
Does a Red House Affect Rhythms in Mice with a Corrupted Circadian System?

 and
and
Abstract
:1. Introduction
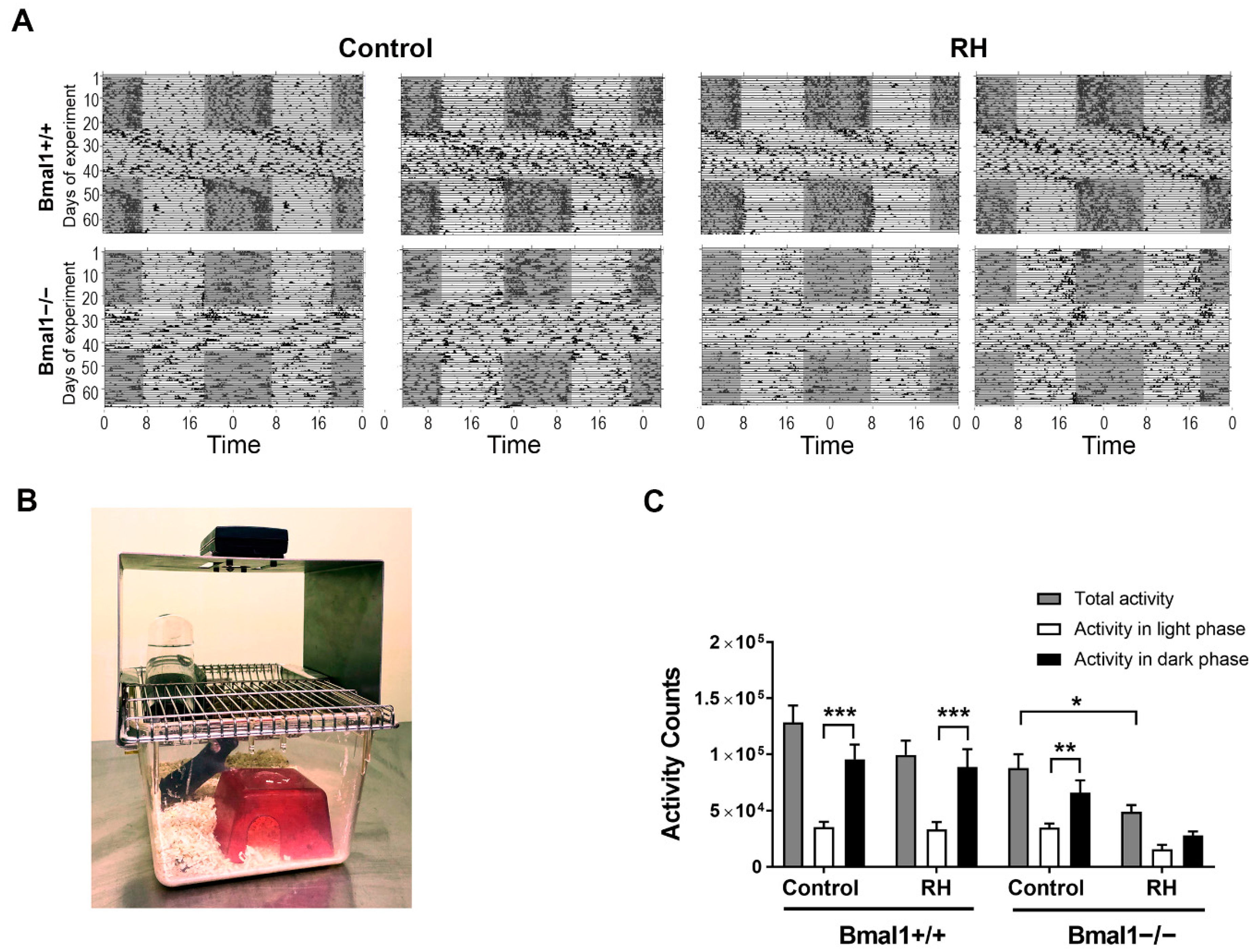
2. Results
2.1. Spontaneous Locomotor Activity Is Affected by RH
2.2. Expression of mPER1 and p-ERK in SCN Is Affected by RH
2.3. Plasma Corticosterone Level Is Not Affected by RH
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Behavioral Analysis
4.3. Tissue Processing
4.4. Immunohistochemistry
4.5. Image Analysis
4.6. Plasma Corticosterone Analysis
4.7. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Balsalobre, A. Resetting of Circadian Time in Peripheral Tissues by Glucocorticoid Signaling. Science 2000, 289, 2344–2347. [Google Scholar] [CrossRef] [Green Version]
- Engeland, W.C.; Massman, L.; Miller, L.; Leng, S.; Pignatti, E.; Pantano, L.; Carlone, D.L.; Kofuji, P.; Breault, D.T. Sex Differences in Adrenal Bmal1 Deletion–Induced Augmentation of Glucocorticoid Responses to Stress and ACTH in Mice. Endocrinology 2019, 160, 2215–2229. [Google Scholar] [CrossRef]
- Dumbell, R.; Leliavski, A.; Matveeva, O.; Blaum, C.; Tsang, A.H.; Oster, H. Dissociation of Molecular and Endocrine Circadian Rhythms in Male Mice Lacking Bmal1 in the Adrenal Cortex. Endocrinology 2016, 157, 4222–4233. [Google Scholar] [CrossRef] [Green Version]
- Gong, S.; Miao, Y.-L.; Jiao, G.-Z.; Sun, M.-J.; Li, H.; Lin, J.; Luo, M.-J.; Tan, J.-H. Dynamics and Correlation of Serum Cortisol and Corticosterone under Different Physiological or Stressful Conditions in Mice. PLoS ONE 2015, 10, e0117503. [Google Scholar] [CrossRef] [PubMed]
- Ralph, C.R.; Tilbrook, A.J. INVITED REVIEW: The usefulness of measuring glucocorticoids for assessing animal welfare. J. Anim. Sci. 2016, 94, 457–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reppert, S.M.; Weaver, D.R. Coordination of circadian timing in mammals. Nature 2002, 418, 935–941. [Google Scholar] [CrossRef] [PubMed]
- Gekakis, N.; Staknis, D.; Nguyen, H.B.; Davis, F.C.; Wilsbacher, L.D.; King, D.P.; Takahashi, J.S.; Weitz, C.J. Role of the CLOCK protein in the mammalian circadian mechanism. Science 1998, 280, 1564–1569. [Google Scholar] [CrossRef]
- HogenEsch, J.B.; Gu, Y.-Z.; Jain, S.; Bradfield, C.A. The basic-helix-loop-helix-PAS orphan MOP3 forms transcriptionally active complexes with circadian and hypoxia factors. Proc. Natl. Acad. Sci. USA 1998, 95, 5474–5479. [Google Scholar] [CrossRef] [Green Version]
- Jin, X.; Shearman, L.P.; Weaver, D.R.; Zylka, M.J.; De Vries, G.J.; Reppert, S.M. A molecular mechanism regulating rhythmic output from the suprachiasmatic circadian clock. Cell 1999, 96, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Kume, K.; Zylka, M.J.; Sriram, S.; Shearman, L.P.; Weaver, D.R.; Jin, X.; Maywood, E.S.; Hastings, M.H.; Reppert, S.M. mCRY1 and mCRY2 Are Essential Components of the Negative Limb of the Circadian Clock Feedback Loop. Cell 1999, 98, 193–205. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.; Etchegaray, J.-P.; Cagampang, F.R.A.; Loudon, A.S.I.; Reppert, S.M. Posttranslational mechanisms regulate the mammalian circadian clock. Cell 2001, 107, 855–867. [Google Scholar] [CrossRef]
- Tosini, G.; Menaker, M. Circadian Rhythms in Cultured Mammalian Retina. Science 1996, 272, 419–421. [Google Scholar] [CrossRef]
- Tosini, G.; Pozdeyev, N.; Sakamoto, K.; Iuvone, P.M. The circadian clock system in the mammalian retina. BioEssays 2008, 30, 624–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baba, K.; Piano, I.; Lyuboslavsky, P.; Chrenek, M.A.; Sellers, J.T.; Zhang, S.; Gargini, C.; He, L.; Tosini, G.; Iuvone, P.M. Removal of clock gene Bmal1 from the retina affects retinal development and accelerates cone photoreceptor degeneration during aging. Proc. Natl. Acad. Sci. USA 2018, 115, 13099–13104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baba, K.; Ribelayga, C.P.; Iuvone, P.M.; Tosini, G. The Retinal Circadian Clock and Photoreceptor Viability. Adv. Exp. Med. Biol. 2018, 1074, 345–350. [Google Scholar] [CrossRef]
- McMahon, D.G.; Iuvone, P.M.; Tosini, G. Circadian organization of the mammalian retina: From gene regulation to physiology and diseases. Prog. Retin. Eye Res. 2014, 39, 58–76. [Google Scholar] [CrossRef] [Green Version]
- Aschoff, J.; Hoffmann, K.; Pohl, H.; Wever, R. Re-entrainment of circadian rhythms after phase-shifts of the Zeitgeber. Chronobiologia 1975, 2, 23–78. [Google Scholar]
- Chen, D.; Buchanan, G.F.; Ding, J.M.; Hannibal, J.; Gillette, M.U. Pituitary adenylyl cyclase-activating peptide: A pivotal modulator of glutamatergic regulation of the suprachiasmatic circadian clock. Proc. Natl. Acad. Sci. USA 1999, 96, 13468–13473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hannibal, J. Neurotransmitters of the retino-hypothalamic tract. Cell Tissue Res. 2002, 309, 73–88. [Google Scholar] [CrossRef]
- Ginty, D.; Kornhauser, J.; Thompson, M.; Bading, H.; Mayo, K.; Takahashi, J.; Greenberg, M. Regulation of CREB phosphorylation in the suprachiasmatic nucleus by light and a circadian clock. Science 1993, 260, 238–241. [Google Scholar] [CrossRef]
- Ding, J.M.; Faiman, L.E.; Hurst, W.J.; Kuriashkina, L.R.; Gillette, M.U. Resetting the Biological Clock: Mediation of Nocturnal CREB Phosphorylation via Light, Glutamate, and Nitric Oxide. J. Neurosci. 1997, 17, 667–675. [Google Scholar] [CrossRef] [PubMed]
- Gau, D.; Lemberger, T.; Von Gall, C.; Kretz, O.; Le Minh, N.; Gass, P.; Schmid, W.; Schibler, U.; Korf, H.W.; Schütz, G. Phosphorylation of CREB Ser142 Regulates Light-Induced Phase Shifts of the Circadian Clock. Neuron 2002, 34, 245–253. [Google Scholar] [CrossRef] [Green Version]
- Coogan, A.N.; Piggins, H.D. Circadian and photic regulation of phosphorylation of ERK1/2 and Elk-1 in the suprachiasmatic nuclei of the Syrian hamster. J. Neurosci. 2003, 23, 3085–3093. [Google Scholar] [CrossRef]
- Reppert, S.M.; Weaver, D.R. Molecular Analysis of Mammalian Circadian Rhythms. Annu. Rev. Physiol. 2001, 63, 647–676. [Google Scholar] [CrossRef]
- Shuboni, D.; Cramm, S.; Yan, L.; Nunez, A.; Smale, L. Acute Behavioral Responses to Light and Darkness in NocturnalMus musculusand DiurnalArvicanthis niloticus. J. Biol. Rhythm. 2012, 27, 299–307. [Google Scholar] [CrossRef]
- Aschoff, J. Masking and Parametric Effects of High-Frequency Light-Dark Cycles. Jpn. J. Physiol. 1999, 49, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Phillips, A.J.K.; Fulcher, B.D.; Robinson, P.A.; Klerman, E.B. Mammalian Rest/Activity Patterns Explained by Physiologically Based Modeling. PLoS Comput. Biol. 2013, 9, e1003213. [Google Scholar] [CrossRef] [Green Version]
- DeCoursey, P.J. Light-sampling behavior in photoentrainment of a rodent circadian rhythm. J. Comp. Physiol. A 1986, 159, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Berson, D.M.; Dunn, F.A.; Takao, M. Phototransduction by Retinal Ganglion Cells That Set the Circadian Clock. Science 2002, 295, 1070–1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hattar, S.; Liao, H.-W.; Takao, M.; Berson, D.M.; Yau, K.-W. Melanopsin-containing retinal ganglion cells: Architecture, projections, and intrinsic photosensitivity. Science 2002, 295, 1065–1070. [Google Scholar] [CrossRef] [Green Version]
- Lucas, R.J.; Douglas, R.H.; Foster, R.G. Characterization of an ocular photopigment capable of driving pupillary constriction in mice. Nat. Neurosci. 2001, 4, 621–626. [Google Scholar] [CrossRef]
- Lall, G.S.; Revell, V.L.; Momiji, H.; Al Enezi, J.; Altimus, C.M.; Güler, A.D.; Aguilar, C.; Cameron, M.A.; Allender, S.; Hankins, M.W.; et al. Distinct Contributions of Rod, Cone, and Melanopsin Photoreceptors to Encoding Irradiance. Neuron 2010, 66, 417–428. [Google Scholar] [CrossRef]
- Bailes, H.J.; Lucas, R.J. Human melanopsin forms a pigment maximally sensitive to blue light (λ max ≈ 479 nm) supporting activation of G q /11 and G i/o signalling cascades. Proc. R. Soc. B Biol. Sci. 2013, 280, 20122987. [Google Scholar] [CrossRef] [Green Version]
- Gooley, J.J.; Rajaratnam, S.M.W.; Brainard, G.C.; Kronauer, R.E.; Czeisler, C.A.; Lockley, S.W. Spectral Responses of the Human Circadian System Depend on the Irradiance and Duration of Exposure to Light. Sci. Transl. Med. 2010, 2, 31ra33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mouland, J.W.; Martial, F.; Watson, A.; Lucas, R.J.; Brown, T.M. Cones Support Alignment to an Inconsistent World by Suppressing Mouse Circadian Responses to the Blue Colors Associated with Twilight. Curr. Biol. 2019, 29, 4260–4267.e4. [Google Scholar] [CrossRef] [Green Version]
- Spitschan, M.; Lucas, R.J.; Brown, T.M. Chromatic clocks: Color opponency in non-image-forming visual function. Neurosci. Biobehav. Rev. 2017, 78, 24–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bunger, M.K.; Wilsbacher, L.D.; Moran, S.M.; Clendenin, C.; Radcliffe, L.A.; HogenEsch, J.B.; Simon, M.C.; Takahashi, J.S.; Bradfield, C.A. Mop3 is an essential component of the master circadian pacemaker in mammals. Cell 2020, 103, 1009–1017. [Google Scholar] [CrossRef] [Green Version]
- Kondratov, R.V.; Kondratova, A.A.; Gorbacheva, V.Y.; Vykhovanets, O.V.; Antoch, M.P. Early aging and age-related pathologies in mice deficient in BMAL1, the core componentof the circadian clock. Genes Dev. 2006, 20, 1868–1873. [Google Scholar] [CrossRef] [Green Version]
- Pfeffer, M.; Korf, H.-W.; Von Gall, C. Chronotype and stability of spontaneous locomotor activity rhythm in BMAL1-deficient mice. Chronobiol. Int. 2014, 32, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Pfeffer, M.; Müller, C.M.; Mordel, J.; Meissl, H.; Ansari, N.; Deller, T.; Korf, H.-W.; Von Gall, C. The Mammalian Molecular Clockwork Controls Rhythmic Expression of Its Own Input Pathway Components. J. Neurosci. 2009, 29, 6114–6123. [Google Scholar] [CrossRef] [PubMed]
- Storch, K.-F.; Paz, C.; Signorovitch, J.; Raviola, E.; Pawlyk, B.; Li, T.; Weitz, C.J. Intrinsic Circadian Clock of the Mammalian Retina: Importance for Retinal Processing of Visual Information. Cell 2007, 130, 730–741. [Google Scholar] [CrossRef] [Green Version]
- Langgartner, D.; Foertsch, S.; Füchsl, A.M.; Reber, S.O. Light and water are not simple conditions: Fine tuning of animal housing in male C57BL/6 mice. Stress 2016, 20, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Scariot, P.P.M.; Manchado-Gobatto, F.B.; Prolla, T.A.; Dos Reis, I.G.M.; Gobatto, C.A. Housing conditions modulate spontaneous physical activity, feeding behavior, aerobic running capacity and adiposity in C57BL/6J mice. Horm. Behav. 2019, 115, 104556. [Google Scholar] [CrossRef]
- Wolfer, D.P.; Litvin, O.; Morf, S.; Nitsch, R.M.; Lipp, H.-P.; Würbel, H.; Wuerbel, H. Cage enrichment and mouse behaviour. Nat. Cell Biol. 2004, 432, 821–822. [Google Scholar] [CrossRef] [PubMed]
- Bayne, K. Environmental enrichment and mouse models: Current perspectives. Anim. Model. Exp. Med. 2018, 1, 82–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wirz, A.; Mandillo, S.; D’Amato, F.R.; Giuliani, A.; Riviello, M.C. Response, use and habituation to a mouse house in C57BL/6J and BALB/c mice. Exp. Anim. 2015, 64, 281–293. [Google Scholar] [CrossRef] [Green Version]
- André, V.; Gau, C.; Scheideler, A.; Aguilar-Pimentel, J.A.; Amarie, O.V.; Becker, L.; Garrett, L.; Hans, W.; Hölter, S.M.; Janik, D.; et al. Laboratory mouse housing conditions can be improved using common environmental enrichment without compromising data. PLoS Biol. 2018, 16, e2005019. [Google Scholar] [CrossRef] [Green Version]
- Storch, K.-F.; Paz, C.; Signorovitch, J.; Raviola, E.; Pawlyk, B.; Li, T.; Weitz, C.J. Physiological Importance of a Circadian Clock Outside the Suprachiasmatic Nucleus. Cold Spring Harb. Symp. Quant. Biol. 2007, 72, 307–318. [Google Scholar] [CrossRef]
- Walmsley, L.; Hanna, L.; Mouland, J.; Martial, F.; West, A.; Smedley, A.R.; Bechtold, D.A.; Webb, A.R.; Lucas, R.J.; Brown, T.M. Colour As a Signal for Entraining the Mammalian Circadian Clock. PLoS Biol. 2015, 13, e1002127. [Google Scholar] [CrossRef] [PubMed]
- Mrosovsky, P.N.; Hattar, S. Impaired Masking Responses to Light in Melanopsin-Knockout Mice. Chronobiol. Int. 2003, 20, 989–999. [Google Scholar] [CrossRef]
- Mrosovsky, N.; Thompson, S. Negative and positive masking responses to light in retinal degenerate slow (rds/rds) mice during aging. Vis. Res. 2008, 48, 1270–1273. [Google Scholar] [CrossRef] [Green Version]
- Hannibal, J.; Norn, T.H.B.; Georg, B.; Fahrenkrug, J. Spatiotemporal expression pattern of PERIOD 1 and PERIOD 2 in the mouse SCN is dependent on VIP receptor 2 signaling. Eur. J. Neurosci. 2019, 50, 3115–3132. [Google Scholar] [CrossRef]
- Hastings, M.H.; Field, M.D.; Maywood, E.S.; Weaver, D.R.; Reppert, S.M. Differential Regulation of mPER1 and mTIM Proteins in the Mouse Suprachiasmatic Nuclei: New Insights into a Core Clock Mechanism. J. Neurosci. 1999, 19, RC11. [Google Scholar] [CrossRef]
- Smith, V.M.; Jeffers, R.T.; Wu, C.; Shankara, J.V.; Antle, M.C. Temporal changes of light-induced proteins in the SCN following treatment with the serotonin mixed agonist/antagonist BMY7378. Exp. Brain Res. 2015, 233, 2723–2731. [Google Scholar] [CrossRef] [PubMed]
- Goldsmith, C.S.; Bell-Pedersen, D. Diverse Roles for MAPK Signaling in Circadian Clocks. Adv. Genet. 2013, 84, 1–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sollars, P.J.; Weiser, M.J.; Kudwa, A.E.; Bramley, J.R.; Ogilvie, M.D.; Spencer, R.L.; Handa, R.J.; Pickard, G.E. Altered Entrainment to the Day/Night Cycle Attenuates the Daily Rise in Circulating Corticosterone in the Mouse. PLoS ONE 2014, 9, e111944. [Google Scholar] [CrossRef]
- Chen, R.; Seo, N.-O.; Bell, E.; Von Gall, C.; Lee, C. Strong Resetting of the Mammalian Clock by Constant Light Followed by Constant Darkness. J. Neurosci. 2008, 28, 11839–11847. [Google Scholar] [CrossRef] [PubMed]
- Ohta, H.; Yamazaki, S.; McMahon, D.G. Constant light desynchronizes mammalian clock neurons. Nat. Neurosci. 2005, 8, 267–269. [Google Scholar] [CrossRef] [PubMed]
- Ingenwerth, M.; Reinbeck, A.L.; Stahr, A.; Partke, H.-J.; Roden, M.; Burkart, V.; Von Gall, C. Perturbation of the molecular clockwork in the SCN of non-obese diabetic mice prior to diabetes onset. Chronobiol. Int. 2016, 33, 1369–1375. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Bmal1+/+ Control | Bmal1+/+ RH | Bmal1−/− Control | Bmal1−/− RH | |
|---|---|---|---|---|
| ZT02 | 2.8 ± 0.9 | 2.7 ± 0.5 | 1.9 ± 0.4 | 1.6 ± 0.2 |
| ZT14 | 7.1 ± 0.1 | 6.9 ± 0.9 | 6.6 ± 1.4 | 7.6 ± 0.7 |
| p-value | 0.03 | 0.008 | 0.02 | 0.008 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Öztürk, M.; Ingenwerth, M.; Sager, M.; von Gall, C.; Ali, A.A.H. Does a Red House Affect Rhythms in Mice with a Corrupted Circadian System? Int. J. Mol. Sci. 2021, 22, 2288. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052288
Öztürk M, Ingenwerth M, Sager M, von Gall C, Ali AAH. Does a Red House Affect Rhythms in Mice with a Corrupted Circadian System? International Journal of Molecular Sciences. 2021; 22(5):2288. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052288
Chicago/Turabian StyleÖztürk, Menekse, Marc Ingenwerth, Martin Sager, Charlotte von Gall, and Amira A. H. Ali. 2021. "Does a Red House Affect Rhythms in Mice with a Corrupted Circadian System?" International Journal of Molecular Sciences 22, no. 5: 2288. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052288