
New Gene Markers Expressed in Porcine Oviductal Epithelial Cells Cultured Primary In Vitro Are Involved in Ontological Groups Representing Physiological Processes of Porcine Oocytes

 , , , , , , , , , , , , and
, , , , , , , , , , , , and
Abstract
:1. Introduction
2. Results
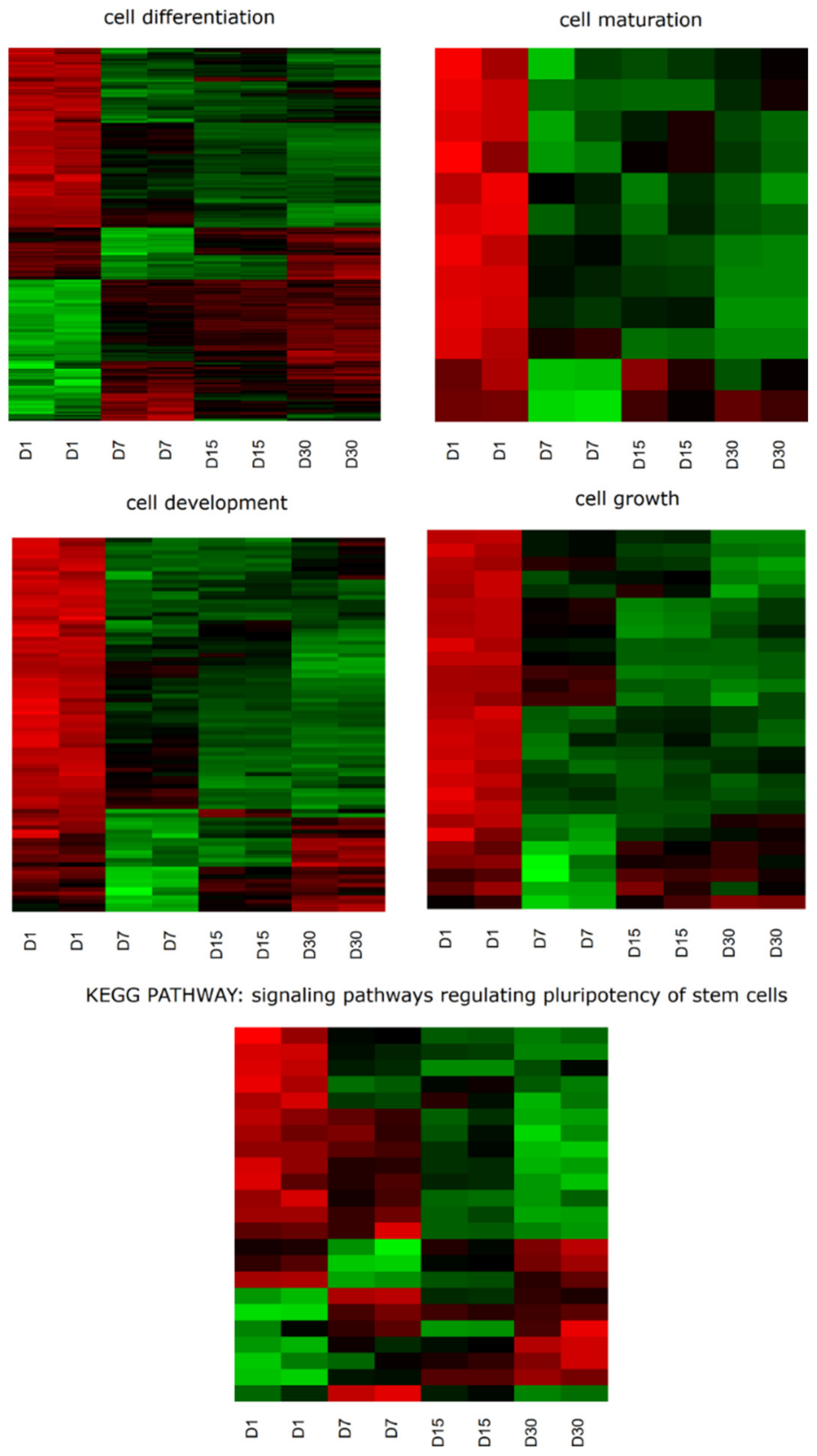
2.1. Whole Transcriptome Profiling
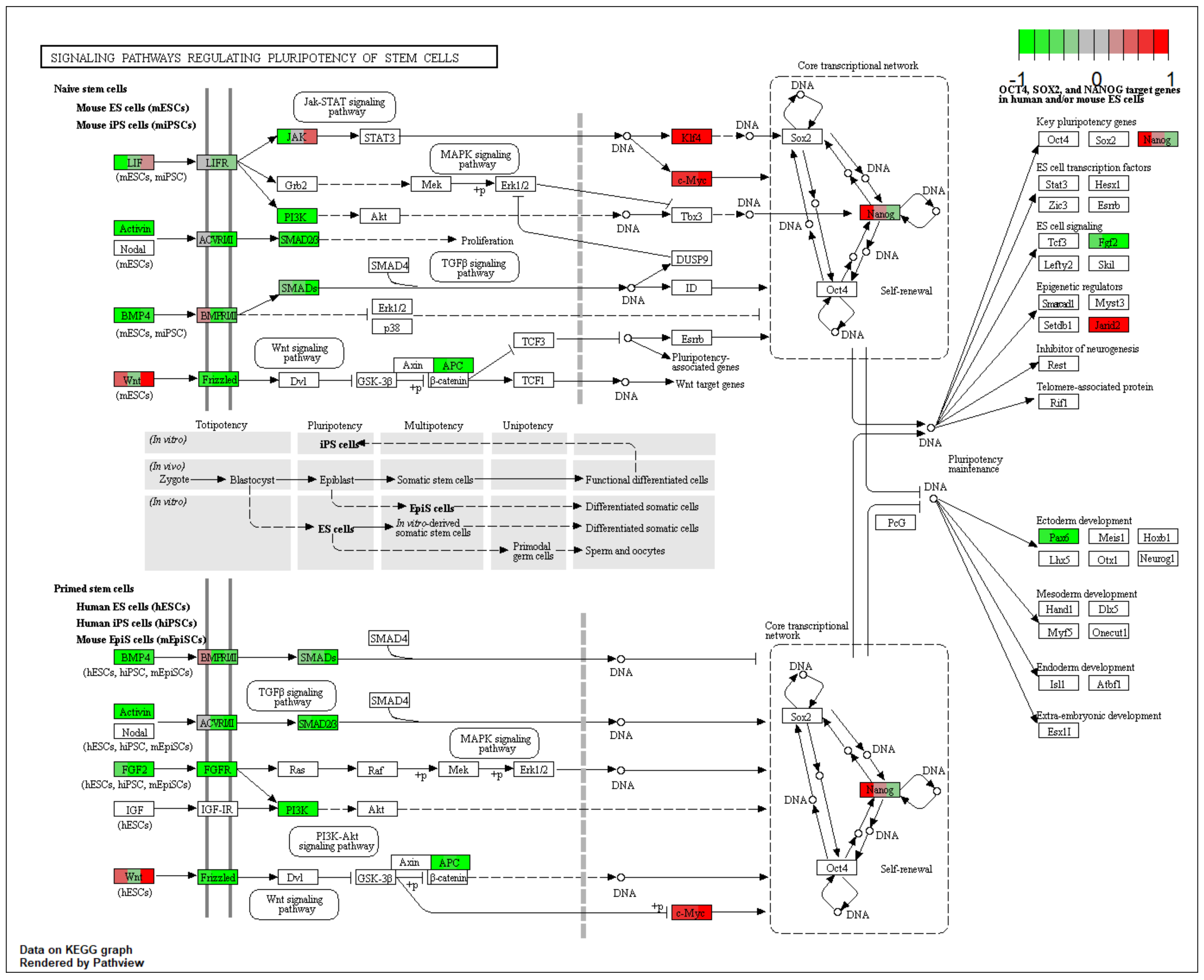
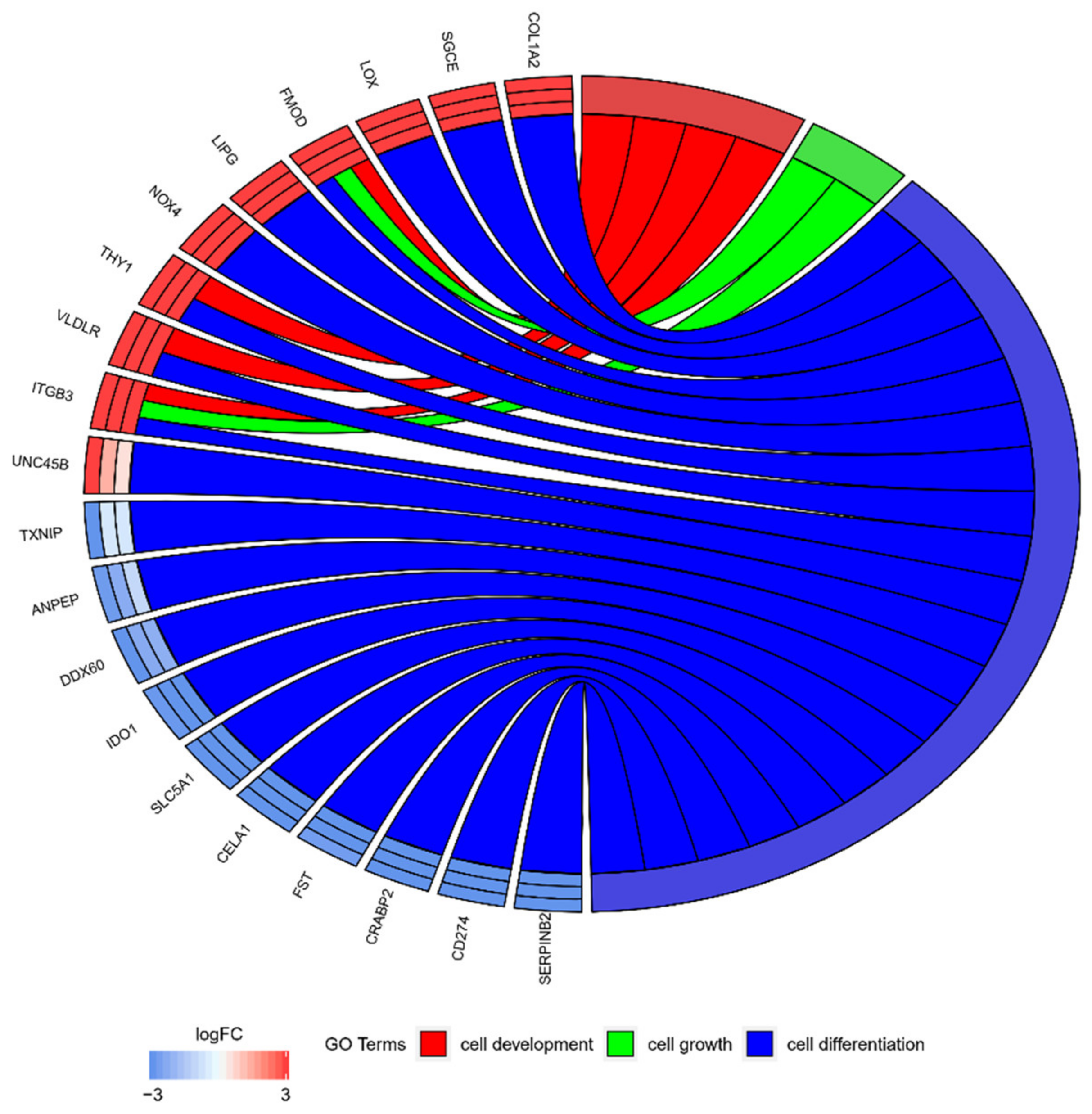
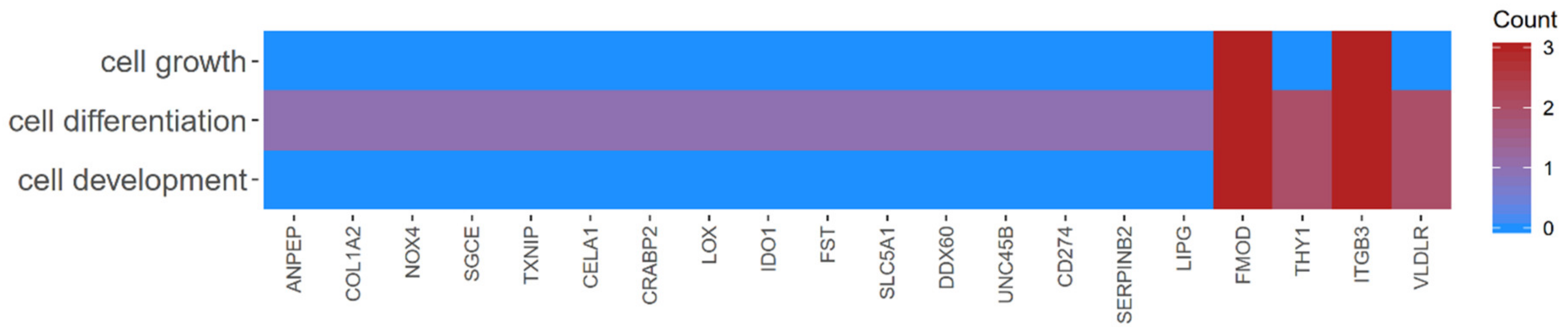
2.2. GO BP Terms and KEGG Pathway Analysis
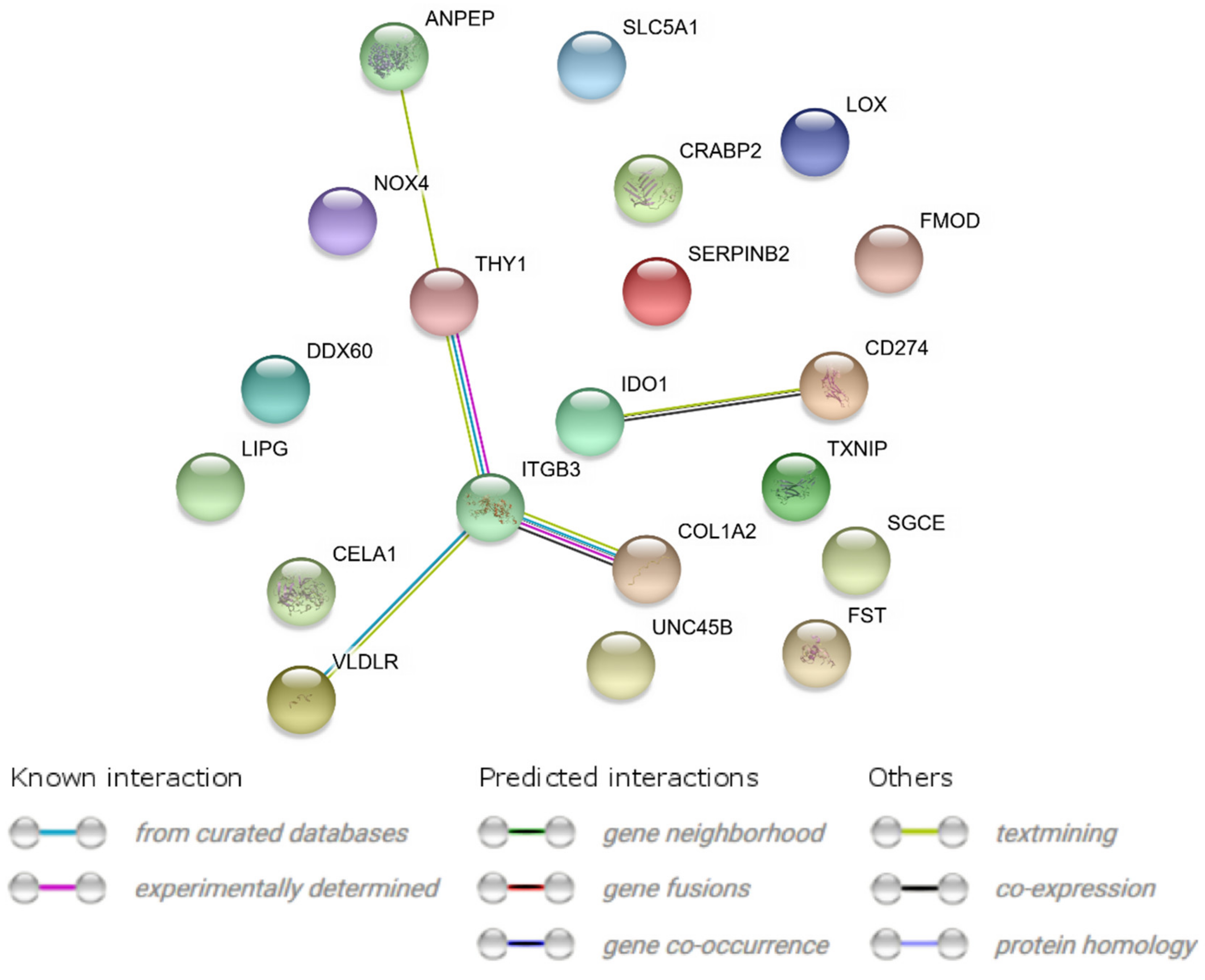
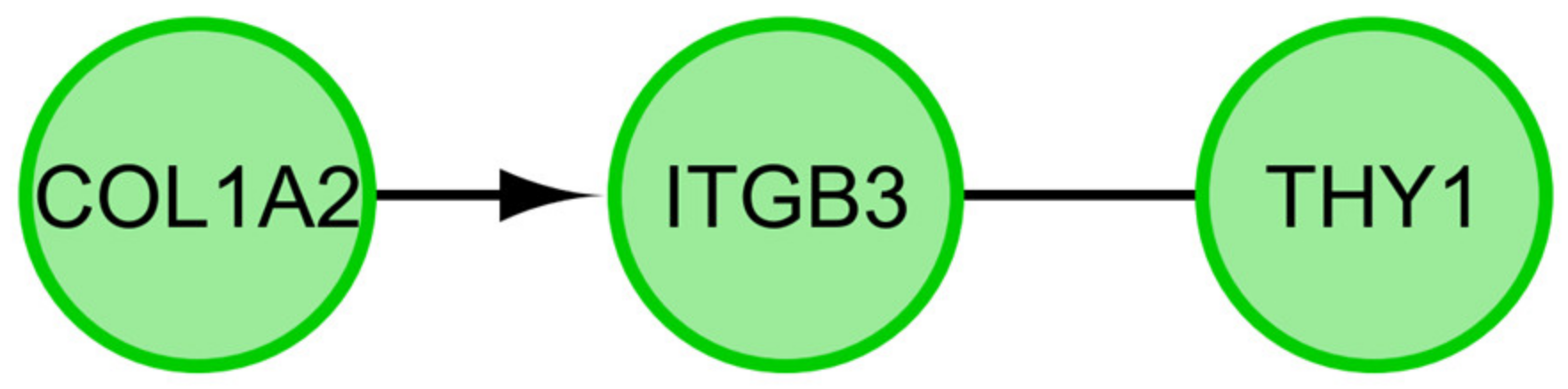
2.3. Interaction Network Analyses
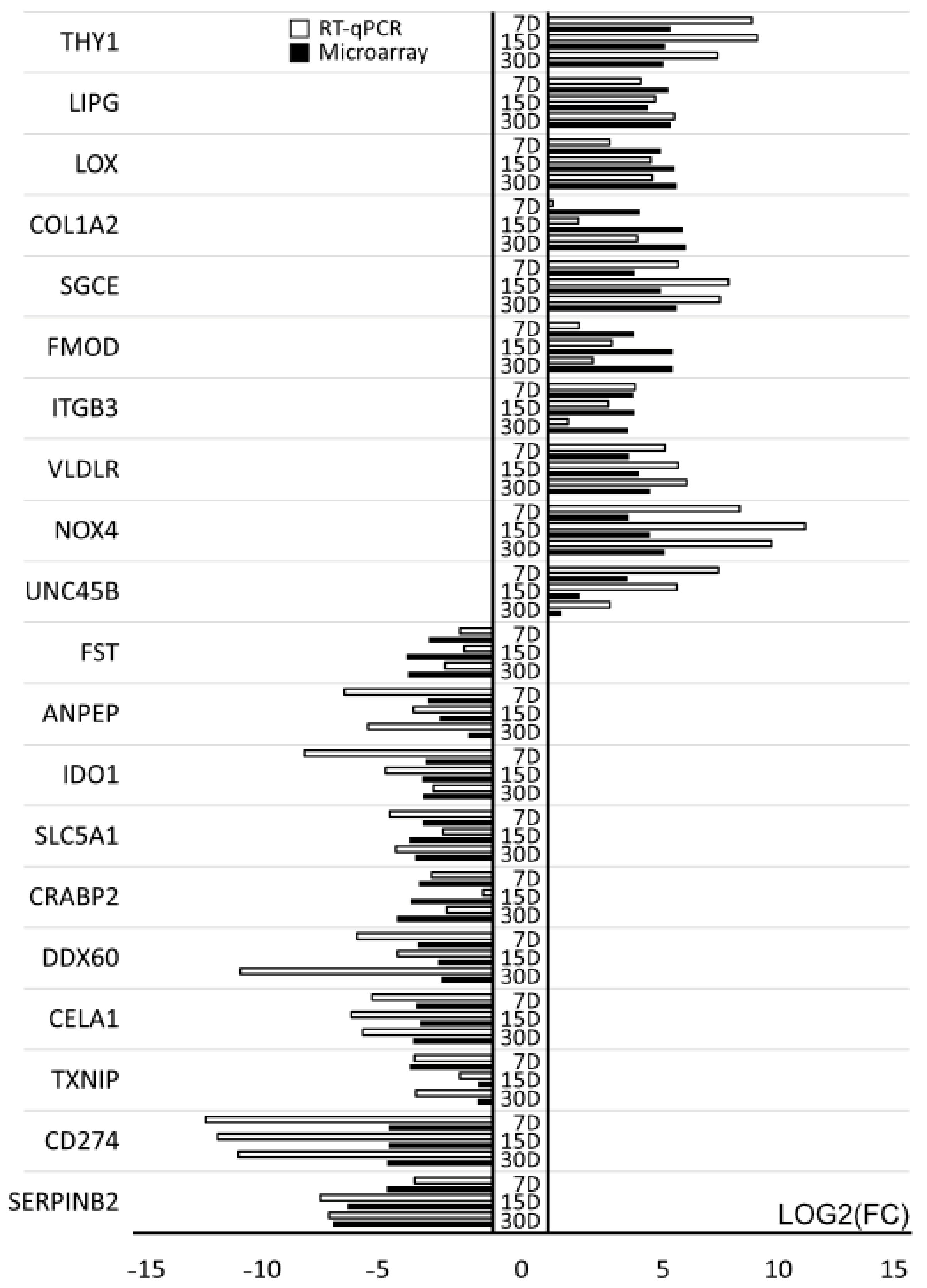
2.4. Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR) Analysis
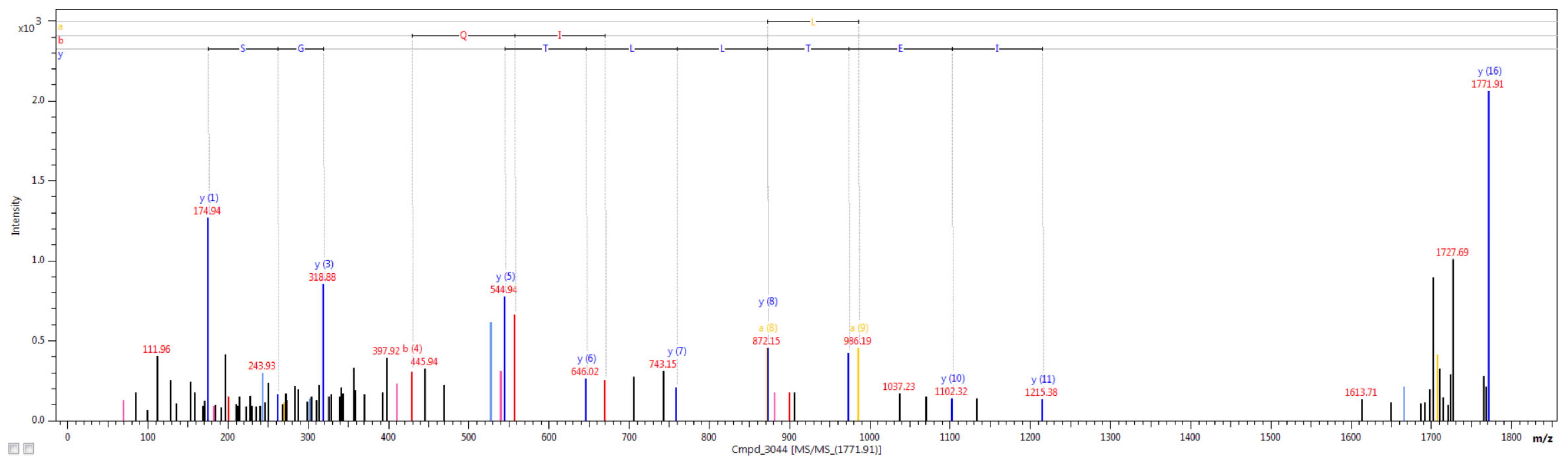
2.5. Nano LC-MALDI-TOF/TOF MS/MS Analysis
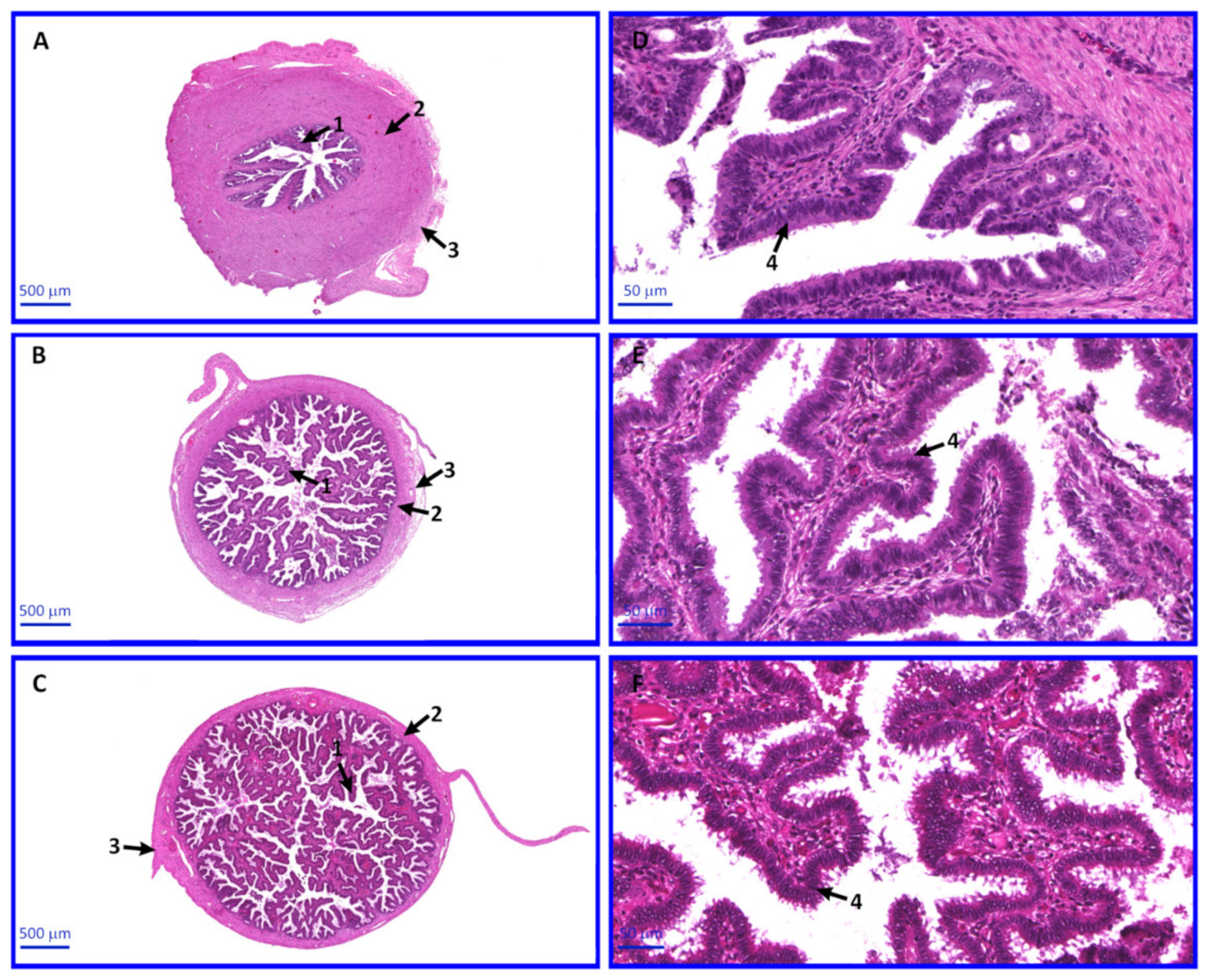
2.6. Morphology of OECs
3. Discussion
4. Materials and Methods
4.1. Animals-Anatomical Structure and Collection of Oviducts
4.2. OEC Selection and Culture
4.3. Hematoxylin and Eosin (H&E) Staining
4.4. RNA Extraction from Porcine OECs
4.5. Microarray Expression Analysis
4.6. Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR) Analysis
4.7. Bioinformatics and Statistical Analysis
4.8. Nano Liquid Chromatography (LC)-MALDI-TOF/TOF MS/MS Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rybska, M.; Knap, S.; Jankowski, M.; Jeseta, M.; Bukowska, D.; Antosik, P.; Nowicki, M.; Zabel, M.; Kempisty, B.; Jaśkowski, J.M. Cytoplasmic and nuclear maturation of oocytes in mammals–living in the shadow of cells developmental capability. Med. J. Cell Biol. 2018, 6, 13–17. [Google Scholar] [CrossRef] [Green Version]
- Kranc, W.; Jankowski, M.; Budna, J.; Celichowski, P.; Khozmi, R.; Bryja, A.; Borys, S.; Dyszkiewicz-Konwińska, M.; Jeseta, M.; Magas, M.; et al. Amino acids metabolism and degradation is regulated during porcine oviductal epithelial cells (OECs) primary culture in vitro–a signaling pathways activation approach. Med. J. Cell Biol. 2018, 6, 18–26. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-M.; Park, J.-E.; Yoo, I.; Han, J.; Kim, N.; Lim, W.-J.; Cho, E.-S.; Choi, B.; Choi, S.; Kim, T.-H.; et al. Integrated transcriptomes throughout swine oestrous cycle reveal dynamic changes in reproductive tissues interacting networks. Sci. Rep. 2018, 8, 5436. [Google Scholar] [CrossRef] [PubMed]
- Budna-Tukan, J.; Światły-Błaszkiewicz, A.; Celichowski, P.; Kałużna, S.; Konwerska, A.; Sujka-Kordowska, P.; Jankowski, M.; Kulus, M.; Jeseta, M.; Piotrowska-Kempisty, H.; et al. “Biological Adhesion” is a Significantly Regulated Molecular Process during Long-Term Primary In vitro Culture of Oviductal Epithelial Cells (Oecs): A Transcriptomic and Proteomic Study. Int. J. Mol. Sci. 2019, 20, 3387. [Google Scholar] [CrossRef] [Green Version]
- Kulus, M.; Józkowiak, M.; Kulus, J.; Popis, M.; Borowiec, B.; Stefańska, K.; Celichowski, P.; Nawrocki, M.J.; Bukowska, D.; Brüssow, K.P.; et al. “Cell cycle process”, “cell division” and “cell proliferation” belong to ontology groups highly regulated during long–term culture of porcine oviductal epithelial cells. Med. J. Cell Biol. 2019, 7, 15–24. [Google Scholar] [CrossRef]
- Kulus, M.; Kulus, J.; Popis, M.; Borowiec, B.; Stefańska, K.; Celichowski, P.; Nawrocki, M.J.; Brüssow, K.P.; Kempisty, B.; Jeseta, M.; et al. “Cell cycle” and ’cell death’- Related genes are differentially expressed during long-Term in vitro real-time cultivation of porcine oviductal epithelial cells. Med. J. Cell Biol. 2019, 7, 90–99. [Google Scholar] [CrossRef] [Green Version]
- Kossowska-Tomaszczuk, K.; De Geyter, C.; De Geyter, M.; Martin, I.; Holzgreve, W.; Scherberich, A.; Zhang, H. The Multipotency of Luteinizing Granulosa Cells Collected from Mature Ovarian Follicles. Stem Cells 2009, 27, 210–219. [Google Scholar] [CrossRef]
- Jankowski, M.; Dyszkiewicz-Konwińska, M.; Budna, J.; Huang, Y.; Knap, S.; Bryja, A.; Borys, S.; Kranc, W.; Jeseta, M.; Magas, M.; et al. Does migrative and proliferative capability of epithelial cells reflect cellular developmental competence? Running title: Epithelial cells in vitro development. Med. J. Cell Biol. 2018, 6, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Jankowski, M.; Dyszkiewicz-Konwińska, M.; Budna, J.; Knap, S.; Bryja, A.; Borys, S.; Kranc, W.; Magas, M.; Jeseta, M.; Bukowska, D.; et al. The differentiation and transdifferentiation of epithelial cells in vitro–is it a new strategy in regenerative biomedicine? Med. J. Cell Biol. 2018, 6, 27–32. [Google Scholar] [CrossRef] [Green Version]
- Rybska, M.; Knap, S.; Jankowski, M.; Jeseta, M.; Bukowska, D.; Antosik, P.; Nowicki, M.; Zabel, M.; Kempisty, B.; Jaśkowski, J.M. Characteristic of factors influencing the proper course of folliculogenesis in mammals. Med. J. Cell Biol. 2018, 6, 33–38. [Google Scholar] [CrossRef] [Green Version]
- Nawrocki, M.J.; Budna, J.; Celichowski, P.; Khozmi, R.; Bryja, A.; Kranc, W.; Borys, S.; Ciesiółka, S.; Knap, S.; Jeseta, M.; et al. Analysis of fructose and mannose–regulatory peptides signaling pathway in porcine epithelial oviductal cells (OECs) primary cultured long-term in vitro. Adv. Cell Biol. 2017, 5. [Google Scholar] [CrossRef] [Green Version]
- Coy, P.; García-Vázquez, F.A.; Visconti, P.E.; Avilés, M. Roles of the oviduct in mammalian fertilization. Reproduction 2012, 144, 649–660. [Google Scholar] [CrossRef] [Green Version]
- Varga, I.; Miko, M.; Kachlík, D.; Žišková, M.; Danihel, L.; Babál, P. How many cell types form the epithelial lining of the human uterine tubes? Revision of the histological nomenclature of the human tubal epithelium. Ann. Anat. Anat. Anz. 2019, 224, 73–80. [Google Scholar] [CrossRef]
- López-Úbeda, R.; García-Vázquez, F.A.; Romar, R.; Gadea, J.; Muñoz, M.; Hunter, R.H.F.; Coy, P. Oviductal Transcriptome Is Modified after Insemination during Spontaneous Ovulation in the Sow. PLoS ONE 2015, 10, e0130128. [Google Scholar]
- López-Úbeda, R.; Muñoz, M.; Vieira, L.; Hunter, R.H.F.; Coy, P.; Canovas, S. The oviductal transcriptome is influenced by a local ovarian effect in the sow. J. Ovarian Res. 2016, 9, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.H.; Oh, H.J.; Kim, M.J.; Setyawan, E.M.N.; Lee, B.C. Interaction of the EGFR signaling pathway with porcine cumulus oocyte complexes and oviduct cells in a coculture system. J. Cell. Physiol. 2018, 234, 4030–4043. [Google Scholar] [CrossRef] [PubMed]
- Moncrieff, L.; Kocherova, I.; Bryja, A.; Kranc, W.; Perek, J.; Kulus, M.; Jeseta, M.; Dompe, C.; Hutchings, G.; Janowicz, K.; et al. Transcriptomic profile of genes encoding proteins responsible for regulation of cells differentiation and neurogenesis in vivo and in vitro–an oocyte model approach. Med. J. Cell Biol. 2020, 8, 1–11. [Google Scholar] [CrossRef]
- Jiménez-Marín, A.; Yubero, N.; Esteso, G.; Moreno, A.; de las Mulas, J.M.; Morera, L.; Llanes, D.; Barbancho, M.; Garrido, J.J. Molecular characterization and expression analysis of the gene coding for the porcine β3 integrin subunit (CD61). Gene 2008, 408, 9–17. [Google Scholar] [CrossRef]
- Tang, Q.; Zhang, Y.; Xu, Y.; He, L.; Dai, C.; Sun, P. Up-regulation of integrin β3 expression in porcine vascular endothelial cells cultured in vitro by classical swine fever virus. Vet. Immunol. Immunopathol. 2010, 133, 237–242. [Google Scholar] [CrossRef]
- Li, W.; Wang, G.; Liang, W.; Kang, K.; Guo, K.; Zhang, Y. Integrin β3 Is Required in Infection and Proliferation of Classical Swine Fever Virus. PLoS ONE 2014, 9, e110911. [Google Scholar] [CrossRef] [Green Version]
- Kulus, M.; Sujka-kordowska, P.; Konwerska, A.; Celichowski, P.; Kranc, W.; Kulus, J.; Piotrowska-Kempisty, H.; Antosik, P. New Molecular Markers Involved in Regulation of Ovarian Granulosa Cell Morphogenesis, Development and Di ff erentiation during Short-Term Primary In vitro Culture—Transcriptomic and Histochemical Study Based on Ovaries and Individual Separated Follicle. Int. J. Mol. Sci. 2019, 20, 3966. [Google Scholar] [CrossRef] [Green Version]
- Kulus, M.; Kranc, W.; Sujka-Kordowska, P.; Mozdziak, P.; Jankowski, M.; Konwerska, A.; Kulus, J.; Bukowska, D.; Skowroński, M.; Piotrowska-Kempisty, H.; et al. The processes of cellular growth, aging, and programmed cell death are involved in lifespan of ovarian granulosa cells during short-term IVC–Study based on animal model. Theriogenology 2020, 148, 76–88. [Google Scholar] [CrossRef] [PubMed]
- Uenishi, H.; Eguchi-Ogawa, T.; Shinkai, H.; Okumura, N.; Suzuki, K.; Toki, D.; Hamasima, N.; Awata, T. PEDE (Pig EST Data Explorer) has been expanded into Pig Expression Data Explorer, including 10 147 porcine full-length cDNA sequences. Nucleic Acids Res. 2007, 35, D650–D653. [Google Scholar] [CrossRef] [PubMed]
- Tormin, A.; Brune, J.C.; Olsson, E.; Valcich, J.; Neuman, U.; Olofsson, T.; Jacobsen, S.-E.; Scheding, S. Characterization of bone marrow-derived mesenchymal stromal cells (MSC) based on gene expression profiling of functionally defined MSC subsets. Cytotherapy 2009, 11, 114–128. [Google Scholar] [CrossRef]
- Zheng, Z.; Jian, J.; Velasco, O.; Hsu, C.; Zhang, K.; Levin, A.; Murphy, M.; Zhang, X.; Ting, K.; Soo, C. Fibromodulin Enhances Angiogenesis during Cutaneous Wound Healing. Plast. Reconstr. Surg. Glob. Open 2014, 2, e275. [Google Scholar] [CrossRef] [PubMed]
- Tiebel, O.; Oka, K.; Robinson, K.; Sullivan, M.; Martinez, J.; Nakamuta, M.; Ishimura-Oka, K.; Chan, L. Mouse very low-density lipoprotein receptor (VLDLR): Gene structure, tissue-specific expression and dietary and developmental regulation. Atherosclerosis 1999, 145, 239–251. [Google Scholar] [CrossRef]
- Zheng, Y.; He, Y.; An, J.; Qin, J.; Wang, Y.; Zhang, Y.; Tian, X.; Zeng, W. THY1 is a surface marker of porcine gonocytes. Reprod. Fertil. Dev. 2014, 26, 533. [Google Scholar] [CrossRef]
- Orsi, P.R.; Landim-Alvarenga, F.C.; Justulin, L.A.; Kaneno, R.; de Assis Golim, M.; Dos Santos, D.C.; Creste, C.F.Z.; Oba, E.; Maia, L.; Barraviera, B.; et al. A unique heterologous fibrin sealant (HFS) as a candidate biological scaffold for mesenchymal stem cells in osteoporotic rats. Stem Cell Res. Ther. 2017, 8, 205. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.M.; Zhao, H.Y.; Zhao, X.E.; Quan, F.S.; Hua, S.; He, X.Y.; Liu, J.; He, X.N.; Lin, H. Development of cloned embryos from porcine neural stem cells and amniotic fluid-derived stem cells transfected with enhanced green fluorescence protein gene. Reproduction 2009, 137, 793–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, S.Y.; Park, H.J.; Hong, J.-Y.; Lee, H.-J.; Lee, S.K. Down-regulation of SerpinB2 is associated with gefitinib resistance in non-small cell lung cancer and enhances invadopodia-like structure protrusions. Sci. Rep. 2016, 6, 32258. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.A.; Yerbury, J.J.; Farrawell, N.; Shearer, R.F.; Constantinescu, P.; Hatters, D.M.; Schroder, W.A.; Suhrbier, A.; Wilson, M.R.; Saunders, D.N.; et al. SerpinB2 (PAI-2) Modulates Proteostasis via Binding Misfolded Proteins and Promotion of Cytoprotective Inclusion Formation. PLoS ONE 2015, 10, e0130136. [Google Scholar] [CrossRef] [Green Version]
- Patwari, P.; Higgins, L.J.; Chutkow, W.A.; Yoshioka, J.; Lee, R.T. The interaction of thioredoxin with Txnip. Evidence for formation of a mixed disulfide by disulfide exchange. J. Biol. Chem. 2006, 281, 21884–21891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salhab, M.; Dhorne-Pollet, S.; Auclair, S.; Guyader-Joly, C.; Brisard, D.; Dalbies-Tran, R.; Dupont, J.; Ponsart, C.; Mermillod, P.; Uzbekova, S. In vitro maturation of oocytes alters gene expression and signaling pathways in bovine cumulus cells. Mol. Reprod. Dev. 2013, 80, 166–182. [Google Scholar] [CrossRef] [PubMed]
- Ożegowska, K.; Dyszkiewicz-Konwińska, M.; Celichowski, P.; Nawrocki, M.J.; Bryja, A.; Jankowski, M.; Kranc, W.; Brązert, M.; Knap, S.; Jeseta, M.; et al. Expression pattern of new genes regulating female sex differentiation and in vitro maturational status of oocytes in pigs. Theriogenology 2018, 121, 122–133. [Google Scholar] [CrossRef]
- Kulus, M.; Kranc, W.; Sujka-Kordowska, P.; Celichowski, P.; Konwerska, A.; Jankowski, M.; Jeseta, M.; Skowroński, M.T.; Piotrowska-Kempisty, H.; Bukowska, D.; et al. Transcriptomic analysis of expression of genes regulating cell cycle progression in porcine ovarian granulosa cells during short-term in vitro primary culture. Histochem. Cell Biol. 2020, 153, 397–412. [Google Scholar] [CrossRef] [PubMed]
- Napoli, J.L. Cellular retinoid binding-proteins, CRBP, CRABP, FABP5: Effects on retinoid metabolism, function and related diseases. Pharmacol. Ther. 2017, 173, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruberte, E.; Friederich, V.; Morriss-Kay, G.; Chambon, P. Differential distribution patterns of CRABP-I and CRABP-II transcripts during mouse embryogenesis. Development 1992, 115, 973–989. [Google Scholar]
- Gao, H.-F.; Chen, L.-Y.; Cheng, C.-S.; Chen, H.; Meng, Z.-Q.; Chen, Z. SLC5A1 promotes growth and proliferation of pancreatic carcinoma via glucose-dependent AMPK/mTOR signaling. Cancer Manag. Res. 2019, 11, 3171. [Google Scholar] [CrossRef] [Green Version]
- Ohta, T.; Isselbacher, K.J.; Rhoads, D.B. Regulation of Glucose Transporters in LLC-PK1 Cells: Effects of D-Glucose and Monosaccharides. Mol. Cell. Biol. 1990, 10, 6491–6499. [Google Scholar] [CrossRef] [Green Version]
- Kotlo, K.; Shukla, S.; Tawar, U.; Skidgel, R.A.; Danziger, R.S. Aminopeptidase N reduces basolateral Na + -K + -ATPase in proximal tubule cells. Am. J. Physiol. Physiol. 2007, 293, F1047–F1053. [Google Scholar] [CrossRef] [Green Version]
- Cheng, G.F.Y.; Yuen, C.W.; Ge, W. Evidence for the existence of a local activin-follistatin negative feedback loop in the goldfish pituitary and its regulation by activin and gonadal steroids. J. Endocrinol. 2007, 195, 373–384. [Google Scholar] [CrossRef] [Green Version]
- Yao, H.H.C.; Matzuk, M.M.; Jorgez, C.J.; Menke, D.B.; Page, D.C.; Swain, A.; Capel, B. Follistatin operates downstream ofWnt4 in mammalian ovary organogenesis. Dev. Dyn. 2004, 230, 210–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- George, J.; Saito, M.; Tsuta, K.; Iwakawa, R.; Shiraishi, K.; Scheel, A.H.; Uchida, S.; Watanabe, S.-I.; Nishikawa, R.; Noguchi, M.; et al. Genomic Amplification of CD274 (PD-L1) in Small-Cell Lung Cancer. Clin. Cancer Res. 2017, 23, 1220–1226. [Google Scholar] [CrossRef] [Green Version]
- Wirthgen, E.; Tuchscherer, M.; Otten, W.; Domanska, G.; Wollenhaupt, K.; Tuchscherer, A.; Kanitz, E. Activation of indoleamine 2,3-dioxygenase by LPS in a porcine model. Innate Immun. 2014, 20, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Oshiumi, H.; Miyashita, M.; Okamoto, M.; Morioka, Y.; Okabe, M.; Matsumoto, M.; Seya, T. DDX60 Is Involved in RIG-I-Dependent and Independent Antiviral Responses, and Its Function Is Attenuated by Virus-Induced EGFR Activation. Cell Rep. 2015, 11, 1193–1207. [Google Scholar] [CrossRef]
- Oliveira, T.F.; Mulchande, J.; Moreira, R.; Iley, J.; Archer, M. Crystallization and preliminary diffraction studies of porcine pancreatic elastase in complex with a novel inhibitor. Protein Pept. Lett. 2007, 14, 93–95. [Google Scholar] [CrossRef]
- Li, M.; Chen, L.; Tian, S.; Lin, Y.; Tang, Q.; Zhou, X.; Li, D.; Yeung, C.K.L.; Che, T.; Jin, L.; et al. Comprehensive variation discovery and recovery of missing sequence in the pig genome using multiple de novo assemblies. Genome Res. 2017, 27, 865–874. [Google Scholar] [CrossRef] [Green Version]
- Kumari, S.; Panda, T.K.; Pradhan, T. Lysyl Oxidase: Its Diversity in Health and Diseases. Indian J. Clin. Biochem. 2017, 32, 134. [Google Scholar] [CrossRef]
- Huang, H.; Chen, S.; Zhang, W.; Wang, S. Induction of EMT-like response by BMP4 via up-regulation of lysyl oxidase is required for adipocyte lineage commitment. Stem Cell Res. 2013, 10, 278–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erler, J.T.; Bennewith, K.L.; Cox, T.R.; Lang, G.; Bird, D.; Koong, A.; Le, Q.-T.; Giaccia, A.J. Hypoxia-induced lysyl oxidase is a critical mediator of bone marrow cell recruitment to form the premetastatic niche. Cancer Cell 2009, 15, 35–44. [Google Scholar] [CrossRef] [Green Version]
- Rozycki, M.; Bialik, J.F.; Speight, P.; Dan, Q.; Knudsen, T.E.T.; Szeto, S.G.; Yuen, D.A.; Szászi, K.; Pedersen, S.F.; Kapus, A. Myocardin-related Transcription Factor Regulates Nox4 Protein Expression: Linking cytoskeletal organization to redox state. J. Biol. Chem. 2016, 291, 227–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cucoranu, I.; Clempus, R.; Dikalova, A.; Phelan, P.J.; Ariyan, S.; Dikalov, S.; Sorescu, D. NAD(P)H Oxidase 4 Mediates Transforming Growth Factor- 1-Induced Differentiation of Cardiac Fibroblasts Into Myofibroblasts. Circ. Res. 2005, 97, 900–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bondi, C.D.; Manickam, N.; Lee, D.Y.; Block, K.; Gorin, Y.; Abboud, H.E.; Barnes, J.L. NAD(P)H oxidase mediates TGF-beta1-induced activation of kidney myofibroblasts. J. Am. Soc. Nephrol. 2010, 21, 93–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basuroy, S.; Tcheranova, D.; Bhattacharya, S.; Leffler, C.W.; Parfenova, H. Nox4 NADPH oxidase-derived reactive oxygen species, via endogenous carbon monoxide, promote survival of brain endothelial cells during TNF-α-induced apoptosis. Am. J. Physiol. Physiol. 2011, 300, C256–C265. [Google Scholar] [CrossRef] [Green Version]
- Basuroy, S.; Bhattacharya, S.; Leffler, C.W.; Parfenova, H. Nox4 NADPH oxidase mediates oxidative stress and apoptosis caused by TNF-α in cerebral vascular endothelial cells. Am. J. Physiol. Physiol. 2009, 296, C422–C432. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.L.; Li, K.; Peng, Z.Z.; Zhao, S.H.; Yu, M.; Fan, B.; Zhu, M.J.; Xu, S.P.; Du, Y.Q.; Liu, B. Molecular characterization, expression and association analysis of the porcine CMYA 4 gene with carcass traits. J. Anim. Breed. Genet. 2008, 125, 234–239. [Google Scholar] [CrossRef]
- Price, M.G.; Landsverk, M.L.; Barral, J.M.; Epstein, H.F. Two mammalian UNC-45 isoforms are related to distinct cytoskeletal and muscle-specific functions. J. Cell Sci. 2002, 115, 413–4023. [Google Scholar] [CrossRef] [Green Version]
- Sovic, A.; Panzenboeck, U.; Wintersperger, A.; Kratzer, I.; Hammer, A.; Levak-Frank, S.; Frank, S.; Rader, D.J.; Malle, E.; Sattler, W. Regulated expression of endothelial lipase by porcine brain capillary endothelial cells constituting the blood-brain barrier. J. Neurochem. 2005, 94, 109–119. [Google Scholar] [CrossRef]
- Bischoff, S.R.; Tsai, S.; Hardison, N.; Motsinger-Reif, A.A.; Freking, B.A.; Nonneman, D.; Rohrer, G.; Piedrahita, J.A. Characterization of conserved and nonconserved imprinted genes in swine. Biol. Reprod. 2009, 81, 906–920. [Google Scholar] [CrossRef] [Green Version]
- Fagerberg, L.; Hallström, B.M.; Oksvold, P.; Kampf, C.; Djureinovic, D.; Odeberg, J.; Habuka, M.; Tahmasebpoor, S.; Danielsson, A.; Edlund, K.; et al. Analysis of the Human Tissue-specific Expression by Genome-wide Integration of Transcriptomics and Antibody-based Proteomics. Mol. Cell. Proteomics 2014, 13, 397–406. [Google Scholar] [CrossRef] [Green Version]
- Esapa, C.T.; Waite, A.; Locke, M.; Benson, M.A.; Kraus, M.; McIlhinney, R.A.J.; Sillitoe, R.V.; Beesley, P.W.; Blake, D.J. SGCE missense mutations that cause myoclonus-dystonia syndrome impair ε-sarcoglycan trafficking to the plasma membrane: Modulation by ubiquitination and torsinA. Hum. Mol. Genet. 2007, 16, 327–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulus, M.; Borowiec, B.; Popis, M.; Celichowski, P.; Jeseta, M.; Bukowska, D.; Piotrowska-Kempisty, H.; Bruska, M.; Zabel, M.; Nowicki, M.; et al. Genes encoding proteins regulating fatty acid metabolism and cellular response to lipids are differentially expressed in porcine luminal epithelium during long-term culture. Med. J. Cell Biol. 2019, 7, 58–65. [Google Scholar] [CrossRef] [Green Version]
- Budna, J.; Celichowski, P.; Knap, S.; Jankowski, M.; Magas, M.; Nawrocki, M.J.; Ramlau, P.; Nowicki, A.; Rojewska, M.; Chermuła, B.; et al. Fatty Acids Related Genes Expression Undergo Substantial Changes in Porcine Oviductal Epithelial Cells During Long-Term Primary Culture. Med. J. Cell Biol. 2018, 6, 39–47. [Google Scholar] [CrossRef] [Green Version]
- Rao, X.; Huang, X.; Zhou, Z.; Lin, X. An improvement of the 2ˆ(-delta delta CT) method for quantitative real-time polymerase chain reaction data analysis. Biostat. Bioinforma Biomath. 2013, 3, 71–85. [Google Scholar]
- Gautier, L.; Cope, L.; Bolstad, B.M.; Irizarry, R.A. Affy--analysis of Affymetrix GeneChip data at the probe level. Bioinformatics 2004, 20, 307–315. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Tan, Q.; Kir, J.; Liu, D.; Bryant, D.; Guo, Y.; Stephens, R.; Baseler, M.W.; Lane, H.C.; et al. DAVID Bioinformatics Resources: Expanded annotation database and novel algorithms to better extract biology from large gene lists. Nucleic Acids Res. 2007, 35, W169–W175. [Google Scholar] [CrossRef] [PubMed]
- Nawrocki, M.J.; Sibiak, R.; Kałużna, S.; Brązert, M.; Celichowski, P.; Moncrieff, L.; Kempisty, B.; Mozdziak, P. Nucleotide, ribonucleotide and ribonucleoside binding belongs to differentially expressed genes in porcine epithelial oviductal cells during long- term primary cultivation. Med. J. Cell Biol. 2019, 7, 161–169. [Google Scholar] [CrossRef] [Green Version]
- Luo, W.; Brouwer, C. Pathview: An R/Bioconductor package for pathway-based data integration and visualization. Bioinformatics 2013, 29, 1830–1831. [Google Scholar] [CrossRef] [Green Version]
- Walter, W.; Sánchez-Cabo, F.; Ricote, M. GOplot: An R package for visually combining expression data with functional analysis. Bioinformatics 2015, 31, 2912–2914. [Google Scholar] [CrossRef]
- Von Mering, C.; Jensen, L.J.; Snel, B.; Hooper, S.D.; Krupp, M.; Foglierini, M.; Jouffre, N.; Huynen, M.A.; Bork, P. STRING: Known and predicted protein-protein associations, integrated and transferred across organisms. Nucleic Acids Res. 2005, 33, D433–D437. [Google Scholar] [CrossRef] [PubMed]
- Kocherova, I.; Brązert, M.; Sujka-kordowska, P.; Konwerska, A.; Chermuła, B.; Celichowski, P.; Piotrowska-kempisty, H.; Antosik, P.; Bukowska, D.; Bruska, M.; et al. Genes regulating biochemical pathways of oxygen metabolism in porcine oviductal epithelial cells during long-term IVC. Med. J. Cell Biol. 2019, 7, 39–47. [Google Scholar] [CrossRef] [Green Version]
- Matysiak, J.; Hajduk, J.; Mayer, F.; Hebeler, R.; Kokot, Z.J. Hyphenated LC–MALDI–ToF/ToF and LC–ESI–QToF approach in proteomic characterization of honeybee venom. J. Pharm. Biomed. Anal. 2016, 121, 69–76. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Fold Change D7/D1 | Fold Change D15/D1 | Fold Change D30/D1 | Adjusted p-Value D7/D1 | Adjusted p-Value D15/D1 | Adjusted p-Value D30/D1 |
|---|---|---|---|---|---|---|
| SERPINB2 | 0.041199 | 0.012677 | 0.008239 | 4.60 × 10−6 | 7.45 × 10−7 | 2.31 × 10−7 |
| CD274 | 0.04424 | 0.04443 | 0.041834 | 2.50 × 10−6 | 7.45 × 10−7 | 2.31 × 10−7 |
| TXNIP | 0.082179 | 0.64453 | 0.63523 | 1.11 × 10−5 | 0.024019 | 0.017645 |
| CELA1 | 0.10027 | 0.111224 | 0.091977 | 4.67 × 10−5 | 3.35 × 10−5 | 1.41 × 10−5 |
| DDX60 | 0.104831 | 0.194221 | 0.213218 | 7.61 × 10−5 | 0.000244 | 0.000232 |
| CRABP2 | 0.108046 | 0.084638 | 0.056801 | 3.11 × 10−5 | 1.05 × 10−5 | 2.84 × 10−6 |
| SLC5A1 | 0.122502 | 0.081281 | 0.097509 | 3.30 × 10−5 | 7.64 × 10−6 | 6.67 × 10−6 |
| IDO1 | 0.134544 | 0.12111 | 0.123331 | 7.22 × 10−5 | 3.53 × 10−5 | 2.36 × 10−5 |
| ANPEP | 0.143464 | 0.201711 | 0.482675 | 1.74 × 10−5 | 1.96 × 10−5 | 0.000794 |
| FST | 0.148055 | 0.075627 | 0.078555 | 1.84 × 10−5 | 2.99 × 10−6 | 1.32 × 10−6 |
| UNC45B | 10.72559 | 2.574362 | 1.464151 | 1.45 × 10−5 | 0.000645 | 0.040563 |
| NOX4 | 11.05623 | 21.12763 | 32.07053 | 4.90 × 10−6 | 9.73 × 10−7 | 2.31 × 10−7 |
| VLDLR | 11.18756 | 15.02697 | 21.33103 | 1.03 × 10−5 | 3.25 × 10−6 | 8.85 × 10−7 |
| ITGB3 | 12.67365 | 13.15962 | 10.9347 | 4.27 × 10−6 | 1.55 × 10−6 | 8.03 × 10−7 |
| FMOD | 12.87587 | 41.88541 | 41.90401 | 2.50 × 10−6 | 6.02 × 10−7 | 2.31 × 10−7 |
| SGCE | 13.222 | 29.18235 | 46.77017 | 5.88 × 10−6 | 9.76 × 10−7 | 2.51 × 10−7 |
| COL1A2 | 15.57291 | 56.37555 | 61.32906 | 3.63 × 10−6 | 7.45 × 10−7 | 2.31 × 10−7 |
| LOX | 29.0517 | 43.84613 | 46.65297 | 7.88 × 10−6 | 2.86 × 10−6 | 1.06 × 10−6 |
| LIPG | 36.5469 | 19.67815 | 39.01801 | 2.50 × 10−6 | 1.39 × 10−6 | 2.57 × 10−7 |
| THY1 | 38.75283 | 32.74287 | 31.30186 | 3.01 × 10−6 | 1.39 × 10−6 | 5.59 × 10−7 |
| Gene | Gene ID | Primer Sequence (5′–3′) | Product Size (bp) |
|---|---|---|---|
| THY1 | 100271931 | CCAAAGATGAGGGGATCTACG CCAAAGATGAGGGGATCTACG | 100 |
| LIPG | 100155736 | ATCCTGAGAACACCCGCATA AGGATGCTCCACAGTTGGAC | 104 |
| LOX | 100525278 | CCAGAGGAGAGTGGCTGAAG CTGGGGTTCACACTGACCTT | 216 |
| COL1A2 | 100626716 | GTCAGACTGGTCCTGCTGGT GTCAGACTGGTCCTGCTGGT | 163 |
| SGCE | 100240725 | CCAACAATCATTGAGATAACTGC GCTGGCCAACATTTCTTCTA | 156 |
| FMOD | 100526237 | TGCTCACTGGGTCTGTGAAG CCTCAAAGATAGGGGCTTCC | 194 |
| ITGB3 | 397063 | GGCTTCAAAGACAGCCTCAC AGTCCTTTTCCGAGCACTCA | 175 |
| VLDLR | 733630 | TGAGCCTTCCCAATTCCAGT CATATGGCACTGTTCTGGGC | 232 |
| NOX4 | 100523323 | ACAACTGTTCCTGGCCTGAC CAGCCCTCCTGAAACATGTAA | 168 |
| UNC45B | 100134956 | GCCTGAAAACGGAGAGCTATG ACAACGCTGCACGTCCTT | 152 |
| FST | 445002 | CTGAGCACCTCCGACGAG ACGTTTCTTTACATGGGATGC | 100 |
| ANPEP | 397520 | CCACCATCTACTGCAATGCC CGTCTTGCTTCCGAATGAGG | 195 |
| IDO1 | 100519877 | CACTGTGGGTGGAGTTCCTT TTTCGCAGGGATACCATAGC | 198 |
| SLC5A1 | 397113 | CTTTGCCATCATCCTCTTTG CCAACACAGGCGGTAGAGAT | 100 |
| CRABP2 | 100155151 | GAGCCTGGTAAAATGGGAGA CTCGGACATAGACCCTGGTG | 164 |
| DDX60 | 100158037 | GCTCATGCTCCTGGCTTC CTTGACCAGGAATTGCAGTG | 154 |
| CELA1 | 396766 | GCTCTGGATGTCAGGGTGAT GCTCTGGATGTCAGGGTGAT | 176 |
| TXNIP | 733688 | CAAGCCAGCCAACTCAAGAG TTCGAGCAGAGACAGACACC | 212 |
| CD274 | 574058 | GTGTTGGTCATCCCAGAACC TATCTCGGCTGCCACATTTT | 157 |
| SERPINB2 | 100519286 | GGAAGAATACATTCGACTCTCCA GGAAGAATACATTCGACTCTCCA | 170 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kulus, M.; Kranc, W.; Wojtanowicz-Markiewicz, K.; Celichowski, P.; Światły-Błaszkiewicz, A.; Matuszewska, E.; Sujka-Kordowska, P.; Konwerska, A.; Zdun, M.; Bryl, R.; et al. New Gene Markers Expressed in Porcine Oviductal Epithelial Cells Cultured Primary In Vitro Are Involved in Ontological Groups Representing Physiological Processes of Porcine Oocytes. Int. J. Mol. Sci. 2021, 22, 2082. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22042082
Kulus M, Kranc W, Wojtanowicz-Markiewicz K, Celichowski P, Światły-Błaszkiewicz A, Matuszewska E, Sujka-Kordowska P, Konwerska A, Zdun M, Bryl R, et al. New Gene Markers Expressed in Porcine Oviductal Epithelial Cells Cultured Primary In Vitro Are Involved in Ontological Groups Representing Physiological Processes of Porcine Oocytes. International Journal of Molecular Sciences. 2021; 22(4):2082. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22042082
Chicago/Turabian StyleKulus, Magdalena, Wiesława Kranc, Katarzyna Wojtanowicz-Markiewicz, Piotr Celichowski, Agata Światły-Błaszkiewicz, Eliza Matuszewska, Patrycja Sujka-Kordowska, Aneta Konwerska, Maciej Zdun, Rut Bryl, and et al. 2021. "New Gene Markers Expressed in Porcine Oviductal Epithelial Cells Cultured Primary In Vitro Are Involved in Ontological Groups Representing Physiological Processes of Porcine Oocytes" International Journal of Molecular Sciences 22, no. 4: 2082. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22042082