
Resveratrol as Chemosensitizer Agent: State of Art and Future Perspectives

 , , , and
, , , and
Abstract
:1. Introduction
2. Natural Compounds as a Strategy for Chemosensitization
3. Mechanism of Action of Resveratrol in Cancer
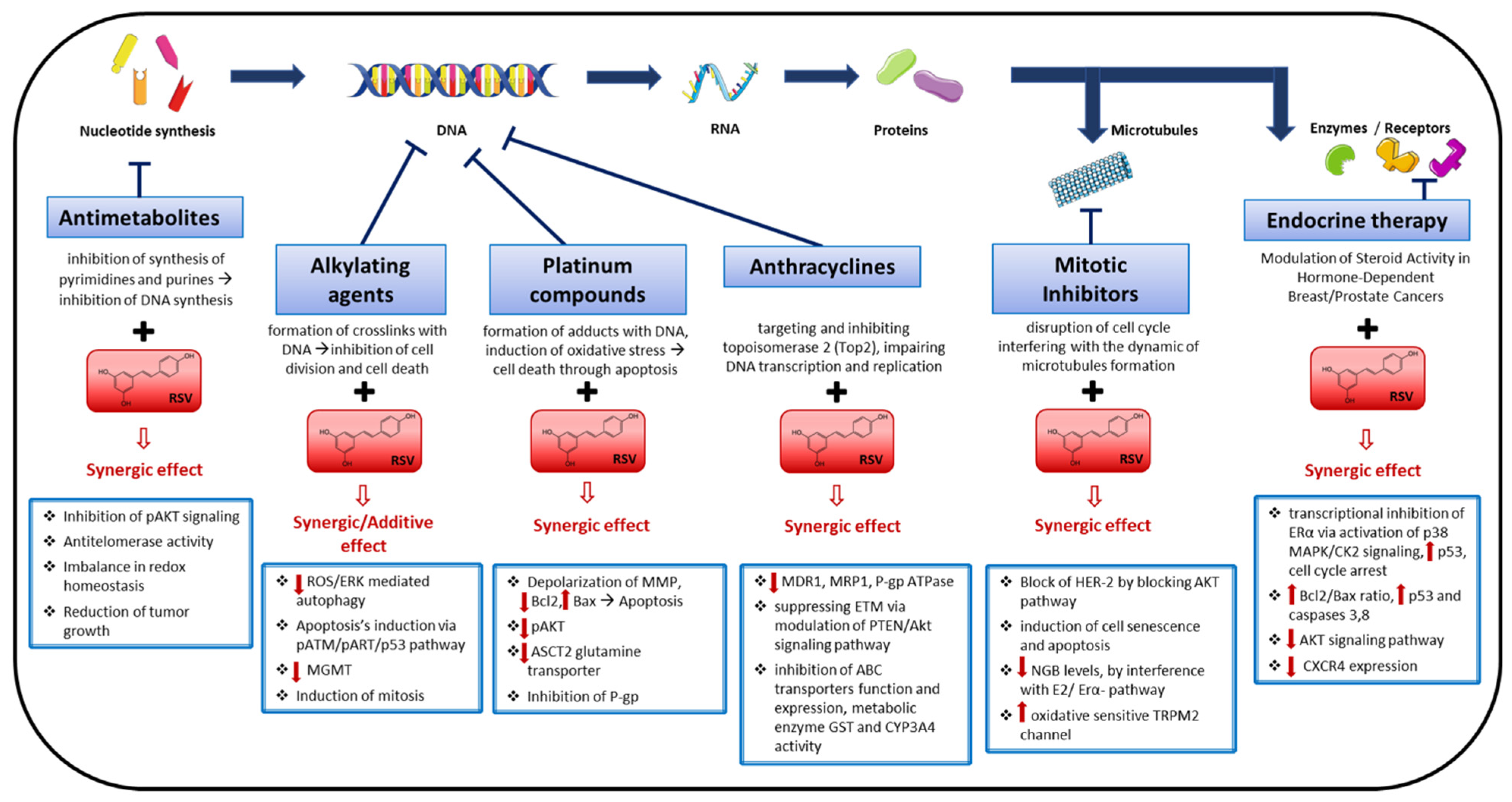
4. Resveratrol as a Chemosensitizer Agent
4.1. Alkylating Agents
4.2. Platinum Compounds
4.3. Anthracyclines
4.4. Antimetabolites
4.5. Mitotic Inhibitors
4.6. Endocrine Therapy
5. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Sun, J.; Wei, Q.; Zhou, Y.; Wang, J.; Liu, Q.; Xu, H. A systematic analysis of FDA-approved anticancer drugs. BMC Syst. Biol. 2017, 11, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Kunnumakkara, A.B.; Bordoloi, D.; Sailo, B.L.; Roy, N.K.; Thakur, K.K.; Banik, K.; Shakibaei, M.; Gupta, S.C.; Aggarwal, B.B. Cancer drug development: The missing links. Exp. Biol. Med. 2019, 244, 663–689. [Google Scholar] [CrossRef]
- Król, M.; Pawłowski, K.M.; Majchrzak, K.; Szyszko, K.; Motyl, T. Why Chemotherapy Can Fail? Polish J. Vet. Sci. 2010, 13, 399–406. [Google Scholar]
- Zheng, H.-C. The molecular mechanisms of chemoresistance in cancers. Oncotarget 2017, 8, 59950–59964. [Google Scholar] [CrossRef] [Green Version]
- Brasseur, K.; Gévry, N.; Asselin, E. Chemoresistance and targeted therapies in ovarian and endometrial cancers. Oncotarget 2016, 8, 4008–4042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morandi, A.; Indraccolo, S. Linking metabolic reprogramming to therapy resistance in cancer. Biochim. Biophys. Acta (BBA) Bioenerg. 2017, 1868, 1–6. [Google Scholar] [CrossRef]
- Cairns, R.A.; Harris, I.S.; Mak, T.W. Regulation of cancer cell metabolism. Nat. Rev. Cancer 2011, 11, 85–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, S.C.; Kannappan, R.; Reuter, S.; Kim, J.H.; Aggarwal, B.B. Chemosensitization of tumors by resveratrol. Ann. N. Y. Acad. Sci. 2011, 1215, 150–160. [Google Scholar] [CrossRef] [Green Version]
- Cragg, G.M.; Pezzuto, J.M. Natural Products as a Vital Source for the Discovery of Cancer Chemotherapeutic and Chemopreventive Agents. Med. Princ. Pract. 2015, 25, 41–59. [Google Scholar] [CrossRef] [PubMed]
- Catanzaro, D.; Gabbia, D.; Cocetta, V.; Biagi, M.; Ragazzi, E.; Montopoli, M.; Carrara, M. Silybin counteracts doxorubicin resistance by inhibiting GLUT1 expression. Fitoterapia 2018, 124, 42–48. [Google Scholar] [CrossRef]
- Montopoli, M.; Ragazzi, E.; Froldi, G.; Caparrotta, L. Cell-cycle inhibition and apoptosis induced by curcumin and cisplatin or oxaliplatin in human ovarian carcinoma cells. Cell Prolif. 2009, 42, 195–206. [Google Scholar] [CrossRef] [Green Version]
- Berretta, M.; Quagliariello, V.; Maurea, N.; Di Francia, R.; Sharifi, S.; Facchini, G.; Rinaldi, L.; Piezzo, M.; Manuela, C.; Nunnari, G.; et al. Multiple Effects of Ascorbic Acid against Chronic Diseases: Updated Evidence from Preclinical and Clinical Studies. Antioxidants 2020, 9, 1182. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.-H.; Sethi, G.; Um, J.-Y.; Shanmugam, M.K.; Arfuso, F.; Kumar, A.P.; Bishayee, A.; Ahn, K.S. The Role of Resveratrol in Cancer Therapy. Int. J. Mol. Sci. 2017, 18, 2589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burns, J.; Yokota, T.; Ashihara, H.; Lean, M.E.J.; Crozier, A. Plant Foods and Herbal Sources of Resveratrol. J. Agric. Food Chem. 2002, 50, 3337–3340. [Google Scholar] [CrossRef]
- Dercks, W.; Creasy, L. The significance of stilbene phytoalexins in the Plasmopara viticola-grapevine interaction. Physiol. Mol. Plant Pathol. 1989, 34, 189–202. [Google Scholar] [CrossRef]
- Park, E.-J.; Pezzuto, J.M. The pharmacology of resveratrol in animals and humans. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2015, 1852, 1071–1113. [Google Scholar] [CrossRef] [Green Version]
- Burkon, A.; Somoza, V. Quantification of free and protein-boundtrans-resveratrol metabolites and identification oftrans-resveratrol-C/O-conjugated diglucuronides—Two novel resveratrol metabolites in human plasma. Mol. Nutr. Food Res. 2008, 52, 549–557. [Google Scholar] [CrossRef]
- Lançon, A.; Hanet, N.; Jannin, B.; Delmas, M.; Heydel, J.-M.; Lizard, G.; Chagnon, M.-C.; Artur, Y.; Latruffe, N. Resveratrol in Human Hepatoma HepG2 Cells: Metabolism and Inducibility of Detoxifying Enzymes. Drug Metab. Dispos. 2007, 35, 699–703. [Google Scholar] [CrossRef] [Green Version]
- Jeandet, P.; Bessis, R.; Gautheron, B. The Production of Resveratrol (3,5,4′-Trihydroxystilbene) by Grape Berries in Different Developmental Stages. Am. J. Enol Vitic. 1991, 42, 41–46. [Google Scholar]
- Goldberg, D.M.; Yan, J.; Ng, E.; Diamandis, E.P.; Karumanchiri, A.; Soleas, G.; Waterhouse, A.L. A Global Survey of Trans-Resveratrol Concentrations in Commercial Wines. Am. J. Enol Vitic. 1995, 46, 159–165. [Google Scholar]
- Renaud, S.; De Lorgeril, M. Wine, alcohol, platelets, and the French paradox for coronary heart disease. Lancet 1992, 339, 1523–1526. [Google Scholar] [CrossRef]
- Jang, M.; Cai, L.; Udeani, G.O.; Slowing, K.V.; Thomas, C.F.; Beecher, C.W.W.; Fong, H.H.S.; Farnsworth, N.R.; Kinghorn, A.D.; Mehta, R.G.; et al. Cancer Chemopreventive Activity of Resveratrol, a Natural Product Derived from Grapes. Science 1997, 275, 218–220. [Google Scholar] [CrossRef] [Green Version]
- Kuršvietienė, L.; Stanevičienė, I.; Mongirdienė, A.; Bernatonienė, J. Multiplicity of effects and health benefits of resveratrol. Rev. Esp. Med. Nucl. Imagen Mol. 2016, 52, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Singh, C.K.; Ndiaye, M.A.; Ahmad, N. Resveratrol and cancer: Challenges for clinical translation. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2015, 1852, 1178–1185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ndiaye, M.; Kumar, R.; Ahmad, N. Resveratrol in cancer management: where are we and where we go from here? Ann. N. Y. Acad. Sci. 2011, 1215, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Carter, L.G.; D’Orazio, J.A.; Pearson, K.J. Resveratrol and cancer: focus on in vivo evidence. Endocr. Relat. Cancer 2014, 21, R209–R225. [Google Scholar] [CrossRef] [Green Version]
- Kotecha, R.; Takami, A.; Espinoza, J.L. Dietary phytochemicals and cancer chemoprevention: a review of the clinical evidence. Oncotarget 2016, 7, 52517–52529. [Google Scholar] [CrossRef] [Green Version]
- Nawaz, W.; Zhou, Z.; Deng, S.; Ma, X.; Ma, X.; Li, C.; Shu, X. Therapeutic Versatility of Resveratrol Derivatives. Nutrients 2017, 9, 1188. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.; Namkoong, K.; Shin, M.; Park, J.; Yang, E.; Ihm, J.; Thu, V.T.; Kim, H.K.; Han, J. Cardiovascular Protective Effects and Clinical Applications of Resveratrol. J. Med. Food 2017, 20, 323–334. [Google Scholar] [CrossRef]
- Cai, J.; Liu, W.; Lu, F.; Kong, W.; Zhou, X.; Miao, P.; Lei, C.; Wang, Y. Resveratrol attenuates neurological deficit and neuroinflammation following intracerebral hemorrhage. Exp. Ther. Med. 2018, 15, 4131–4138. [Google Scholar] [CrossRef]
- Berretta, M.; Bignucolo, A.; Di Francia, R.; Comello, F.; Facchini, G.; Ceccarelli, M.; Iaffaioli, R.V.; Quagliariello, V.; Maurea, N. Resveratrol in Cancer Patients: From Bench to Bedside. Int. J. Mol. Sci. 2020, 21, 2945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, W.K. Recent Advances in Chemoprevention of Cancer. Science 1997, 278, 1073–1077. [Google Scholar] [CrossRef]
- Sethi, G.; Shanmugam, M.K.; Ramachandran, L.; Kumar, A.P.; Tergaonkar, V. Multifaceted link between cancer and inflammation. Biosci. Rep. 2011, 32, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brockmueller, A.; Sameri, S.; Liskova, A.; Zhai, K.; Varghese, E.; Samuel, S.M.; Büsselberg, D.; Kubatka, P.; Shakibaei, M. Resveratrol’s Anti-Cancer Effects through the Modulation of Tumor Glucose Metabolism. Cancers 2021, 13, 188. [Google Scholar] [CrossRef]
- Shakibaei, M.; Harikumar, K.B.; Aggarwal, B.B. Resveratrol addiction: To die or not to die. Mol. Nutr. Food Res. 2008, 53, 115–128. [Google Scholar] [CrossRef]
- Han, Y.; Jo, H.; Cho, J.H.; Dhanasekaran, D.N.; Song, Y.S. Resveratrol as a Tumor-Suppressive Nutraceutical Modulating Tumor Microenvironment and Malignant Behaviors of Cancer. Int. J. Mol. Sci. 2019, 20, 925. [Google Scholar] [CrossRef] [Green Version]
- Lohse, I.; Wildermuth, E.; Brothers, S.P. Naturally occurring compounds as pancreatic cancer therapeutics. Oncotarget 2018, 9, 35448–35457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buhrmann, C.; Shayan, P.; Popper, B.; Goel, A.; Shakibaei, M. Sirt1 Is Required for Resveratrol-Mediated Chemopreventive Effects in Colorectal Cancer Cells. Nutrients 2016, 8, 145. [Google Scholar] [CrossRef] [Green Version]
- Buhrmann, C.; Shayan, P.; Goel, A.; Shakibaei, M. Resveratrol Regulates Colorectal Cancer Cell Invasion by Modulation of Focal Adhesion Molecules. Nutrients 2017, 9, 1073. [Google Scholar] [CrossRef] [PubMed]
- Busch, F.; Mobasheri, A.; Shayan, P.; Lueders, C.; Stahlmann, R.; Shakibaei, M. Resveratrol Modulates Interleukin-1β-induced Phosphatidylinositol 3-Kinase and Nuclear Factor κB Signaling Pathways in Human Tenocytes. J. Biol. Chem. 2012, 287, 38050–38063. [Google Scholar] [CrossRef] [Green Version]
- Bourguignon, L.Y.W.; Xia, W.; Wong, G. Hyaluronan-mediated CD44 Interaction with p300 and SIRT1 Regulates β-Catenin Signaling and NFκB-specific Transcription Activity Leading to MDR1 and Bcl-xL Gene Expression and Chemoresistance in Breast Tumor Cells. J. Biol. Chem. 2009, 284, 2657–2671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Jiang, S.; Yu, F.; Zhou, L.; Wang, K. Noncoding RNAs as Molecular Targets of Resveratrol Underlying Its Anticancer Effects. J. Agric. Food Chem. 2019, 67, 4709–4719. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Sun, S.; Yu, W.; Jiang, J.; Zhuo, F.; Qiu, G.; Xu, S.; Jiang, X. Altered expression of long non-coding RNAs during genotoxic stress-induced cell death in human glioma cells. J. Neuro-Oncol. 2015, 122, 283–292. [Google Scholar] [CrossRef]
- Yang, Y.; Xu, H.; Huang, W.; Ding, M.; Xiao, J.; Yang, D.; Li, H.; Liu, X.; Chu, L. Targeting lung cancer stem-like cells with TRAIL gene armed oncolytic adenovirus. J. Cell. Mol. Med. 2015, 19, 915–923. [Google Scholar] [CrossRef] [PubMed]
- Ji, Q.; Liu, X.; Fu, X.; Zhang, L.; Sui, H.; Zhou, L.; Sun, J.; Cai, J.; Qin, J.; Ren, J.; et al. Resveratrol Inhibits Invasion and Metastasis of Colorectal Cancer Cells via MALAT1 Mediated Wnt/β-Catenin Signal Pathway. PLoS ONE 2013, 8, e78700. [Google Scholar] [CrossRef]
- A Benitez, D.; Pozo-Guisado, E.; Clementi, M.; Castellón, E.; Fernandez-Salguero, P.M. Non-genomic action of resveratrol on androgen and oestrogen receptors in prostate cancer: modulation of the phosphoinositide 3-kinase pathway. Br. J. Cancer 2007, 96, 1595–1604. [Google Scholar] [CrossRef] [Green Version]
- Sarkaria, J.N.; Kitange, G.J.; James, C.D.; Plummer, R.; Calvert, H.; Weller, M.; Wick, W. Mechanisms of Chemoresistance to Alkylating Agents in Malignant Glioma. Clin. Cancer Res. 2008, 14, 2900–2908. [Google Scholar] [CrossRef] [Green Version]
- Kiskova, T.; Kubatka, P.; Büsselberg, D.; Kassayova, M. The Plant-Derived Compound Resveratrol in Brain Cancer: A Review. Biomolecules 2020, 10, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiapaer, S.; Furuta, T.; Tanaka, S.; Kitabayashi, T.; Nakada, M. Potential Strategies Overcoming the Temozolomide Resistance for Glioblastoma. Neurol. Medico-Chir. 2018, 58, 405–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, G.-Z.; Huang, G.; Guo, M.; Zhang, X.; Wang, H.; Deng, S.; Li, Y.; Xiang, W.; Chen, Z.; Pan, J.; et al. Acquired temozolomide resistance in MGMT-deficient glioblastoma cells is associated with regulation of DNA repair by DHC2. Brain 2019, 142, 2352–2366. [Google Scholar] [CrossRef]
- Rath, G.K.; Mallick, S.; Gandhi, A.K. Therapeutic approach beyond conventional temozolomide for newly diagnosed glioblastoma: Review of the present evidence and future direction. Indian J. Med Paediatr. Oncol. 2015, 36, 229. [Google Scholar] [CrossRef] [Green Version]
- Decleves, X.; Amiel, A.; Scherrmann, J.-Y.D.; Scherrmann, J.M. Role of ABC Transporters in the Chemoresistance of Human Gliomas. Available online: https://www.eurekaselect.com/56392/article (accessed on 15 December 2020).
- Yuan, Y.; Xue, X.; Guo, R.-B.; Sun, X.-L.; Hu, G. Resveratrol Enhances the Antitumor Effects of Temozolomide in Glioblastoma via ROS-dependent AMPK-TSC-mTOR Signaling Pathway. CNS Neurosci. Ther. 2012, 18, 536–546. [Google Scholar] [CrossRef]
- Li, H.; Liu, Y.; Jiao, Y.; Guo, A.; Xu, X.; Qu, X.; Wang, S.; Zhao, J.; Li, Y.; Cao, Y. Resveratrol sensitizes glioblastoma-initiating cells to temozolomide by inducing cell apoptosis and promoting differentiation. Oncol. Rep. 2015, 35, 343–351. [Google Scholar] [CrossRef] [Green Version]
- Özdemir, F.; Apaydın, E.; Önder, N.I.; Şen, M.; Ayrım, A.; Öğünç, Y.; Incesu, Z. Apoptotic effects of ε-viniferin in combination with cis-platin in C6 cells. Cytotechnology 2018, 70, 1061–1073. [Google Scholar] [CrossRef]
- Zhang, X.; Huang, H.; Lin, H.; Li, J. Resveratrol reverses temozolomide resistance by downregulation of MGMT in T98G glioblastoma cells by the NF-κB-dependent pathway. Oncol. Rep. 2012, 27, 2050–2056. [Google Scholar] [CrossRef]
- Yang, H.-C.; Wang, J.-Y.; Bu, X.-Y.; Yang, B.; Wang, B.-Q.; Hu, S.; Yan, Z.-Y.; Gao, Y.-S.; Han, S.-Y.; Qu, M.-Q. Resveratrol restores sensitivity of glioma cells to temozolamide through inhibiting the activation of Wnt signaling pathway. J. Cell. Physiol. 2019, 234, 6783–6800. [Google Scholar] [CrossRef] [PubMed]
- Filippi-Chiela, E.C.; Thomé, M.P.; E Silva, M.M.B.; Pelegrini, A.L.; Ledur, P.F.; Garicochea, B.; Zamin, L.L.; Lenz, G. Resveratrol abrogates the Temozolomide-induced G2 arrest leading to mitotic catastrophe and reinforces the Temozolomide-induced senescence in glioma cells. BMC Cancer 2013, 13, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makovec, T. Cisplatin and beyond: molecular mechanisms of action and drug resistance development in cancer chemotherapy. Radiol. Oncol. 2019, 53, 148–158. [Google Scholar] [CrossRef] [Green Version]
- Galluzzi, L.; Vitale, I.; Michels, J.; Brenner, C.; Szabadkai, G.; Harelbellan, A.; Castedo, M.; Kroemer, G. Systems biology of cisplatin resistance: past, present and future. Cell Death Dis. 2014, 5, e1257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galluzzi, L.; Senovilla, L.; Vitale, I.; Michels, J.J.; Martins, I.; Kepp, O.; Castedo, M.; Kroemer, G. Molecular mechanisms of cisplatin resistance. Oncogene 2011, 31, 1869–1883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cocetta, V.; Ragazzi, E.; Montopoli, M. Links between cancer metabolism and cisplatin resistance. Int. Rev. Cell Mol. Biol. 2020, 354, 107–164. [Google Scholar] [CrossRef] [PubMed]
- Cocetta, V.; Ragazzi, E.; Montopoli, M. Mitochondrial Involvement in Cisplatin Resistance. Int. J. Mol. Sci. 2019, 20, 3384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giacomini, I.; Cocetta, V.; Carrara, M.; Ragazzi, E.; Montopoli, M. Plumbagin Induces Cell Cycle Arrest and Apoptosis in A431 Cisplatin-Resistant Cancer Cells. Nat. Prod. Commun. 2020, 15. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Li, W.; Wang, R.; Nan, Y.; Wang, Q.; Liu, W.; Jin, F. Resveratrol enhanced anticancer effects of cisplatin on non-small cell lung cancer cell lines by inducing mitochondrial dysfunction and cell apoptosis. Int. J. Oncol. 2015, 47, 1460–1468. [Google Scholar] [CrossRef] [Green Version]
- Xing, C.; Zhu, B.; Liu, H.; Yao, H.; Zhang, L. Class I phosphatidylinositol 3-kinase inhibitor LY294002 activates autophagy and induces apoptosis through p53 pathway in gastric cancer cell line SGC7901. Acta Biochim. Biophys. Sin. 2008, 40, 194–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hui, K.; Li, X.; Xu, R.; Ye, L.; Kong, H.; Zeng, X.; Wang, H.; Xie, W. The synergistic effect of resveratrol in combination with cisplatin on apoptosis via modulating autophagy in A549 cells. Acta Biochim. Biophys. Sin. 2016, 48, 528–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wise, D.R.; Thompson, C.B. Glutamine addiction: A new therapeutic target in cancer. Trends Biochem. Sci. 2010, 35, 427–433. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Peng, Q.; Li, Y.; Gao, Y. Resveratrol enhances cisplatin-induced apoptosis in human hepatoma cells via glutamine metabolism inhibition. BMB Rep. 2018, 51, 474–479. [Google Scholar] [CrossRef] [Green Version]
- Nessa, M.U.; Beale, P.; Chan, C.; Yu, J.Q.; Huq, F. Combinations of resveratrol, cisplatin and oxaliplatin applied to human ovarian cancer cells. Anticancer Res. 2012, 32, 53–59. [Google Scholar]
- Engelke, L.H.; Hamacher, A.; Proksch, P.; Kassack, M.U. Ellagic Acid and Resveratrol Prevent the Development of Cisplatin Resistance in the Epithelial Ovarian Cancer Cell Line A2780. J. Cancer 2016, 7, 353–363. [Google Scholar] [CrossRef] [Green Version]
- Rezk, Y.A.; Balulad, S.S.; Keller, R.S.; Bennett, J.A. Use of Resveratrol to improve the effectiveness of cisplatin and doxorubicin: Study in human gynecologic cancer cell lines and in rodent heart. Am. J. Obstet. Gynecol. 2006, 194, e23–e26. [Google Scholar] [CrossRef]
- Björklund, M.; Roos, J.; Gogvadze, V.; Shoshan, M. Resveratrol induces SIRT1- and energy–stress-independent inhibition of tumor cell regrowth after low-dose platinum treatment. Cancer Chemother. Pharmacol. 2011, 68, 1459–1467. [Google Scholar] [CrossRef] [PubMed]
- Daniel, N.P.; Ezekiel, U. Antiproliferative Effect of Resveratrol and Oxaliplatin on Colon Cancer Cells. FASEB J. 2016, 30, 1090.11. [Google Scholar] [CrossRef]
- Park, D.-G. Antichemosensitizing effect of resveratrol in cotreatment with oxaliplatin in HCT116 colon cancer cell. Ann. Surg. Treat. Res. 2014, 86, 68–75. [Google Scholar] [CrossRef] [Green Version]
- Kaminski, B.M.; Weigert, A.; Scherzberg, M.-C.; Ley, S.; Gilbert, B.; Brecht, K.; Brüne, B.; Steinhilber, D.; Stein, J.; Ulrich-Rückert, S. Resveratrol-induced potentiation of the antitumor effects of oxaliplatin is accompanied by an altered cytokine profile of human monocyte-derived macrophages. Apoptosis 2014, 19, 1136–1147. [Google Scholar] [CrossRef]
- Osman, A.-M.M.; A Telity, S.; A Damanhouri, Z.; E Al-Harthy, S.; Al-Kreathy, H.M.; Ramadan, W.S.; ElShal, M.F.; Khan, L.M.; Kamel, F.; A Telity, S. Chemosensitizing and nephroprotective effect of resveratrol in cisplatin—Treated animals. Cancer Cell Int. 2015, 15, 6. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.-J.; Chang, M.-Y.; Chang, W.-W.; Wang, W.-K.; Liu, C.-F.; Lin, S.-T.; Lee, C.-H. Resveratrol Enhances Chemosensitivity in Mouse Melanoma Model Through Connexin 43 Upregulation. Environ. Toxicol. 2015, 30, 877–886. [Google Scholar] [CrossRef]
- Murabito, A.; Hirsch, E.; Ghigo, A. Mechanisms of Anthracycline-Induced Cardiotoxicity: Is Mitochondrial Dysfunction the Answer? Front. Cardiovasc. Med. 2020, 7, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broxterman, H.J.; Gotink, K.J.; Verheul, H.M. Understanding the causes of multidrug resistance in cancer: a comparison of doxorubicin and sunitinib. Drug Resist. Updat. 2009, 12, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.H.; Shin, Y.J.; Won, A.J.; Lee, B.M.; Choi, W.S.; Jung, J.H.; Chung, H.Y.; Kim, H.S. Resveratrol enhances chemosensitivity of doxorubicin in multidrug-resistant human breast cancer cells via increased cellular influx of doxorubicin. Biochim. Biophys. Acta (BBA) Gen. Subj. 2014, 1840, 615–625. [Google Scholar] [CrossRef]
- Rai, G.; Mishra, S.; Suman, S.; Shukla, Y. Resveratrol improves the anticancer effects of doxorubicin in vitro and in vivo models: A mechanistic insight. Phytomedicine 2016, 23, 233–242. [Google Scholar] [CrossRef]
- Fischer, K.R.; Durrans, A.; Lee, S.; Sheng, J.; Li, F.; Wong, S.T.C.; Choi, H.; El Rayes, T.; Ryu, S.; Troeger, J.; et al. Epithelial-to-mesenchymal transition is not required for lung metastasis but contributes to chemoresistance. Nature 2015, 527, 472–476. [Google Scholar] [CrossRef]
- Xu, J.; Liu, D.; Niu, H.; Zhu, G.; Xu, Y.; Ye, D.; Li, J.; Zhang, Q. Resveratrol reverses Doxorubicin resistance by inhibiting epithelial-mesenchymal transition (EMT) through modulating PTEN/Akt signaling pathway in gastric cancer. J. Exp. Clin. Cancer Res. 2017, 36, 19. [Google Scholar] [CrossRef] [Green Version]
- Jin, X.; Wei, Y.; Liu, Y.; Lu, X.; Ding, F.; Wang, J.; Yang, S. Resveratrol promotes sensitization to Doxorubicin by inhibiting epithelial-mesenchymal transition and modulating SIRT1/β-catenin signaling pathway in breast cancer. Cancer Med. 2019, 8, 1246–1257. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Jiang, H.; Chen, Y.; Ren, F. Resveratrol chemosensitizes adriamycin-resistant breast cancer cells by modulating miR-122-5p. J. Cell. Biochem. 2019, 120, 16283–16292. [Google Scholar] [CrossRef] [PubMed]
- Barros, A.S.; Costa, E.C.; Nunes, A.S.; De Melo-Diogo, D.; Correia, I.J. Comparative study of the therapeutic effect of Doxorubicin and Resveratrol combination on 2D and 3D (spheroids) cell culture models. Int. J. Pharm. 2018, 551, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Khaleel, S.A.; Al-Abd, A.M.; Ali, A.A.; Abdel-Naim, A.B. Didox and resveratrol sensitize colorectal cancer cells to doxorubicin via activating apoptosis and ameliorating P-glycoprotein activity. Sci. Rep. 2016, 6, 36855. [Google Scholar] [CrossRef]
- El-Readi, M.Z.; Eid, S.; Abdelghany, A.A.; Al-Amoudi, H.S.; Efferth, T.; Wink, M. Resveratrol mediated cancer cell apoptosis, and modulation of multidrug resistance proteins and metabolic enzymes. Phytomedicine 2019, 55, 269–281. [Google Scholar] [CrossRef]
- Kinsella, A.R.; Smith, D. Tumor Resistance To Antimetabolites. Gen. Pharmacol. Vasc. Syst. 1998, 30, 623–626. [Google Scholar] [CrossRef]
- Luqmani, Y.A. Mechanisms of Drug Resistance in Cancer Chemotherapy. Med. Princ. Pract. 2005, 14, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Miura, K.; Kinouchi, M.; Ishida, K.; Fujibuchi, W.; Naitoh, T.; Ogawa, H.; Ando, T.; Yazaki, N.; Watanabe, K.; Haneda, S.; et al. 5-FU Metabolism in Cancer and Orally-Administrable 5-FU Drugs. Cancers 2010, 2, 1717–1730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, N.; Yin, Y.; Xu, S.-J.; Chen, W.-S. 5-Fluorouracil: Mechanisms of Resistance and Reversal Strategies. Molecules 2008, 13, 1551–1569. [Google Scholar] [CrossRef] [Green Version]
- Chung, S.S.; Dutta, P.; Austin, D.; Wang, P.; Awad, A.; Vadgama, J.V. Combination of resveratrol and 5-flurouracil enhanced anti-telomerase activity and apoptosis by inhibiting STAT3 and Akt signaling pathways in human colorectal cancer cells. Oncotarget 2018, 9, 32943–32957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santandreu, F.M.; Valle, A.; Oliver, J.; Roca, P. Resveratrol Potentiates the Cytotoxic Oxidative Stress Induced by Chemotherapy in Human Colon Cancer Cells. Cell. Physiol. Biochem. 2011, 28, 219–228. [Google Scholar] [CrossRef]
- Lee, S.H.; Koo, B.S.; Park, S.Y.; Kim, Y.M. Anti-angiogenic effects of resveratrol in combination with 5-fluorouracil on B16 murine melanoma cells. Mol. Med. Rep. 2015, 12, 2777–2783. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.-L.; Sun, Z.-J.; Yu, L.; Meng, K.-W.; Qin, X.-L.; Pan, C.-E. Effect of resveratrol and in combination with 5-FU on murine liver cancer. World J. Gastroenterol. 2004, 10, 3048–3052. [Google Scholar] [CrossRef]
- Buhrmann, C.; Yazdi, M.; Popper, B.; Shayan, P.; Goel, A.; Aggarwal, B.B.; Shakibaei, M. Resveratrol Chemosensitizes TNF-β-Induced Survival of 5-FU-Treated Colorectal Cancer Cells. Nutrients 2018, 10, 888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buhrmann, C.; Shayan, P.; Kraehe, P.; Popper, B.; Goel, A.; Shakibaei, M. Resveratrol induces chemosensitization to 5-fluorouracil through up-regulation of intercellular junctions, Epithelial-to-mesenchymal transition and apoptosis in colorectal cancer. Biochem. Pharmacol. 2015, 98, 51–68. [Google Scholar] [CrossRef]
- Amrutkar, M.; Gladhaug, I.P. Pancreatic Cancer Chemoresistance to Gemcitabine. Cancers 2017, 9, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, C.; Qian, W.; Ma, J.; Cheng, L.; Jiang, Z.; Yan, B.; Li, J.; Duan, W.; Sun, L.; Cao, J.; et al. Resveratrol enhances the chemotherapeutic response and reverses the stemness induced by gemcitabine in pancreatic cancer cells via targeting SREBP1. Cell Prolif. 2019, 52, e12514. [Google Scholar] [CrossRef]
- Harikumar, K.B.; Kunnumakkara, A.B.; Sethi, G.; Diagaradjane, P.; Anand, P.; Pandey, M.K.; Gelovani, J.; Krishnan, S.; Guha, S.; Aggarwal, B.B. Resveratrol, a multitargeted agent, can enhance antitumor activity of gemcitabinein vitroand in orthotopic mouse model of human pancreatic cancer. Int. J. Cancer 2009, 127, 257–268. [Google Scholar] [CrossRef] [Green Version]
- Cheng, L.; Yan, B.; Chen, K.; Jiang, Z.; Zhou, C.; Cao, J.; Qian, W.; Li, J.; Sun, L.; Ma, J.; et al. Resveratrol-Induced Downreg-ulation of NAF-1 Enhances the Sensitivity of Pancreatic Cancer Cells to Gemcitabine via the ROS/Nrf2 Signaling Pathways. Available online: https://www.hindawi.com/journals/omcl/2018/9482018/ (accessed on 17 December 2020).
- Jiang, Z.; Chen, X.; Chen, K.; Sun, L.; Gao, L.; Zhou, C.; Lei, M.; Duan, W.; Wang, Z.; Ma, Q.; et al. YAP Inhibition by Resveratrol via Activation of AMPK Enhances the Sensitivity of Pancreatic Cancer Cells to Gemcitabine. Nutrients 2016, 8, 546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Tian, W.; Yang, L.; Zhang, Q.; Zhu, M.; Liu, Y.; Li, J.; Yang, L.; Liu, J.; Shen, Y.; et al. Gemcitabine potentiates anti-tumor effect of resveratrol on pancreatic cancer via down-regulation of VEGF-B. J. Cancer Res. Clin. Oncol. 2021, 147, 93–103. [Google Scholar] [CrossRef]
- Cho, C.; Yang, C.; Wu, C.; Ho, J.; Yu, C.; Wu, S.; Yu, D. The modulation study of multiple drug resistance in bladder cancer by curcumin and resveratrol. Oncol. Lett. 2019, 18, 6869–6876. [Google Scholar] [CrossRef]
- A Sprowl, J.; Reed, K.; Armstrong, S.R.; Lanner, C.; Guo, B.; Kalatskaya, I.; Stein, L.; Hembruff, S.L.; Tam, A.; Parissenti, A.M. Alterations in tumor necrosis factor signaling pathways are associated with cytotoxicity and resistance to taxanes: a study in isogenic resistant tumor cells. Breast Cancer Res. 2012, 14, R2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinod, B.S.; Nair, H.H.; Vijayakurup, V.; Shabna, A.; Shah, S.; Krishna, A.; Pillai, K.S.; Thankachan, S.; Anto, R.J. Resveratrol chemosensitizes HER-2-overexpressing breast cancer cells to docetaxel chemoresistance by inhibiting docetaxel-mediated activation of HER-2–Akt axis. Cell Death Discov. 2015, 1, 15061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- A Sprouse, A.; Herbert, B.-S. Resveratrol augments paclitaxel treatment in MDA-MB-231 and paclitaxel-resistant MDA-MB-231 breast cancer cells. Anticancer Res. 2014, 34, 5363–5374. [Google Scholar]
- Jazirehi, A.R.; Bonavida, B. Resveratrol modifies the expression of apoptotic regulatory proteins and sensitizes non-Hodgkin’s lymphoma and multiple myeloma cell lines to paclitaxel-induced apoptosis. Mol. Cancer Ther. 2004, 3, 71–84. [Google Scholar]
- Cipolletti, M.; Montalesi, E.; Nuzzo, M.T.; Fiocchetti, M.; Ascenzi, P.; Marino, M. Potentiation of paclitaxel effect by resveratrol in human breast cancer cells by counteracting the 17β-estradiol/estrogen receptor α/neuroglobin pathway. J. Cell. Physiol. 2019, 234, 3147–3157. [Google Scholar] [CrossRef]
- Öztürk, Y.; Günaydın, C.; Yalçın, F.; Nazıroğlu, M.; Braidy, N. Resveratrol Enhances Apoptotic and Oxidant Effects of Paclitaxel through TRPM2 Channel Activation in DBTRG Glioblastoma Cells. Oxidative Med. Cell. Longev. 2019, 2019, 1–13. [Google Scholar] [CrossRef]
- Tong, C.W.S.; Wu, M.; Cho, W.C.S.; To, K.K.W. Recent Advances in the Treatment of Breast Cancer. Front. Oncol. 2018, 8, 227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nevedomskaya, E.; Baumgart, S.J.; Haendler, B. Recent Advances in Prostate Cancer Treatment and Drug Discovery. Int. J. Mol. Sci. 2018, 19, 1359. [Google Scholar] [CrossRef] [Green Version]
- De Amicis, F.; Chimento, A.; Montalto, F.I.; Casaburi, I.; Sirianni, R.; Pezzi, V. Steroid Receptor Signallings as Targets for Resveratrol Actions in Breast and Prostate Cancer. Int. J. Mol. Sci. 2019, 20, 1087. [Google Scholar] [CrossRef] [Green Version]
- Levenson, A.S.; Kliakhandler, I.L.; Svoboda, K.M.; Pease, K.M.; A Kaiser, S.; E Ward, I.J.; Jordan, V.C. Molecular classification of selective oestrogen receptor modulators on the basis of gene expression profiles of breast cancer cells expressing oestrogen receptor α. Br. J. Cancer 2002, 87, 449–456. [Google Scholar] [CrossRef] [Green Version]
- Gehm, B.D.; McAndrews, J.M.; Chien, P.-Y.; Jameson, J.L. Resveratrol, a polyphenolic compound found in grapes and wine, is an agonist for the estrogen receptor. Proc. Natl. Acad. Sci. USA 1997, 94, 14138–14143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, R.; Serrero, G. Resveratrol, a natural product derived from grape, exhibits antiestrogenic activity and inhibits the growth of human breast cancer cells. J. Cell. Physiol. 1999, 179, 297–304. [Google Scholar] [CrossRef]
- De Amicis, F.; Giordano, F.; Vivacqua, A.; Pellegrino, M.; Panno, M.L.; Tramontano, D.; Fuqua, S.A.W.; Andò, S. Resveratrol, through NF-Y/p53/Sin3/HDAC1 complex phosphorylation, inhibits estrogen receptor α gene expression via p38 MAPK /CK2 signaling in human breast cancer cells. FASEB J. 2011, 25, 3695–3707. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, G.F.S.; Silva, G.D.B.; Pavan, A.R.; Chiba, D.E.; Chin, C.M.; Dos Santos, J.L. Epigenetic Regulatory Mechanisms Induced by Resveratrol. Nutrients 2017, 9, 1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirzapur, P.; Khazaei, M.R.; Moradi, M.T.; Khazaei, M. Apoptosis induction in human breast cancer cell lines by synergic effect of raloxifene and resveratrol through increasing proapoptotic genes. Life Sci. 2018, 205, 45–53. [Google Scholar] [CrossRef]
- Kuwajerwala, N.; Cifuentes, E.; Gautam, S.; Menon, M.; Barrack, E.R.; Reddy, G.P.V. Resveratrol induces prostate cancer cell entry into s phase and inhibits DNA synthesis. Cancer Res. 2002, 62, 2488–2492. [Google Scholar]
- Jang, Y.-G.; Go, R.-E.; Hwang, K.-A.; Choi, K.-C. Resveratrol inhibits DHT-induced progression of prostate cancer cell line through interfering with the AR and CXCR4 pathway. J. Steroid Biochem. Mol. Biol. 2019, 192, 105406. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Kandagatla, P.; Singareddy, R.; Kropinski, A.; Sheng, S.; Cher, M.L.; Chinni, S.R. Androgens Induce Functional CXCR4 through ERG Factor Expression in TMPRSS2-ERG Fusion-Positive Prostate Cancer Cells. Transl. Oncol. 2010, 3, 195–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Havens, A.M.; Jung, Y.; Sun, Y.X.; Wang, J.; Shah, R.B.; Bühring, H.J.; Pienta, K.; Taichman, R.S. The role of sialomucin CD164 (MGC-24v or endolyn) in prostate cancer metastasis. BMC Cancer 2006, 6, 195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, M.; Tian, H.; Lin, S.; Mo, J.; Li, Z.; Chen, X.; Liu, J. Resveratrol inhibits proliferation and promotes apoptosis via the androgen receptor splicing variant 7 and PI3K/AKT signaling pathway in LNCaP prostate cancer cells. Oncol. Lett. 2020, 20, 1. [Google Scholar] [CrossRef] [PubMed]
- Samuel, V.P.; Gupta, G.; Dahiya, R.; Jain, D.A.; Mishra, A.; Dua, K. Current Update on Preclinical and Clinical Studies of Resveratrol, a Naturally Occurring Phenolic Compound. Crit. Rev. Eukaryot. Gene Expr. 2019, 29, 529–537. [Google Scholar] [CrossRef]
- Franciosoa, A.; Mastromarino, P.; Masci, A.; D’Erme, M.; Mosca, L. Chemistry, Stability and Bioavailability of Resveratrol. Med. Chem. 2014, 10, 237–245. [Google Scholar] [CrossRef]
- Kashyap, D.; Tuli, H.S.; Yerer, M.B.; Sharma, A.; Sak, K.; Srivastava, S.; Pandey, A.; Garg, V.K.; Sethi, G.; Bishayee, A. Natural product-based nanoformulations for cancer therapy: Opportunities and challenges. Semin. Cancer Biol. 2019. [Google Scholar] [CrossRef]
- Zhao, Y.; Huan, M.-L.; Liu, M.; Cheng, Y.; Sun, Y.; Cui, H.; Liu, D.-Z.; Mei, Q.-B.; Zhou, S.-Y. Doxorubicin and resveratrol co-delivery nanoparticle to overcome doxorubicin resistance. Sci. Rep. 2016, 6, 35267. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Guo, F.; Xu, H.; Liang, W.; Wang, C.; Yang, X.-D. Combination Therapy using Co-encapsulated Resveratrol and Paclitaxel in Liposomes for Drug Resistance Reversal in Breast Cancer Cells in vivo. Sci. Rep. 2016, 6, 22390. [Google Scholar] [CrossRef]
- Berretta, M.; Rinaldi, L.; Taibi, R.; Tralongo, P.; Fulvi, A.; Montesarchio, V.; Madeddu, G.; Magistri, P.; Bimonte, S.; Trovò, M.; et al. Physician Attitudes and Perceptions of Complementary and Alternative Medicine (CAM): A Multicentre Italian Study. Front. Oncol. 2020, 10. [Google Scholar] [CrossRef]
- Berretta, M.; Di Francia, R.; Cazzavillan, S.; Rossi, P.; Scorba, A.; Sciacca, P.G.; Santagà, D. Integrative Medicine in Oncology, Milan 19th October 2019. World Cancer Res. J. 2019, 6. [Google Scholar] [CrossRef]
- İnci, H.; İnci, F. Complementary and Alternative Medicine Awareness in Cancer Patients Receiving Chemotherapy. World Cancer Res. J. 2020, 7. [Google Scholar] [CrossRef]
- Berretta, M.; Della Pepa, C.; Tralongo, P.; Fulvi, A.; Martellotta, F.; Lleshi, A.; Nasti, G.; Fisichella, R.; Romano, C.; De Divitiis, C.; et al. Use of Complementary and Alternative Medicine (CAM) in cancer patients: An Italian multicenter survey. Oncotarget 2017, 8, 24401–24414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dehghan, M.; Ghaedi Heidari, F.; Malakoutikhah, A.; Mokhtarabadi, S. Complementary and Alternative Medicine Usage and Its Determinant Factors among Iranian Patients with Cancer. World Cancer Res. J. 2019, 6. [Google Scholar] [CrossRef]
- Antiga, E.; Bonciolini, V.; Volpi, W.; Del Bianco, E.; Caproni, M. Oral Curcumin (Meriva) Is Effective as an Adjuvant Treatment and Is Able to Reduce IL-22 Serum Levels in Patients with Psoriasis Vulgaris. BioMed Res. Int. 2015, 2015, 1–7. [Google Scholar] [CrossRef]
- Stohs, S.J.; Chen, O.; Ray, S.D.; Ji, J.; Bucci, L.R.; Preuss, H.G. Highly Bioavailable Forms of Curcumin and Promising Avenues for Curcumin-Based Research and Application: A Review. Antioxidants 2020, 25, 1397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quagliariello, V.; Vecchione, R.; Coppola, C.; Di Cicco, C.; De Capua, A.; Piscopo, G.; Paciello, R.; Narciso, V.; Formisano, C.; Taglialatela-Scafati, O.; et al. Cardioprotective Effects of Nanoemulsions Loaded with Anti-Inflammatory Nutraceuticals against Doxorubicin-Induced Cardiotoxicity. Nutrients 2018, 10, 1304. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Drug Class | Drug | Cancer Model | Effect in Combination with Resveratrol | References |
|---|---|---|---|---|
| Alkylating agents | Temozolomide | SHG44 GBM cell line In vivo xenograft mouse models | Additive effect by ROS-dependent AMPK-TSC-mTOR signaling pathway Reduction of tumor growth | [53] |
| GBM-initiating cells (GICs) | Activation of the DNA double strands pATM/pART/p53 pathway | [54] | ||
| GBM cells | By downregulating MGMT via the NF-KB dependent pathway and via the repression of the activated Wnt signaling pathway | [56,57] | ||
| Glioma cells | Mitotic catastrophe and senescence | [58] | ||
| Platinum compounds | Cisplatin | Non-small lung cancer cells (NSCLC) H838 and H520 | Depolarization of MMP and apoptosis | [65] |
| A549 lung carcinoma cells | Decreases the phosphorylation of AKT thus inducing autophagy | [67] | ||
| Hepatoma cells C3A and SMCC7721 | Reduction of transporter ASCT2 imbalance in the redox homeostasis | [69] | ||
| A2780, OVCAR-3 CaOv-4 | Proapoptotic effect | [70,71,72,73] | ||
| EAC tumor-bearing mice | Activation of proapoptotic family members | [77] | ||
| Melanoma models | Upregulation of connexin 43 expression | [78] | ||
| Oxaliplatin | Colon cancer cell line | Inhibition of cell growth | [74] | |
| Colon cancer cell line Caco2 | Interfering with caspase-3-activation, PARP cleavage, and depolarization of mitochondrial membrane potential | [76] | ||
| HCT116 cells | Reestablishment of surviving protein expression | [75] | ||
| Anthracyclines | Doxorubicin (Adriamycin) | MDA-MB-231 and MCF-7/Adr breast cancer cells MCF-7/Adr cell xenograft model | Decreases the expression of MDR1 and MRP1; decreases P-gp ATPase activity Reduces tumor volume and expression levels of MDR1 and MRP1 | [81] |
| MCF-7 and MDA-MB-231 cell lines Ehrlich ascitic carcinoma cells-bearing mice | Growth inhibition, decreased clonogenic potential, inhibition of inflammatory response, induction of apoptosis. Reduced tumor volume, increased lifespan | [82] | ||
| SGC7901 gastric cancer and SGC7901/DOX in vivo xenograft tumor model | Suppressing ETM via modulation of PTEN/Akt signaling pathway | [84] | ||
| MCF7/Adr breast cancer cells | Regulation of SIRT1/ β-catenin pathway | [85] | ||
| MCF-7/Adr breast cancer cells | Targeting miR-122-5p, thus regulating apoptosis-inhibitory proteins Bcl-2 and CDKs | [86] | ||
| Spheroids of PANC-1 pancreatic cells | Reduced P-glycoprotein mediated efflux of the drug | [87] | ||
| HCT116 colon cancer cell lines | Increased expression of Bax gene and blocked the efflux activity of p-gp | [88] | ||
| Caco-2 colorectal and CEM/ADR5000 T lymphoblastoid cell line | Inhibition of the ABC transporters, metabolic enzyme GST and CYP3A4 activity | [89] | ||
| Antimetabolites | 5-Fluorouracil | Colorectal cancer cells | Inhibiting pAkt signaling pathway Reduction of TNF-β-induced epithelial-mesenchymal transition and downregulation of NF-KB | [94] [98,99] |
| Colorectal cancer cell lines | Imbalance in redox homeostasis linked to inhibition of Akt and STAT3 levels | [95] | ||
| B16 murine melanoma cells B16-tumors model in mice | Regulation of levels of AMPK, COX-2, VASP and VEGF Reduces tumor growth | [96] | ||
| Murine model of liver | Reduces tumor growth | [97] | ||
| Gemcitabine | Advanced pancreatic cancer (PC) | Downregulation of SREBP1 | [101] | |
| PaCa xenografts in nude mice | Reduces tumor growth | [102] | ||
| Pancreatic cancer cells | ROS accumulation, activation of Nrf-2 signaling, suppression of NAF-1 | [103] | ||
| Pancreatic cancer cells | Downregulation of YAP | [104] | ||
| Pancreatic cancer cells and in vivo model | Downregulation of VEGF-B and GSK3β | [105] | ||
| Human bladder cancer cell line | Modulation of ABCC2, DCK, TK1 and TK2 function and increased PARP cleavage and apoptosis | [106] | ||
| Mitotic Inhibitors | Docetaxel | HER-2-overexpressing SK-BR-3 breast cancer cells | Blocks the enhancement and activation of HER-2, in addition to blocking AKT pathway | [108] |
| Paclitaxel | MDA-MB-231 breast cancer cell line | Induction of cell senescence and apoptosis | [109] | |
| Paclitaxel-resistant non-Hodgkin’s lymphoma and multiple myeloma | Decreases Bcl-x, upregulates Bax and Apaf-1 expression. | [110] | ||
| MCF-7 and T47D ERα positive breast cancer cells | Decreases NGB levels, via interference with E2/Erα pathway | [111] | ||
| DBTRG glioblastoma cell lines | Stimulation and activation of the oxidative sensitive TRPM2 channel | [112] | ||
| Endocrine therapy | Tamoxifen | MCF-7 Tamoxifen-resistant cancer cells | Transcriptional inhibition of ERα via blockade of cell cycle, activation of p38 MAPK/CK2 signaling and induction of p53 | [119] |
| 4-hydroxy -tamoxifen (4-OHT) | MCF-7-TR breast cancer cells | Inhibition of ER transcription via p38 MAPK/casein kinase II signaling, p53, binding with the transcription nuclear factor Y (NF-Y) to the ER proximal promoter | [119] | |
| Raloxifene | MCF7 cells estrogen-receptor positive | Increases the Bcl2/Bax ratio and expression of p53 and caspases 3,8 | [121] | |
| Bicalutamide | Prostate cancer | Via downregulation of AKT signaling pathway, suppresses CXCR4 expression | [123,125] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cocetta, V.; Quagliariello, V.; Fiorica, F.; Berretta, M.; Montopoli, M. Resveratrol as Chemosensitizer Agent: State of Art and Future Perspectives. Int. J. Mol. Sci. 2021, 22, 2049. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22042049
Cocetta V, Quagliariello V, Fiorica F, Berretta M, Montopoli M. Resveratrol as Chemosensitizer Agent: State of Art and Future Perspectives. International Journal of Molecular Sciences. 2021; 22(4):2049. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22042049
Chicago/Turabian StyleCocetta, Veronica, Vincenzo Quagliariello, Francesco Fiorica, Massimiliano Berretta, and Monica Montopoli. 2021. "Resveratrol as Chemosensitizer Agent: State of Art and Future Perspectives" International Journal of Molecular Sciences 22, no. 4: 2049. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22042049