
Trogocytosis between Non-Immune Cells for Cell Clearance, and among Immune-Related Cells for Modulating Immune Responses and Autoimmunity

 , , , ,
, , , ,
Abstract
:1. Introduction
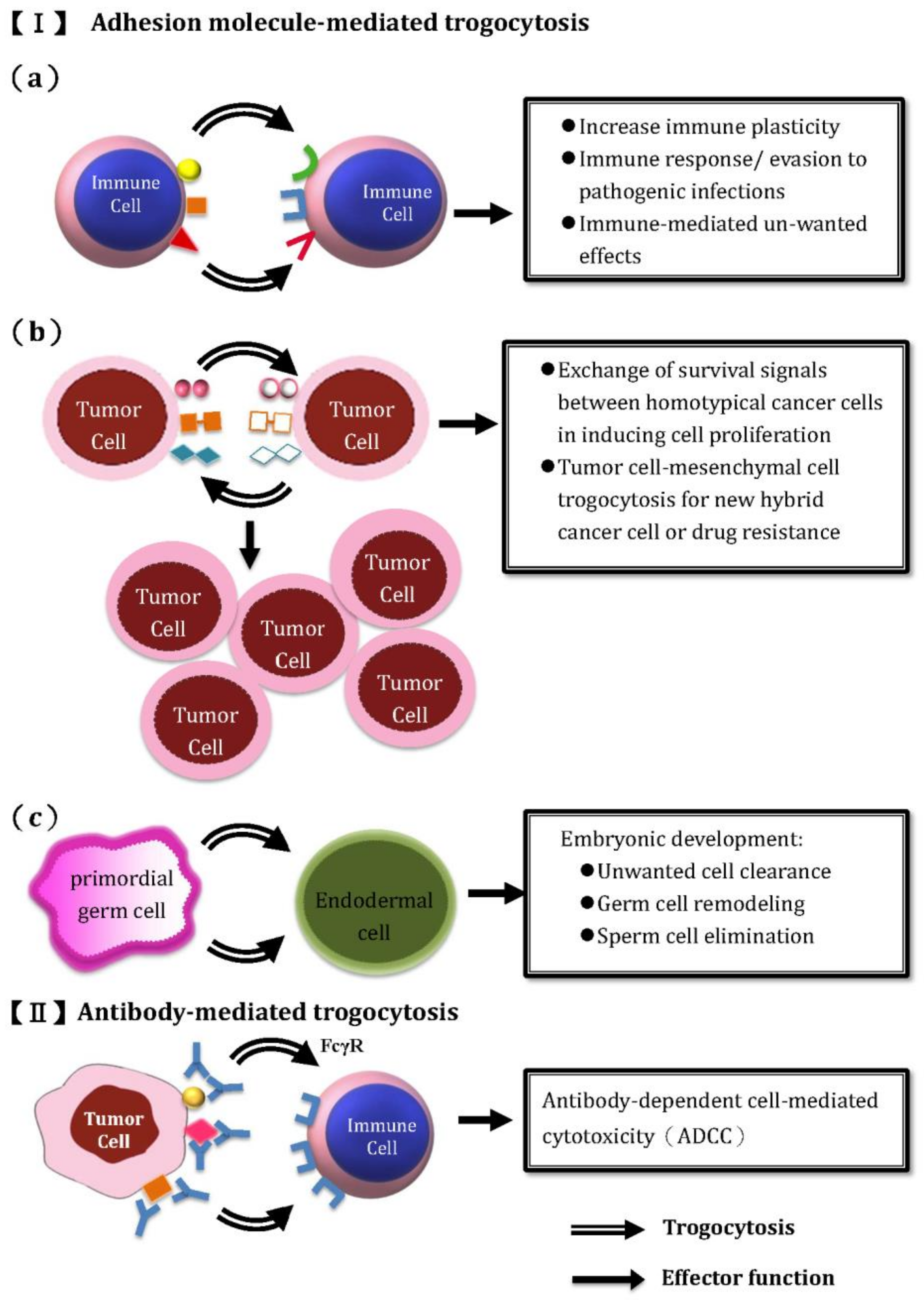
2. Trogocytosis between Embryonic Cells for Un-Wanted Cell Clearance, Germ Cell Remodeling and Sperm Elimination in Embryonic Development
3. Trogocytosis-Associated Cytopathic Effects, Immune Evasion and Immune Response Depending on Different Pathogenic Microbes
3.1. Cytopathic Effects by Eukaryotic Amoebic Parasites via Trogocytosis
3.2. Immune Evasion Induced by Pathogenic Microbes via Trogocytosis
3.2.1. Immune Evasion Induced by Eh via Trogocytosis
3.2.2. Intercellular Transfer of Intracellular Microbial Pathogens via Trogocytosis between Macrophages
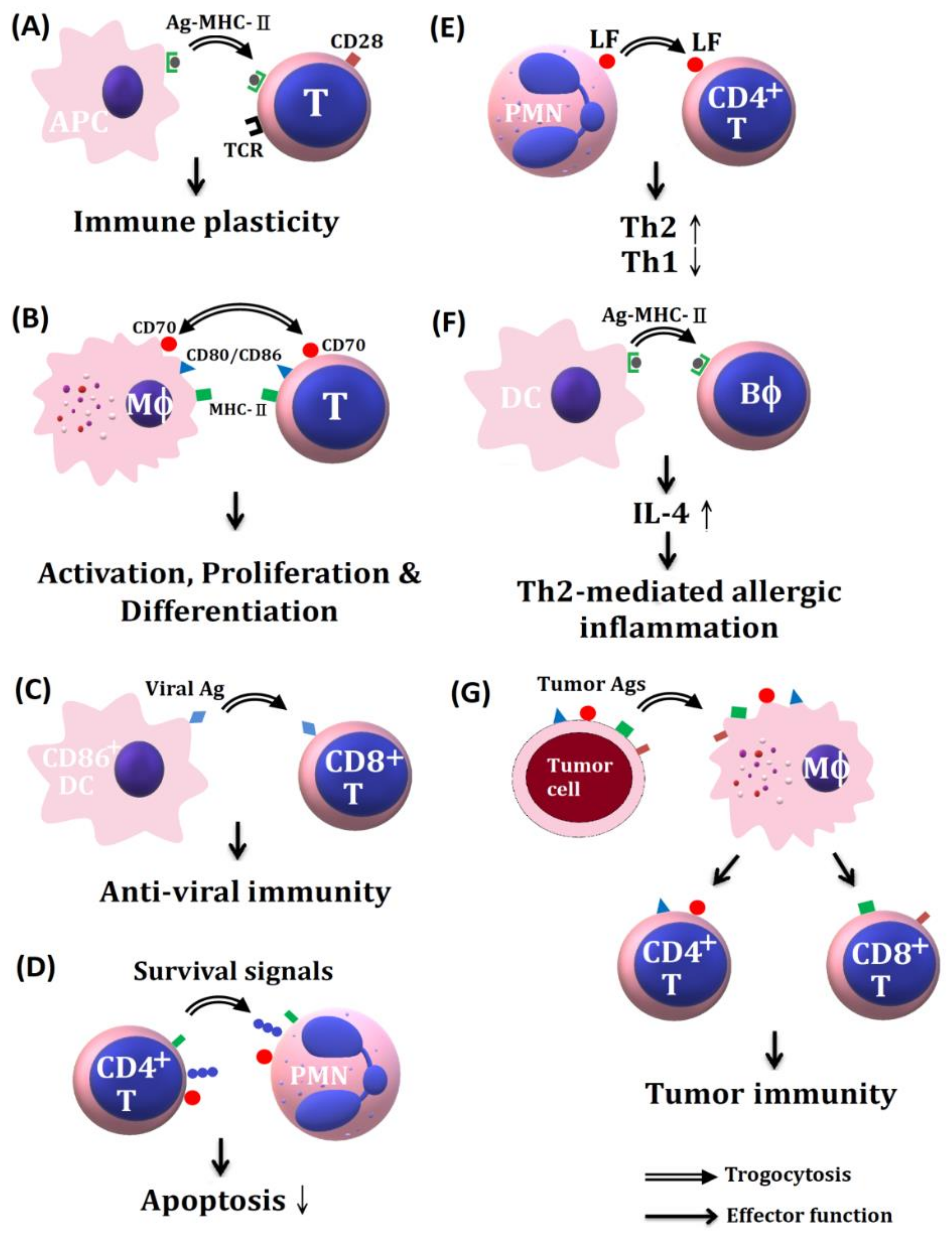
4. The Trogocytosis among Immune Cells for Information Exchange and Generation of Immune Plasticity in the Immune System
4.1. The Molecules on Immune Cells Involved in Trogocytosis
4.2. Intercellular Exchange of Membrane Patches Highlights the Next Level of Immune Plasticity
5. Trogocytosis between Basophils and Dendritic Cells in Supporting Th2-Mediated Allergic and Inflammatory Reactions
6. The Effects of Trogocytosis on the Allogeneic Transplantation in Maintaining Allograft Survive
7. Trogocytosis among Immune-Related Cells and Tumor Cells in Mediating Immune Surveillance and Immune Evasion
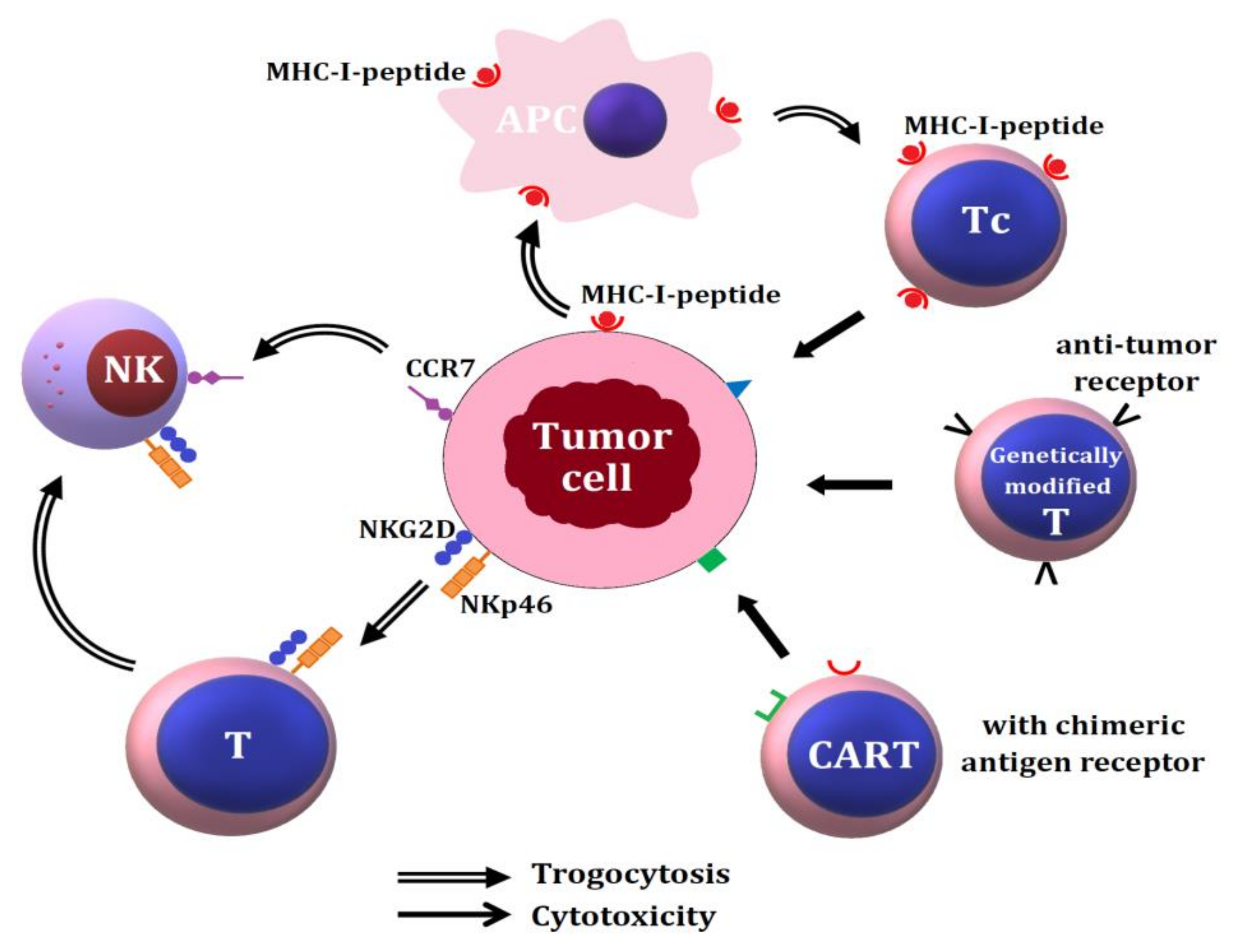
7.1. Capture of Tumor Cell Membranes by CD8+ Cytotoxic T Lymphocytes, NK Cells and Chimeric Antigen Receptor T Cells (CART) for Tumor Surveillance
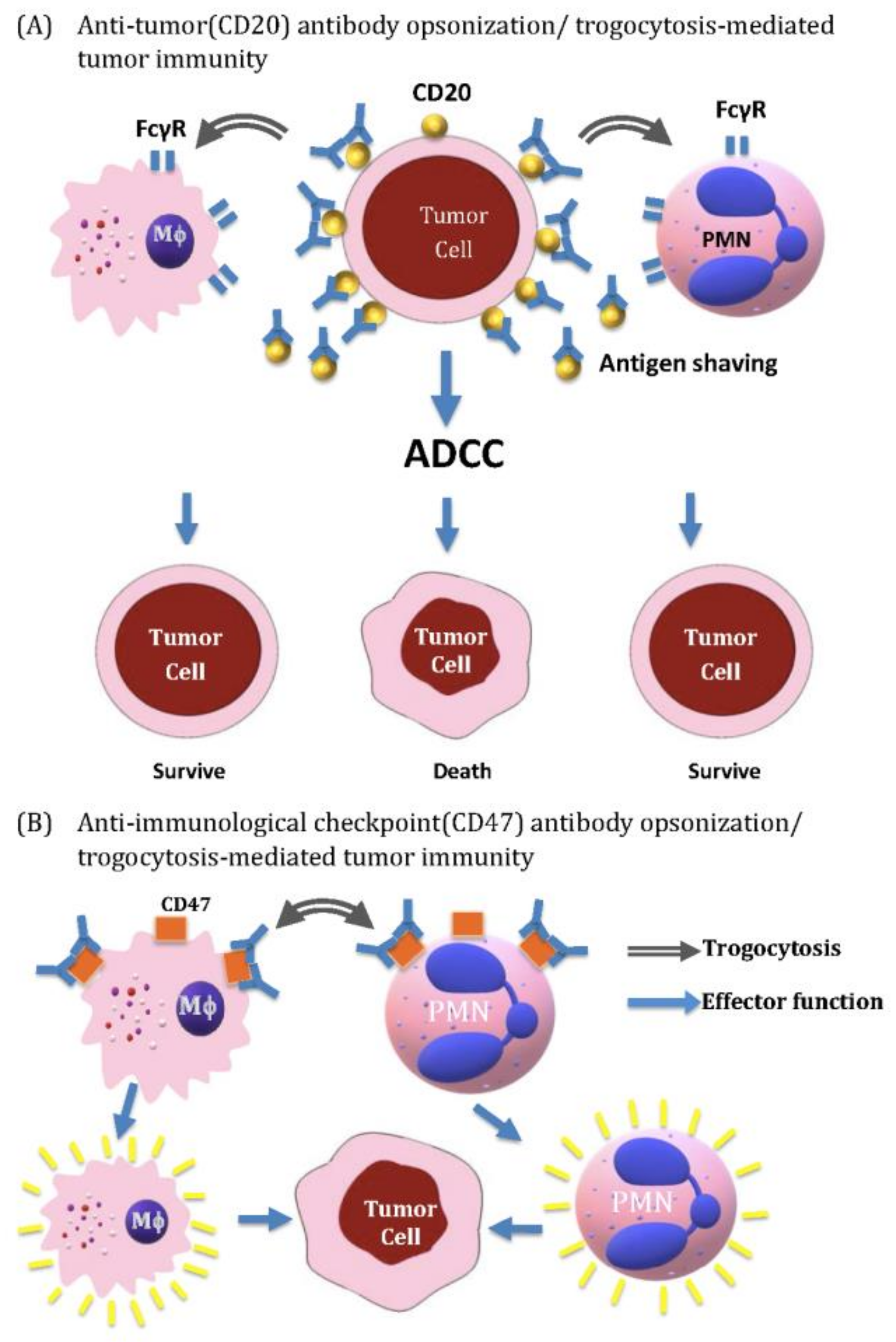
7.2. Antibody-Opsonized Tumor Cell Killing by Innate Immune Cells and Augmented by Trogocytosis and Other Immune-Mediated Mechanisms
7.2.1. Reactivation of Anti-Cancer Immunity of Macrophages by Using Anti-CD47 Antibody and Other Anti-Cancer Antibodies with Augmentation by Trogocytosis
7.2.2. Antibody-Opsonized Cancer Cell Killing by ADCC of Innate Immune Cells and Augmented by Trogocytosis
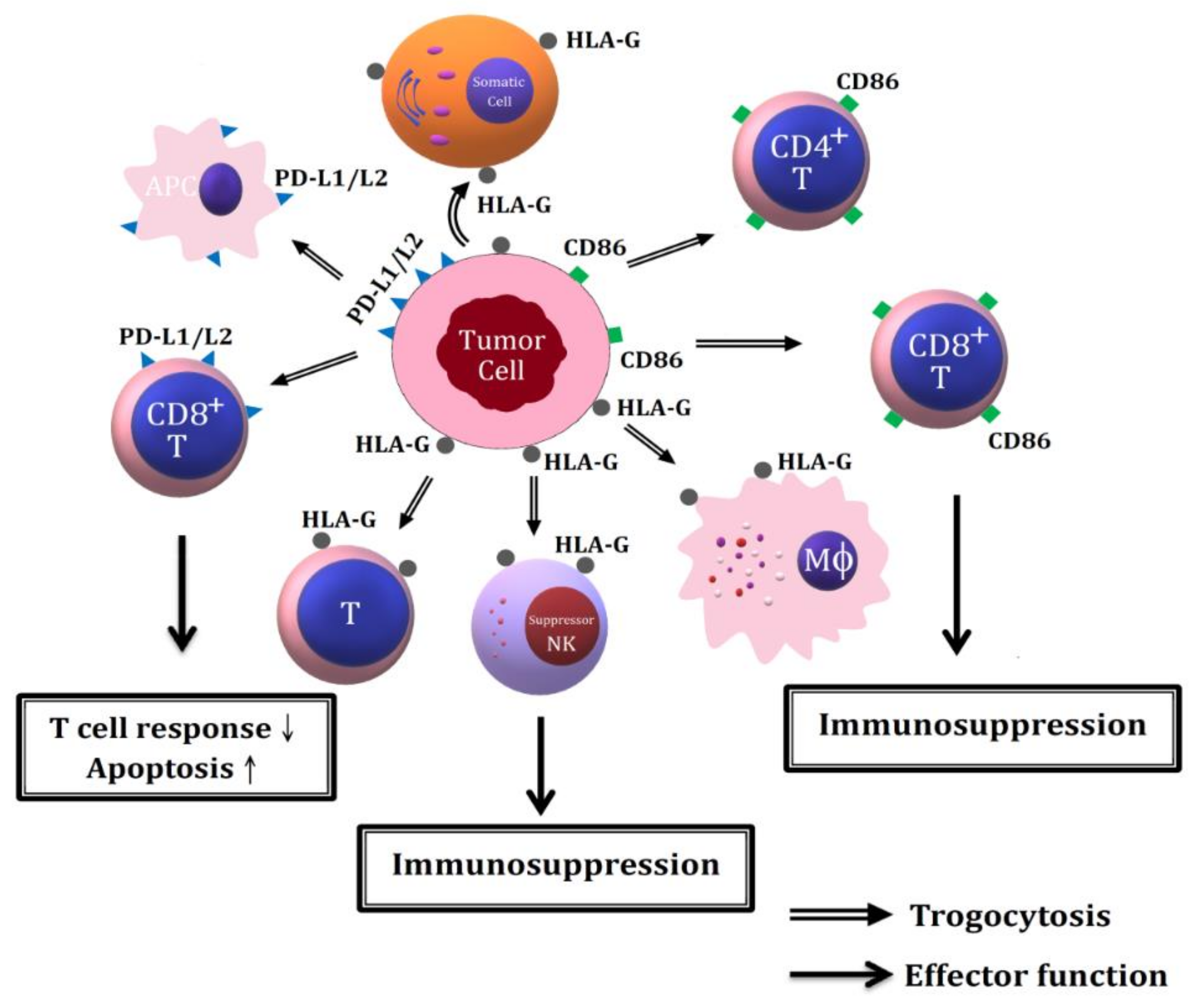
7.2.3. Immune Evasion via Trogocytosis of Immunosuppressive Molecules among Tumor Cells, Immune-Related Cells, and Mesenchymal Stroma/Stem Cells (MSC)
7.2.3.1. Trogocytosis of HLA-G from Tumor Cells to NK Cells, Macrophages and T Cells
7.2.3.2. Trogocytosis of PD-L1/L2 Molecules among Tumor Cells, APC and CD8+ T Cells in Facilitating Immune Evasion to Tumor Cells
7.2.3.3. Oncologic Trogocytosis between Tumor Cells and Microenvironmental Mesenchymal Stroma/Stem Cells in Increasing Tumor Heterogeneity
8. Abnormal Trogocytosis in Systemic Autoimmune and Immune-Mediated Inflammatory Diseases
8.1. Decreased Expr#Ession of HLA-G Mole#Cule on # in the Patients with Systemic Lupus Erythematosus (SLE)
8.2. Aberrant Trogocytosis in Other Autoimmune and Immune-Mediated Inflammatory Diseases
9. Conclusions and Perspectives
- (1)
- The real molecular mechanism of cell-cell membrane transfer and the repulsive force after cell-cell contact.
- (2)
- The real molecular mechanism of homotypic tumor-tumor and tumor-MSC trogocytosis in hybrid cancer cell formation and for novel cancer therapy in future.
- (3)
- The appropriate use of cytoskeletal protein blockade to suppress trogocytosis of immunomodulatory molecules (HLA-G, PD-L1/L2 and CD37) between tumor and immune cells.
- (4)
- The selective modulation of stimulatory or inhibitory IgG-Fc receptor expression in adjusting cell-cell membrane transfer remains elucidation.
- (5)
- The best strategy of antibody-opsonized tumor cell killing by immune-related cells remain evaluation.
- (6)
- CART immunotherapy may also face several challenges including therapeutic potency, impaired trafficking to solid tumor, local immunosuppression within tumor microenvironment, and toxicity associated with CART cells. Although Petty et al. [165] proposed to use CAR-NK or CAR-NKT cell instead of CAR-T cells. Combination therapy of CAR cells and anti-tumor antibody may potentially improve the efficacy for CAR cells trafficking into solid tumors.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ADCC | antibody-dependent cell-mediated cytotoxicity |
| AIHA | antibody-induced hemolytic anemia |
| APC | Adenosine diphosphate ribose |
| Bϕ | basophil |
| CART | chimeric antigen receptor T lymphocyte |
| CCR | C-C chemokine receptor |
| CD4+ T | helper T lymphocyte |
| CD8+T | cytotoxic T lymphocyte |
| CTL | cytotoxic T lymphocyte |
| DC | dendritic cell |
| Eh | Entamoeba histolytica |
| Gal/GalNAc | galactose/N-acetyl-galactose |
| HLA-G | human leukocyte antigen-G |
| iTreg | inducible regulatory T cell |
| IL | interleukin |
| ILT | immunoglobulin-like transcript |
| IUHCT | in utero hematopoietic cellular transplantation |
| KIR | killer immunoglobulin-like receptor |
| MCP | macrophage chemotactic protein |
| Mϕ | macrophage |
| MHC-I | major histocompatibility complex class I antigen |
| MHC-II | major histocompatibility complex class II antigen |
| MSC | mesenchymal stroma/stem cell |
| NK | natural killer cell |
| PGC | primordial germ cell |
| PD-1 | programmed cell death-1 |
| PD-L1/L2 | programmed cell death ligand 1/2 |
| SCID | severe combined immunodeficiency |
| SLE | systemic lupus erythematosus |
| Tc | cytotoxic T lymphocyte |
| Th1 | helper T cell type1 |
| Th2 | helper T cell type2 |
References
- Joly, E.; Hudrisier, D. What is trogocytosis and what is its purpose? Nat. Immunol. 2003, 4, 815. [Google Scholar] [CrossRef]
- Abdu, Y.; Maniscalco, C.; Heddleston, J.M.; Chew, T.L. Developmentally programmed germ cell remodelling by endodermal cell cannibalism. Nat. Cell Biol. 2016, 18, 1302–1310. [Google Scholar] [CrossRef]
- Kinnear, F.B. Cytopathogenicity of acanthamoeba, vahlkampfia and hartmannella: Quantitative & qualitative in vitro studies on keratocytes. J. Infect. 2003, 46, 228–237. [Google Scholar] [PubMed]
- Miller, H.W.; Suleiman, R.L.; Ralston, K.S. Trogocytosis by Entamoeba histolytica mediates acquisition and display of human cell membrane proteins and evasion of lysis by human serum. Am. Soc. Microbiol. 2019, 10, e00068-19. [Google Scholar] [CrossRef] [Green Version]
- Hudrisier, D.; Riond, J.; Mazarguil, H.; Gairin, J.E.; Joly, E. Cutting edge: CTLs rapidly capture membrane fragments from target cells in a TCR signaling-dependent manner. J. Immunol. 2001, 166, 3645–3649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dance, A.; Writer, S. Cells nibble one another via the under-appreciated process of trogocytosis. Proc. Natl. Acad. Sci. USA 2019, 116, 17608–17610. [Google Scholar] [CrossRef] [Green Version]
- Arandjelovic, S.; Ravichandran, K.S. Phagocytosis of apoptotic cells in homeostasis. Nat. Immunol. 2015, 16, 907–917. [Google Scholar] [CrossRef] [Green Version]
- Elliott, M.R.; Ravichandran, K.S. The dynamics of apoptotic cell Clearance. Dev. Cell 2016, 38, 147–160. [Google Scholar] [CrossRef] [Green Version]
- Gordon, S. Phagocytosis: An Immunobiologic Process. Immunity 2016, 44, 463–475. [Google Scholar] [CrossRef] [Green Version]
- Perry, J.S.; Ravichandran, K.S. Embryonic Trogocytosis: Neighborly nibbling during development. Curr. Biol. 2017, 27, R68–R70. [Google Scholar] [CrossRef] [PubMed]
- Weihard, L.; di Bartolomei, G.; Bolasco, G.; Machado, P.; Schieber, N.L.; Neniskyte, Y.; Exiga, M.; Nadisiute, A.; Raggioli, A.; Schertel, A.; et al. Microglia remodel synapses by presynaptic trogocytosis and spine head filopodia induction. Nat. Commun. 2018, 9, 1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villani, A.; Peri, F. Microglia: Picky brain eaters. Dev. Cell 2018, 48, 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riccomagno, M.M.; Kolodkin, A.L. Sculpting neural circuits by axon and dendrite pruning. Annu. Rev. Cell. Dev. Biol. 2015, 31, 779–805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, J.W.; Winklbauer, R. Ingression-type cell migration drives vegetal endoderm internalization in the Xenopus gastrula. Elife 2017, 6, e27190. [Google Scholar] [CrossRef] [PubMed]
- Batlle, E.; Wilkinson, D.G. Molecular mechanisms of cell segregation and boundary formation in development and tumorigenesis. Cold Spring Harb. Perspect. Biol. 2012, 4, a008227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ventrella, R.; Kaplan, N.; Getsios, S. Asymmetry at cell-cell interfaces direct cell sorting, boundary formation, and tissue morphogenesis. Exp. Cell Res. 2017, 358, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Ralston, K.S. Taking a bite: Amoebic trogocytosis in Entamoeba histolytica and beyond. Curr. Opin. Microbiol. 2015, 28, 26–35. [Google Scholar] [CrossRef] [Green Version]
- Gong, J.; Gaitanos, T.N.; Luu, O.; Huang, Y.; Gaitanos, L.; Lindner, J.; Winklbauer, R.; Klein, R. Gulp1 controls Eph/ephrin trogocytosis and is important for cell rearrangements during development. J. Cell Biol. 2019, 218, 3455–3471. [Google Scholar] [CrossRef] [Green Version]
- Valgardsdottir, R.; Cattaneo, I.; Klein, C.; Introna, M.; Figliuzzi, M.; Golay, J. Human neutrophils mediate trogocytosis rather than phagocytosis of CLL B cells opsonized with anti-CD20 antibodies. Blood 2017, 129, 2636–2644. [Google Scholar] [CrossRef] [Green Version]
- Mercer, F.; Ng, S.H.; Brown, T.M.; Boatman, G.; Johnson, P.J. Neutrophils kill the parasite Trichomonas vaginalis using trogocytosis. PLoS Biol. 2018, 16, e2003885. [Google Scholar] [CrossRef]
- Olivera-Valle, I.; Latorre, M.C.; Calvo, M.; Gaspar, B.; Gómez-Oro, C.; Collazos, A.; Breton, A.; Caballero-Campo, P.; Ardoy, M.; Asensio, F.; et al. Vaginal neutrophils eliminate sperm by trogocytosis. Hum. Reprod. 2020, 35, 2567–2578. [Google Scholar] [CrossRef] [PubMed]
- Brown, T. Observations by immunofluorescence microscopy and electron microscopy on the cytopathogenicity of Naegleria fowleri in mouse embryo-cell cultures. J. Med. Microbiol. 1979, 12, 363–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ralston, K.S.; Solga, M.D.; Mackey-Lawrence, N.M.; Bhattacharya, A.; Petri, W.A., Jr. Trogocytosis by Entamoeba histolytica contributes to cell killing and tissue invasion. Nature 2014, 508, 526–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ralston, K.S. Chew on this: Amoebic trogocytosis and host cell killing by Entamoeba histolytica. Trends Parasitol. 2015, 31, 442–452. [Google Scholar] [CrossRef] [Green Version]
- Nakada-Tsukui, K.; Nozaki, T. AGC Family kinase 1 participates in trogocytosis but not in phagocytosis in Entamoeba histolytica. Nat. Commun. 2017, 8, 101. [Google Scholar]
- Bettadapur, A.; Ralston, K.S. Direct and high-throughput assays for human cell killing through trogocytosis by Entamoeba histolytica. Mol. Biochem. Parasitol. 2020, 239, 111301. [Google Scholar] [CrossRef] [PubMed]
- Reed, S.L.; Keene, W.E.; McKerrow, J.H.; Gigli, I. Cleavage of C3 by a neutral cysteine proteinase of Entamoeba histolytica. J. Immunol. 1989, 143, 189–195. [Google Scholar] [PubMed]
- Reed, S.L.; Gigli, I. Lysis of complement-sensitive Entamoeba histolytica by activated terminal complement components. Initiation of complement activation by an extracellular neutral cysteine proteinase. J. Clin. Investig. 1990, 86, 1815–1822. [Google Scholar] [CrossRef]
- Reed, S.L.; Ember, J.A.; Herdman, D.S.; DiScipio, R.G.; Hugli, T.E.; Gigli, I. The extracellular neutral cysteine proteinase of Entamoeba histolytica degrades anaphylatoxins C3a and C5a. J. Immunol. 1995, 155, 266–274. [Google Scholar] [PubMed]
- Braga, L.L.; Ninomiya, H.; McCoy, J.J.; Eacker, S.; Wiedmer, T.; Pham, C.; Wood, S.; Sims, P.J.; Petri, W.A., Jr. Inhibition of the complement membrane attack complex by the galactose-specific adhesion of Entamoeba histolytica. J. Clin. Investig. 1992, 90, 1131–1137. [Google Scholar] [CrossRef] [Green Version]
- Begum, S.; Quach, J.; Chadee, K. Immune evasion mechanisms of Entamoeba histolytica: Progression to disease. Front. Microbiol. 2015, 6, 1394. [Google Scholar] [CrossRef]
- Steele, S.; Radlinski, L.; Taft-Benz, S.; Brunton, J.; Kawula, T.H. Trogocytosis-associated cell to cell spread of intracellular bacterial pathogens. Elife 2016, 5, e10625. [Google Scholar] [CrossRef] [PubMed]
- Steele, S.P.; Chamberlain, Z.; Park, J.; Kawula, T.H. Francisella tularensis enters a double membrane compartment following cell-cell transfer. Elife 2019, 8, e45252. [Google Scholar] [CrossRef] [PubMed]
- Kongsomros, S.; Thanunchai, M.; Manopwisedjaroen, S.; Na-Ek, P.; Wang, S.F.; Taechalertpaisarn, T.; Thitithanyanont, A. Trogocytosis with monocytes associated with increased alpha2,3 sialic acid expression on B cells during H5N1 influenza virus infection. PLoS ONE 2020, 15, e0239488. [Google Scholar] [CrossRef]
- Batista, F.D.; Iber, D.; Neuberger, M.S. B cells acquire antigen from target cells after synapse formation. Nature 2001, 411, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Hudrisier, D.; Aucher, A.; Puaux, A.L.; Bordier, C.; Joly, E. Capture of target cell membrane components via trogocytosis is triggered by a selected set of surface molecules on T or B cells. J. Immunol. 2007, 178, 3637–3647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakayama, M.; Takeda, K.; Kawano, M.; Takai, T.; Ishii, N.; Ogasawara, K. Natural killer (NK)-dendritic cell interactions generate MHC class II-dressed NK cells that regulate CD4+T cells. Proc. Natl. Acad. Sci. USA 2011, 108, 18360–18365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonaccorsi, I.; Morandi, B.; Antsiferova, O.; Costa, G.; Oliver, D.; Conte, R.; Pezzino, G.; Vermiglio, G.; Anastasi, G.P.; Navarra, G.; et al. Membrane transfer from tumor cells overcomes deficient phagocytic ability of plasmacytoid dendritic cells for the acquisition and presentation of tumor antigens. J. Immunol. 2014, 192, 824–832. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.; Mero, P.; Booth, J.W. Dynamics of macrophage trogocytosis of rituximab-coated B cells. PLoS ONE 2011, 6, e14498. [Google Scholar] [CrossRef] [Green Version]
- Li, K.J.; Wu, C.H.; Shen, C.Y.; Kuo, Y.M.; Yu, C.L.; Hsieh, S.C. Membrane transfer from mononuclear cells to polymorphonuclear neutrophils transduces cell survival and activation signals in the recipient cells via anti-extrinsic apoptotic and MAP kinase signaling pathways. PLoS ONE 2016, 11, e0156262. [Google Scholar] [CrossRef] [Green Version]
- Miyake, K.; Shiozawa, N.; Nagao, T.; Yoshikawa, S.; Yamanishi, Y.; Karasuyama, H. Trogocytosis of peptide-MHC class II complexes from dendritic cells confers antigen-presenting ability on basophils. Proc. Natl. Acad. Sci. USA 2017, 114, 1111–1116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.F.; Yang, Y.; Sepulveda, H.; Shi, W.; Hwang, I.; Peterson, P.A.; Jackson, M.R.; Sprent, J.; Cai, Z. TCR-mediated internalization of peptide-MHC complexes acquired by T cells. Science 1999, 286, 952–954. [Google Scholar] [CrossRef]
- Rosenits, K.; Keppler, S.J.; Vucikuja, S.; Aichele, P. T cells acquire cell surface determinants of APC via in vivo trogocytosis during viral infections. Eur. J. Immunol. 2010, 40, 3450–3457. [Google Scholar] [CrossRef]
- Son, J.; Ha, S.J. Extrinsic acquisition of CD80 by antigen-specific CD8+T Cells regulates their recall immune responses to acute viral infection. Immune Netw. 2019, 19, e25. [Google Scholar] [CrossRef]
- Tabiasco, J.; Espinosa, E.; Hudrisier, D.; Joly, E.; Fournié, J.J.; Vercellone, A. Active trans-synaptic capture of membrane fragments by natural killer cells. Eur. J. Immunol. 2002, 32, 1502–1508. [Google Scholar] [CrossRef]
- Daubeuf, S.; Lindorfer, M.A.; Taylor, R.P.; Joly, E.; Hudrisier, D. The direction of plasma membrane exchange between lymphocytes and accessory cells by trogocytosis is influenced by the nature of accessory cell. J. Immunol. 2010, 184, 1897–1908. [Google Scholar] [CrossRef]
- Daubeuf, S.; Puaux, A.-L.; Joly, E.; Hudrisier, D. A simple trogocytosis-based method to detect quantify, characterize and purify antigen-specific live lymphocytes by flow cytometry, via their capture of membrane fragments from antigen-presenting cells. Nat. Protoc. 2006, 1, 2536–2542. [Google Scholar] [CrossRef] [PubMed]
- Daubeuf, S.; Aucher, A.; Sampathkumar, S.-G.; Preville, X.; Yarema, K.J.; Hudrisier, D. Chemical labels metabolically installed into the glycoconjugates of the target cell surface can be used to track lymphocyte/target cell interplay via trogocytosis: Comparisons with lipophilic dyes and biotin. Immunol. Invest. 2007, 36, 687–712. [Google Scholar] [CrossRef] [PubMed]
- Wetzel, S.; McKeiththan, T.; Parker, D. Peptide-specific intercellular transfer of MHC class II to CD4 T cells directly from the immunological synapse upon cellular dissociation. J. Immunol. 2005, 174, 80–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chattpadhyay, P.K.; Yu, J.; Roederer, M.A. A live-cell assay to detect antigen-specific CD4+T cells with diverse cytokine profiles. Nat. Med. 2005, 11, 1113–1117. [Google Scholar] [CrossRef]
- Daubeuf, S.; Bordier, C.; Hudrisier, D.; Joly, E. Suitability of various membrane lipophilic probes for the detection of trogocytosis by flow cytometry. Cytometry Part A 2009, 75, 380–389. [Google Scholar] [CrossRef]
- Lorber, M.I.; Loken, M.R.; Stall, A.M.; Fitch, F.W. I-A antigens on cloned alloreactive murine T lymphocytes are acquired passively. J. Immunol. 1982, 128, 2798–2803. [Google Scholar] [PubMed]
- Hwang, I.; Huang, J.F.; Kishimoto, H.; Brunmark, A.; Peterson, P.A.; Jackson, M.R.; Surh, C.D.; Cai, Z.; Sprent, J. T cells can use either T cell receptor or CD28 receptors to absorb and internalize cell surface molecules derived from antigen-presenting cells. J. Exp. Med. 2000, 191, 1137–1148. [Google Scholar] [CrossRef]
- Ramming, A.; Thümmler, K.; Schulze-Koops, H.; Skapenko, A. Homotypic T-cell/T-cell interaction induces T-cell activation, proliferation, and differentiation. Hum. Immunol. 2009, 70, 873–881. [Google Scholar] [CrossRef]
- Alegre, E.; Howangyin, K.Y.; Favier, B.; Baudhuin, J.; Lesport, E.; Daouya, M.; Gonzalez, A.; Carosella, E.D.; Lemaoult, J. Membrane redistributions through multi-intercellular exchanges and serial trogocytosis. Cell Res. 2010, 20, 1239–1251. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, S.; Masuda, S.; Baba, T.; Tomaru, U.; Katsumata, K.; Kasahara, M.; Ishizu, A. Plasma-dependent, antibody- and Fcγ receptor-mediated translocation of CD8 molecules from T cells to monocytes. Cytometry Part A 2011, 79A, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Aucher, A.; Magdeleine, E.; Joly, E.; Hudrisier, D. Capture of plasma membrane fragments from target cells by trogocytosis requires signaling in T cells but not in B cells. Blood 2008, 111, 5621–5628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, K.J.; Lu, M.C.; Hsieh, S.C.; Wu, C.H.; Yu, H.S.; Tsai, C.Y.; Yu, C.L. Release of surface-expressed lactoferrin from polymorphonuclear neutrophils after contact with CD4+T cells and its modulation on Th1/Th2 cytokine production. J. Leukoc. Biol. 2006, 80, 350–358. [Google Scholar] [CrossRef]
- Reed, J.; Wetzel, S.A. Trogocytosis-mediated intracellular signaling in CD4+T cells drives TH2-associated effector cytokine production and differentiation. J. Immunol. 2019, 202, 2873–2887. [Google Scholar] [CrossRef] [PubMed]
- LeMaoult, J.; Caumartin, J.; Carosella, E.D. Exchanges of membrane patches (trogocytosis) split theoretical and actual functions of immune cells. Hum. Immunol. 2007, 68, 240–243. [Google Scholar] [CrossRef] [PubMed]
- Riond, J.; Elhmouzi, J.; Hudrisier, D.; Gairin, J.E. Capture of membrane components via trogocytosis occurs in vivo during both dendritic cells and target cells encounter by CD8(+) T cells. Scand. J. Immunol. 2007, 66, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, M.; Watanabe, M.; Uchimaru, K.; Horie, R. Trogocytosis of ligand-receptor complex and its intracellular transport in CD30 signaling. Biol. Cell 2018, 110, 109–124. [Google Scholar] [CrossRef] [PubMed]
- Sokol, C.L.; Barton, G.M.; Farr, A.G.; Medzhitov, R.A. A mechanism for the initiation of allergen-induced T helper type 2 responses. Nat. Immunol. 2008, 9, 310–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshimoto, T.; Yasuda, K.; Tanaka, H.; Nakahira, M.; Imai, Y.; Fujimori, Y.; Nakanishi, K. Basophils contribute to T(H)2-IgE responses in vivo via IL-4 production and presentation of peptide-MHC class II complexes to CD4+T cells. Nat. Immunol. 2009, 10, 706–712. [Google Scholar] [CrossRef] [PubMed]
- Yoshimoto, T.; Tsutsui, H.; Tominaga, K.; Hoshino, K.; Okamura, H.; Akira, S.; Paul, W.E.; Nakanishi, K. IL-18, although antiallergic when administered with IL-12, stimulates IL-4 and histamine release by basophils. Proc. Natl. Acad. Sci. USA 1999, 96, 13962–13966. [Google Scholar] [CrossRef] [Green Version]
- Kondo, Y.; Yoshimoto, T.; Yasuda, K.; Futatsugi-Yumikura, S.; Morimoto, M.; Hayashi, N.; Hoshino, T.; Fujimoto, J.; Nakanishi, K. Administration of IL-33 induces airway hyperresponsiveness and goblet cell hyperplasia in the lungs in the absence of adaptive immune system. Int. Immunol. 2008, 20, 791–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamanishi, Y.; Miyake, K.; Iki, M.; Tsutsui, H.; Karasuyama, H. Recent advances in understanding basophil-mediated Th2 immune responses. Immunol. Rev. 2017, 278, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Karasuyama, H.; Miyake, K.; Yoshikawa, S.; Kawano, Y.; Yamanishi, Y. How do basophils contribute to Th2 cell differentiation and allergic responses? Int. Immunol. 2018, 30, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Ohlén, C.; Kling, G.; Höglund, P.; Hansson, M.; Scangos, G.; Bieberich, C.; Jay, G.; Kärre, K. Prevention of allogeneic bone marrow graft rejection by H-2 transgene in donor mice. Science 1989, 246, 666–668. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, N.; Wong, C.J.; Gertsenstein, M.; Casper, R.F.; Nagy, A.; Rogers, I.M. Bone marrow transplantation results in human donor blood cells acquiring and displaying mouse recipient class I MHC and CD45 antigens on their surface. PLoS ONE 2009, 4, e8489. [Google Scholar] [CrossRef] [Green Version]
- Chow, T.; Whiteley, J.; Li, M.; Rogers, I.M. The transfer of host MHC class I protein protects donor cells from NK cell and macrophage-mediated rejection during hematopoietic stem cell transplantation and engraftment in mice. Stem Cells 2013, 31, 2242–2252. [Google Scholar] [CrossRef] [PubMed]
- Rogers, I.M. Trogocytosis in allogeneic transplants: Donor cells take on the recipients identity. Chimerism 2013, 4, 142–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durkin, E.T.; Jones, K.A.; Rajesh, D.; Shaaban, A.F. Early chimerism threshold predicts sustained engraftment and NK-cell tolerance in prenatal allogeneic chimeras. Blood 2008, 112, 5245–5253. [Google Scholar] [CrossRef] [Green Version]
- Alhajjat, A.M.; Strong, B.S.; Durkin, E.T.; Turner, L.E.; Wadhwani, R.K.; Midura, E.F.; Keswani, S.G.; Shaaban, A.F. Trogocytosis as a mechanistic link between chimerism and prenatal tolerance. Chimerism 2013, 4, 126–131. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, K.A.; Munegowda, M.A.; Xie, Y.; Xiang, J. Intercellular trogocytosis plays an important role in modulation of immune responses. Cell. Mol. Immunol. 2008, 5, 261–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rechavi, O.; Goldstein, I.; Kloog, Y. Intercellular exchange of proteins: The immune cell habit of sharing. FEBS Lett. 2009, 583, 1792–1799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhainaut, M.; Moser, M. Regulation of immune reactivity by intercellular transfer. Front. Immunol. 2014, 5, 112. [Google Scholar] [CrossRef] [Green Version]
- Barinov, A.; Galgano, A.; Krenn, G.; Tanchot, C.; Vasseur, F.; Rocha, B. CD4/CD8/Dendritic cell complexes in the spleen: CD8+T cells can directly bind CD4+T cells and modulate their response. PLoS ONE 2017, 12, e0180644. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Schwarz, H. The role of trogocytosis in immune surveillance of Hodgkin lymphoma. Oncoimmunology 2020, 9, 1781334. [Google Scholar] [CrossRef] [PubMed]
- Stinchcombe, J.C.; Bossi, G.; Booth, S.; Griffiths, G.M. The immunological synapse of CTL contains a secretory domain and membrane bridges. Immunity 2001, 15, 751–761. [Google Scholar] [CrossRef] [Green Version]
- Machlenkin, A.; Uzana, R.; Frankenburg, S.; Eisenberg, G.; Eisenbach, L.; Pitcovski, J.; Gorodetsky, R.; Nissan, A.; Peretz, T.; Lotem, M. Capture of tumor cell membranes by trogocytosis facilitates detection and isolation of tumor-specific functional CTLs. Cancer Res. 2008, 68, 2006–2013. [Google Scholar] [CrossRef] [Green Version]
- Uzana, R.; Eisenberg, G.; Sagi, Y.; Frankenburg, S.; Merims, S.; Amariglio, N.; Yefenof, E.; Peretz, T.; Machlenkin, A.; Lotem, M. Trogocytosis is a gateway to characterize functional diversity in melanoma-specific CD8+T cell clones. J. Immunol. 2012, 188, 632–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uzana, R.; Eisenberg, G.; Merims, S.; Frankenburg, S.; Pato, A.; Yefenof, E.; Engelstein, R.; Peretz, T.; Machlenkin, A.; Lotem, M. Human T cell crosstalk is induced by tumor membrane transfer. PLoS ONE 2015, 10, e0118244. [Google Scholar] [CrossRef]
- Ljunggren, H.G.; Malmberg, K.J. Prospects for the use of NK cells in immunotherapy of human cancer. Nat. Rev. Immunol. 2007, 7, 329–339. [Google Scholar] [CrossRef]
- Williams, G.S.; Collinson, L.M.; Brzostek, J.; Eissmann, P.; Almeida, C.R.; McCann, F.E.; Burshtyn, D.; Davis, D.M. Membranous structures transfer cell surface proteins across NK cell immune synapses. Traffic 2007, 8, 1190–1204. [Google Scholar] [CrossRef]
- Marcenaro, E.; Cantoni, C.; Pesce, S.; Prato, C.; Pende, D.; Agaugué, S.; Moretta, L.; Moretta, A. Uptake of CCR7 and acquisition of migratory properties by human KIR+NK cells interacting with monocyte-derived DC or EBV cell lines: Regulation by KIR/HLA-class I interaction. Blood 2009, 114, 4108–4116. [Google Scholar] [CrossRef]
- Domaica, C.I.; Fuertes, M.B.; Rossi, L.E.; Girart, M.V.; Avila, D.E.; Rabinovich, G.A.; Zwirner, N.W. Tumour-experienced T cells promote NK cell activity through trogocytosis of NKG2D and NKp46 ligands. EMBO Rep. 2009, 10, 908–915. [Google Scholar] [CrossRef] [Green Version]
- Cho, F.N.; Chang, T.H.; Shu, C.W.; Ko, M.C.; Liao, S.K.; Wu, K.H.; Yu, M.S.; Lin, S.J.; Hong, Y.C.; Chen, C.H.; et al. Enhanced cytotoxicity of natural killer cells following the acquisition of chimeric antigen receptors through trogocytosis. PLoS ONE 2014, 9, e109352. [Google Scholar] [CrossRef] [PubMed]
- Sadelain, M.; Rivière, I.; Riddell, S. Therapeutic T cell engineering. Nature 2017, 545, 423–431. [Google Scholar] [CrossRef] [Green Version]
- Brudno, J.N.; Maric, I.; Hartman, S.; Rose, J.J.; Wang, M.; Lam, N.; Stetler-Stevenson, M.; Salem, D.; Yuan, C.; Pavletic, S.; et al. T Cells genetically modified to express an anti-B-cell maturation antigen chimeric antigen receptor cause remissions of poor-prognosis relapsed multiple myeloma. J. Clin. Oncol. 2018, 36, 2267–2280. [Google Scholar] [CrossRef] [PubMed]
- Orlando, E.J.; Han, X.; Tribouley, C.; Wood, P.A.; Leary, R.J.; Riester, M.; Levine, J.E.; Qayed, M.; Grupp, S.A.; Boyer, M.; et al. Genetic mechanisms of target antigen loss in CAR19 therapy of acute lymphoblastic leukemia. Nat. Med. 2018, 24, 1504–1506. [Google Scholar] [CrossRef] [PubMed]
- Hamieh, M.; Dobrin, A.; Cabriolu, A.; van der Stegen, S.J.C.; Giavridis, T.; Mansilla-Soto, J.; Eyquem, J.; Zhao, Z.; Whitlock, B.M.; Miele, M.M.; et al. CAR T cell trogocytosis and cooperative killing regulate tumour antigen escape. Nature 2019, 568, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Sakuishi, K.; Apetoh, L.; Sullivan, J.M.; Blazar, B.R.; Kuchroo, V.K.; Anderson, A.C. Targeting Tim-3 and PD-1 pathways to reverse T cell exhaustion and restore anti-tumor immunity. J. Exp. Med. 2010, 207, 2187–2194. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.J.; Frazier, W.A. Integrin-associated protein (CD47) and its ligands. Trends Cell Biol. 2001, 11, 130–135. [Google Scholar] [CrossRef]
- Blazar, B.R.; Lindberg, F.P.; Ingulli, E.; Panoskaltsis-Mortari, A.; Oldenborg, P.A.; Iizuka, K.; Yokoyama, W.M.; Taylor, P.A. CD47 (integrin-associated protein) engagement of dendritic cell and macrophage counterreceptors is required to prevent the clearance of donor lymphohematopoietic cells. J. Exp. Med. 2001, 194, 541–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majeti, R.; Chao, M.P.; Alizadeh, A.A.; Pang, W.W.; Jaiswal, S.; Gibbs, K.D., Jr.; van Rooijen, N.; Weissman, I.L. CD47 is an adverse prognostic factor and therapeutic antibody target on human acute myeloid leukemia stem cells. Cell 2009, 138, 286–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiskopf, K.; Jahchan, N.S.; Schnorr, P.J.; Cristea, S.; Ring, A.M.; Maute, R.L.; Volkmer, A.K.; Volkmer, J.P.; Liu, J.; Lim, J.S.; et al. CD47-blocking immunotherapies stimulate macrophage-mediated destruction of small-cell lung cancer. J. Clin. Investig. 2016, 126, 2260–2610. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, A.D.; Beum, P.V.; Solga, M.D.; DiLillo, D.J.; Lindorfer, M.A.; Hess, C.E.; Densmore, J.J.; Williams, M.E.; Taylor, R.P. Rituximab infusion promotes rapid complement depletion and acute CD20 loss in chronic lymphocytic leukemia. J. Immunol. 2004, 172, 3280–3288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beum, P.V.; Mack, D.A.; Pawluczkowycz, A.W.; Lindorfer, M.A.; Taylor, R.P. Binding of rituximab, trastuzumab, cetuximab, or mAbT101 to cancer cells promotes trogocytosis mediated by THP-1 cells and monocytes. J. Immunol. 2008, 181, 8120–8132. [Google Scholar] [CrossRef] [Green Version]
- Boross, P.; Jansen, J.H.; Pastula, A.; van der Poel, C.E.; Leusen, J.H. Both activating and inhibitory Fc gamma receptors mediate rituximab-induced trogocytosis of CD20 in mice. Immunol. Lett. 2012, 143, 44–52. [Google Scholar] [CrossRef]
- Velmurugan, R.; Challa, D.K.; Ram, S.; Ober, R.J.; Ward, E.S. Macrophage-mediated trogocytosis leads to death of antibody-opsonized tumor cells. Mol. Cancer Ther. 2016, 15, 1879–1889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerrard, T.L.; Cohen, D.J.; Kaplan, A.M. Human neutrophil-mediated cytotoxicity to tumor cells. J. Natl. Cancer Inst. 1981, 66, 483–488. [Google Scholar]
- Vols, S.; Sionov, R.V.; Granot, Z. Always look on the bright side: Anti-tumor functions of neutrophils. Curr. Pharm. Des. 2017, 23, 4862–4892. [Google Scholar] [CrossRef]
- Eruslanov, E.B.; Bhojnagarwala, P.S.; Quatromoni, J.G.; Stephen, T.L.; Ranganathan, A.; Deshpande, C.; Akimova, T.; Vachani, A.; Litzky, L.; Hancock, W.W.; et al. Tumor-associated neutrophils stimulate T cell responses in early-stage human lung cancer. J. Clin. Investig. 2014, 124, 5466–5480. [Google Scholar] [CrossRef] [Green Version]
- Sagiv, J.Y.; Michaeli, J.; Assi, S.; Mishalian, I.; Kisos, H.; Levy, L.; Damti, P.; Lumbroso, D.; Polyansky, L.; Sionov, R.V.; et al. Phenotypic diversity and plasticity in circulating neutrophil subpopulations in cancer. Cell Rep. 2015, 10, 562–573. [Google Scholar] [CrossRef] [Green Version]
- Horner, H.; Frank, C.; Dechant, C.; Repp, R.; Glennie, M.; Herrmann, M.; Stockmeyer, B. Intimate cell conjugate formation and exchange of membrane lipids precede apoptosis induction in target cells during antibody-dependent, granulocyte-mediated cytotoxicity. J. Immunol. 2007, 179, 337–345. [Google Scholar] [CrossRef] [Green Version]
- Matlung, H.L.; Babes, L.; Zhao, X.W.; van Houdt, M.; Treffers, L.W.; van Rees, D.J.; Franke, K.; Schornagel, K.; Verkuijlen, P.; Janssen, H.; et al. Neutrophils kill antibody-opsonized cancer cells by trogoptosis. Cell Rep. 2018, 23, 3946–3959. [Google Scholar] [CrossRef] [PubMed]
- Treffers, L.W.; Ten Broeke, T.; Rösner, T.; Jansen, J.H.M.; van Houdt, M.; Kahle, S.; Schornagel, K.; Verkuijlen, P.J.J.H.; Prins, J.M.; Franke, K.; et al. IgA-mediated killing of tumor cells by neutrophils is enhanced by CD47-SIRPα checkpoint inhibition. Cancer Immunol. Res. 2020, 8, 120–130. [Google Scholar] [CrossRef] [Green Version]
- Ustyanovska Avtenyuk, N.; Visser, N.; Bremer, E.; Wiersma, V.R. The neutrophil: The underdog that packs a punch in the fight against cancer. Int. J. Mol. Sci. 2020, 21, 7820. [Google Scholar] [CrossRef] [PubMed]
- Poupot, M.; Fournié, J.J. Spontaneous membrane transfer through homotypic synapses between lymphoma cells. J. Immunol. 2003, 171, 2517–2523. [Google Scholar] [CrossRef] [Green Version]
- LeMaoult, J.; Caumartin, J.; Daouya, M.; Switala, M.; Rebmann, V.; Arnulf, B.; Carosella, E.D. Trogocytic intercellular membrane exchanges among hematological tumors. J. Hematol. Oncol. 2015, 8, 24. [Google Scholar] [CrossRef] [Green Version]
- Long, E.O. Tumor cell recognition by natural killer cells. Semin. Cancer Biol. 2002, 12, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Moretta, L.; Bottino, C.; Pende, D.; Vitale, M.; Mingari, M.C.; Moretta, A. Human natural killer cells: Molecular mechanisms controlling NK cell activation and tumor cell lysis. Immunol. Lett. 2005, 100, 7–13. [Google Scholar] [CrossRef]
- Riteau, B.; Menier, C.; Khalil-Daher, I.; Martinozzi, S.; Pla, M.; Dausset, J.; Carosella, E.D.; Rouas-Freiss, N. HLA-G1 co-expression boosts the HLA class I-mediated NK lysis inhibition. Int. Immunol. 2001, 13, 193–201. [Google Scholar] [CrossRef] [Green Version]
- Caumartin, J.; Favier, B.; Daouya, M.; Guillard, C.; Moreau, P.; Carosella, E.D.; LeMaoult, J. Trogocytosis-based generation of suppressive NK cells. EMBO J. 2007, 26, 1423–1433. [Google Scholar] [CrossRef] [Green Version]
- HoWangYin, K.Y.; Alegre, E.; Daouya, M.; Avier, B.; Carosella, E.D.; LeMaoult, J. Different functional outcomes of intercellular membrane transfers to monocytes and T cells. Cell. Mol. Life Sci. 2010, 67, 1133–1145. [Google Scholar] [CrossRef] [PubMed]
- LeMaoult, J.; Caumartin, J.; Daouya, M.; Favier, B.; Le Rond, S.; Gonzalez, A.; Carosella, E.D. Immune regulation by pretenders: Cell-to-cell transfers of HLA-G make effector T cells act as regulatory cells. Blood 2007, 109, 2040–2048. [Google Scholar] [CrossRef]
- Joshua, D.E.; Brown, R.D.; Ho, P.J.; Gibson, J. Regulatory T cells and multiple myeloma. Clin. Lymphoma Myeloma 2008, 8, 283–286. [Google Scholar] [CrossRef]
- Brown, R.; Kabani, K.; Favaloro, J.; Yang, S.; Ho, P.J.; Gibson, J.; Fromm, P.; Suen, H.; Woodland, N.; Nassif, N.; et al. CD86+ or HLA-G+ can be transferred via trogocytosis from myeloma cells to T cells and are associated with poor prognosis. Blood 2012, 120, 2055–2063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, R.; Suen, H.; Favaloro, J.; Yang, S.; Ho, P.J.; Gibson, J.; Joshua, D. Trogocytosis generates acquired regulatory T cells adding further complexity to the dysfunctional immune response in multiple myeloma. Oncoimmunology 2012, 1, 1658–1660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, A.; Yan, W.H. Intercellular transfer of HLA-G: Its potential in cancer immunology. Clin. Transl. Immunol. 2019, 8, e1077. [Google Scholar] [CrossRef] [Green Version]
- Carter, L.; Fouser, L.A.; Jussif, J.; Fitz, L.; Deng, B.; Wood, C.R.; Collins, M.; Honjo, T.; Freeman, G.J.; Carreno, B.M. PD-1: PD-L inhibitory pathway affects both CD4(+) and CD8(+) T cells and is overcome by IL-2. Eur. J. Immunol. 2002, 32, 634–643. [Google Scholar] [CrossRef]
- Francisco, L.M.; Salinas, V.H.; Brown, K.E.; Vanguri, V.K.; Freeman, G.J.; Kuchroo, V.K.; Sharpe, A.H. PD-L1 regulates the development, maintenance, and function of induced regulatory T cells. J. Exp. Med. 2009, 206, 3015–3029. [Google Scholar] [CrossRef] [PubMed]
- Amarnath, S.; Costanzo, C.M.; Mariotti, J.; Ullman, J.L.; Telford, W.G.; Kapoor, V.; Riley, J.L.; Levine, B.L.; June, C.H.; Fong, T.; et al. Regulatory T cells and human myeloid dendritic cells promote tolerance via programmed death ligand-1. PLoS Biol. 2010, 8, e1000302. [Google Scholar] [CrossRef] [PubMed]
- Hino, R.; Kabashima, K.; Kato, Y.; Yagi, H.; Nakamura, M.; Honjo, T.; Okazaki, T.; Tokura, Y. Tumor cell expression of programmed cell death-1 ligand 1 is a prognostic factor for malignant melanoma. Cancer 2010, 116, 1757–1766. [Google Scholar] [CrossRef]
- Gary, R.; Voelkl, S.; Palmisano, R.; Ullrich, E.; Bosch, J.J.; Mackensen, A. Antigen-specific transfer of functional programmed death ligand 1 from human APCs onto CD8+T cells via trogocytosis. J. Immunol. 2012, 188, 744–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawashima, M.; Carreras, J.; Higuchi, H.; Kotaki, R.; Hoshina, T.; Okuyama, K.; Suzuki, N.; Kakizaki, M.; Miyatake, Y.; Ando, K.; et al. PD-L1/L2 protein levels rapidly increase on monocytes via trogocytosis from tumor cells in classical Hodgkin lymphoma. Leukemia 2020, 34, 2405–2417. [Google Scholar] [CrossRef] [PubMed]
- Rafii, A.; Mirshahi, P.; Poupot, M.; Faussat, A.M.; Simon, A.; Ducros, E.; Mery, E.; Couderc, B.; Lis, R.; Capdet, J.; et al. Oncologic trogocytosis of an original stromal cells induces chemoresistance of ovarian tumours. PLoS ONE 2008, 3, e3894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Otte, A.; Hass, R. Human mesenchymal stroma/stem cells exchange membrane proteins and alter functionality during interaction with different tumor cell lines. Stem Cells Dev. 2015, 24, 1205–1222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melzer, C.; von der Ohe, J.; Hass, R. Concise review: Crosstalk of mesenchymal stroma/stem-like cells with cancer cells provides therapeutic potential. Stem Cells 2018, 36, 951–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melzer, C.; Ohe, J.V.; Hass, R. Altered tumor plasticity after different cancer cell fusions with MSC. Int. J. Mol. Sci. 2020, 21, 8347. [Google Scholar] [CrossRef]
- Hass, R. Role of MSC in the tumor microenvironment. Cancers 2020, 12, 2107. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Zong, S.; Wu, X.; Wei, Y.; Xiang, J. CD4+T cell acquisition of the bystander pMHC I colonizing in the same immunological synapse comprising pMHC II and costimulatory CD40, CD54, CD80, OX40L, and 41BBL. Biochem. Biophys. Res. Commun. 2007, 362, 822–828. [Google Scholar] [CrossRef]
- Osborne, D.G.; Wetzel, S.A. Trogocytosis results in sustained intracellular signaling in CD4(+) T cells. J. Immunol. 2012, 189, 4728–4739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsang, J.Y.; Cha, J.G.; Lechle, R. Antigen presentation by mouse CD4+T cells involving acquired MHC class II: Peptide complexes: Another mechanism to limit clonal expansion? Blood 2003, 101, 2704–2710. [Google Scholar] [CrossRef] [PubMed]
- Carlin, L.M.; Yanagi, K.; Verhoef, A.; Nolte-’t Hoen, E.N.; Yates, J.; Gardner, L.; Lamb, J.; Lombardi, G.; Dallman, M.J.; Davis, D.M. Secretion of IFN-gamma and not IL-2 by anergic human T cells correlates with assembly of an immature immune synapse. Blood 2005, 106, 3874–3879. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, K.A.; Xiang, J. Mechanisms of cellular communication through intercellular protein transfer. J. Cell. Mol. Med. 2011, 15, 1458–1473. [Google Scholar] [CrossRef]
- Masuda, S.; Iwasaki, S.; Tomaru, U.; Baba, T.; Katsumata, K.; Ishizu, A. Possible implication of Fcγ receptor-mediated trogocytosis in susceptibility to systemic autoimmune disease. Clin. Dev. Immunol. 2013, 2013, 345745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haastert, B.; Mellanby, R.J.; Anderton, S.M.; O’Connor, R.A. T cells at the site of autoimmune inflammation show increased potential for trogocytosis. PLoS ONE 2013, 8, e81404. [Google Scholar] [CrossRef]
- LeMaoult, J.; Le Discorde, M.; Rouas-Freiss, N.; Moreau, P.; Menier, C.; McCluskey, J.; Carosella, E.D. Biology and functions of human leukocyte antigen-G in health and sickness. Tissue Antigens 2003, 62, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Carosella, E.D.; Moreau, P.; Lemaoult, J.; Rouas-Freiss, N. HLA-G: From biology to clinical benefits. Trends Immunol. 2008, 29, 125–132. [Google Scholar] [CrossRef]
- Kovats, S.; Main, E.K.; Librach, C.; Stubblebine, M.; Fisher, S.J.; DeMars, R. A class I antigen, HLA-G, expressed in human trophoblasts. Science 1990, 248, 220–223. [Google Scholar] [CrossRef] [PubMed]
- Crisa, L.; McMaster, M.T.; Ishii, J.K.; Fisher, S.J.; Salomon, D.R. Identification of a thymic epithelial cell subset sharing expression of the class Ib HLA-G molecule with fetal trophoblasts. J. Exp. Med. 1997, 186, 289–298. [Google Scholar] [CrossRef] [Green Version]
- Le Discorde, M.; Moreau, P.; Sabatier, P.; Legeais, J.M.; Carosella, E.D. Expression of HLA-G in human cornea, an immune-privileged tissue. Hum. Immunol. 2003, 64, 1039–1044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirszenbaum, M.; Moreau, P.; Gluckman, E.; Dausset, J.; Carosella, E. An alternatively spliced form of HLA-G mRNA in human trophoblasts and evidence for the presence of HLA-G transcript in adult lymphocytes. Proc. Natl. Acad. Sci. USA 1994, 91, 4209–4213. [Google Scholar] [CrossRef] [Green Version]
- Colonna, M.; Samaridis, J.; Cella, M.; Angman, L.; Allen, R.L.; O’Callaghan, C.A.; Dunbar, R.; Ogg, G.S.; Cerundolo, V.; Rolink, A. Human myelomonocytic cells express an inhibitory receptor for classical and nonclassical MHC class I molecules. J. Immunol. 1998, 160, 3096–3100. [Google Scholar] [PubMed]
- Rajagopalan, S.; Long, E.O. A human histocompatibility leukocyte antigen (HLA)-G-specific receptor expressed on all natural killer cells. J. Exp. Med. 1999, 189, 1093–1100. [Google Scholar] [CrossRef] [Green Version]
- LeMaoult, J.; Krawice-Radanne, I.; Dausset, J.; Carosella, E.D. HLA-G1-expressing antigen-presenting cells induce immunosuppressive CD4+T cells. Proc. Natl. Acad. Sci. USA 2004, 101, 7064–7069. [Google Scholar] [CrossRef] [Green Version]
- Monsiváis-Urenda, A.E.; Baranda, L.; Alvarez-Quiroga, C.; Abud-Mendoza, C.; González-Amaro, R. Expression and functional role of HLA-G in immune cells from patients with systemic lupus erythematosus. J. Clin. Immunol. 2011, 31, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Rosado, S.; Perez-Chacon, G.; Mellor-Pita, S.; Sanchez-Vegazo, I.; Bellas-Menendez, C.; Citores, M.J.; Losada-Fernandez, I.; Martin-Donaire, T.; Rebolleda, N.; Perez-Aciego, P. Expression of human leukocyte antigen-G in systemic lupus erythematosus. Hum. Immunol. 2008, 69, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Negrini, S.; Contini, P.; Pupo, F.; Greco, M.; Murdaca, G.; Puppo, F. Expression of membrane-bound human leucocyte antigen-G in systemic sclerosis and systemic lupus erythematosus. Hum. Immunol. 2020, 81, 162–167. [Google Scholar] [CrossRef]
- Rossi, E.A.; Chang, C.H.; Goldenberg, D.M. Anti-CD22/CD20 bispecific antibody with enhanced trogocytosis for treatment of lupus. PLoS ONE 2014, 9, e98315. [Google Scholar] [CrossRef] [Green Version]
- Wiendl, H.; Hohlfeld, R.; Kieseier, B.C. Muscle-derived positive and negative regulators of the immune response. Curr. Opin. Rheumatol. 2005, 17, 714–719. [Google Scholar] [CrossRef]
- Arahata, K.; Engel, A.G. Monoclonal antibody analysis of mononuclear cells in myopathies. I: Quantitation of subsets according to diagnosis and sites of accumulation and demonstration and counts of muscle fibers invaded by T cells. Ann. Neurol. 1984, 16, 193–208. [Google Scholar] [CrossRef] [PubMed]
- Waschbisch, A.; Meuth, S.G.; Herrmann, A.M.; Wrobel, B.; Schwab, N.; Lochmüller, H.; Wiendl, H. Intercellular exchanges of membrane fragments (trogocytosis) between human muscle cells and immune cells: A potential mechanism for the modulation of muscular immune responses. J. Neuroimmunol. 2009, 209, 131–138. [Google Scholar] [CrossRef]
- Clark, D.A.; Dessypris, E.N.; Jenkins, D.E., Jr.; Krantz, S.B. Acquired immune hemolytic anemia associated with IgA erythrocyte coating: Investigation of hemolytic mechanisms. Blood 1984, 64, 1000–1005. [Google Scholar] [CrossRef] [PubMed]
- Chadebech, P.; Michel, M.; Janvier, D.; Yamada, K.; Copie-Bergman, C.; Bodivit, G.; Bensussan, A.; Fournie, J.J.; Godeau, B.; Bierling, P.; et al. IgA-mediated human autoimmune hemolytic anemia as a result of hemagglutination in the spleen, but independent of complement activation and FcαRI. Blood 2010, 116, 4141–4147. [Google Scholar] [CrossRef] [PubMed]
- Shoelson, S.E.; Herrero, L.; Naaz, A. Obesity, inflammation, and insulin resistance. Gastroenterology 2007, 132, 2169–2180. [Google Scholar] [CrossRef] [PubMed]
- Sárvári, A.K.; Doan-Xuan, Q.M.; Bacsó, Z.; Csomós, I.; Balajthy, Z.; Fésüs, L. Interaction of differentiated human adipocytes with macrophages leads to trogocytosis and selective IL-6 secretion. Cell Death Dis. 2015, 6, e1613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engin, A.B. Adipocyte-macrophage cross-talk in obesity. Adv. Exp. Med. Biol. 2017, 960, 327–343. [Google Scholar]
- Lu, Y.; Ma, J.; Zhao, J.; Song, Z.; Zhou, C.; Liu, X.; Teng, W.; Wang, W.; Zhang, Q.; Yan, W.; et al. The Role of MKP-5 in adipocyte-macrophage interactions during obesity. Obes. Facts 2020, 13, 86–101. [Google Scholar] [CrossRef] [PubMed]
- Monnet, C.; Jorieux, S.; Souyris, N.; Zaki, O.; Jacquet, A.; Fournier, N.; Crozet, F.; de Romeuf, C.; Bouayadi, K.; Urbain, R.; et al. Combined glycol- and protein-Fc engineering simultaneously enhance cytotoxicity and half-life of a therapeutic antibody. MAbs 2014, 6, 422–436. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Xu, L.; Tao, F.; Xie, K.; Wu, Z.; Li, Y.; Li, J.; Chen, K.; Pi, C.; Mendelsohn, A.; et al. Simultaneous exposure to FcγR and FcαR on monocytes and macrophages enhances antitumor activity in vivo. Oncotarget 2017, 8, 39356–39366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Bethune, M.; Wong, S.; Joglekar, A.V.; Leonard, M.T.; Wang, J.K.; Kim, J.T.; Cheng, D.; Peng, S.; Zaretsky, J.M. T cell antigen discovery via trogocytosis. Nat. Methods 2019, 15, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Petty, A.J.; Heyman, B.; Yang, Y. Chimeric antigen receptor cell therapy: Overcoming obstacles to battle cancer. Cancers 2020, 12, 842. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Disease Entity | Abnormal Trogocytosis-Mediated Adverse Effects |
|---|---|
| Systemic lupus erythematosus | -Decreased HLA-G molecule expression on monocytes and CD83+ dendritic cell [149] -Diminished lymphocytes trogocytosis of HLA-G molecules from autologous monocytes [149] -Increased % of HLA-G expressing PBMC reflects a compensatory down-regulated hyperactive immune status [150,151] |
| Autoimmune myopathy | -Increased trogocytosis between myoblasts and T lymphocytes [155] |
| IgA autoantibody –mediated hemolytic anemia | -Increased RBC membrane trogocytosis to monocytes and sequestration in spleen [157] |
| Obesity | -Increased adipocyte to macrophage trogocytosis in enhancing IL-6 and MCP-1 production by macrophages [158,159] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, K.-J.; Wu, C.-H.; Lu, C.-H.; Shen, C.-Y.; Kuo, Y.-M.; Tsai, C.-Y.; Hsieh, S.-C.; Yu, C.-L. Trogocytosis between Non-Immune Cells for Cell Clearance, and among Immune-Related Cells for Modulating Immune Responses and Autoimmunity. Int. J. Mol. Sci. 2021, 22, 2236. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052236
Li K-J, Wu C-H, Lu C-H, Shen C-Y, Kuo Y-M, Tsai C-Y, Hsieh S-C, Yu C-L. Trogocytosis between Non-Immune Cells for Cell Clearance, and among Immune-Related Cells for Modulating Immune Responses and Autoimmunity. International Journal of Molecular Sciences. 2021; 22(5):2236. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052236
Chicago/Turabian StyleLi, Ko-Jen, Cheng-Han Wu, Cheng-Hsun Lu, Chieh-Yu Shen, Yu-Min Kuo, Chang-Youh Tsai, Song-Chou Hsieh, and Chia-Li Yu. 2021. "Trogocytosis between Non-Immune Cells for Cell Clearance, and among Immune-Related Cells for Modulating Immune Responses and Autoimmunity" International Journal of Molecular Sciences 22, no. 5: 2236. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052236