
Development of Antimicrobial Phototreatment Tolerance: Why the Methodology Matters

 ,
,
Abstract
:1. Introduction
- -
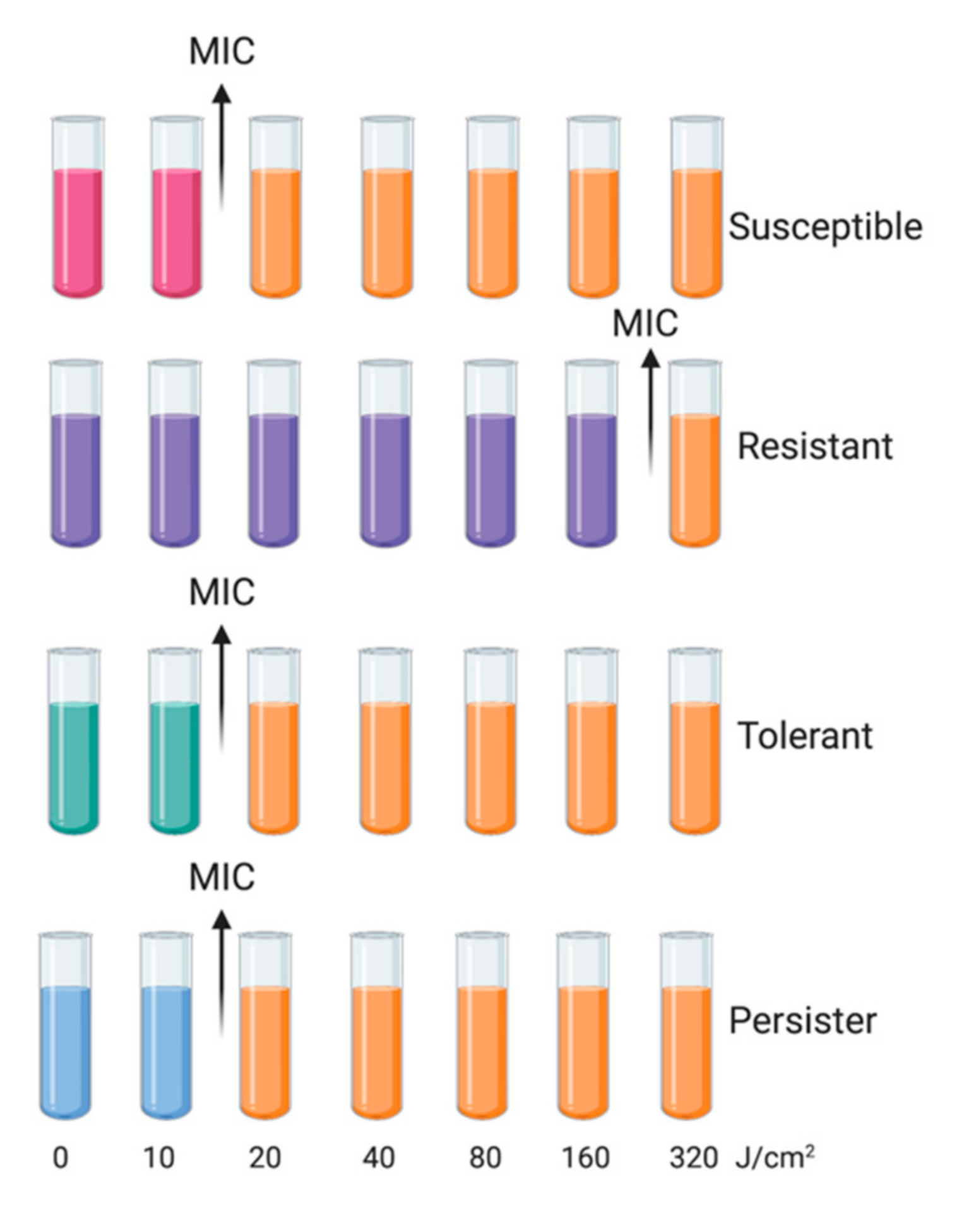
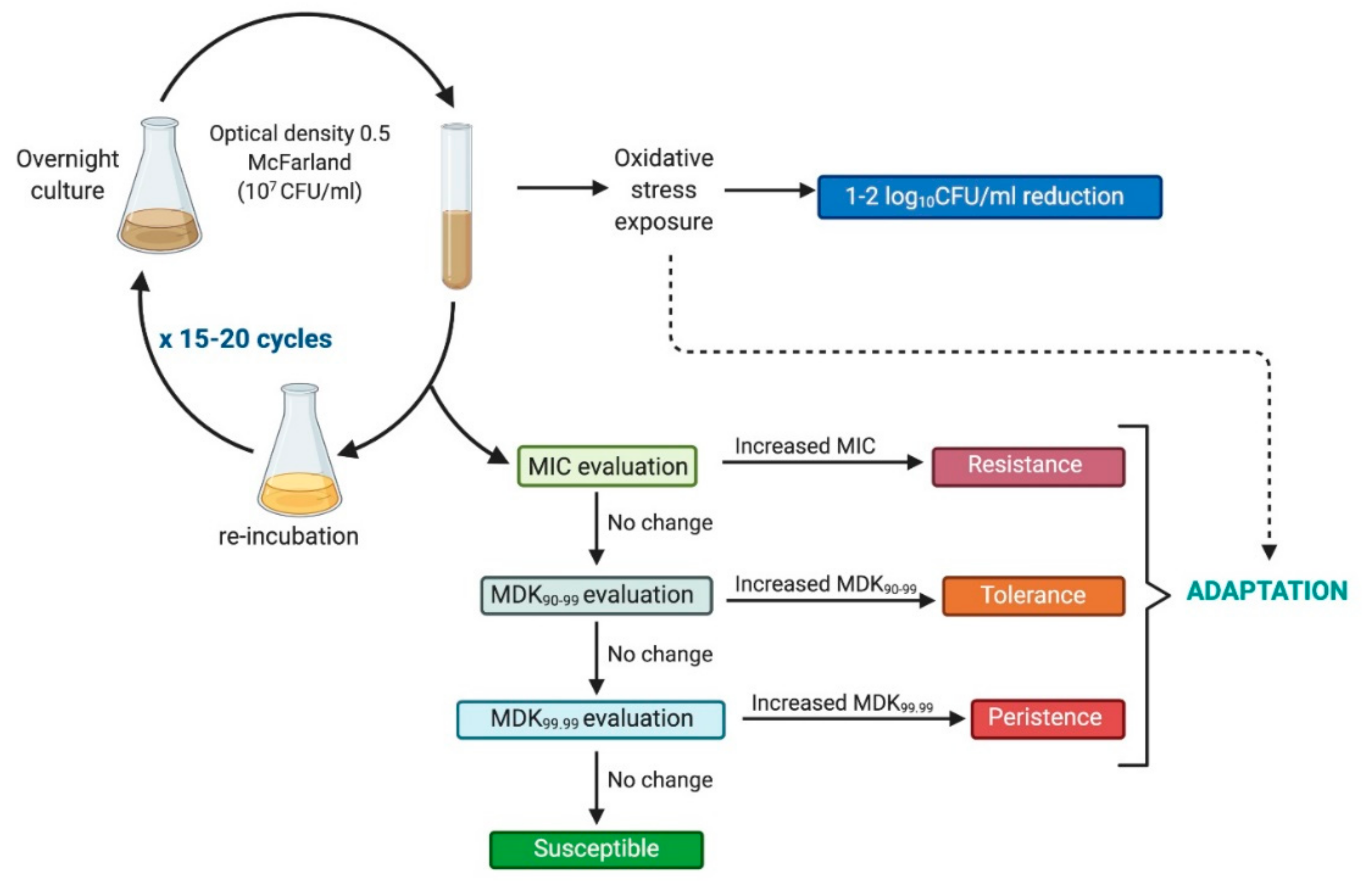
- Resistance is an acquired and inherited decline in the effectiveness of a given treatment, resulting in the need for high concentrations of a photosensitizing agent and/or longer exposure to the treatment; it should be a stable feature, observed in the next consecutive cycles. In the case of phototreatments, due to the unspecific mechanism, this is rare, and has not been observed to date.
- -
- Tolerance is an acquired stable feature, whereby longer minimum treatment duration (e.g., irradiation time) is needed to achieve the same killing efficacy regardless of the concentration of the photosensitizing agent; it is characterized by being stable and, thus, is observed in subsequent consecutive cycles.
- -
- Persistence is a nonheritable and dormant phenotypic state (transient tolerance) represented by a small subpopulation (about 0.1–1%) of bacterial cells that are killed at a slower rate than susceptible cells [17]; it can be observed as unstable tolerance, occurring in a few cycles of photoinactivation and vanishing in subsequent cycles.
- (i)
- Inoculation from a single surviving colony: This procedure is burdened with the risk of very low probability of detecting tolerance or resistance due to the fact that the majority of surviving bacterial cells that could carry genetic alterations are omitted and not included in the next cycle. Single surviving colonies were used as inoculation sources for the next cycle in numerous studies [19,24,27,29,30,32]. However, few studies have described the use of treated suspensions for re-inoculation [20,26,28,31,33,35].
- (ii)
- Application of lethal rather than sublethal doses or irradiation times longer than the minimal duration for killing 99% of cells (MDK99): Too-high doses of light-based treatment could lead to irreversible changes in bacterial cells and contribute to the state that most bacterial cells would not be able to recover and form tolerant/resistant phenotypes. Most of the published studies describe the use of lethal instead of sublethal conditions [19,20,21,25,29,32,36]. Sublethal treatment was applied in just a few studies [18,24,26,27,28,30].
- (iii)
- Too few consecutive passages (<15 cycles).
- (iv)
- Lack of verification regarding whether the adaptation is stable and the change has a genetic basis or is due to persistence. In a study by Zhang et al. (2016), reduced aBL susceptibility with increasing number of cycles (fourth and fifth passages) was observed in Candida albicans. However, there was no statistically significant difference in the post-aBL survival rate of C. albicans between the first and last passage (P > 0.05). Leanse et al. (2018) observed an unstable decrease in aBL efficacy in A. baumannii in the 9th, 16th, and 17th cycles. The temporary, unstable reduction of susceptibility observed in these two studies may indicate the appearance of persister cells due to oxidative stress and phenotype switching [12]; thus, it is necessary to validate whether the observed decrease in treatment susceptibility is a stable feature.
- (v)
- Lack of untreated control, in order to exclude the phenomenon of naturally occurring mutations due to cell aging with an increasing number of passages.
- -
- -
- -
- -

2. State of the Art
2.1. Antimicrobial Blue Light (aBL)
2.2. Antimicrobial Photodynamic Inactivation (aPDI)
2.2.1. Xanthene Photosensitizers
2.2.2. Phenothiazine Photosensitizers
2.2.3. Phthalocyanines
2.2.4. Porphyrins and Porphyrin Derivatives
2.2.5. Other Photosensitizing Compounds

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ref. | Species | PS | Light Source | Methodology | Log10 Reduction | Inoculation Source | Tolerance |
|---|---|---|---|---|---|---|---|
| Bacteria | |||||||
| [36] | P. micros A. actinomycetemcomitans | GlamTMPn BOTHMPn | 4 × 250 W tungsten lamps | 10 cycles | Lethal a | Solid agar plates | No |
| [18] | E. coli S. aureus | MB | Non-thermal diode laser, 670 nm | 11 cycles (E. coli) 25 cycles (S. aureus) | Sublethal | Surviving colonies from previous cycle | No |
| [19] | V. fischeri E. coli | Tri-Py+-Me-PF | 13 × 18W OSRAM 21 lamps, 380–700 nm | 10 cycles | Lethal | Single colony survivors from previous treatment used further as overnight culture | No |
| [20] | S. aureus P. aeruginosa | RLP068/Cl | Non-coherent halogen lamp, 600–700 nm | 20 cycles | Lethal | From each sample, 10 µL was subcultured to perform subsequent cycles | No |
| [23] | S. aureus | - | SLD light probe/405 nm | 7 cycles | No data | Irradiated cells were growing in solid medium, then subcultured on another fresh medium and finally used for next experimental stage | Yes |
| [22] | S. aureus | - | Tri-wave light ultrasound device, 464 nm, 850 nm; SLD light probe, 405 nm | 7 cycles | No data | Irradiated cells grown in solid medium then subcultured on another fresh medium and used for next experimental stage | Yes |
| [24] | A. baumannii | - | Omnilux clear-U light- emitting diode array, 415 nm | 10 cycles | Sublethal | Surviving bacterial cells from agar were collected and recultured for next cycle | No |
| [26] | P. aeruginosa | - | LED, 415 nm | Irradiation of Petri dish containing bacterial suspension (10 cycles) | Sublethal and lethal | Treated suspension | No (after 9th cycle surviving fraction was increased by 2 log10) |
| [27] | S. aureus | - | LED, 405 nm | 15 cycles | Sublethal | Surviving colonies | No |
| [28] | S. aureus E. coli | ZnTnHex-2-PyP | Overhead projector OHP-3100 p, broad spectrum | Continuous growth under sublethal conditions for 48 h (10–20 cycles) | Sublethal | Treated suspension | No |
| [29] | P. aeruginosa A. baumannii E. coli | - | LED, 405 nm | 20 cycles | Lethal | Single surviving colony | No (increased unstable aBL tolerance in 9th, 16th, and 17th cycles) |
| P. aeruginosa (mouse skin abrasion wounds) | - | - | 5 cycles | Sublethal | - | No | |
| [30] | E. faecalis | Ce6 MB | No data | 4 cycles | Sublethal and lethal | Single surviving colony from agar plate | No |
| [31] | S. aureus | RB - | LED, 515 nm LED, 411 nm | 15 cycles | Sublethal | Treated suspension | Stable tolerance to RB-aPDI and aBL |
| [33] | S. agalactiae | RB | LED, 515 nm | 15 cycles | Sublethal | Treated suspension | Stable tolerance |
| [32] | S. aureus E. coli | Zn-TBut4PyP | Tungsten lamp, 320–780 nm | 10 cycles | Lethal | Single surviving colony | No |
| [35] | S. aureus | MB | Broadband visible light, 575–700 nm | 7 cycles | Sublethal | Treated suspension | Stable tolerance (cross-tolerance to TBO-aPDI) |
| Viruses | |||||||
| [21] | T4-like phage | Tri-Py+-Me-PF | 13 OSRAM 21 fluorescent lamps (18 W), 380–700 nm | 10 cycles | Lethal | Phage suspension prepared from previous experimental cycle sample | No |
| Yeast | |||||||
| [25] | C. albicans | - | LED, 415 nm | 10 cycles | Lethal | Surviving cells | Reduced aBL susceptibility with increasing number of cycles |
| [20] | C. albicans | RLP068/Cl | Non-coherent halogen lamp, 600–700 nm | 20 cycles | Lethal | From each sample, 10 µL was subcultured to perform subsequent cycles | No |
2.3. Pulsed Light
2.4. Cold Atmospheric Plasma (CAP)
| Ref. | Species | Plasma Source | Methodology | Log10 Reduction | Inoculation Source | Tolerance |
|---|---|---|---|---|---|---|
| [54] | E. coli E. mundtii | Plasma device (HandPlaSter) | 4 cycles | Lethal | Surviving cells on solid medium | No |
| [58] | S. aureus | Radio frequency plasma pen (kinpen09®) | 6 repetitions for period of 6 h | No data | Same medium with microorganisms for all experimental procedures | No |
| [50] | S. aureus (MRSA) P. aeruginosa | Radiofrequency source with helium as working gas | 6 cycles | Sublethal (S. aureus) Lethal (P. aeruginosa) | Same bacterial suspension for all cycles | No |
2.5. Ultraviolet (UV) Light
| Ref. | Species | Light Source | Methodology | Log10 Reduction | Inoculation Source | Tolerance |
|---|---|---|---|---|---|---|
| Bacteria | ||||||
| [64] | E. coli | 15 W Hg vapor UV germicidal lamp, 254 nm | 80 cycles | Initially sublethal (increasing 2-fold every 10 cycles) | Cells after irradiation inoculated in fresh LB medium | Yes (resistance) |
| [46] | L. monocytogenes | Xenon flash lamp, spectrum from UV-C to IR | 13 cycles | No data/lethal | Single colony survivors on solid medium plate used for preparation of inoculum for next experimental cycle | No |
| [49] | E. coli C. jejuni L. monocytogenes | Xenon flash lamp, UVC-UV-IR | 20 cycles | Sublethal | Cocktail of strains after PL treatment incubated in fresh medium | Yes (resistance) |
| [65] | E. coli | UVP Chromato-Vue TM-36 transilluminator (UVP Inc., Upland, CA, USA), 302 nm | 60 cycles | Sublethal | LB agar plates with surviving colonies | Yes (resistance) |
| [47] | P. aeruginosa | 4 × xenon flash lamps, white (200–1100 nm) and UV (200–400 nm) light | 1 cycle (2 treatments) | Sublethal/ lethal | None | Yes |
| [48] | L. monocytogenes L. innocua E. coli | Xenon flash lamp (SteriPulse system), 200–1100 nm | 10 cycles | No data | Single colony survivors replated on fresh solid medium and used for preparation of inoculum for next cycle | No |
| [44] | L. monocytogenes | SteriPulse-XL RS-3000C (xenon) pulsed light device | 20 cycles | Lethal | TSA + YE plates with single colony survivors used for inoculation | Not stable tolerance (declined after deep-freeze storage) |
| Viruses | ||||||
| [66] | Bacteriophage T7 | UVP® transilluminator (bulbs, UVP 34-0042-01) | 30 cycles | Lethal | A lysate of phage survivors added to a culture of cells and grown to culture lysis | Yes (resistance) |
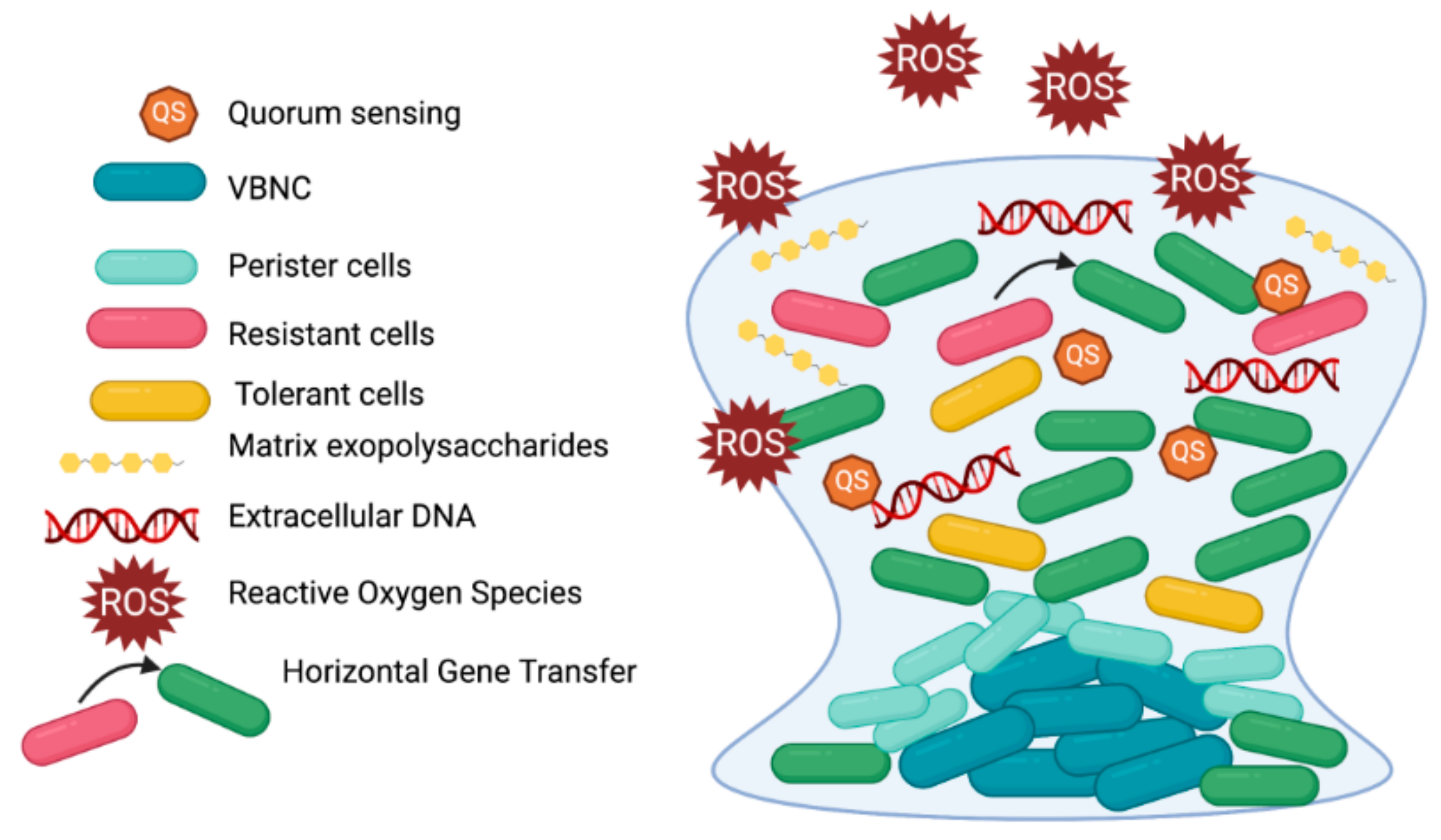
3. Biofilm Tolerance
4. Discussion
- -
- the subculture originated from the treated suspension (not from a single surviving colony),
- -
- the treatment condition resulted in the reduction in viable counts that left sufficient survivors for tolerance development (approx. 1–2 log10 units reduction),
- -
- the experiment was conducted up to 7–15 cycles,
- -
- phenotypic stability testing was performed.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- English, B.K.; Gaur, A.H. The use and abuse of antibiotics and the development of antibiotic resistance. In Hot Topics in Infection and Immunity in Children VI; Springer: New York, NY, USA, 2010; pp. 73–82. [Google Scholar]
- Enwemeka, C.S. Antimicrobial blue light: An emerging alternative to antibiotics. Photomed Laser Surg. 2013, 31, 509–511. [Google Scholar] [CrossRef]
- O’Neill, J. Tackling Drug-resistant Infections Globally: Final Report and Recommendations-The Review on Antimicrobial Resistance Chaired by Jim O’Neill; Government WTaH: London, UK, 2016. [Google Scholar]
- Bush, K.; Courvalin, P.; Dantas, G.; Davies, J.; Eisenstein, B.; Huovinen, P.; Jacoby, G.A.; Kishony, R.; Kreiswirth, B.N.; Kutter, E.; et al. Tackling antibiotic resistance. Nat. Rev. Microbiol. 2011, 9, 894–896. [Google Scholar] [CrossRef]
- Brauner, A.; Fridman, O.; Gefen, O.; Balaban, N.Q. Distinguishing between resistance, tolerance and persistence to antibiotic treatment. Nat. Rev. Microbiol. 2016, 14, 320–330. [Google Scholar] [CrossRef]
- Safety SCoC. Opinion on Triclosan Antimicrobial Resistance European Commission; European Commission: Brussels, Belgium, 2010. [Google Scholar]
- Kester, J.C.; Fortune, S.M. Persisters and beyond: Mechanisms of phenotypic drug resistance and drug tolerance in bacteria. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 91–101. [Google Scholar] [CrossRef]
- Charpentier, E.; Tuomanen, E. Mechanisms of antibiotic resistance and tolerance in Streptococcus pneumoniae. Microbes Infect. 2000, 2, 1855–1864. [Google Scholar] [CrossRef]
- Fridman, O.; Goldberg, A.; Ronin, I.; Shoresh, N.; Balaban, N.Q. Optimization of lag time underlies antibiotic tolerance in evolved bacterial populations. Nature 2014, 513, 418–421. [Google Scholar] [CrossRef]
- Balaban, N.Q.; Merrin, J.; Chait, R.; Kowalik, L.; Leibler, S. Bacterial persistence as a phenotypic switch. Science 2004, 305, 1622–1625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helaine, S.; Kugelberg, E. Bacterial persisters: Formation, eradication, and experimental systems. Trends Microbiol. 2014, 22, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Grant, S.S.; Hung, D.T. Persistent bacterial infections, antibiotic tolerance, and the oxidative stress response. Virulence 2013, 4, 273–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuchscherr, L.; Medina, E.; Hussain, M.; Völker, W.; Heitmann, V.; Niemann, S.; Holzinger, D.; Roth, J.; Proctor, R.A.; Becker, K.; et al. Staphylococcus aureus phenotype switching: An effective bacterial strategy to escape host immune response and establish a chronic infection. EMBO Mol. Med. 2011, 3, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Starkey, M.; Hickman, J.H.; Ma, L.; Zhang, N.; De Long, S.; Hinz, A.; Palacios, S.; Manoil, C.; Kirisits, M.J.; Starner, T.D.; et al. Pseudomonas aeruginosa rugose small-colony variants have adaptations that likely promote persistence in the cystic fibrosis lung. J. Bacteriol. 2009, 191, 3492–3503. [Google Scholar] [CrossRef] [Green Version]
- Imlay, J.A.; Linn, S. Mutagenesis and stress responses induced in Escherichia coli by hydrogen peroxide. J. Bacteriol. 1987, 169, 2967–2976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Painter, K.L.; Strange, E.; Parkhill, J.; Bamford, K.B.; Armstrong-James, D.; Edwards, A.M. Staphylococcus aureus adapts to oxidative stress by producing H2O2-resistant small-colony variants via the SOS response. Infect. Immun. 2015, 83, 1830–1844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gefen, O.; Balaban, N.Q. The importance of being persistent: Heterogeneity of bacterial populations under antibiotic stress. FEMS Microbiol. Rev. 2009, 33, 704–717. [Google Scholar] [CrossRef] [PubMed]
- Pedigo, L.A.; Gibbs, A.J.; Scott, R.J.; Street, C.N. Absence of bacterial resistance following repeat exposure to photodyanmic therapy. In Photodynamic Therapy: Back to the Future; SPIE: Bellingham, WA, USA, 2009; Volume 7380. [Google Scholar]
- Tavares, A.; Carvalho, C.; Faustino, M.A.; Neves, M.G.; Tomé, J.P.; Tomé, A.C.; Cavaleiro, J.A.; Cunha, Â.; Gomes, N.; Alves, E.; et al. Antimicrobial photodynamic therapy: Study of bacterial recovery viability and potential development of resistance after treatment. Mar. Drugs 2010, 8, 91–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giuliani, F.; Martinelli, M.; Cocchi, A.; Arbia, D.; Fantetti, L.; Roncucci, G. In vitro resistance selection studies of RLP068/Cl, a new Zn(II) phthalocyanine suitable for antimicrobial photodynamic therapy. Antimicrob. Agents Chemother. 2010, 54, 637–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, L.; Tomé, J.P.; Neves, M.G.; Tomé, A.C.; Cavaleiro, J.A.; Faustino, M.A.; Cunha, Â.; Gomes, N.C.; Almeida, A. Evaluation of resistance development viability recovery by a non-enveloped virus after repeated cycles of, aPDT. Antivir. Res. 2011, 91, 278–282. [Google Scholar] [CrossRef]
- Guffey, J.S.; Payne, W.; Martin, K.; Dodson, C. Delaying the Onset of Resistance Formation: Effect of Manipulating Dose, Wavelength, and Rate of Energy Delivery of 405-, 464-, and 850-Nanometer Light for Staphylococcus aureus. Wounds 2014, 26, 95–100. [Google Scholar]
- Guffey, J.S.; Payne, W.; Jones, T.; Martin, K. Evidence of resistance development by Staphylococcus aureus to an in vitro, multiple stage application of 405 nm light from a supraluminous diode array. Photomed. Laser Surg. 2013, 31, 179–182. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhu, Y.; Gupta, A.; Huang, Y.; Murray, C.K.; Vrahas, M.S.; Sherwood, M.E.; Baer, D.G.; Hamblin, M.R.; Dai, T. Antimicrobial blue light therapy for multidrug-resistant Acinetobacter baumannii infection in a mouse burn model: Implications for prophylaxis and treatment of combat-related wound infections. J. Infect. Dis. 2014, 209, 1963–1971. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhu, Y.; Chen, J.; Wang, Y.; Sherwood, M.E.; Murray, C.K.; Vrahas, M.S.; Hooper, D.C.; Hamblin, M.R.; Dai, T. Antimicrobial blue light inactivation of Candida albicans: In vitro and in vivo studies. Virulence 2016, 7, 536–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amin, R.M.; Bhayana, B.; Hamblin, M.R.; Dai, T. Antimicrobial blue light inactivation of Pseudomonas aeruginosa by photo-excitation of endogenous porphyrins: In vitro and in vivo studies. Lasers Surg. Med. 2016, 48, 562–568. [Google Scholar] [CrossRef] [Green Version]
- Tomb, R.M.; Maclean, M.; Coia, J.E.; MacGregor, S.J.; Anderson, J.G. Assessment of the potential for resistance to antimicrobial violet-blue light in Staphylococcus aureus. Antimicrob. Resist. Infect. Control 2017, 6, 100. [Google Scholar] [CrossRef] [Green Version]
- Al-Mutairi, R.; Tovmasyan, A.; Batinic-Haberle, I.; Benov, L. Sublethal Photodynamic Treatment Does Not Lead to Development of Resistance. Front. Microbiol. 2018, 9, 1699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leanse, L.G.; Harrington, O.D.; Fang, Y.; Ahmed, I.; Goh, X.S.; Dai, T. Evaluating the Potential for Resistance Development to Antimicrobial Blue Light (at 405 nm) in Gram-Negative Bacteria: In vitro and in vivo Studies. Front. Microbiol. 2018, 9, 2403. [Google Scholar] [CrossRef] [Green Version]
- de Freitas, L.M.; Blanco, A.L.; Fontana, C.R. Antimicrobial photodynamic therapy proved not to induce bacterial resistance. In Light-Based Diagnosis and Treatment of Infectious Diseases; SPIE: Bellingham, WA, USA, 2018; Volume 10479. [Google Scholar]
- Rapacka-Zdonczyk, A.; Wozniak, A.; Pieranski, M.; Woziwodzka, A.; Bielawski, K.P.; Grinholc, M. Development of Staphylococcus aureus tolerance to antimicrobial photodynamic inactivation and antimicrobial blue light upon sub-lethal treatment. Sci. Rep. 2019, 9, 9423. [Google Scholar] [CrossRef] [Green Version]
- Paronyan, M.H.; Koloyan, H.O.; Avetisyan, S.V.; Aganyants, H.A.; Hovsepyan, A.S. Study of the possible development of bacterial resistance to photodynamic inactivation. Biol. J. Armen. 2019, 71, 17–22. [Google Scholar]
- Pieranski, M.; Sitkiewicz, I.; Grinholc, M. Increased photoinactivation stress tolerance of Streptococcus agalactiae upon consecutive sublethal phototreatments. Free Radic. Biol. Med. 2020, 160, 657–669. [Google Scholar] [CrossRef] [PubMed]
- Kashef, N.; Hamblin, M.R. Can microbial cells develop resistance to oxidative stress in antimicrobial photodynamic inactivation? Drug Resist. Updat. 2017, 31, 31–42. [Google Scholar] [CrossRef]
- Snell, S.B.; Gill, A.L.; Haidaris, C.G.; Foster, T.H.; Baran, T.M.; Gill, S.R. Staphylococcus aureus Tolerance and Genomic Response to Photodynamic Inactivation. Msphere 2021, 6. [Google Scholar] [CrossRef]
- Lauro, F.M.; Pretto, P.; Covolo, L.; Jori, G.; Bertoloni, G. Photoinactivation of bacterial strains involved in periodontal diseases sensitized by porphycene-polylysine conjugates. Photochem. Photobiol. Sci. 2002, 1, 468–470. [Google Scholar] [CrossRef]
- Ruiz, G.T.T.; Nelson, E.; Sparks, L.; Langland, J. Bacterial development of resistance to botanical antimicrobials. J. Evol. Health 2017, 2. [Google Scholar] [CrossRef] [Green Version]
- Wesgate, R.; Grasha, P.; Maillard, J.Y. Use of a predictive protocol to measure the antimicrobial resistance risks associated with biocidal product usage. Am. J. Infect. Control 2016, 44, 458–464. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, S.; Horinouchi, T.; Furusawa, C. Acceleration and suppression of resistance development by antibiotic combinations. BMC Genom. 2017, 18, 328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brook, I.; Elliott, T.B.; Pryor, H.I., II; Sautter, T.E.; Gnade, B.T.; Thakar, J.H.; Knudson, G.B. In vitro resistance of Bacillus anthracis Sterne to doxycycline, macrolides and quinolones. Int. J. Antimicrob. Agents 2001, 18, 559–562. [Google Scholar] [CrossRef]
- Knapp, L.; Amezquita, A.; McClure, P.; Stewart, S.; Maillard, J.Y. Development of a protocol for predicting bacterial resistance to microbicides. Appl Environ. Microbiol. 2015, 81, 2652–2659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, T.; Gupta, A.; Murray, C.K.; Vrahas, M.S.; Tegos, G.P.; Hamblin, M.R. Blue light for infectious diseases: Propionibacterium acnes, Helicobacter pylori, and beyond? Drug Resist. Updat. 2012, 15, 223–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wang, Y.; Wang, Y.; Murray, C.K.; Hamblin, M.R.; Hooper, D.C.; Dai, T. Antimicrobial blue light inactivation of pathogenic microbes: State of the art. Drug Resist. Updat. 2017, 33, 1–22. [Google Scholar]
- Heinrich, V.; Zunabovic, M.; Petschnig, A.; Müller, H.; Lassenberger, A.; Reimhult, E.; Kneifel, W. Previous Homologous and Heterologous Stress Exposure Induces Tolerance Development to Pulsed Light in Listeria monocytogenes. Front. Microbiol. 2016, 7, 490. [Google Scholar] [CrossRef]
- Bhavya, M.L.; Umesh Hebbar, H. Pulsed light processing of foods for microbial safety. Food Qual. Saf. 2017, 1, 187–201. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Lopez, V.M.; Devlieghere, F.; Bonduelle, V.; Debevere, J. Factors affecting the inactivation of micro-organisms by intense light pulses. J. Appl. Microbiol. 2005, 99, 460–470. [Google Scholar] [CrossRef] [PubMed]
- Massier, S.; Rince, A.; Maillot, O.; Feuilloley, M.G.; Orange, N.; Chevalier, S. Adaptation of Pseudomonas aeruginosa to a pulsed light-induced stress. J. Appl. Microbiol. 2012, 112, 502–511. [Google Scholar] [CrossRef] [PubMed]
- Uesugi, A.R.; Hsu, L.; Moraru, C.I. Effect of pulsed light treatments on the growth and resistance behavior of Listeria monocytogenes 10403S, Listeria innocua, and Escherichia coli ATCC 25922 in a liquid substrate. J. Food Prot. 2013, 76, 435–439. [Google Scholar] [CrossRef] [PubMed]
- Rajkovic, A.; Smigic, N.; Uyttendaele, M.; Medic, H.; de Zutter, L.; Devlieghere, F. Resistance of Listeria monocytogenes, Escherichia coli O157:H7 and Campylobacter jejuni after exposure to repetitive cycles of mild bactericidal treatments. Food Microbiol. 2009, 26, 889–895. [Google Scholar] [CrossRef]
- Brun, P.; Bernabè, G.; Marchiori, C.; Scarpa, M.; Zuin, M.; Cavazzana, R.; Zaniol, B.; Martines, E. Antibacterial efficacy and mechanisms of action of low power atmospheric pressure cold plasma: Membrane permeability, biofilm penetration and antimicrobial sensitization. J. Appl. Microbiol. 2018, 125, 398–408. [Google Scholar] [CrossRef]
- Theinkom, F.; Singer, L.; Cieplik, F.; Cantzler, S.; Weilemann, H.; Cantzler, M.; Karl-Anton Hiller Maisch, T.; Julia, L.; Zimmermann, J.L. Antibacterial efficacy of cold atmospheric plasma against Enterococcus faecalis planktonic cultures and biofilms in vitro. PLoS ONE 2019, 14, e0223925. [Google Scholar] [CrossRef] [Green Version]
- Kong, M.G.; Kroesen, G.; Morfill, G.; Nosenko, T.; Shimizu, T.; Van Dijk, J.; Zimmermann, J.L. Plasma medicine: An introductory review. New J. Phys. 2009, 11, 115012. [Google Scholar] [CrossRef]
- Dijksteel, G.S.; Ulrich, M.M.W.; Vlig, M.; Sobota, A.; Middelkoop, E.; Boekema, B.K.H.L. Safety and bactericidal efficacy of cold atmospheric plasma generated by a flexible surface Dielectric Barrier Discharge device against Pseudomonas aeruginosa in vitro and in vivo. Ann. Clin. Microbiol. Antimicrob. 2020, 19, 37. [Google Scholar] [CrossRef]
- Zimmermann, J.L.; Shimizu, T.; Schmidt, H.U.; Li, Y.F.; Morfill, G.E.; Isbary, G. Test for bacterial resistance build-up against plasma treatment. New J. Phys. 2012, 14, 073037. [Google Scholar] [CrossRef] [Green Version]
- Tseng, S.; Abramzon, N.; Jackson, J.O.; Lin, W.J. Gas discharge plasmas are effective in inactivating Bacillus and Clostridium spores. Appl. Microbiol. Biotechnol. 2012, 93, 2563–2570. [Google Scholar] [CrossRef]
- Maisch, T.; Shimizu, T.; Li, Y.F.; Heinlin, J.; Karrer, S.; Morfill, G.; Zimmermann, J.L. Decolonisation of MRSA, S. aureus and E. coli by Cold-Atmospheric Plasma Using a Porcine Skin Model In Vitro. PLoS ONE 2012, 7, e34610. [Google Scholar]
- Beyrer, M.; Smeu, I.; Martinet, D.; Howling, A.; Pina-Pérez, M.C.; Ellert, C. Cold Atmospheric Plasma Inactivation of Microbial Spores Compared on Reference Surfaces and Powder Particles. Food Bioprocess Technol. 2020, 13, 827–837. [Google Scholar] [CrossRef]
- Matthes, R.; Assadian, O.; Kramer, A. Repeated applications of cold atmospheric pressure plasma does not induce resistance in Staphylococcus aureus embedded in biofilms. GMS Hyg. Infect. Control 2014, 9, Doc17. [Google Scholar] [PubMed]
- Visser, P.M.; Poos, J.J.; Scheper, B.B.; Boelen, P.; van Duyl, F.C. Diurnal variations in depth profiles of UV—Induced DNA damage and inhibition of bacterioplankton production in tropical coastal waters. Mar. Ecol. Prog. Ser. 2002, 228, 25–33. [Google Scholar] [CrossRef]
- Sinha, R.P.; Häder, D. UV—Induced DNA damage and repair: A review. Photo-Chem. Photobiol. Sci. 2002, 1, 225–236. [Google Scholar] [CrossRef]
- Lindahl, T.; Wood, R.D. Quality control by DNA repair. Science 1999, 286, 1897–1905. [Google Scholar] [CrossRef]
- Li, G.Q.; Wang, W.; Huo, Z.; Lu, Y.; Hu, H. Comparison of UV--LED and low pressure UV for water disinfection: Photoreactivation and dark repair of Escherichia coli. Water Res. 2017, 126, 134–143. [Google Scholar] [CrossRef]
- Green, A.; Popovi, V.; Pierscianowski, J.; Biancaniello, M.; Warriner, K.; Koutchma, T. Inactivation of Escherichia coli, Listeriaand Salmonella by single and multiple wavelength ultraviolet--light emitting diodes. Innov. Food Sci. Emerg. Technol. 2018, 47, 353–361. [Google Scholar] [CrossRef]
- Alcantara-Diaz, D.; Brena-Valle, M.; Serment-Guerrero, J. Divergent adaptation of Escherichia coli to cyclic ultraviolet light exposures. Mutagenesis 2004, 19, 349–354. [Google Scholar] [CrossRef]
- Goldman, R.P.; Travisano, M. Experimental evolution of ultraviolet radiation resistance in Escherichia coli. Evolution 2011, 65, 3486–3498. [Google Scholar] [CrossRef]
- Tom, E.F.; Molineux, I.J.; Paff, M.L.; Bull, J.J. Experimental evolution of UV resistance in a phage. PeerJ 2018, 6, e5190. [Google Scholar] [CrossRef]
- Cieplik, F.; Tabenski, L.; Buchalla, W.; Maisch, T. Antimicrobial photodynamic therapy for inactivation of biofilms formed by oral key pathogens. Front. Microbiol. 2014, 5, 405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceri, H.; Olson, M.E.; Stremick, C.; Read, R.R.; Morck, D.; Buret, A. The Calgary Biofilm Device: New technology for rapid determination of antibiotic susceptibilities of bacterial biofilms. J. Clin. Microbiol. 1999, 37, 1771–1776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shani, S.; Friedman, M.; Steinberg, D. The anticariogenic effect of amine fluorides on Streptococcus sobrinus and glucosyltransferase in biofilms. Caries Res. 2000, 34, 260–267. [Google Scholar] [CrossRef]
- Gulías, Ò.; McKenzie, G.; Bayó, M.; Agut, M.; Nonell, S. Effective photodynamic inactivation of 26 Escherichia coli strains with different antibiotic susceptibility profiles: A planktonic and biofilm study. Antibiotics 2020, 9, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, P.S.; Costerton, J.W. Antibiotic resistance of bacteria in biofilms. Lancet 2001, 358, 135–138. [Google Scholar] [CrossRef]
- Kolter, R.; Greenberg, E.P. Microbial sciences: The superficial life of microbes. Nature 2006, 441, 300–302. [Google Scholar] [CrossRef]
- Boles, B.R.; Thoendel, M.; Singh, P.K. Self-generated diversity produces “insurance effects” in biofilm communities. Proc. Natl. Acad. Sci. USA 2004, 101, 16630–16635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shemesh, M.; Tam, A.; Steinberg, D. Differential gene expression profiling of Streptococcus mutans cultured under biofilm and planktonic conditions. Microbiology 2007, 153, 1307–1317. [Google Scholar] [CrossRef] [Green Version]
- Lo, A.W.; Seers, C.A.; Boyce, J.D.; Dashper, S.G.; Slakeski, N.; Lissel, J.P.; Reynolds, E.C. Comparative transcriptomic analysis of Porphyromonas gingivalis biofilm and planktonic cells. BMC Microbiol. 2009, 9, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waters, E.M.; Rowe, S.E.; O’Gara, J.P.; Conlon, B.P. Convergence of Staphylococcus aureus persister and biofilm research: Can biofilms be defined as communities of adherent persister cells? PLoS Pathog. 2016, 12, e1006012. [Google Scholar] [CrossRef]
- Stewart, P.S.; Franklin, M.J. Physiological heterogeneity in biofilms. Nat. Rev. Microbiol. 2008, 6, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Hall, C.W.; Mah, T.F. Molecular mechanisms of biofilm-based antibiotic resistance and tolerance in pathogenic bacteria. FEMS Microbiol. Rev. 2017, 41, 276–301. [Google Scholar] [CrossRef] [PubMed]
- Balaure, P.C.; Grumezescu, A.M. Recent advances in surface nanoengineering for biofilm prevention and control. Part I: Molecular basis of biofilm recalcitrance. passive anti-biofouling nanocoatings. Nanomaterials 2020, 10, 1230. [Google Scholar] [CrossRef]
- Mah, T.F.C.; O’Toole, G.A. Mechanisms of biofilm resistance to antimicrobial agents. Trends Microbiol. 2001, 9, 34–39. [Google Scholar] [CrossRef]
- Stewart, P.S. Antimicrobial tolerance in biofilms. Microbial Biofilms. 2015, 269–285. [Google Scholar] [CrossRef] [Green Version]
- Lebeaux, D.; Ghigo, J.M.; Beloin, C. Biofilm-related infections: Bridging the gap between clinical management and fundamental aspects of recalcitrance toward antibiotics. Microbiol. Mol. Biol. Rev. 2014, 78, 510–543. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.S.; Heo, P.; Yang, T.J.; Lee, K.S.; Jin, Y.S.; Kim, S.K.; Kweon, D.H. Bacterial persisters tolerate antibiotics by not producing hydroxyl radicals. Biochem. Biophys. Res. Commun. 2011, 413, 105–110. [Google Scholar] [CrossRef]
- Stewart, P.S.; Davison, W.M.; Steenbergen, J.N. Daptomycin rapidly penetrates a Staphylococcus epidermidis biofilm. Antimicrob. Agents Chemother. 2009, 53, 3505–3507. [Google Scholar] [CrossRef] [Green Version]
- Spoering, A.L.; Lewis, K.I.M. Biofilms and planktonic cells of Pseudomonas aeruginosa have similar resistance to killing by antimicrobials. J. Bacteriol. 2001, 183, 6746–6751. [Google Scholar] [CrossRef] [Green Version]
- Ryder, V.J.; Ian, C.; O’Neill, A.J. Increased mutability of Staphylococci in biofilms as a consequence of oxidative stress. PLoS ONE 2012, 7, e47695. [Google Scholar] [CrossRef] [Green Version]
- Driffield, K.; Miller, K.; Bostock, J.M.; O’neill, A.J.; Chopra, I. Increased mutability of Pseudomonas aeruginosa in biofilms. J. Antimicrob. Chemother. 2008, 61, 1053–1056. [Google Scholar] [CrossRef] [Green Version]
- Boles, B.R.; Singh, P.K. Endogenous oxidative stress produces diversity and adaptability in biofilm communities. Proc. Natl. Acad. Sci. USA 2008, 105, 12503–12508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakubowski, W.; Walkowiak, B. Resistance of Oxidative Stress in Biofilm and Planktonic Cells. Arch. Biol. Technol. 2015, 58, 300–308. [Google Scholar] [CrossRef] [Green Version]
- Savage, V.J.; Chopra, I.; O’Neill, A.J. Staphylococcus aureus biofilms promote horizontal transfer of antibiotic resistance. Antimicrob. Agents Chemother. 2013, 57, 1968–1970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cook, L.C.; Dunny, G.M. Effects of biofilm growth on plasmid copy number and expression of antibiotic resistance genes in Enterococcus faecalis. Antimicrob. Agents Chemother. 2013, 57, 850–1856. [Google Scholar] [CrossRef] [Green Version]
- Strugeon, E.; Tilloy, V.; Ploy, M.C.; Da Re, S. The stringent response promotes antibiotic resistance dissemination by regulating integron integrase expression in biofilms. MBio 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Cieplik, F.; Späth, A.; Regensburger, J.; Gollmer, A.; Tabenski, L.; Hiller, K.A.; Bäumler, W.; Maisch, T.; Schmalz, G. Photodynamic biofilm inactivation by SAPYR--an exclusive singlet oxygen photosensitizer. Free Radic. Biol. Med. 2013, 65, 477–487. [Google Scholar] [CrossRef] [Green Version]
- Biel, M.A.; Pedigo, L.; Gibbs, A.; Loebel, N. Photodynamic therapy of antibiotic-resistant biofilms in a maxillary sinus model. Int. Forum Allergy Rhinol. 2013, 3, 468–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Zhou, Y.; Wang, L.; Han, L.; Ishaq, H.M.; Nair, S.P.; Xu, J. Photodynamic inactivation of Klebsiella pneumoniae biofilms and planktonic cells by 5-aminolevulinic acid and 5-aminolevulinic acid methyl ester. Lasers Med. Sci. 2016, 31, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Halstead, F.D.; Thwaite, J.E.; Burt, R.; Laws, T.R.; Raguse, M.; Moeller, R.; Webber, M.A.; Oppenheim, B.A. The antibacterial activity of blue light against nosocomial wound pathogens growing planktonically and as mature biofilms. Appl. Environ. Microbiol. 2016, 82, 4006–4016. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wu, X.; Chen, J.; Amin, R.; Lu, M.; Bhayana, B.; Zhao, J.; Murray, C.K.; Hamblin, M.R.; Hooper, D.C.; et al. Antimicrobial Blue Light Inactivation of Gram-Negative Pathogens in Biofilms: In Vitro and In Vivo Studies. J. Infect. Dis. 2016, 213, 1380–1387. [Google Scholar] [CrossRef] [Green Version]
- Orlandi, V.T.; Rybtke, M.; Caruso, E.; Banfi, S.; Tolker-Nielsen, T.; Barbieri, P. Antimicrobial and anti-biofilm effect of a novel BODIPY photosensitizer against Pseudomonas aeruginosa PAO1. Biofouling 2014, 30, 883–891. [Google Scholar] [CrossRef]
- Rothschild, L.J.; Cockell, C.S. Radiation: Microbial evolution, ecology, and relevance to Mars missions. Mutat. Res. Fundam. Mol. Mech. Mutagenesis 1999, 430, 281–291. [Google Scholar] [CrossRef]
- Goosen, N.; Geri, F.M. Repair of UV damage in bacteria. DNA Repair 2008, 7, 353–379. [Google Scholar] [CrossRef] [PubMed]
- Joux, F.; Jeffrey, W.H.; Lebaron, P.; Mitchell, D.L. Marine bacterial isolates display diverse responses to UV-B radiation. Appl. Environ. Microbiol. 1999, 65, 3820–3827. [Google Scholar] [CrossRef] [Green Version]
- Zelle, M.R.; Ogg, J.E. Radiation resistance and genetic segregation in a large cell possibly polyploid strain of Escherichia coli. J. Bacteriol. 1957, 74, 485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karentz, D.; Cleaver, J.E.; Mitchell, D.L. Cell survival characteristics and molecular responses of Antarctic phytoplankton to ultraviolet-B radiation. J. Phycol. 1991, 27, 326–341. [Google Scholar] [CrossRef]
- Davies, R.; Sinskey, A.J. Radiation-resistant mutants of Salmonella typhimurium LT2: Development and characterization. J. Bacteriol. 1973, 113, 133–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grinholc, M.; Rodziewicz, A.; Forys, K.; Rapacka-Zdonczyk, A.; Kawiak, A.; Domachowska, A.; Golunski, G.; Wolz, C.; Mesak, L.; Becker, K.; et al. Fine-tuning recA expression in Staphylococcus aureus for antimicrobial photoinactivation: Importance of photo-induced DNA damage in the photoinactivation mechanism. Appl Microbiol Biotechnol. 2015, 99, 9161–9176. [Google Scholar] [CrossRef] [Green Version]
- Grinholc, M.; Rodziewicz, A.; Forys, K.; Rapacka-Zdonczyk, A.; Kawiak, A.; Domachowska, A.; Golunski, G.; Wolz, C.; Mesak, L.; Becker, K.; et al. Antimicrobial photodynamic therapy with fulleropyrrolidine: Photoinactivation mechanism of Staphylococcus aureus, in vitro and in vivo studies. Appl. Microbiol. Biotechnol. 2015, 99, 4031–4043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakonieczna, J.; Rapacka-Zdonczyk, A.; Kawiak, A.; Bielawski, K.P.; Grinholc, M. Sub-lethal photodynamic inactivation renders Staphylococcus aureus susceptible to silver nanoparticles. Photochem. Photobiol. Sci. 2013, 12, 1622–1627. [Google Scholar] [CrossRef]
- Levin, B.R.; Rozen, D.E. Non-inherited antibiotic resistance. Nat. Rev. Microbiol. 2006, 4, 556–562. [Google Scholar] [CrossRef] [PubMed]
- Sandberg, A.; Jensen, K.S.; Baudoux, P.; Van Bambeke, F.; Tulkens, P.M.; Frimodt-Moller, N. Intra- and extracellular activities of dicloxacillin against Staphylococcus aureus in vivo and in vitro. Antimicrob Agents Chemother. 2010, 54, 2391–2400. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rapacka-Zdonczyk, A.; Wozniak, A.; Nakonieczna, J.; Grinholc, M. Development of Antimicrobial Phototreatment Tolerance: Why the Methodology Matters. Int. J. Mol. Sci. 2021, 22, 2224. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22042224
Rapacka-Zdonczyk A, Wozniak A, Nakonieczna J, Grinholc M. Development of Antimicrobial Phototreatment Tolerance: Why the Methodology Matters. International Journal of Molecular Sciences. 2021; 22(4):2224. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22042224
Chicago/Turabian StyleRapacka-Zdonczyk, Aleksandra, Agata Wozniak, Joanna Nakonieczna, and Mariusz Grinholc. 2021. "Development of Antimicrobial Phototreatment Tolerance: Why the Methodology Matters" International Journal of Molecular Sciences 22, no. 4: 2224. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22042224