
Effects of Different Short-Term UV-B Radiation Intensities on Metabolic Characteristics of Porphyra haitanensis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
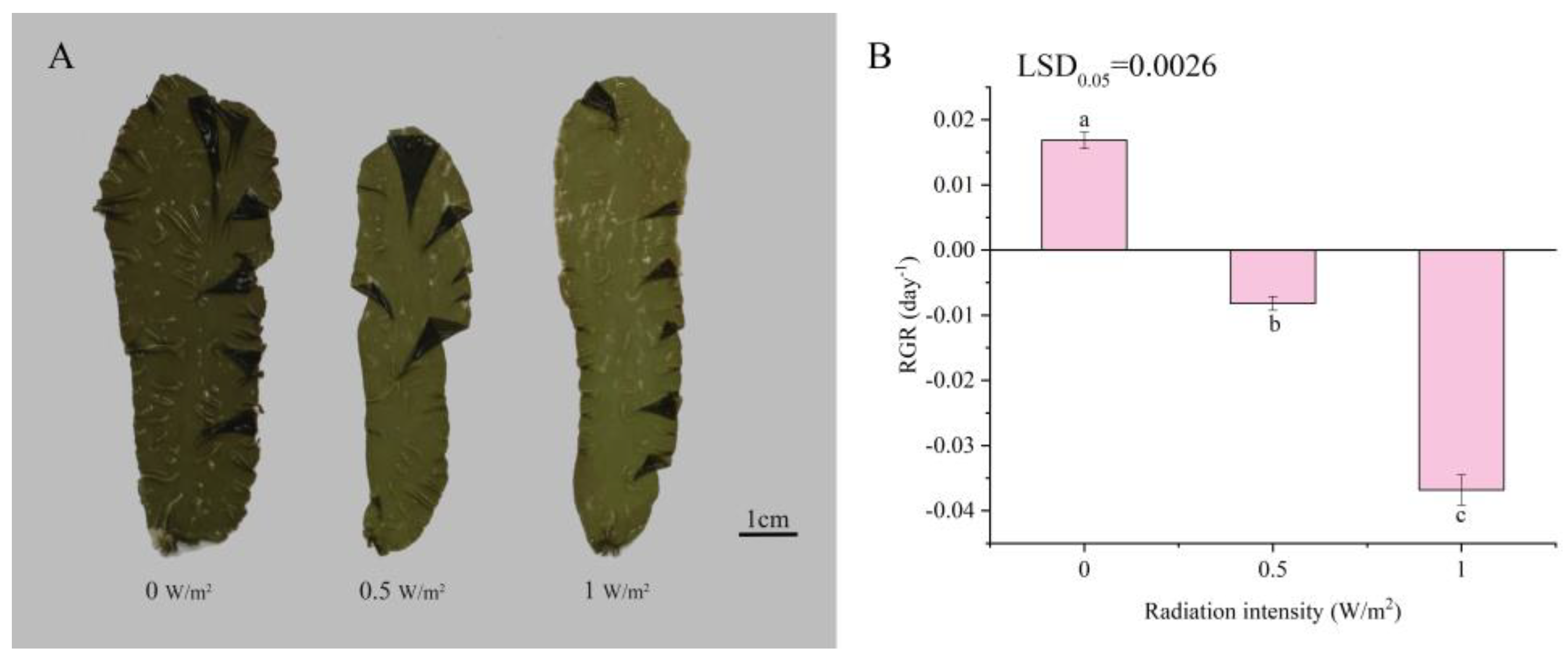
2.1. Effects of UV-B Radiation on the Morphology and Relative Growth Rate of P. haitanensis
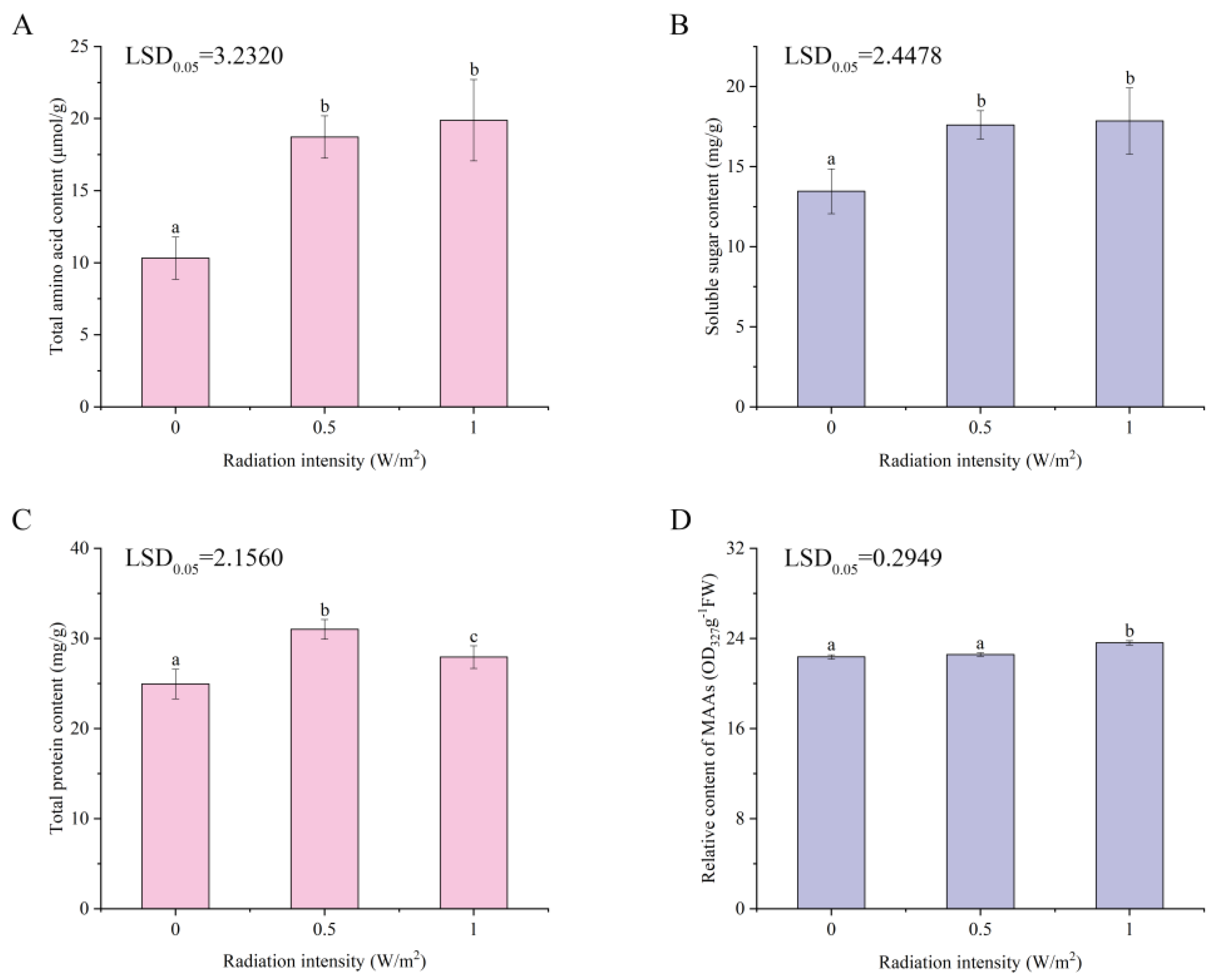
2.2. Effects of UV-B Irradiation on Total Amino Acids, Soluble Sugar, Total Protein, and MAAs of P. haitanensis
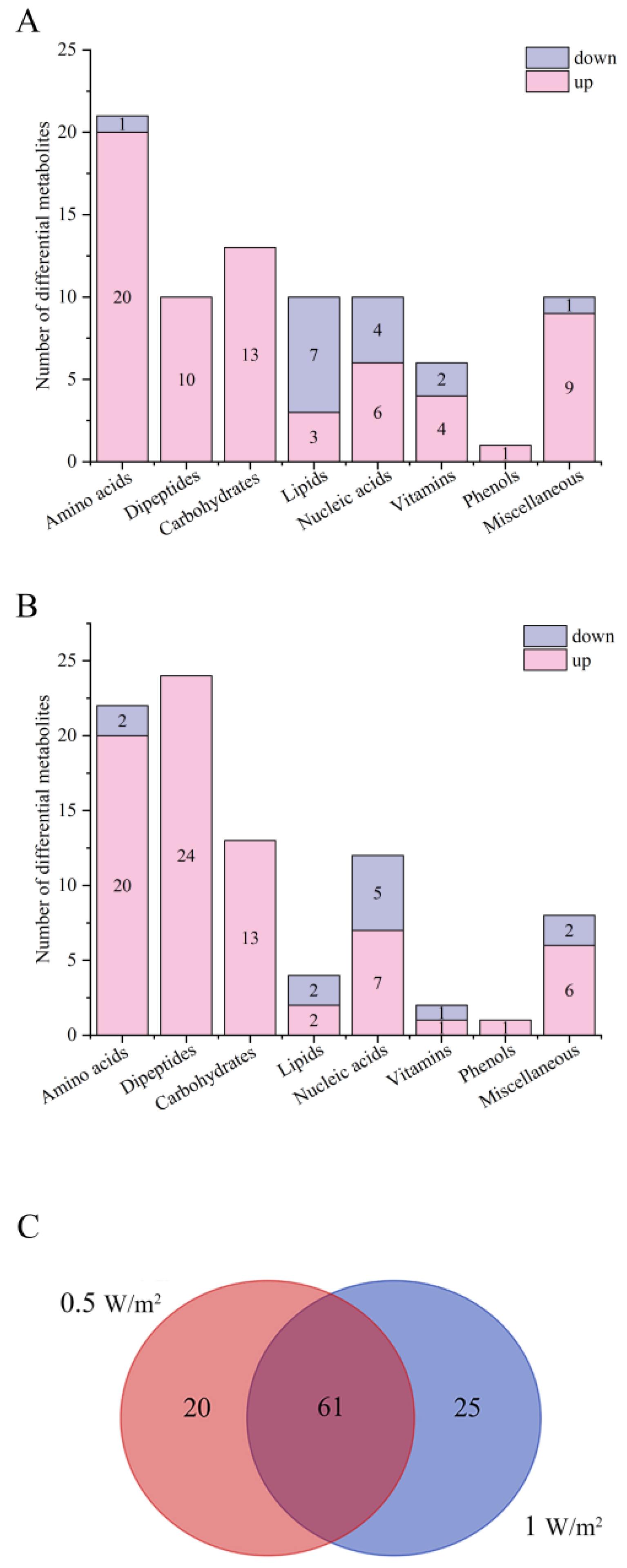
2.3. Analysis of Differential Metabolites of P. haitanensis under Different UV-B Radiation Intensities
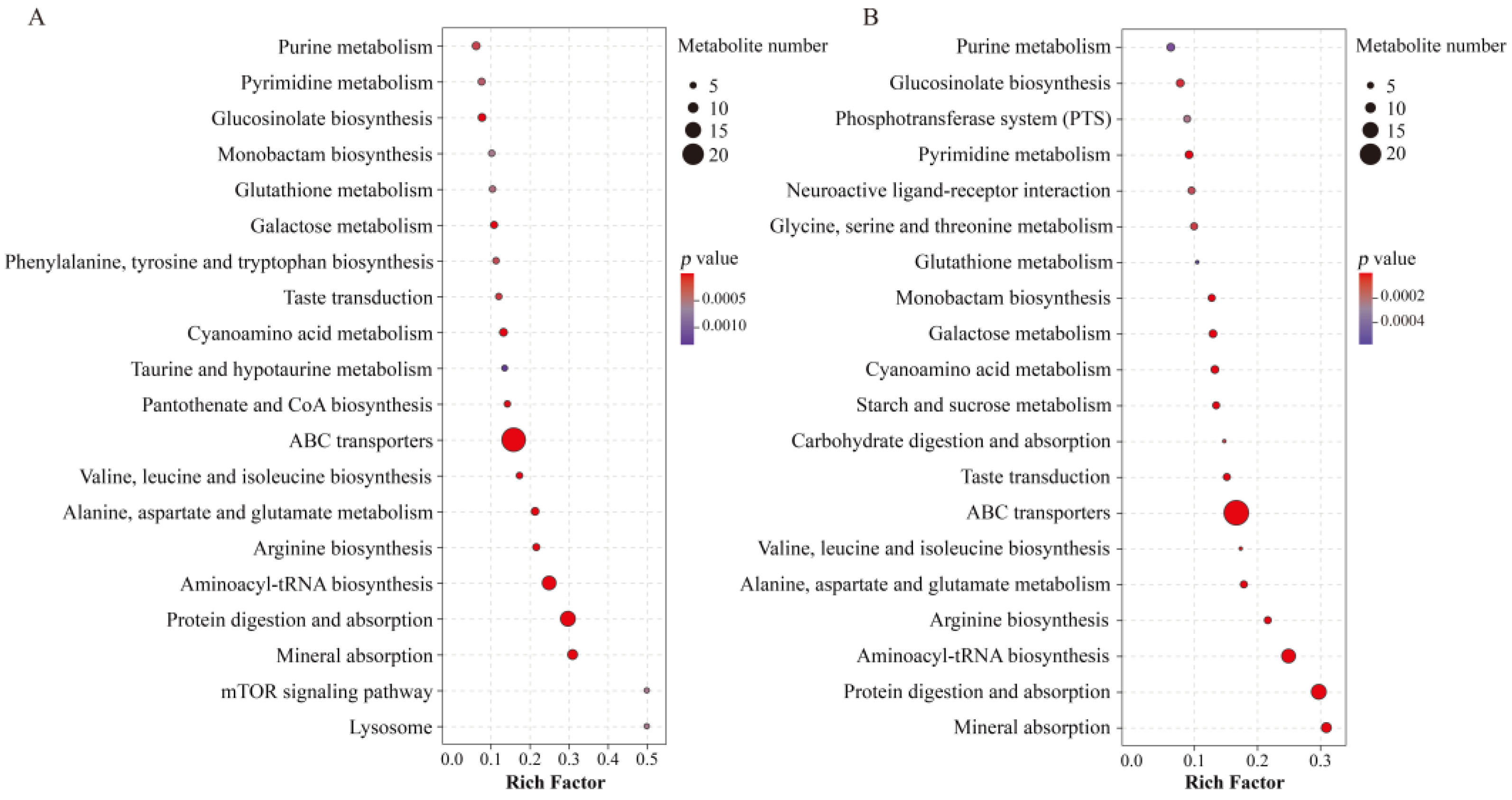
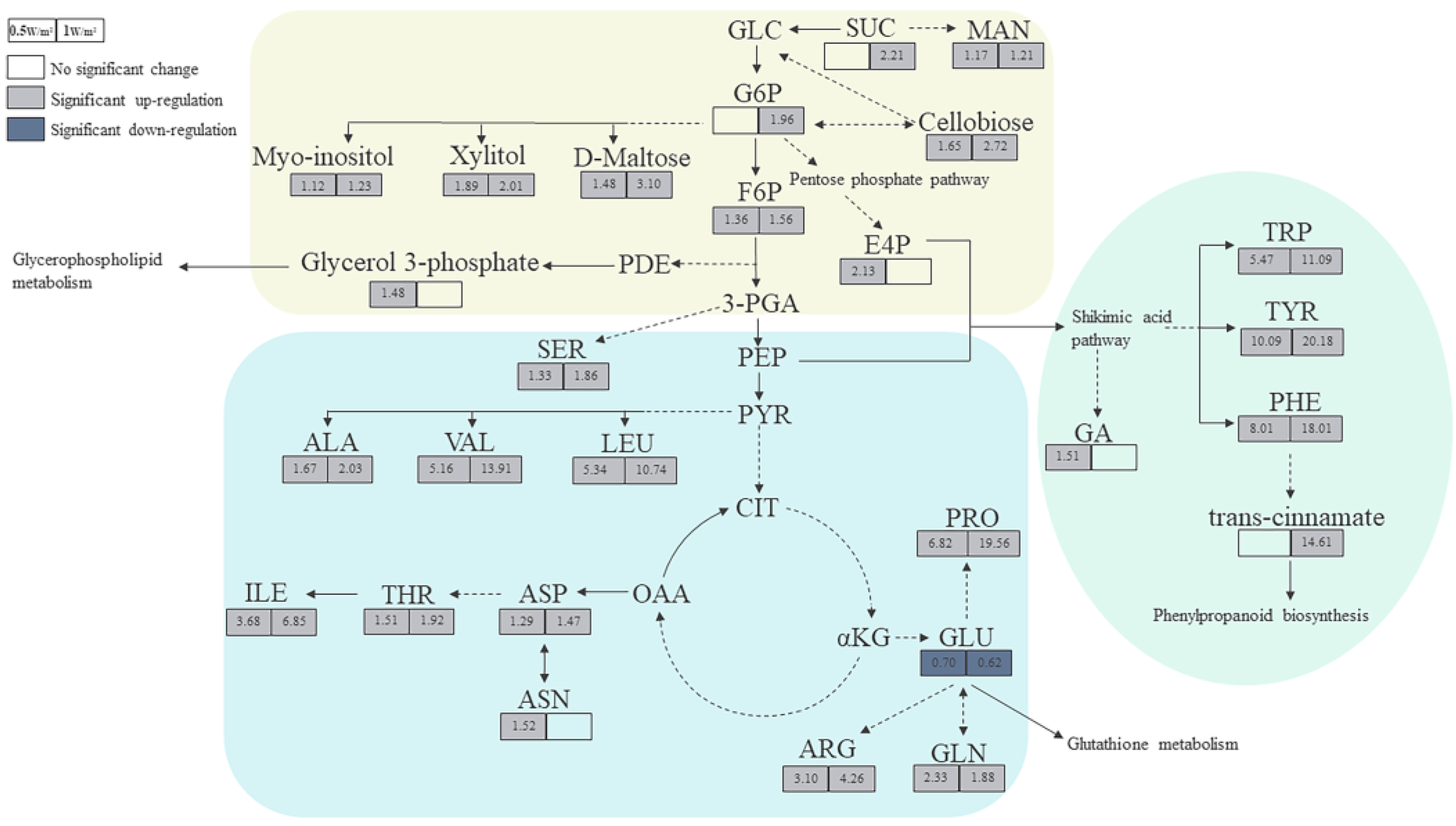
2.4. Metabolic Pathway Analysis of Differential Metabolites of P. haitanensis under Different UV-B Radiation Intensities
3. Discussion
4. Materials and Methods
4.1. Algal Materials and Growth Conditions
4.2. UV-B Treatments
4.3. Physiological Analysis
4.3.1. Relative Growth Rate
4.3.2. Total Amino Acids, Soluble Sugar, Total Protein, and MAAs
4.4. Metabolic Analysis and Data Processing
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- McFarland, M.; Kaye, J. Chlorofluorocarbons and Ozone. Photochem. Photobiol. 1992, 55, 911–929. [Google Scholar] [CrossRef]
- Anderson, J.G.; Toohey, D.W.; Brune, W.H. Free Radicals Within the Antarctic Vortex: The Role of CFCs in Antarctic Ozone Loss. Science 1991, 251, 39–46. [Google Scholar] [CrossRef] [Green Version]
- Pessoa, M.F. Harmful effects of UV radiation in Algae and aquatic macrophytes—A review. Emir. J. Food Agric. 2012, 24, 510–526. [Google Scholar] [CrossRef]
- Dahms, H.-U.; Lee, J.-S. UV radiation in marine ectotherms: Molecular effects and responses. Aquat. Toxicol. 2010, 97, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Short, F.T.; Neckles, H.A. The effects of global climate change on seagrasses. Aquat. Bot. 1999, 63, 169–196. [Google Scholar] [CrossRef]
- Häder, D.-P.; Helbling, E.W.; Williamson, C.E.; Worrest, R.C. Effects of UV radiation on aquatic ecosystems and interactions with climate change. Photochem. Photobiol. Sci. 2011, 10, 242–260. [Google Scholar] [CrossRef]
- Warner, M.E.; Madden, M.L. The impact of shifts to elevated irradiance on the growth and photochemical activity of the harmful algae Chattonella subsalsa and Prorocentrum minimum from Delaware. Harmful Algae 2007, 6, 332–342. [Google Scholar] [CrossRef]
- Jiang, H.; Zou, D.; Lou, W.; Deng, Y.; Zeng, X. Effects of seawater acidification and alkalization on the farmed seaweed, Pyropia haitanensis (Bangiales, Rhodophyta), grown under different irradiance conditions. Algal Res. 2018, 31, 413–420. [Google Scholar] [CrossRef]
- Zou, D.; Gao, K. Effects of elevated CO2 on the red seaweed Gracilaria lemaneiformis (Gigartinales, Rhodophyta) grown at different irradiance levels. Phycologia 2009, 48, 510–517. [Google Scholar] [CrossRef] [Green Version]
- Becker, S.; Wälter, B.; Bischof, K. Freezing tolerance and photosynthetic performance of polar seaweeds at low temperatures. Bot. Mar. 2009, 52, 609–616. [Google Scholar] [CrossRef]
- Nalley, J.O.; O’Donnell, D.R.; Litchman, E. Temperature effects on growth rates and fatty acid content in freshwater algae and cyanobacteria. Algal Res. 2018, 35, 500–507. [Google Scholar] [CrossRef]
- Cui, J.; Zhang, J.; Huo, Y.; Zhou, L.; Wu, Q.; Chen, L.; Yu, K.; He, P. Adaptability of free-floating green tide algae in the Yellow Sea to variable temperature and light intensity. Mar. Pollut. Bull. 2015, 101, 660–666. [Google Scholar] [CrossRef] [PubMed]
- Altamirano, M.; Flores-Moya, A.; Figueroa, F.L. Effects of UV radiation and temperature on growth of germlings of three species of Fucus (Phaeophyceae). Aquat. Bot. 2003, 75, 9–20. [Google Scholar] [CrossRef]
- Roleda, M.Y.; Wiencke, C.; Hanelt, D. Thallus morphology and optical characteristics affect growth and DNA damage by UV radiation in juvenile Arctic Laminaria sporophytes. Planta 2005, 223, 407–417. [Google Scholar] [CrossRef] [Green Version]
- Han, T.; Han, Y.-S.; Kim, K.-Y.; Kim, J.-H.; Shin, H.-W.; Kain, J.M.; Callow, J.A.; Callow, M.E. Influences of light and UV-B on growth and sporulation of the green alga Ulva pertusa Kjellman. J. Exp. Mar. Biol. Ecol. 2003, 290, 115–131. [Google Scholar] [CrossRef]
- Wang, Q.-F.; Hou, Y.-H.; Miao, J.-L.; Li, G.-Y. Effect of UV-B radiation on the growth and antioxidant enzymes of Antarctic sea ice microalgae Chlamydomonas sp. ICE-L. Acta Physiol. Plant. 2009, 31, 1097–1102. [Google Scholar] [CrossRef]
- Helbling, E.W.; Buma, A.G.; Van De Poll, W.; Zenoff, M.V.F.; Villafañe, V.E. UVR-induced photosynthetic inhibition dominates over DNA damage in marine dinoflagellates exposed to fluctuating solar radiation regimes. J. Exp. Mar. Biol. Ecol. 2008, 365, 96–102. [Google Scholar] [CrossRef]
- Aguilera, J.; Jiménez, C.; Figueroa, F.L.; Lebert, M.; Häder, D.-P. Effect of ultraviolet radiation on thallus absorption and photosynthetic pigments in the red alga Porphyra umbilicalis. J. Photochem. Photobiol. B Biol. 1999, 48, 75–82. [Google Scholar] [CrossRef]
- Holzinger, A.; Roleda, M.Y.; Lütz, C. The vegetative arctic freshwater green alga Zygnema is insensitive to experimental UV exposure. Micron 2009, 40, 831–838. [Google Scholar] [CrossRef] [Green Version]
- Bischof, K.; Hanelt, D.; Wiencke, C. Effects of ultraviolet radiation on photosynthesis and related enzyme reactions of marine macroalgae. Planta 2000, 211, 555–562. [Google Scholar] [CrossRef]
- Tian, J.; Yu, J. Changes in ultrastructure and responses of antioxidant systems of algae (Dunaliella salina) during acclimation to enhanced ultraviolet-B radiation. J. Photochem. Photobiol. B Biol. 2009, 97, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Shiu, C.-T.; Lee, T.-M. Ultraviolet-B-induced oxidative stress and responses of the ascorbate–glutathione cycle in a marine macroalga Ulva fasciata. J. Exp. Bot. 2005, 56, 2851–2865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leu, E.; Falk-Petersen, S.; Hessen, D.O. Ultraviolet radiation negatively affects growth but not food quality of arctic diatoms. Limnol. Oceanogr. 2007, 52, 787–797. [Google Scholar] [CrossRef]
- Liang, Y.; Beardall, J.; Heraud, P. Effects of nitrogen source and UV radiation on the growth, chlorophyll fluorescence and fatty acid composition of Phaeodactylum tricornutum and Chaetoceros muelleri (Bacillariophyceae). J. Photochem. Photobiol. B Biol. 2006, 82, 161–172. [Google Scholar] [CrossRef]
- Van De Poll, W.H.; Eggert, A.; Buma, A.G.J.; Breeman, A.M. Effects of Uv-B-Induced Dna Damage and Photoinhibition on Growth of Temperate Marine Red Macrophytes: Habitat-Related Differences in Uv-B Tolerance. J. Phycol. 2001, 37, 30–38. [Google Scholar] [CrossRef]
- Van de Poll, W.H.; Hanelt, D.; Hoyer, K.; Buma, A.G.; Breeman, A.M. Ultraviolet-B-induced cyclobutane-pyrimidine dimer formation and repair in Arctic marine macrophytes. Photochem. Photobiol. 2002, 76, 493–500. [Google Scholar] [CrossRef]
- Lee, T.-M.; Shiu, C.-T. Implications of mycosporine-like amino acid and antioxidant defenses in UV-B radiation tolerance for the algae species Ptercladiella capillacea and Gelidium amansii. Mar. Environ. Res. 2009, 67, 8–16. [Google Scholar] [CrossRef] [Green Version]
- Klisch, M.; Häder, D.-P. Mycosporine-Like Amino Acids and Marine Toxins—The Common and the Different. Mar. Drugs 2008, 6, 147–163. [Google Scholar] [CrossRef]
- Carreto, J.I.; Carignan, M.O. Mycosporine-Like Amino Acids: Relevant Secondary Metabolites. Chemical and Ecological Aspects. Mar. Drugs 2011, 9, 387–446. [Google Scholar] [CrossRef]
- Duval, B.; Shetty, K.; Thomas, W.H. Phenolic compounds and antioxidant properties in the snow alga Chlamydomonas nivalis after exposure to UV light. Environ. Boil. Fishes 1999, 11, 559–566. [Google Scholar] [CrossRef]
- Sass, L.; Spetea, C.; Máté, Z.; Nagy, F.; Vass, I. Repair of UV-B induced damage of Photosystem II via de novo synthesis of the D1 and D2 reaction centre subunits in Synechocystis sp. PCC 6803. Photosynth. Res. 1997, 54, 55–62. [Google Scholar] [CrossRef]
- Aguilera, J.; Dummermuth, A.; Karsten, U.; Schriek, R.; Wiencke, C. Enzymatic defences against photooxidative stress induced by ultraviolet radiation in Arctic marine macroalgae. Polar Biol. 2002, 25, 432–441. [Google Scholar] [CrossRef] [Green Version]
- Zeeshan, M.; Prasad, S. Differential response of growth, photosynthesis, antioxidant enzymes and lipid peroxidation to UV-B radiation in three cyanobacteria. S. Afr. J. Bot. 2009, 75, 466–474. [Google Scholar] [CrossRef] [Green Version]
- Pidatala, V.R.; Li, K.; Sarkar, D.; Ramakrishna, W.; Datta, R. Identification of Biochemical Pathways Associated with Lead Tolerance and Detoxification in Chrysopogon zizanioides L. Nash (Vetiver) by Metabolic Profiling. Environ. Sci. Technol. 2016, 50, 2530–2537. [Google Scholar] [CrossRef] [PubMed]
- Ryan, D.; Robards, K. Metabolomics: The Greatest Omics of Them All? Anal. Chem. 2006, 78, 7954–7958. [Google Scholar] [CrossRef]
- Xiaolei, F.; Yongjun, F.; Songnian, H.; Guangce, W. Generation and analysis of 5318 expressed sequence tags from the filamentous sporophyte of Porphyra haitanensis (Rhodophyta) 1. J. Phycol. 2007, 43, 1287–1294. [Google Scholar] [CrossRef]
- Niu, J.; Xie, S.-W.; Fang, H.-H.; Xie, J.-J.; Guo, T.-Y.; Zhang, Y.-M.; Liu, Z.-L.; Liao, S.-Y.; He, J.-Y.; Tian, L.-X.; et al. Dietary values of macroalgae Porphyra haitanensis in Litopenaeus vannamei under normal rearing and WSSV challenge conditions: Effect on growth, immune response and intestinal microbiota. Fish Shellfish. Immunol. 2018, 81, 135–149. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.-Y.; Chen, X.-Q.; Liu, Y.; Cheong, K.-L. Ultrasonic/microwave-assisted extraction, simulated digestion, and fermentation in vitro by human intestinal flora of polysaccharides from Porphyra haitanensis. Int. J. Biol. Macromol. 2020, 152, 748–756. [Google Scholar] [CrossRef]
- Khan, B.M.; Qiu, H.-M.; Xu, S.-Y.; Liu, Y.; Cheong, K.-L. Physicochemical characterization and antioxidant activity of sulphated polysaccharides derived from Porphyra haitanensis. Int. J. Biol. Macromol. 2020, 145, 1155–1161. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, J.; Karsten, U.; Lippert, H.; Vögele, B.; Philipp, E.; Hanelt, D.; Wiencke, C. Effects of solar radiation on growth, photosynthesis and respiration of marine macroalgae from the Arctic. Mar. Ecol. Prog. Ser. 1999, 191, 109–119. [Google Scholar] [CrossRef]
- Grobe, C.W.; Murphy, T.M. Artificial ultraviolet-B radiation and cell expansion in the intertidal alga Ulva expansa (Setch.) S. and G. (Chlorophyta). J. Exp. Mar. Biol. Ecol. 1997, 217, 209–223. [Google Scholar] [CrossRef]
- Hildebrandt, T.M.; Nesi, A.N.; Araújo, W.L.; Braun, H.-P. Amino Acid Catabolism in Plants. Mol. Plant 2015, 8, 1563–1579. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Lüscher, J.; Torres, N.; Hilbert, G.; Richard, T.; Sánchez-Díaz, M.; Delrot, S.; Aguirreolea, J.; Pascual, I.; Gomès, E. Ultraviolet-B radiation modifies the quantitative and qualitative profile of flavonoids and amino acids in grape berries. Phytochemistry 2014, 102, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Yue, M.; Li, Y.; Wang, X. Effects of enhanced ultraviolet-B radiation on plant nutrients and decomposition of spring wheat under field conditions. Environ. Exp. Bot. 1998, 40, 187–196. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Kim, S.; Lee, H.; Kim, K.H. Metabolomic elucidation of recovery of Melissa officinalis from UV-B irradiation stress. Ind. Crop. Prod. 2018, 121, 428–433. [Google Scholar] [CrossRef]
- Hu, X.; Ouyang, S.; Mu, L.; An, J.; Zhou, Q. Effects of Graphene Oxide and Oxidized Carbon Nanotubes on the Cellular Division, Microstructure, Uptake, Oxidative Stress, and Metabolic Profiles. Environ. Sci. Technol. 2015, 49, 10825–10833. [Google Scholar] [CrossRef] [PubMed]
- Han, T.; Liang, Y.; Wu, Z.; Zhang, L.; Liu, Z.; Li, Q.; Chen, X.; Guo, W.; Jiang, L.; Pan, F.; et al. Effects of tetracycline on growth, oxidative stress response, and metabolite pattern of ryegrass. J. Hazard. Mater. 2019, 380, 120885. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, J.; Wang, H.-Z.; Wu, K.-X.; Guo, X.-R.; Mu, L.-Q.; Tang, Z.-H. Comparison of the global metabolic responses to UV-B radiation between two medicinal Astragalus species: An integrated metabolomics strategy. Environ. Exp. Bot. 2020, 176, 104094. [Google Scholar] [CrossRef]
- Hollósy, F. Effects of ultraviolet radiation on plant cells. Micron 2002, 33, 179–197. [Google Scholar] [CrossRef]
- Ma, M.; Zhang, H.; Xie, Y.; Yang, M.; Tang, J.; Wang, P.; Yang, R.; Gu, Z. Response of nutritional and functional composition, anti-nutritional factors and antioxidant activity in germinated soybean under UV-B radiation. LWT 2020, 118, 108709. [Google Scholar] [CrossRef]
- De Vries, J.; De Vries, S.; Slamovits, C.H.; Rose, L.E.; Archibald, J.M. How Embryophytic is the Biosynthesis of Phenylpropanoids and their Derivatives in Streptophyte Algae? Plant Cell Physiol. 2017, 58, 934–945. [Google Scholar] [CrossRef] [PubMed]
- Escobar, A.L.; Silva, F.M.D.O.; Acevedo, P.; Nunes-Nesi, A.; Alberdi, M.; Reyes-Díaz, M. Different levels of UV-B resistance in Vaccinium corymbosum cultivars reveal distinct backgrounds of phenylpropanoid metabolites. Plant Physiol. Biochem. 2017, 118, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Huovinen, P.; Gómez, I.; Figueroa, F.L.; Ulloa, N.; Morales, V.; Lovengreen, C. Ultraviolet-absorbing mycosporine-like amino acids in red macroalgae from Chile. Bot. Mar. 2004, 47, 21–29. [Google Scholar] [CrossRef]
- Karsten, U.; Bischof, K.; Hanelt, D.; Tüg, H.; Wiencke, C. The effect of ultraviolet radiation on photosynthesis and ultraviolet-absorbing substances in the endemic Arctic macroalga Devaleraea ramentacea (Rhodophyta). Physiol. Plant. 1999, 105, 58–66. [Google Scholar] [CrossRef]
- Wada, N.; Sakamoto, T.; Matsugo, S. Mycosporine-Like Amino Acids and Their Derivatives as Natural Antioxidants. Antioxidants 2015, 4, 603–646. [Google Scholar] [CrossRef]
- Cho, Y.-H.; Yoo, S.-D. Signaling Role of Fructose Mediated by FINS1/FBP in Arabidopsis thaliana. PLoS Genet. 2011, 7, e1001263. [Google Scholar] [CrossRef] [Green Version]
- Rolland, F.; Baena-Gonzalez, E.; Sheen, J. Sugar Sensing and Signaling in Plants: Conserved and Novel Mechanisms. Annu. Rev. Plant Biol. 2006, 57, 675–709. [Google Scholar] [CrossRef] [Green Version]
- Ende, W.V.D.; Valluru, R. Sucrose, sucrosyl oligosaccharides, and oxidative stress: Scavenging and salvaging? J. Exp. Bot. 2008, 60, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Furlan, A.; Llanes, A.; Luna, V.; Castro, S. Physiological and Biochemical Responses to Drought Stress and Subsequent Rehydration in the Symbiotic Association Peanut-Bradyrhizobium sp. ISRN Agron. 2012, 2012, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Mortimer, M.; Kasemets, K.; Vodovnik, M.; Marinšek-Logar, R.; Kahru, A. Exposure to CuO Nanoparticles Changes the Fatty Acid Composition of Protozoa Tetrahymena thermophila. Environ. Sci. Technol. 2011, 45, 6617–6624. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Marin, E.; Martínez, A. Carbohydrates and Their Free Radical Scavenging Capability: A Theoretical Study. J. Phys. Chem. B 2012, 116, 9668–9675. [Google Scholar] [CrossRef]
- Gillaspy, G.E. The cellular language of myo-inositol signaling. New Phytol. 2011, 192, 823–839. [Google Scholar] [CrossRef]
- Laakso, K.; Kinnunen, H.; Huttunen, S. The glutathione status of mature Scots pines during the third season of UV-B radiation exposure. Environ. Pollut. 2001, 111, 349–354. [Google Scholar] [CrossRef]
- Noctor, G.; Foyer, C.H. Ascorbate and Glutathione: Keeping Active Oxygen Under Control. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef]
- Smirnoff, N.; Wheeler, G.L. Ascorbic Acid in Plants: Biosynthesis and Function. Crit. Rev. Biochem. Mol. Biol. 2000, 35, 291–314. [Google Scholar] [CrossRef]
- Asensi-Fabado, M.A.; Munné-Bosch, S. Vitamins in plants: Occurrence, biosynthesis and antioxidant function. Trends Plant Sci. 2010, 15, 582–592. [Google Scholar] [CrossRef]
- Prasad, S.M.; Kumar, S.; Parihar, P.; Singh, R. Interactive effects of herbicide and enhanced UV-B on growth, oxidative damage and the ascorbate-glutathione cycle in two Azolla species. Ecotoxicol. Environ. Saf. 2016, 133, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Scherer, S.; Chen, T.; Böger, P. A New UV-A/B Protecting Pigment in the Terrestrial Cyanobacterium Nostoc commune. Plant Physiol. 1988, 88, 1055–1057. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, X.; Yang, S.; Yuan, Y. Lignin Involvement in Programmed Changes in Peach-Fruit Texture Indicated by Metabolite and Transcriptome Analyses. J. Agric. Food Chem. 2018, 66, 12627–12640. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, S.; Xue, S.; Chen, J.; Shang, S.; Xiao, H.; Zang, Y.; Tang, X. Effects of Different Short-Term UV-B Radiation Intensities on Metabolic Characteristics of Porphyra haitanensis. Int. J. Mol. Sci. 2021, 22, 2180. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22042180
Fu S, Xue S, Chen J, Shang S, Xiao H, Zang Y, Tang X. Effects of Different Short-Term UV-B Radiation Intensities on Metabolic Characteristics of Porphyra haitanensis. International Journal of Molecular Sciences. 2021; 22(4):2180. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22042180
Chicago/Turabian StyleFu, Shimei, Song Xue, Jun Chen, Shuai Shang, Hui Xiao, Yu Zang, and Xuexi Tang. 2021. "Effects of Different Short-Term UV-B Radiation Intensities on Metabolic Characteristics of Porphyra haitanensis" International Journal of Molecular Sciences 22, no. 4: 2180. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22042180