
Development of a Specific Monoclonal Antibody to Detect Male Cells Expressing the RPS4Y1 Protein


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. RPS4Y1 Is a Marker of Male Cells
2.2. Selection of RPS4Y1-Antigen Peptides
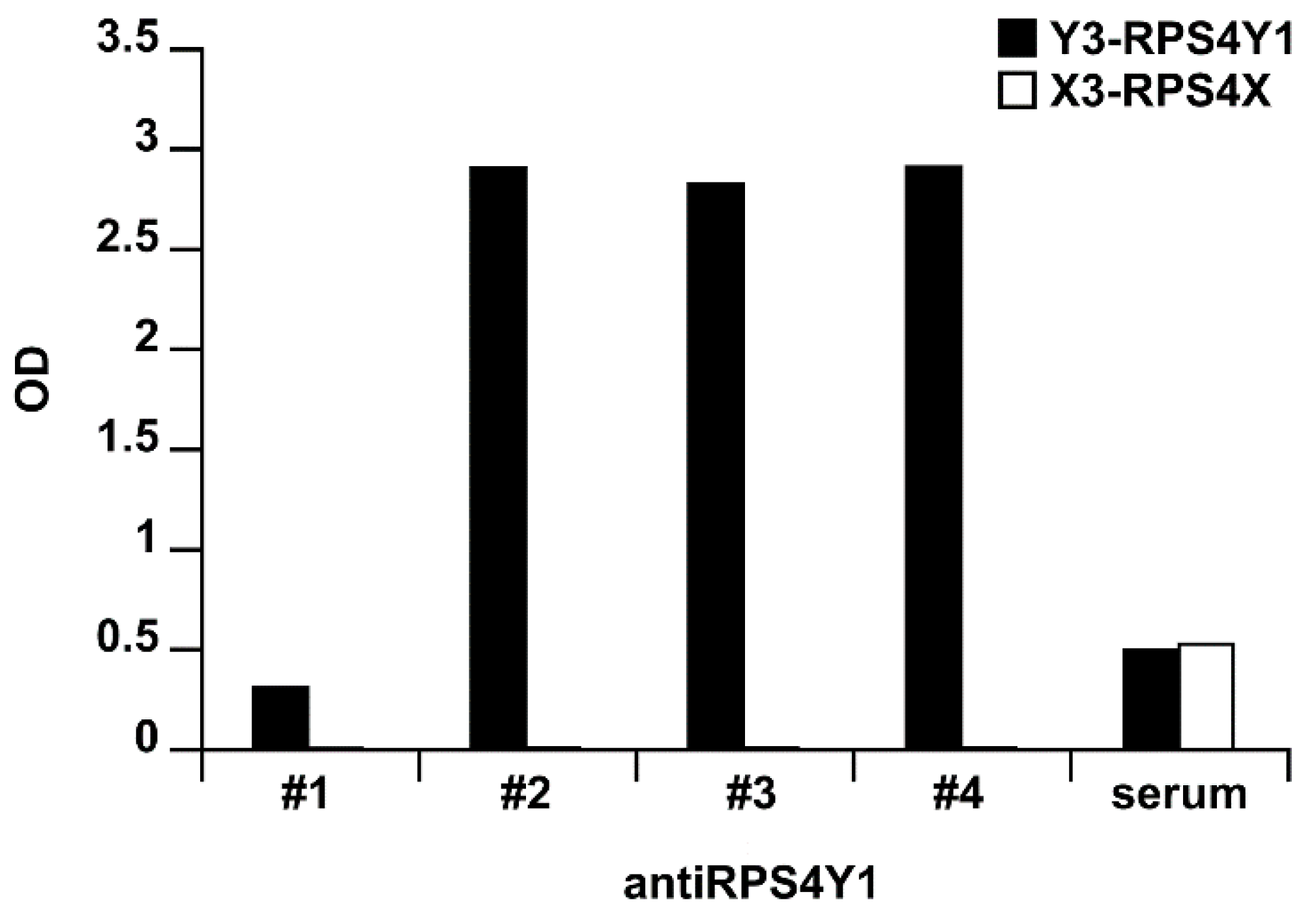
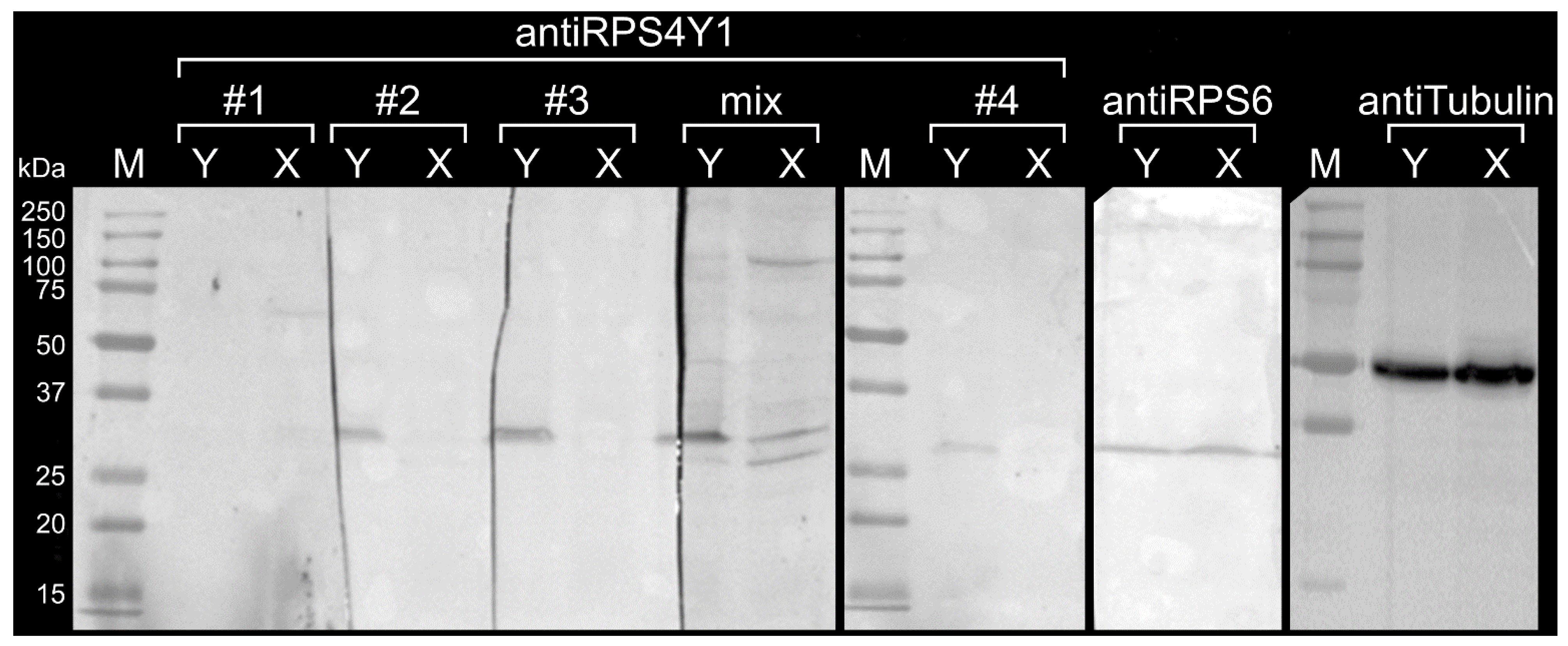
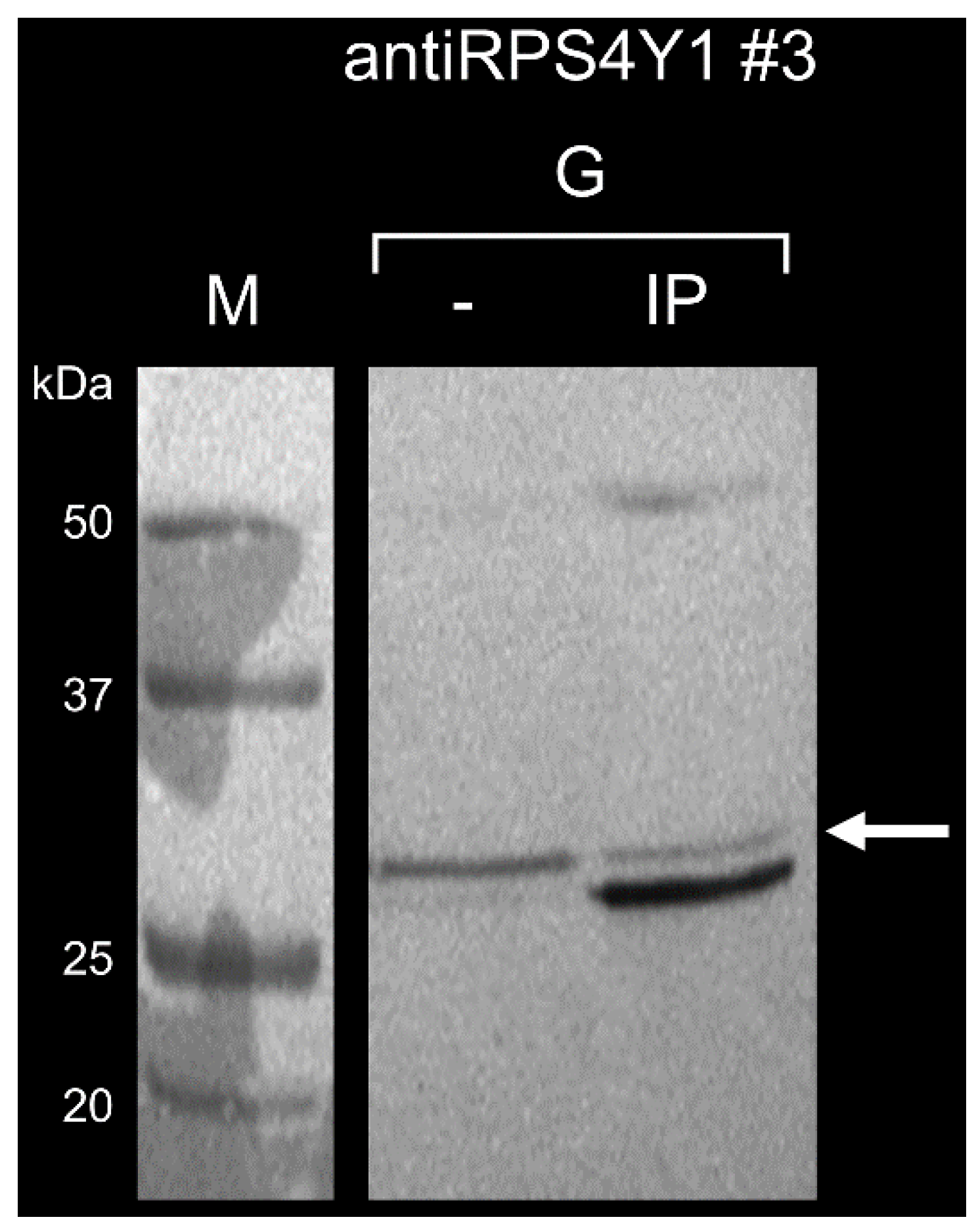
2.3. AntiRPS4Y1 Antibodies Are Specific for a RPS4Y1-Antigen Peptide and the RPS4Y1 Protein
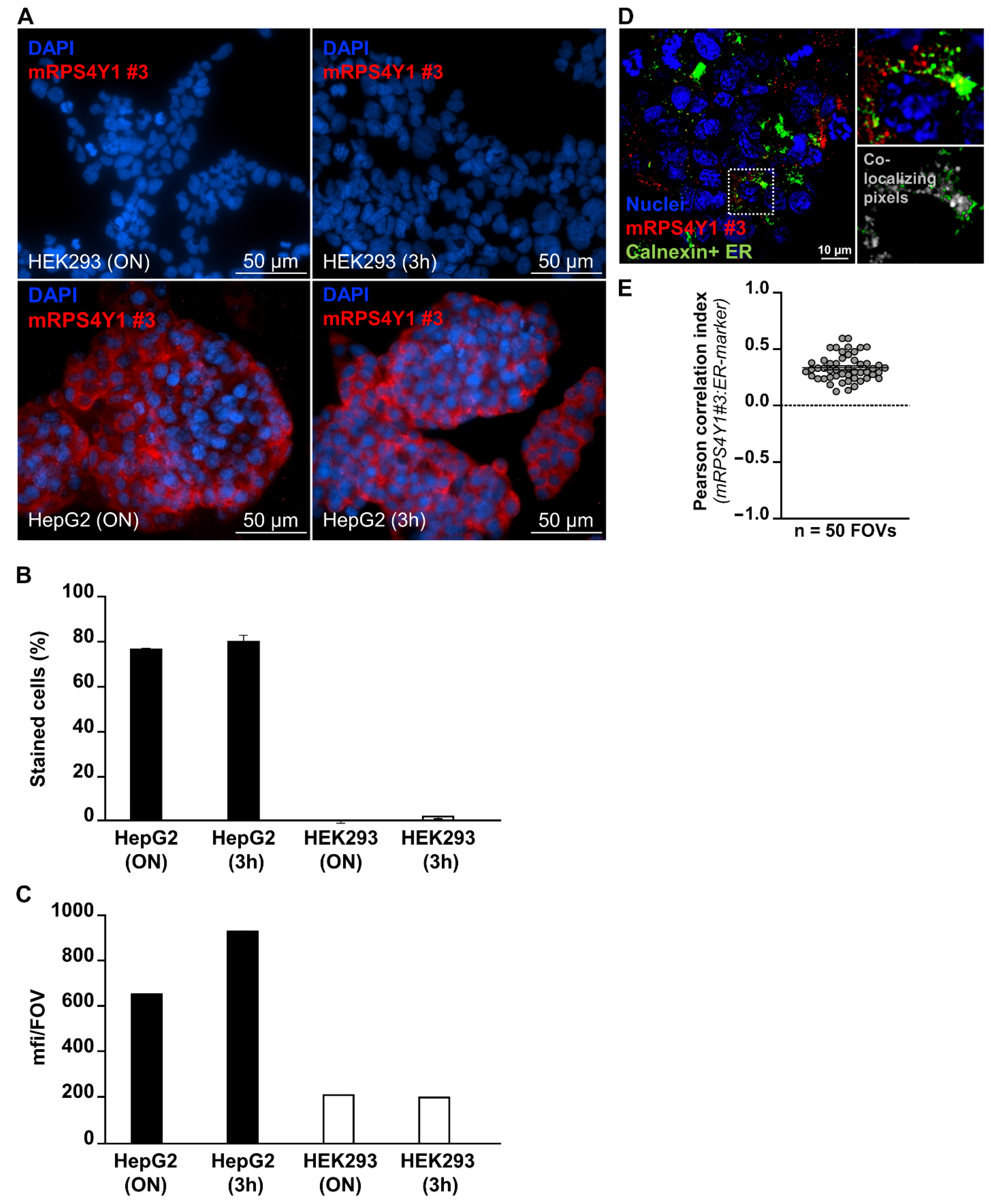
2.4. AntiRPS4Y1 Antibody Is Specific for Male Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Mononuclear Cell Isolation
4.2. Protein Extraction and Quantification
4.3. RNA Extraction and Reverse Transcription
4.4. PCR and qPCR
4.5. Sequencing
4.6. Peptide Production and Mice Immunization
4.7. Antibody Screening by ELISA
4.8. Detection of RPS4Y1 Protein by Western Blotting
4.9. Detection of RPS4Y1 Protein by Immunoprecipitation
4.10. Detection of RPS4Y1 Protein by Immunofluorescence
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mannucci, P.M.; Tuddenham, E.G. The hemophilias—From royal genes to gene therapy. N. Engl. J. Med. 2001, 344, 1773–1779. [Google Scholar] [CrossRef] [PubMed]
- Lo, Y.M.; Corbetta, N.; Chamberlain, P.F.; Rai, V.; Sargent, I.L.; Redman, C.W.; Wainscoat, J.S. Presence of fetal DNA in maternal plasma and serum. Lancet 1997, 350, 485–487. [Google Scholar] [CrossRef]
- Devaney, S.A.; Palomaki, G.E.; Scott, J.A.; Bianchi, D.W. Noninvasive fetal sex determination using cell-free fetal DNA: A systematic review and meta-analysis. JAMA 2011, 306, 627–636. [Google Scholar] [CrossRef] [PubMed]
- Mokari-Zadeh, N.; Mesbah-Namin, S.A. Evaluation of an Improved Non-invasive Fetal Sex Determination in Hemophilia A Patients. J. Clin. Diagn. Res. 2015, 9, GC01–GC04. [Google Scholar] [PubMed]
- Tsui, N.B.; Kadir, R.A.; Chan, K.C.; Chi, C.; Mellars, G.; Tuddenham, E.G.; Leung, T.Y.; Lau, T.K.; Chiu, R.W.; Lo, Y.M. Noninvasive prenatal diagnosis of hemophilia by microfluidics digital PCR analysis of maternal plasma DNA. Blood 2011, 117, 3684–3691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudecova, I.; Jiang, P.; Davies, J.; Lo, Y.M.D.; Kadir, R.A.; Chiu, R.W.K. Noninvasive detection of F8 int22h-related inversions and sequence variants in maternal plasma of hemophilia carriers. Blood 2017, 130, 340–347. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Sun, J.; Yang, Y.; Jiang, L.; Guo, F.; Zhu, Y.; Li, D.; Wu, R.; Lu, R.; Zhao, M.; et al. Noninvasive prenatal diagnosis of hemophilia A by a haplotype-based approach using cell-free fetal DNA. Biotechniques 2020, 68, 117–121. [Google Scholar] [CrossRef]
- Beta, J.; Lesmes-Heredia, C.; Bedetti, C.; Akolekar, R. Risk of miscarriage following amniocentesis and chorionic villus sampling: A systematic review of the literature. Minerva Ginecol. 2018, 70, 215–219. [Google Scholar]
- Ljung, R.C. Prenatal diagnosis of hemophilia. Hemophilia 1999, 5, 84–87. [Google Scholar] [CrossRef]
- Fiddler, M. Fetal Cell Based Prenatal Diagnosis: Perspectives on the Present and Future. J. Clin. Med. 2014, 3, 972–985. [Google Scholar] [CrossRef]
- Mergenthaler, S.; Babochkina, T.; Kiefer, V.; Lapaire, O.; Holzgreve, W.; Hahn, S. FISH analysis of all fetal nucleated cells in maternal whole blood: Improved specificity by the use of two Y-chromosome probes. J. Histochem. Cytochem. 2005, 53, 319–322. [Google Scholar] [CrossRef]
- Rezaei, M.; Winter, M.; Zander-Fox, D.; Whitehead, C.; Liebelt, J.; Warkiani, M.E.; Hardy, T.; Thierry, B. A Reappraisal of Circulating Fetal Cell Noninvasive Prenatal Testing. Trends Biotechnol. 2019, 37, 632–644. [Google Scholar] [CrossRef]
- Gänshirt-Ahlert, D.; Burschyk, M.; Garritsen, H.S.; Helmer, L.; Miny, P.; Horst, J.; Schneider, H.P.; Holzgreve, W. Magnetic cell sorting and the transferrin receptor as potential means of prenatal diagnosis from maternal blood. Am. J. Obstet. Gynecol. 1992, 166, 1350–1355. [Google Scholar] [CrossRef]
- Bianchi, D.W.; Zickwolf, G.K.; Yih, M.C.; Flint, A.F.; Geifman, O.H.; Erikson, M.S.; Williams, J.M. Erythroid-specific antibodies enhance detection of fetal nucleated erythrocytes in maternal blood. Prenat. Diagn. 1993, 13, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Mavrou, A.; Kolialexi, A.; Antsaklis, A.; Korantzis, A.; Metaxotou, C. Identification of fetal nucleated red blood cells in the maternal circulation during pregnancy using anti-hemoglobin-epsilon antibody. Fetal. Diagn. Ther. 2003, 18, 309–313. [Google Scholar] [CrossRef]
- Sekizawa, A.; Purwosunu, Y.; Farina, A.; Okai, T.; Takabayashi, H.; Kita, M.; Yura, H.; Kitagawa, M. Development of noninvasive fetal DNA diagnosis from nucleated erythrocytes circulating in maternal blood. Prenat. Diagn. 2007, 27, 846–848. [Google Scholar] [CrossRef]
- Zimmermann, S.; Hollmann, C.; Stachelhaus, S.A. Unique monoclonal antibodies specifically bind surface structures on human fetal erythroid blood cells. Exp. Cell. Res. 2013, 319, 2700–2707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, L.; Zhu, X.; Li, R.; Wu, H.; Chen, W.; Chen, J.; Liu, H.; Li, S.; Liu, P. A novel method for noninvasive diagnosis of monogenic diseases from circulating fetal cells. Prenat. Diagn. 2020, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hatt, L.; Brinch, M.; Singh, R.; Møller, K.; Lauridsen, R.H.; Uldbjerg, N.; Huppertz, B.; Christensen, B.; Kølvraa, S. Characterization of fetal cells from the maternal circulation by microarray gene expression analysis—Could the extravillous trophoblasts be a target for future cell-based non-invasive prenatal diagnosis? Fetal Diagn. Ther. 2014, 35, 218–227. [Google Scholar] [CrossRef]
- Kølvraa, S.; Singh, R.; Normand, E.A.; Qdaisat, S.; van den Veyver, I.B.; Jackson, L.; Hatt, L.; Schelde, P.; Uldbjerg, N.; Vestergaard, E.M.; et al. Genome-wide copy number analysis on DNA from fetal cells isolated from the blood of pregnant women. Prenat. Diagn. 2016, 36, 1127–1134. [Google Scholar] [CrossRef]
- Brinch, M.; Hatt, L.; Singh, R.; Møller, K.; Sommer, S.; Uldbjerg, N.; Christensen, B.; Kølvraa, S. Identification of circulating fetal cell markers by microarray analysis. Prenat. Diagn. 2012, 32, 742–751. [Google Scholar] [CrossRef]
- Mouawia, H.; Saker, A.; Jais, J.P.; Benachi, A.; Bussières, L.; Lacour, B.; Bonnefont, J.P.; Frydman, R.; Simpson, J.L.; Paterlini-Brechot, P. Circulating trophoblastic cells provide genetic diagnosis in 63 fetuses at risk for cystic fibrosis or spinal muscular atrophy. Reprod. Biomed. Online. 2012, 25, 508–520. [Google Scholar] [CrossRef] [Green Version]
- Breman, A.M.; Chow, J.C.; U’Ren, L.; Normand, E.A.; Qdaisat, S.; Zhao, L.; Henke, D.M.; Chen, R.; Shaw, C.A.; Jackson, L.; et al. Evidence for feasibility of fetal trophoblastic cell-based noninvasive prenatal testing. Prenat. Diagn. 2016, 36, 1009–1019. [Google Scholar] [CrossRef] [Green Version]
- Hou, S.; Chen, J.F.; Song, M.; Zhu, Y.; Jan, Y.J.; Chen, S.H.; Weng, T.H.; Ling, D.A.; Chen, S.F.; Ro, T.; et al. Imprinted NanoVelcro Microchips for Isolation and Characterization of Circulating Fetal Trophoblasts: Toward Noninvasive Prenatal Diagnostics. ACS Nano. 2017, 11, 8167–8177. [Google Scholar] [CrossRef] [PubMed]
- Zinn, A.R.; Alagappan, R.K.; Brown, L.G.; Wool, I.; Page, D.C. Structure and function of ribosomal protein S4 genes on the human and mouse sex chromosomes. Mol. Cell. Biol. 1994, 14, 2485–2492. [Google Scholar] [CrossRef] [Green Version]
- Lopes, A.M.; Miguel, R.N.; Sargent, C.A.; Ellis, P.J.; Amorim, A.; Affara, N.A. The human RPS4 paralogue on Yq11.223 encodes a structurally conserved ribosomal protein and is preferentially expressed during spermatogenesis. BMC Mol. Biol. 2010, 11, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, K.; McCormack, C.E.; Bradbury, N.A. Do you know the sex of your cells? Am. J. Physiol. Cell. Physiol. 2014, 306, C3–C18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peyvandi, F.; Garagiola, I.; Boscarino, M.; Ryan, A.; Hermans, C.; Makris, M. Real-life experience in switching to new extended half-life products at European hemophilia centres. Hemophilia 2019, 25, 946–952. [Google Scholar] [CrossRef]
- Witmer, C.; Young, G. Factor VIII inhibitors in hemophilia A: Rationale and latest evidence. Ther. Adv. Hematol. 2013, 4, 59–72. [Google Scholar] [CrossRef] [Green Version]
- Batty, P.; Lillicrap, D. Advances and challenges for hemophilia gene therapy. Hum. Mol. Genet. 2019, 28, R95–R101. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Chang, S.P.; Ma, G.C.; Lin, W.H.; Chen, H.F.; Chen, S.U.; Tsai, H.D.; Tsai, F.P.; Shen, M.C. Preimplantation genetic diagnosis of hemophilia A. Thromb. J. 2016, 14 (Suppl. 1), 33. [Google Scholar] [CrossRef] [Green Version]
- Bowman-Smart, H.; Savulescu, J.; Gyngell, C.; Mand, C.; Delatycki, M.B. Sex selection and non-invasive prenatal testing: A review of current practices, evidence, and ethical issues. Prenat. Diagn. 2020, 40, 398–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackie, F.L.; Hemming, K.; Allen, S.; Morris, R.K.; Kilby, M.D. The accuracy of cell-free fetal DNA-based non-invasive prenatal testing in singleton pregnancies: A systematic review and bivariate meta-analysis. BJOG 2017, 124, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Daley, R.; Hill, M.; Chitty, L.S. Non-invasive prenatal diagnosis: Progress and potential. Arch. Dis. Child Fetal Neonatal Ed. 2014, 99, F426–F430. [Google Scholar] [CrossRef] [PubMed]
- Lun, F.M.; Chiu, R.W.; Chan, K.C.; Leung, T.Y.; Lau, T.K.; Lo, Y.M. Microfluidics digital PCR reveals a higher than expected fraction of fetal DNA in maternal plasma. Clin. Chem. 2008, 54, 1664–1672. [Google Scholar] [CrossRef] [Green Version]
- Breveglieri, G.; D’Aversa, E.; Finotti, A.; Borgatti, M. Non-invasive Prenatal Testing Using Fetal DNA. Mol. Diagn. Ther. 2019, 23, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yang, Y.; Li, X.; Shi, Y.; Hu, B.; An, Y.; Zhu, Z.; Hong, G.; Yang, C.J. Frequency-enhanced transferrin receptor antibody-labelled microfluidic chip (FETAL-Chip) enables efficient enrichment of circulating nucleated red blood cells for non-invasive prenatal diagnosis. Lab. Chip. 2018, 18, 2749–2756. [Google Scholar] [CrossRef]
- Zhou, Q.; Wang, T.; Leng, L.; Zheng, W.; Huang, J.; Fang, F.; Yang, L.; Chen, F.; Lin, G.; Wang, W.J.; et al. Single-cell RNA-seq reveals distinct dynamic behavior of sex chromosomes during early human embryogenesis. Mol. Reprod. Dev. 2019, 86, 871–882. [Google Scholar] [CrossRef]
- Chen, X.; Tong, C.; Li, H.; Peng, W.; Li, R.; Luo, X.; Ge, H.; Ran, Y.; Li, Q.; Liu, Y.; et al. Dysregulated Expression of RPS4Y1 (Ribosomal Protein S4, Y-Linked 1) Impairs STAT3 (Signal Transducer and Activator of Transcription 3) Signaling to Suppress Trophoblast Cell Migration and Invasion in Preeclampsia. Hypertension 2018, 71, 481–490. [Google Scholar] [CrossRef]
- Winkelmann, D.A.; Kahan, L.; Lake, J.A. Ribosomal protein S4 is an internal protein: Localization by immunoelectron microscopy on protein-deficient subribosomal particles. Proc. Natl. Acad. Sci. USA 1982, 79, 5184–5188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, M.; Zinn, A.R.; Page, D.C.; Nishimoto, T. Functional equivalence of human X- and Y-encoded isoforms of ribosomal protein S4 consistent with a role in Turner syndrome. Nat. Genet. 1993, 4, 268–271. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spena, S.; Cordiglieri, C.; Garagiola, I.; Peyvandi, F. Development of a Specific Monoclonal Antibody to Detect Male Cells Expressing the RPS4Y1 Protein. Int. J. Mol. Sci. 2021, 22, 2001. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22042001
Spena S, Cordiglieri C, Garagiola I, Peyvandi F. Development of a Specific Monoclonal Antibody to Detect Male Cells Expressing the RPS4Y1 Protein. International Journal of Molecular Sciences. 2021; 22(4):2001. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22042001
Chicago/Turabian StyleSpena, Silvia, Chiara Cordiglieri, Isabella Garagiola, and Flora Peyvandi. 2021. "Development of a Specific Monoclonal Antibody to Detect Male Cells Expressing the RPS4Y1 Protein" International Journal of Molecular Sciences 22, no. 4: 2001. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22042001