
Sex Differences in Photoprotective Responses to 1,25-Dihydroxyvitamin D3 in Mice Are Modulated by the Estrogen Receptor-β

 ,
,
Abstract
:1. Introduction
2. Results
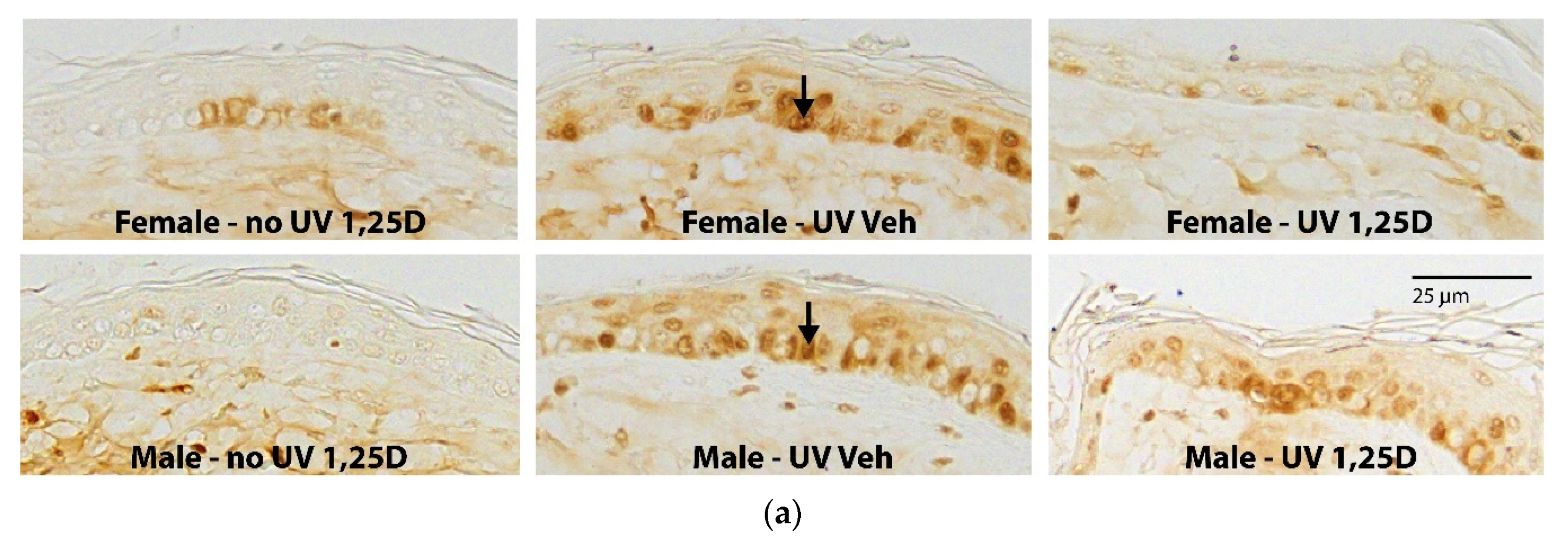
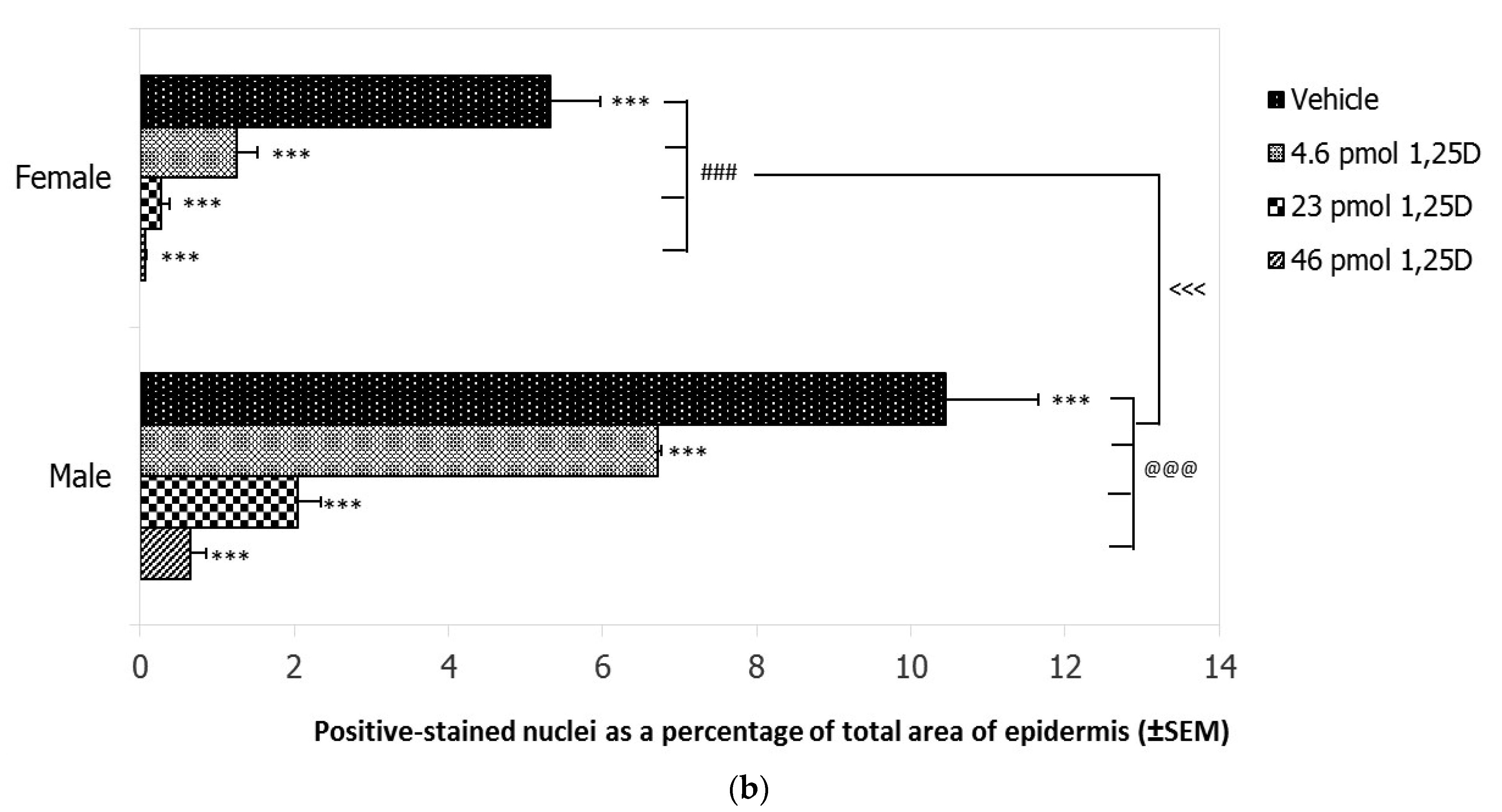
2.1. Effect of 1,25(OH)2D on Photoprotection against CPDs in Female and Male Skh:hr1 Mice
2.2. Effect of ER Blockade on 1,25(OH)2D Photoprotection against CPDs in Female
Skh:hr1 Mice
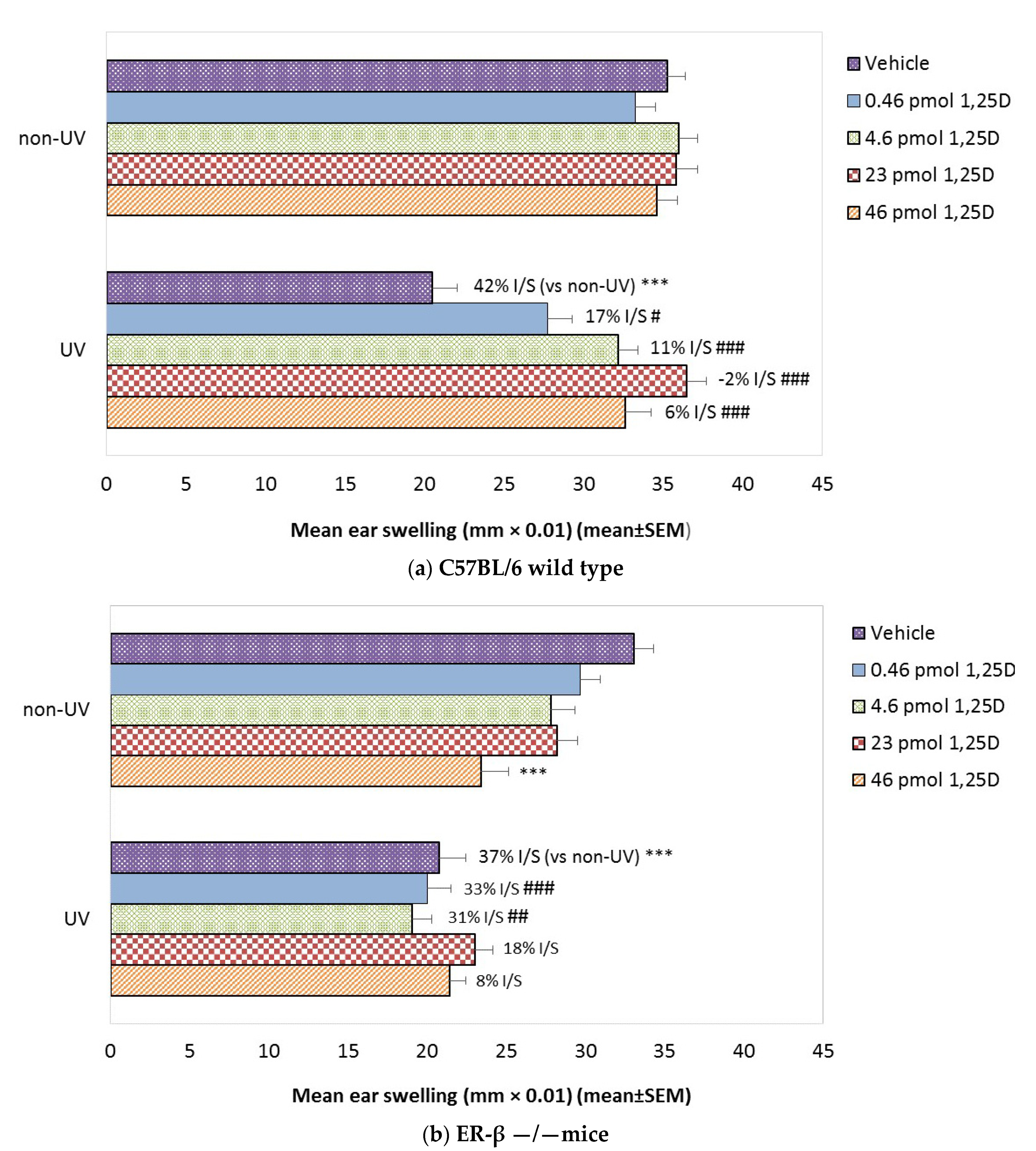
2.3. Effect of 1,25(OH)2D on SSUV-Induced CPDs in ER-β−/−Mice
2.4. Effect of SSUV on Sunburn Cells in Female and Male Skh:hr1 Mice
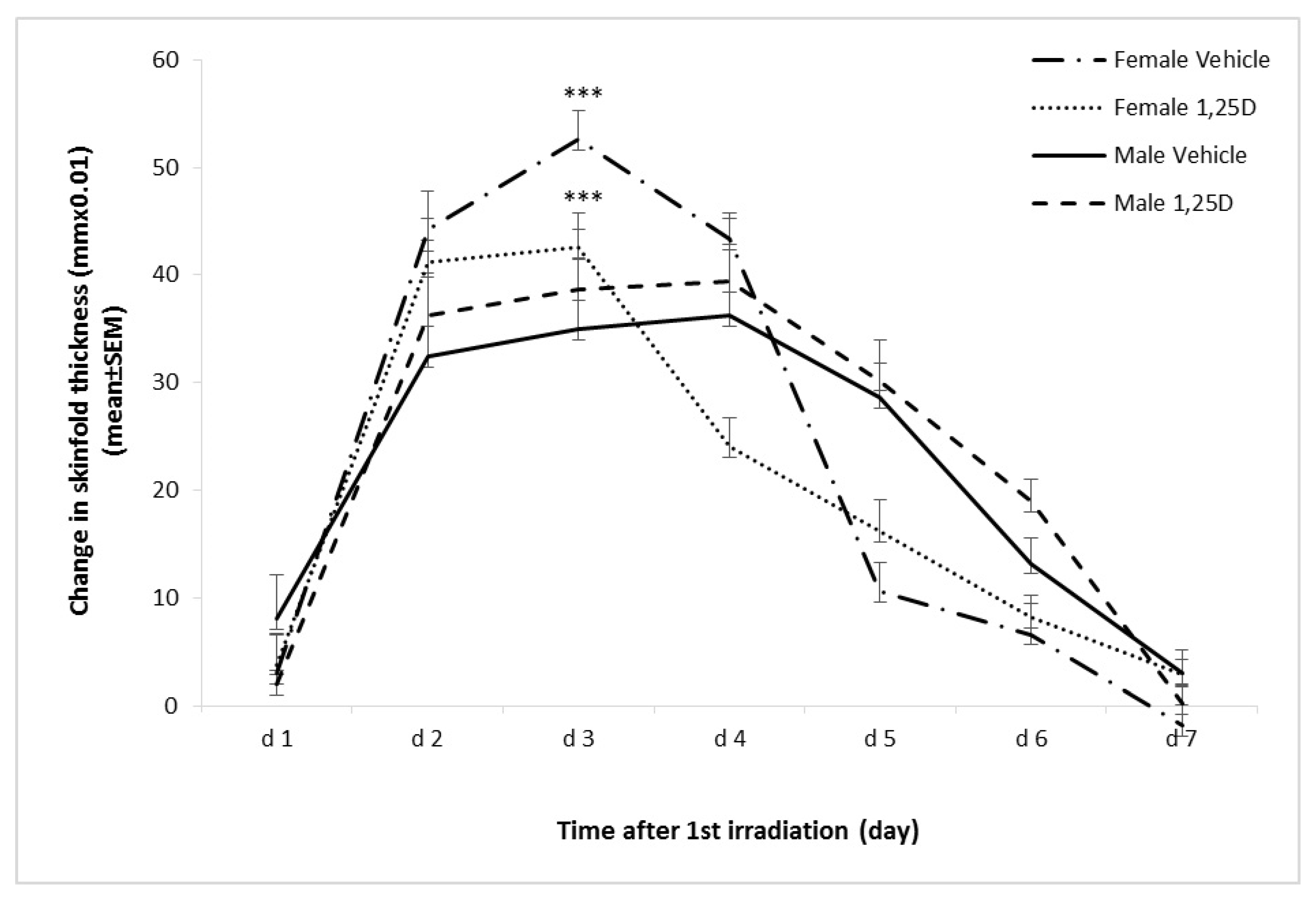
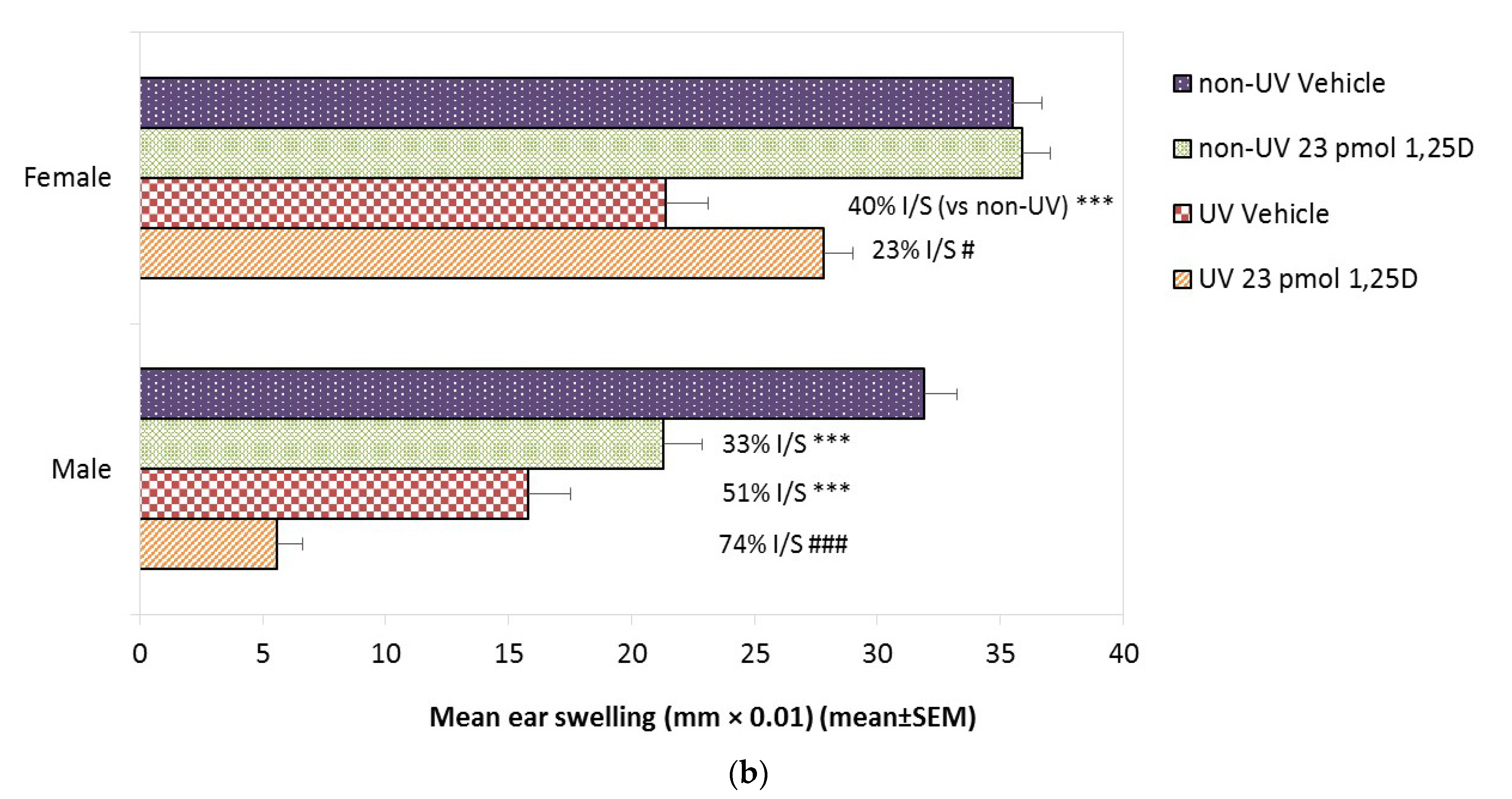
2.5. Effect of 1,25(OH)2D on SSUV-Induced Inflammatory Edema in Female and Male Skh:hr1 Mice
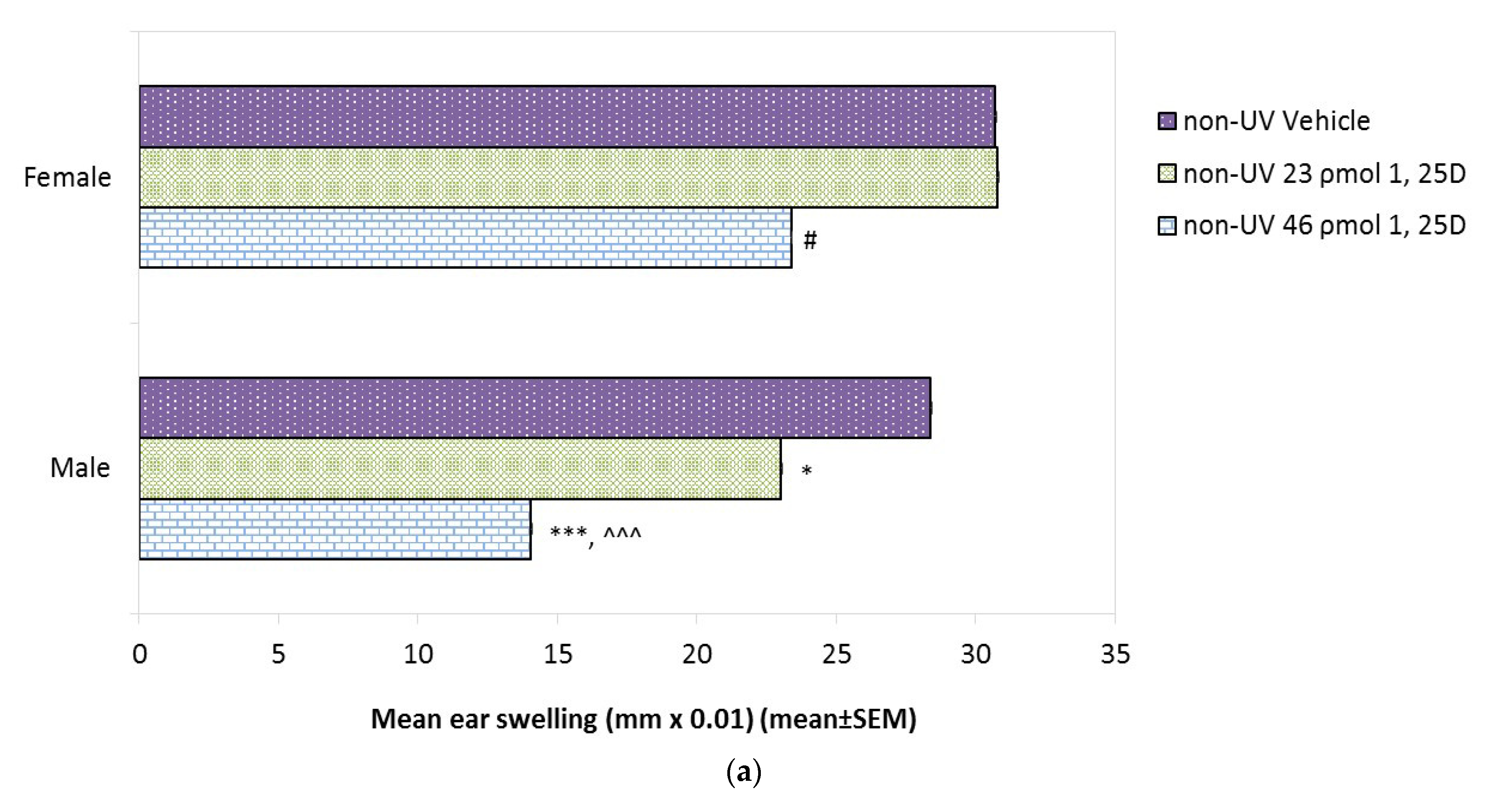
2.6. Effect of 1,25(OH)2D Alone on Basal CHS in Male and Female Skh:hr1 Mice
2.7. Effect of 1,25(OH)2D on Photoimmune Suppression in Female and Male Skh:hr1 Mice
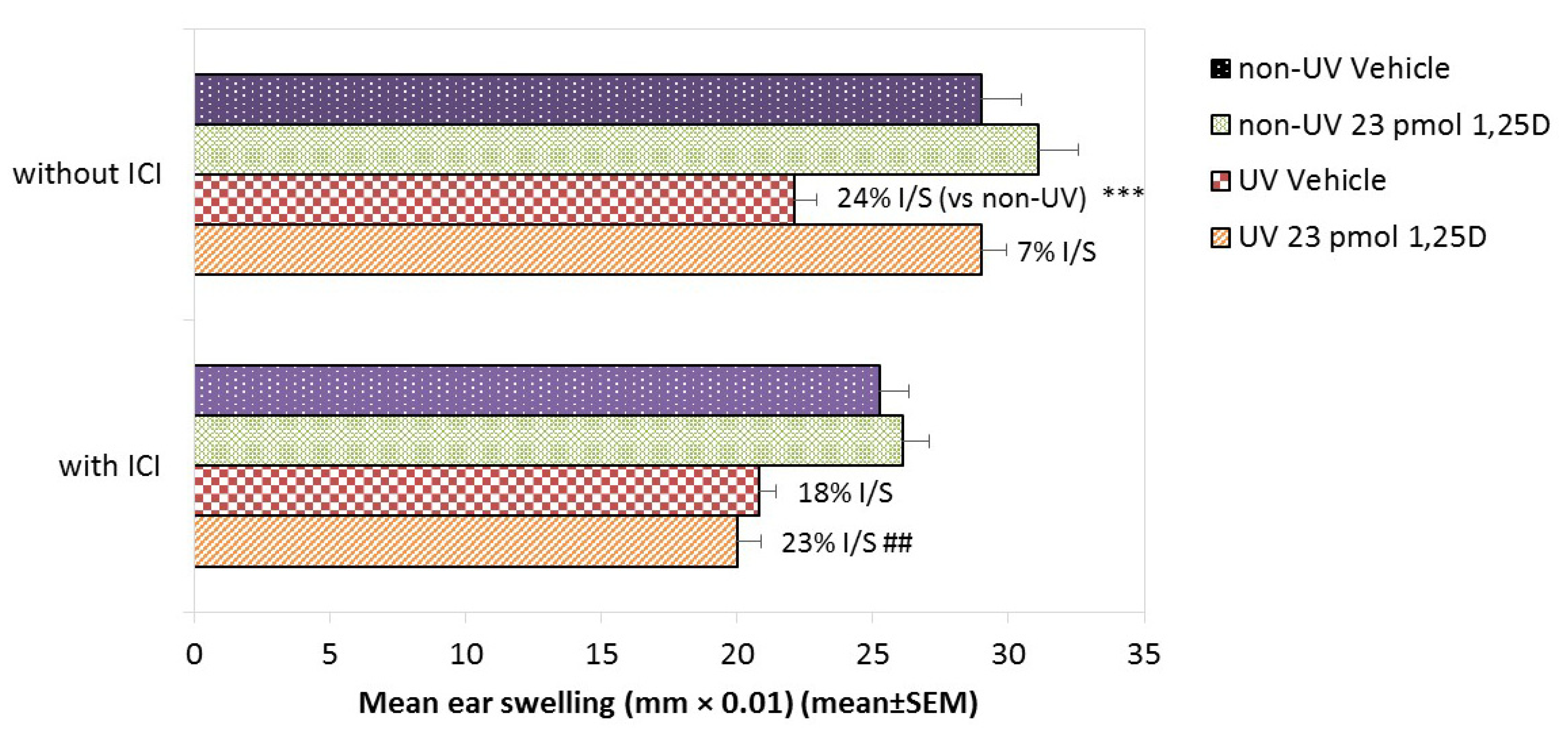
2.8. Effect of ER-β Signalling Blockade on Photoimmune Protection by 1,25(OH)2D in Female Skh:hr1 Mice
2.9. Effect of Genetic Deletion of ER-β on Photoimmune Protection by 1,25(OH)2D
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. UV Irradiation
4.3. Immunohistochemical Detection of CPDs and Image Analysis
4.4. Sunburn Cells
4.5. The Inflammatory Edema Response
4.6. Induction of Contact Hypersensitivity (CHS)
4.7. Topical Treatments
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bouillon, R.; Marcocci, C.; Carmeliet, G.; Bikle, D.; White, J.H.; Dawson-Hughes, B.; Lips, P.; Munns, C.F.; Lazaretti-Castro, M.; Giustina, A.; et al. Skeletal and Extraskeletal Actions of Vitamin D: Current Evidence and Outstanding Questions. Endocr. Rev. 2019, 40, 1109–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plum, L.A.; DeLuca, H.F. Vitamin D, disease and therapeutic opportunities. Nat. Rev. Drug Discov. 2010, 9, 941–955. [Google Scholar] [CrossRef] [PubMed]
- Caraba, A.; Crişan, V.; Romoşan, I.; Mozoş, I.; Murariu, M. Vitamin D Status, Disease Activity, and Endothelial Dysfunction in Early Rheumatoid Arthritis Patients. Dis. Markers 2017, 2017, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Prietl, B.; Treiber, G.; Pieber, T.R.; Amrein, K. Vitamin D and Immune Function. Nutrients 2013, 5, 2502–2521. [Google Scholar] [CrossRef] [PubMed]
- Mason, R.S.; Reichrath, J. Sunlight vitamin D and skin cancer. Anticancer Agents Med. Chem. 2013, 13, 83–97. [Google Scholar] [CrossRef]
- Kim, J.S.; Jung, M.; Yoo, J.; Choi, E.H.; Park, B.C.; Kim, M.H.; Hong, S.P. Protective Effect of Topical Vitamin D3 against Photocarcinogenesis in a Murine Model. Ann. Derm. 2016, 28, 304–313. [Google Scholar] [CrossRef] [Green Version]
- Kripke, M.L.; Cox, P.A.; Alas, L.G.; Yarosh, D.B. Pyrimidine dimers in DNA initiate systemic immunosuppression in UV-irradiated mice. Proc. Natl. Acad. Sci. USA 1992, 89, 7516–7520. [Google Scholar] [CrossRef] [Green Version]
- Nishigori, C.; Yarosh, D.B.; Ullrich, S.E.; Vink, A.A.; Bucana, C.D.; Roza, L.; Kripke, M.L. Evidence that DNA damage triggers interleukin 10 cytokine production in UV-irradiated murine keratinocytes. Proc. Natl. Acad. Sci. USA 1996, 93, 10354–10359. [Google Scholar] [CrossRef] [Green Version]
- Ullrich, S.E. Mechanisms underlying UV-induced immune suppression. Mutat. Res. 2005, 571, 185–205. [Google Scholar] [CrossRef]
- Halliday, G.M. Inflammation, gene mutation and photoimmunosuppression in response to UVR-induced oxidative damage contributes to photocarcinogenesis. Mutat. Res. Mol. Mech. Mutagen. 2005, 571, 107–120. [Google Scholar] [CrossRef]
- Halliday, G.M. Common Links among the Pathways Leading to UV-Induced Immunosuppression. J. Investig. Derm. 2010, 130, 1209–1212. [Google Scholar] [CrossRef]
- Bikle, D.D.; Nemanic, M.K.; Whitney, J.O.; Elias, P.W. Neonatal human foreskin keratinocytes produce 1,25-dihydroxyvitamin D3. Biochem. 1986, 25, 1545–1548. [Google Scholar] [CrossRef]
- Lehmann, B.; Genehr, T.; Knuschke, P.; Meurer, M.; Pietzsch, J. UVB-Induced Conversion of 7-Dehydrocholesterol to 1α,25-Dihydroxyvitamin D3 in an In Vitro Human Skin Equivalent Model. J. Investig. Derm. 2001, 117, 1179–1185. [Google Scholar] [CrossRef] [Green Version]
- Rybchyn, M.S.; De Silva, W.G.M.; Sequeira, V.B.; McCarthy, B.Y.; Dilley, A.V.; Dixon, K.M.; Halliday, G.M.; Mason, R.S. Enhanced Repair of UV-Induced DNA Damage by 1,25-Dihydroxyvitamin D3 in Skin Is Linked to Pathways that Control Cellular Energy. J. Investig. Derm. 2018, 138, 1146–1156. [Google Scholar] [CrossRef] [Green Version]
- De Silva, W.G.M.; Abboud, M.; Yang, C.; Dixon, K.M.; Rybchyn, M.S.; Mason, R.S. Protection from Ultraviolet Damage and Photocarcinogenesis by Vitamin D Compounds. Adv. Exp. Med. Biol. 2020, 1268, 227–253. [Google Scholar] [CrossRef]
- Dixon, K.; Deo, S.; Wong, G.; Slater, M.; Norman, A.; Bishop, J.; Posner, G.; Ishizuka, S.; Halliday, G.; Reeve, V.; et al. Skin cancer prevention: A possible role of 1,25dihydroxyvitamin D3 and its analogs. J. Steroid Biochem. Mol. Biol. 2005, 97, 137–143. [Google Scholar] [CrossRef]
- Gupta, R.; Dixon, K.M.; Deo, S.S.; Holliday, C.J.; Slater, M.; Halliday, G.M.; Reeve, V.E.; Mason, R.S. Photoprotection by 1,25 Dihydroxyvitamin D3 Is Associated with an Increase in p53 and a Decrease in Nitric Oxide Products. J. Investig. Derm. 2007, 127, 707–715. [Google Scholar] [CrossRef] [Green Version]
- Damian, D.L.; Kim, Y.J.; Dixon, K.M.; Halliday, G.M.; Javeri, A.; Mason, R.S. Topical calcitriol protects from UV-induced genetic damage but suppresses cutaneous immunity in humans. Exp. Derm. 2009, 19, e23–e30. [Google Scholar] [CrossRef] [PubMed]
- Gordon-Thomson, C.; Gupta, R.; Tongkao-On, W.; Ryan, A.; Halliday, G.M.; Mason, R.S. 1α,25 Dihydroxyvitamin D3 enhances cellular defences against UV-induced oxidative and other forms of DNA damage in skin. Photochem. Photobiol. Sci. 2012, 11, 1837–1847. [Google Scholar] [CrossRef] [PubMed]
- Song, E.J.; Gordon-Thomson, C.; Cole, L.; Stern, H.; Halliday, G.M.; Damian, D.L.; Reeve, V.E.; Mason, R.S. 1α,25-Dihydroxyvitamin D3 reduces several types of UV-induced DNA damage and contributes to photoprotection. J. Steroid Biochem. Mol. Biol. 2013, 136, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Sequeira, V.B.; Rybchyn, M.S.; Gordon-Thomson, C.; Tongkao-On, W.; Mizwicki, M.T.; Norman, A.W.; Reeve, V.E.; Halliday, G.M.; Mason, R.S. Opening of Chloride Channels by 1α,25-Dihydroxyvitamin D 3 Contributes to Photoprotection against UVR-Induced Thymine Dimers in Keratinocytes. J. Investig. Derm. 2013, 133, 776–782. [Google Scholar] [CrossRef] [Green Version]
- Chaiprasongsuk, A.; Janjetovic, Z.; Kim, T.-K.; Jarrett, S.G.; D’Orazio, J.A.; Holick, M.F.; Tang, E.K.; Tuckey, R.C.; Panich, U.; Li, W.; et al. Protective effects of novel derivatives of vitamin D3 and lumisterol against UVB-induced damage in human keratinocytes involve activation of Nrf2 and p53 defense mechanisms. Redox Biol. 2019, 24, 101206. [Google Scholar] [CrossRef]
- Dixon, K.; Deo, S.; Norman, A.; Bishop, J.; Halliday, G.; Reeve, V.; Mason, R. In vivo relevance for photoprotection by the vitamin D rapid response pathway. J. Steroid Biochem. Mol. Biol. 2007, 103, 451–456. [Google Scholar] [CrossRef]
- Dixon, K.M.; Norman, A.W.; Sequeira, V.B.; Mohan, R.; Rybchyn, M.S.; Reeve, V.E.; Halliday, G.M.; Mason, R.S. 1α,25(OH)2-Vitamin D and a Nongenomic Vitamin D Analogue Inhibit Ultraviolet Radiation–Induced Skin Carcinogenesis. Cancer Prev. Res. 2011, 4, 1485–1494. [Google Scholar] [CrossRef] [Green Version]
- Wood, A.W.; Chang, R.L.; Huang, M.-T.; Uskokovic, M.; Conney, A.H. 1α,25-Dihydroxyvitamin D3 inhibits phorbol ester-dependent chemical carcinogenesis in mouse skin. Biochem. Biophys. Res. Commun. 1983, 116, 605–611. [Google Scholar] [CrossRef]
- Kensler, T.W.; Dolan, P.M.; Gange, S.J.; Lee, J.K.; Wang, Q.; Posner, G.H. Conceptually new deltanoids (vitamin D analogs) inhibit multistage skin tumorigenesis. Carcinogenesis. 2000, 21, 1341–1345. [Google Scholar] [CrossRef] [PubMed]
- Ellison, T.I.; Smith, M.K.; Gilliam, A.C.; Macdonald, P.N. Inactivation of the Vitamin D Receptor Enhances Susceptibility of Murine Skin to UV-Induced Tumorigenesis. J. Investig. Derm. 2008, 128, 2508–2517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reeve, V.E.; Allanson, M.; Cho, J.L.; Arun, S.J.; Domanski, D. Interdependence between Heme Oxygenase-1 Induction and Estrogen-Receptor-beta Signaling Mediates Photoimmune Protection by UVA Radiation in Mice. J. Investig. Derm. 2009, 129, 2702–2710. [Google Scholar] [CrossRef] [Green Version]
- Reeve, V.E.; Allanson, M.; Domanski, D.; Painter, N. Gender differences in UV-induced inflammation and immunosuppression in mice reveal male unresponsiveness to UVA radiation. Photochem. Photobiol. Sci. 2012, 11, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Damian, D.L.; Patterson, C.R.S.; Stapelberg, M.; Park, J.; Barnetson, R.S.C.; Halliday, G.M. UV Radiation-Induced Immunosuppression Is Greater in Men and Prevented by Topical Nicotinamide. J. Investig. Derm. 2008, 128, 447–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas-Ahner, J.M.; Wulff, B.C.; Tober, K.L.; Kusewitt, D.F.; Riggenbach, J.A.; Oberyszyn, T.M. Gender Differences in UVB-Induced Skin Carcinogenesis, Inflammation, and DNA Damage. Cancer Res. 2007, 67, 3468–3474. [Google Scholar] [CrossRef] [Green Version]
- Foote, J.A.; Harris, R.B.; Giuliano, A.R.; Roe, D.J.; Moon, T.E.; Cartmel, B.; Alberts, D.S. Predictors for cutaneous basal- and squamous-cell carcinoma among actinically damaged adults. Int. J. Cancer 2001, 95, 7–11. [Google Scholar] [CrossRef] [Green Version]
- Rees, J.R.; Zens, M.S.; Celaya, M.O.; Riddle, B.L.; Karagas, M.R.; Peacock, J.L. Survival after squamous cell and basal cell carcinoma of the skin: A retrospective cohort analysis. Int. J. Cancer 2015, 137, 878–884. [Google Scholar] [CrossRef] [PubMed]
- Friedmann, A.C.; Halliday, G.M.; Barnetson, R.S.; Reeve, V.E.; Walker, C.; Patterson, C.R. The topical isoflavonoid NV-07alpha reduces solar-simulated UV-induced suppression of Mantoux reactions in humans. Photochem. Photobiol. 2004, 80, 416–421. [Google Scholar] [CrossRef]
- Widyarini, S.; Spinks, N.; Husband, A.J.; Reeve, V.E. Isoflavonoid compounds from red clover (Trifolium pratense) protect from inflammation and immune suppression induced by UV radiation. Photochem. Photobiol. 2001, 74, 465–470. [Google Scholar] [CrossRef]
- Widyarini, S.; Domanski, D.; Painter, N.; Reeve, V.E. Estrogen receptor signaling protects against immune suppression by UV radiation exposure. Proc. Natl. Acad. Sci. USA 2006, 103, 12837–12842. [Google Scholar] [CrossRef] [Green Version]
- Widyarini, S.; Husband, A.J.; Reeve, V.E. Protective effect of the isoflavonoid equol against hairless mouse skin carcinogenesis induced by UV radiation alone or with a chemical cocarcinogen. Photochem. Photobiol. 2005, 81, 32–37. [Google Scholar] [CrossRef]
- Cho, J.-L.; Allanson, M.; Domanski, D.; Arun, S.J.; Reeve, V.E. Estrogen receptor-beta signaling protects epidermal cytokine expression and immune function from UVB-induced impairment in mice. Photochem. Photobiol. Sci. 2007, 7, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.L.; Allanson, M.; Reeve, V.E. Oestrogen receptor-beta signalling protects against transplanted skin tumour growth in the mouse. Photochem. Photobiol. Sci. Off. J. Eur. Photochem. Assoc. Eur. Soc. Photobiol. 2010, 9, 608–614. [Google Scholar]
- Gilad, L.A.; Tirosh, O.; Schwartz, B. Phytoestrogens regulate transcription and translation of vitamin D receptor in colon cancer cells. J. Endocrinol. 2006, 191, 387–398. [Google Scholar] [CrossRef] [Green Version]
- Reeve, V.E.; Ley, R.D. Animal models of ultraviolet radiation-induced skin cancer. In Cancer Prevention—Cancer Causes; Springer International Publishing: New York, NY, USA, 2004; Volume 3, pp. 177–194. [Google Scholar]
- Gorman, S.; Kuritzky, L.A.; Judge, M.A.; Dixon, K.M.; McGlade, J.P.; Mason, R.S.; Finlay-Jones, J.J.; Hart, P.H. Topically Applied 1,25-Dihydroxyvitamin D3 Enhances the Suppressive Activity of CD4+CD25+ Cells in the Draining Lymph Nodes. J. Immunol. 2007, 179, 6273–6283. [Google Scholar] [CrossRef] [Green Version]
- Applegate, L.A.; Ley, R.D.; Alcalay, J.; Kripke, M.L. Identification of the molecular target for the suppression of contact hypersensitivity by ultraviolet radiation. J. Exp. Med. 1989, 170, 1117–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mancuso, M.; Gallo, D.; Leonardi, S.; Pierdomenico, M.; Pasquali, E.; De Stefano, I.; Rebessi, S.; Tanori, M.; Scambia, G.; Di Majo, V.; et al. Modulation of basal and squamous cell carcinoma by endogenous estrogen in mouse models of skin cancer. Carcinogenesis 2008, 30, 340–347. [Google Scholar] [CrossRef] [Green Version]
- De Haes, P.; Garmyn, M.; Degreef, H.; Vantieghem, K.; Bouillon, R.; Segaert, S. 1,25-dihydroxyvitamin D3 inhibits ultraviolet B-induced apoptosis, Jun kinase activation, and interleukin-6 production in primary human keratinocytes. J. Cell. Biochem. 2003, 89, 663–673. [Google Scholar] [CrossRef] [PubMed]
- Mason, R.; Sequeira, V.; Dixon, K.; Gordon-Thomson, C.; Pobre, K.; Dilley, A.; Mizwicki, M.; Norman, A.; Feldman, D.; Halliday, G.; et al. Photoprotection by 1α,25-dihydroxyvitamin D and analogs: Further studies on mechanisms and implications for UV-damage. J. Steroid Biochem. Mol. Biol. 2010, 121, 164–168. [Google Scholar] [CrossRef] [PubMed]
- D’Ambrosio, D.; Cippitelli, M.; Cocciolo, M.G.; Mazzeo, D.; Di Lucia, P.; Lang, R.; Sinigaglia, F.; Panina-Bordignon, P. Inhibition of IL-12 production by 1,25-dihydroxyvitamin D3. Involvement of NF-kappaB downregulation in transcriptional repression of the p40 gene. J. Clin. Investig. 1998, 101, 252–262. [Google Scholar]
- Bikle, D. Nonclassic Actions of Vitamin D. J. Clin. Endocrinol. Metab. 2009, 94, 26–34. [Google Scholar] [CrossRef] [Green Version]
- Daniel, C.; Sartory, N.A.; Zahn, N.; Radeke, H.H.; Stein, J.M. Immune Modulatory Treatment of Trinitrobenzene Sulfonic Acid Colitis with Calcitriol Is Associated with a Change of a T Helper (Th) 1/Th17 to a Th2 and Regulatory T Cell Profile. J. Pharm. Exp. 2007, 324, 23–33. [Google Scholar] [CrossRef] [Green Version]
- Penna, G.; Adorini, L. 1α,25-Dihydroxyvitamin D3Inhibits Differentiation, Maturation, Activation, and Survival of Dendritic Cells Leading to Impaired Alloreactive T Cell Activation. J. Immunol. 2000, 164, 2405–2411. [Google Scholar] [CrossRef] [Green Version]
- Hart, P.H.; Gorman, S.; Finlay-Jones, J.J. Modulation of the immune system by UV radiation: More than just the effects of vitamin D? Nat. Rev. Immunol. 2011, 11, 584–596. [Google Scholar] [CrossRef]
- Biggs, L.; Yu, C.; Fedoric, B.; Lopez, A.F.; Galli, S.J.; Grimbaldeston, M.A. Evidence that vitamin D3 promotes mast cell–dependent reduction of chronic UVB-induced skin pathology in mice. J. Exp. Med. 2010, 207, 455–463. [Google Scholar] [CrossRef]
- Hiramoto, K.; Tanaka, H.; Yanagihara, N.; Sato, E.F.; Inoue, M. Effect of 17beta-estradiol on immunosuppression induced by ultraviolet B irradiation. Arch. Derm. Res. 2004, 295, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Sequeira, V.B.; Rybchyn, M.S.; Tongkao-On, W.; Gordon-Thomson, C.; Malloy, P.J.; Nemere, I.; Norman, A.W.; Reeve, V.E.; Halliday, G.M.; Feldman, D.; et al. The Role of the Vitamin D Receptor and ERp57 in Photoprotection by 1α,25-Dihydroxyvitamin D3. Mol. Endocrinol. 2012, 26, 574–582. [Google Scholar] [CrossRef] [Green Version]
- Liel, Y.; Shany, S.; Smirnoff, P.; Schwartz, B. Estrogen Increases 1,25-Dihydroxyvitamin D Receptors Expression and Bioresponse in the Rat Duodenal Mucosa. Endocrinology 1999, 140, 280–285. [Google Scholar] [CrossRef]
- Schwartz, B.; Smirnoff, P.; Shany, S.; Liel, Y. Estrogen controls expression and bioresponse of 1,25-dihydroxyvitamin D receptors in the rat colon. Mol. Cell. Biochem. 2000, 203, 87–93. [Google Scholar] [CrossRef]
- Gilad, L.A.; Bresler, T.; Gnainsky, J.; Smirnoff, P.; Schwartz, B. Regulation of vitamin D receptor expression via estrogen-induced activation of the ERK 1/2 signaling pathway in colon and breast cancer cells. J. Endocrinol. 2005, 185, 577–592. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, A.V.; Swami, S.; Feldman, D. The potential therapeutic benefits of vitamin D in the treatment of estrogen receptor positive breast cancer. Steroids 2012, 77, 1107–1112. [Google Scholar] [CrossRef] [Green Version]
- Kallay, E.; Adlercreutz, H.; Farhan, H.; Lechner, D.; Bajna, E.; Gerdenitsch, W.; Campbell, M.; Cross, H.S. Phytoestrogens Regulate Vitamin D Metabolism in the Mouse Colon: Relevance for Colon Tumor Prevention and Therapy. J. Nutr. 2002, 132, 3490S–3493S. [Google Scholar] [CrossRef] [Green Version]
- Gorman, S.; Scott, N.M.; Tan, D.H.W.; Weeden, C.E.; Tuckey, R.C.; Bisley, J.L.; Grimbaldeston, M.A.; Hart, P.H. Acute Erythemal Ultraviolet Radiation Causes Systemic Immunosuppression in the Absence of Increased 25-Hydroxyvitamin D3 Levels in Male Mice. PLoS ONE 2012, 7, e46006. [Google Scholar] [CrossRef] [Green Version]
- Correale, J.; Ysrraelit, M.C.; Gaitán, M.I. Response to Comment on “Gender Differences in 1,25 Dihydroxyvitamin D3 Immunomodulatory Effects in Multiple Sclerosis Patients and Healthy Subjects”. J. Immunol. 2011, 186, 648. [Google Scholar] [CrossRef] [Green Version]
- Reeve, V.E.; Domanski, D.; Slater, M. Radiation Sources Providing Increased UVA/UVB Ratios Induce Photoprotection Dependent on the UVA Dose in Hairless Mice. Photochem. Photobiol. 2006, 82, 406–411. [Google Scholar] [CrossRef] [PubMed]
- Ibuki, Y.; Allanson, M.; Dixon, K.M.; Reeve, V.E. Radiation Sources Providing Increased UVA/UVB Ratios Attenuate the Apoptotic Effects of the UVB Waveband UVA-Dose-Dependently in Hairless Mouse Skin. J. Investig. Dermatol. 2007, 127, 2236–2244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, S.E. Mechanisms of Photoprotection by 20-Hydroxyvitamin D3, A Naturally Occurring 1alpha25-Dihydroxyvitamin D3 Analogue; University of Sydney: Sydney, Australia, 2014. [Google Scholar]
- Douki, T.; Cadet, J. Individual Determination of the Yield of the Main UV-Induced Dimeric Pyrimidine Photoproducts in DNA Suggests a High Mutagenicity of CC Photolesions. Biochemistry 2001, 40, 2495–2501. [Google Scholar] [CrossRef]
- Okada, T.; Albarenque, S.M.; Yasoshima, A.; Malcotti, V.; Katayama, K.; Uetsuka, K.; Nakayama, H.; Doi, K. Sunburn reaction in the dorsal skin of hypotrichotic WBN/ILA-Ht rats. Histol. Histopathol. 2003, 18, 753–760. [Google Scholar] [PubMed]
- Reeve, V.E. Ultraviolet radiation and the contact hypersensitivity reaction in mice. Methods 2002, 28, 20–24. [Google Scholar] [CrossRef]
- Ashford, J.R.; Colquhoun, D. Lectures on Biostatistics: An Introduction to Statistics with Applications in Biology and Medicine. J. R. Stat. Soc. Ser. A 1972, 135, 606. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| [1,25(OH)2D Dose] pmol/cm2 | Protection % | Protection % | ||
|---|---|---|---|---|
| A | Female Skh:hr1 Mice | versus | Male Skh:hr1 Mice | |
| 0 (vehicle) | 100 ± 13.2 | 198.1± 22.6 | ||
| 4.6 | 23.6± 5.1 | 77 ** | 126.4 ± 0.9 | 36 ** |
| 23 | 5.1 ± 1.9 | 95 | 39.7 ± 5.7 | 80 |
| 46 | 1.3 ± 0.2 | 99 | 12.3 ± 4.0 | 94 |
| B | Female Skh:hr1 Mice —ICI 182,780 versus | +ICI 182,780 | ||
| 0 (vehicle) | 100 ± 20.0 | 90.0 ± 5.0 | ||
| 4.6 | 18.5 ± 0.4 | 81 | 24.0 ± 3.5 | 73 |
| 23 | 16.3 ± 1.5 | 84 | 23.5 ± 2.0 | 74 |
| 46 | 5.9 ± 0.8 | 94 | 23.5 ± 3.0 | 74 |
| C | Female C57BL/6 Mice | versus | Female ERβ−/−Mice | |
| 0 (vehicle) | 100 ± 8.6 | 125.7 ± 14.3 | ||
| 23 | 34.3 ± 2.9 | 66 * | 68.6 ± 8.6 | 45 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tongkao-on, W.; Yang, C.; McCarthy, B.Y.; De Silva, W.G.M.; Rybchyn, M.S.; Gordon-Thomson, C.; Dixon, K.M.; Halliday, G.M.; Reeve, V.E.; Mason, R.S. Sex Differences in Photoprotective Responses to 1,25-Dihydroxyvitamin D3 in Mice Are Modulated by the Estrogen Receptor-β. Int. J. Mol. Sci. 2021, 22, 1962. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041962
Tongkao-on W, Yang C, McCarthy BY, De Silva WGM, Rybchyn MS, Gordon-Thomson C, Dixon KM, Halliday GM, Reeve VE, Mason RS. Sex Differences in Photoprotective Responses to 1,25-Dihydroxyvitamin D3 in Mice Are Modulated by the Estrogen Receptor-β. International Journal of Molecular Sciences. 2021; 22(4):1962. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041962
Chicago/Turabian StyleTongkao-on, Wannit, Chen Yang, Bianca Y. McCarthy, Warusavithana G. Manori De Silva, Mark S. Rybchyn, Clare Gordon-Thomson, Katie M. Dixon, Gary M. Halliday, Vivienne E. Reeve, and Rebecca S. Mason. 2021. "Sex Differences in Photoprotective Responses to 1,25-Dihydroxyvitamin D3 in Mice Are Modulated by the Estrogen Receptor-β" International Journal of Molecular Sciences 22, no. 4: 1962. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22041962