CrossRef]
- Sadek, B.; Kuder, K.; Subramanian, D.; Shafiullah, M.; Stark, H.; Lazewska, D.; Adem, A.; Kiec-Kononowicz, K. Anticonvulsive effect of nonimidazole histamine H3 receptor antagonists. Behav. Pharm. 2014, 25, 245–252. [Google Scholar] [CrossRef]
- Sadek, B.; Schreeb, A.; Schwed, J.S.; Weizel, L.; Stark, H. Drug-likeness approach of 2-aminopyrimidines as histamine H3 receptor ligands. Drug Des. Devel. Ther. 2014, 8, 1499–1513. [Google Scholar] [CrossRef] [PubMed]
- Sadek, B.; Schwed, J.S.; Subramanian, D.; Weizel, L.; Walter, M.; Adem, A.; Stark, H. Non-imidazole histamine H3 receptor ligands incorporating antiepileptic moieties. Eur. J. Med. Chem. 2014, 77, 269–279. [Google Scholar] [CrossRef]
- Angoa-Perez, M.; Kane, M.J.; Briggs, D.I.; Francescutti, D.M.; Kuhn, D.M. Marble burying and nestlet shredding as tests of repetitive, compulsive-like behaviors in mice. J. Vis. Exp. 2013, 82, 50978. [Google Scholar] [CrossRef] [PubMed]
- Thomas, A.; Burant, A.; Bui, N.; Graham, D.; Yuva-Paylor, L.A.; Paylor, R. Marble burying reflects a repetitive and perseverative behavior more than novelty-induced anxiety. Psychopharmacology 2009, 204, 361–373. [Google Scholar] [CrossRef] [Green Version]
- Bahi, A.; Schwed, J.S.; Walter, M.; Stark, H.; Sadek, B. Anxiolytic and antidepressant-like activities of the novel and potent non-imidazole histamine H (3) receptor antagonist ST-1283. Drug Des. Devel. Ther. 2014, 8, 627–637. [Google Scholar]
- Kim, J.W.; Seung, H.; Kwon, K.J.; Ko, M.J.; Lee, E.J.; Oh, H.A.; Choi, C.S.; Kim, K.C.; Gonzales, E.L.; You, J.S.; et al. Subchronic treatment of donepezil rescues impaired social, hyperactive, and stereotypic behavior in valproic acid-induced animal model of autism. PLoS ONE 2014, 9, e104927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amodeo, D.A.; Rivera, E.; Dunn, J.T.; Ragozzino, M.E. M100907 attenuates elevated grooming behavior in the BTBR mouse. Behav. Brain Res. 2016, 313, 67–70. [Google Scholar] [CrossRef]
- Amodeo, D.A.; Pahua, A.E.; Zarate, M.; Taylor, J.A.; Peterson, S.; Posadas, R.; Oliver, B.L.; Amodeo, L.R. Differences in the expression of restricted repetitive behaviors in female and male BTBR T + tf/J mice. Behav. Brain Res. 2019, 372, 112028. [Google Scholar] [CrossRef] [PubMed]
- Steinmetz, A.B.; Stern, S.A.; Kohtz, A.S.; Descalzi, G.; Alberini, C.M. Insulin-Like Growth Factor II Targets the mTOR Pathway to Reverse Autism-Like Phenotypes in Mice. J. Neurosci. 2018, 38, 1015–1029. [Google Scholar] [CrossRef] [PubMed]
- Silverman, J.L.; Tolu, S.S.; Barkan, C.L.; Crawley, J.N. Repetitive self-grooming behavior in the BTBR mouse model of autism is blocked by the mGluR5 antagonist MPEP. Neuropsychopharmacology 2010, 35, 976–989. [Google Scholar] [CrossRef] [PubMed]
- Prut, L.; Belzung, C. The open field as a paradigm to measure the effects of drugs on anxiety-like behaviors: A review. Eur. J. Pharm. 2003, 463, 3–33. [Google Scholar] [CrossRef]
- Kataoka, S.; Takuma, K.; Hara, Y.; Maeda, Y.; Ago, Y.; Matsuda, T. Autism-like behaviours with transient histone hyperacetylation in mice treated prenatally with valproic acid. Int. J. Neuropsychopharmacol. 2013, 16, 91–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ojha, S.; Javed, H.; Azimullah, S.; Abul Khair, S.B.; Haque, M.E. Neuroprotective potential of ferulic acid in the rotenone model of Parkinson’s disease. Drug Des. Devel. Ther. 2015, 9, 5499–5510. [Google Scholar] [PubMed] [Green Version]
- Javed, H.; Azimullah, S.; Abul Khair, S.B.; Ojha, S.; Haque, M.E. Neuroprotective effect of nerolidol against neuroinflammation and oxidative stress induced by rotenone. BMC Neurosci. 2016, 17, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BTBR | ||||||
|---|---|---|---|---|---|---|
| B6 (VEH) | (VEH) | ST-2223 (5 mg/kg) | ST-2223 (5 mg/kg) + RAM (10 mg/kg) | ARP (1 mg/kg) | MEM (5 mg/kg) | |
| MDA | ||||||
| Cerebellum | 29.9 ± 3.3 | 54.0 ± 6.4 ## | 35.2 ± 1.8 ** | 45.0 ± 0.6 $$ | 36.8 ± 1.8 *** | 26.7 ± 3.6 *** |
| Hippocampus | 35.1 ± 3.0 | 53.8 ± 2.9 ## | 38.1 ± 3.1 ** | 40.7 ± 3.3 | 32.9 ± 4.1 *** | 32.8 ± 3.4 *** |
| Prefrontal Cortex | 41.1 ± 7.0 | 58.3 ± 1.8 # | 39.1 ± 3.2 *** | 53.6 ± 1.2 $$ | 43.0 ± 2.5 ** | 40.0 ± 5.7 ** |
| Striatum | 30.8 ± 4.1 | 52.7 ± 3.4 ## | 34.5 ± 5.5 * | 51.4 ± 4.8 $ | 41.2 ± 1.4 * | 32.6 ± 4.1 ** |
| GSH | ||||||
| Cerebellum | 1560 ± 103.5 | 827 ± 93.3 ## | 1108 ± 41.1 * | 1266 ± 70.2 | 1030 ± 36.2 * | 877 ± 55.7 |
| Hippocampus | 2119 ± 425.1 | 1096 ± 16.7 # | 916 ± 4.2 | 643.6 ± 49.9 | 611.5 ± 62.6 *** | 642.6 ± 66.0 *** |
| Prefrontal Cortex | 1776 ± 266.6 | 835 ± 97.4 # | 1212 ± 302.6 * | 1070.4 ± 64.8 $ | 935 ± 98.0 | 719.3 ± 50.0 |
| Striatum | 1128 ± 199.5 | 583 ± 68.8 # | 1007 ± 93.1 * | 735.5 ± 59.9 $ | 1139 ± 74.6 * | 915.5 ± 97.3 * |
| SOD | ||||||
| Cerebellum | 60.6 ± 3.1 | 49.7 ± 1.3 # | 64.3 ± 1.0 *** | 49.2 ± 2.0 $$$ | 56.4 ± 1.1 ** | 64.4 ± 3.1 ** |
| Hippocampus | 66.4 ± 5.4 | 49.5 ± 0.5 # | 48.1 ± 4.2 | 39.5 ± 0.5 $$ | 51.8 ± 2.0 | 41.4 ± 3.4 |
| Prefrontal Cortex | 53.8 ± 2.2 | 45.1 ± 2.8 # | 43.2 ± 3.4 | 42.9 ± 4.2 | 54.0 ± 5.2 ** | 44.9 ± 2.2 |
| Striatum | 68.5 ± 4.7 | 51.8 ± 1.2 # | 51.2 ± 7.7 | 45.0 ± 1.0 | 65.7 ± 7.7 ** | 66.47 ± 4.4 ** |
| CAT | ||||||
| Cerebellum | 156.7 ± 7.29 | 130. 7 ± 3.3 # | 120.6 ± 8.8 | 153.3 ± 5.4 | 159.2 ± 5.1 ** | 132.3 ± 4.5 |
| Hippocampus | 131.7 ± 3.4 | 111.4 ± 4.8 # | 173.0 ± 1.9 * | 100.6 ± 3.6 $$ | 118.3 ± 6.5 | 114.3 ± 11.2 |
| Prefrontal Cortex | 130.3 ± 9.4 | 80.1 ± 14.9 # | 116.8 ± 5.3 * | 70.9± 3.9 $ | 96.2 ± 1.3 | 94.1 ± 4.8 |
| Striatum | 117.7 ± 2.5 | 104.6 ± 2.28 # | 119.1 ± 3.7 * | 76.0 ± 9.3 $$ | 120.5 ± 10.1 * | 108.1 ± 3.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eissa, N.; Venkatachalam, K.; Jayaprakash, P.; Falkenstein, M.; Dubiel, M.; Frank, A.; Reiner-Link, D.; Stark, H.; Sadek, B. The Multi-Targeting Ligand ST-2223 with Histamine H3 Receptor and Dopamine D2/D3 Receptor Antagonist Properties Mitigates Autism-Like Repetitive Behaviors and Brain Oxidative Stress in Mice. Int. J. Mol. Sci. 2021, 22, 1947. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041947
Eissa N, Venkatachalam K, Jayaprakash P, Falkenstein M, Dubiel M, Frank A, Reiner-Link D, Stark H, Sadek B. The Multi-Targeting Ligand ST-2223 with Histamine H3 Receptor and Dopamine D2/D3 Receptor Antagonist Properties Mitigates Autism-Like Repetitive Behaviors and Brain Oxidative Stress in Mice. International Journal of Molecular Sciences. 2021; 22(4):1947. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041947
Chicago/Turabian StyleEissa, Nermin, Karthikkumar Venkatachalam, Petrilla Jayaprakash, Markus Falkenstein, Mariam Dubiel, Annika Frank, David Reiner-Link, Holger Stark, and Bassem Sadek. 2021. "The Multi-Targeting Ligand ST-2223 with Histamine H3 Receptor and Dopamine D2/D3 Receptor Antagonist Properties Mitigates Autism-Like Repetitive Behaviors and Brain Oxidative Stress in Mice" International Journal of Molecular Sciences 22, no. 4: 1947. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041947