
Oxytocin Signaling as a Target to Block Social Defeat-Induced Increases in Drug Abuse Reward


Abstract
:1. Oxytocin as a Promising Therapeutic Treatment for Addiction Disorders
1.1. Endogenous Oxytocin (OTX) System
1.2. Drug Exposure Alters Oxytocin Neurotransmission
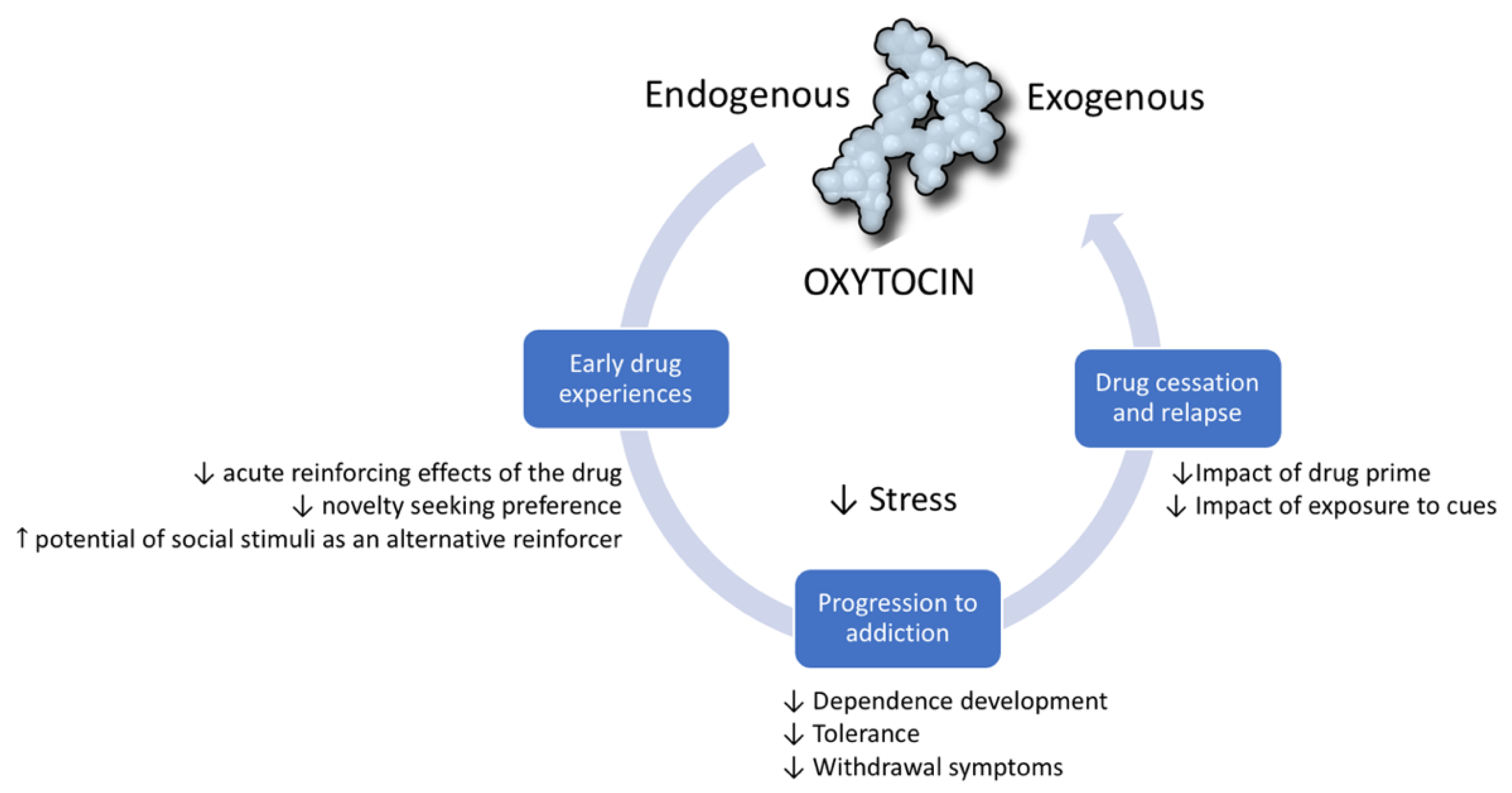
1.3. OXT Modulates the Addictive Cycle
1.3.1. OXT in Early Stages of Addiction
1.3.2. OXT in the Progression to Addiction
1.3.3. OXT in the Prevention of Reinstatement and Relapse into Drug Use
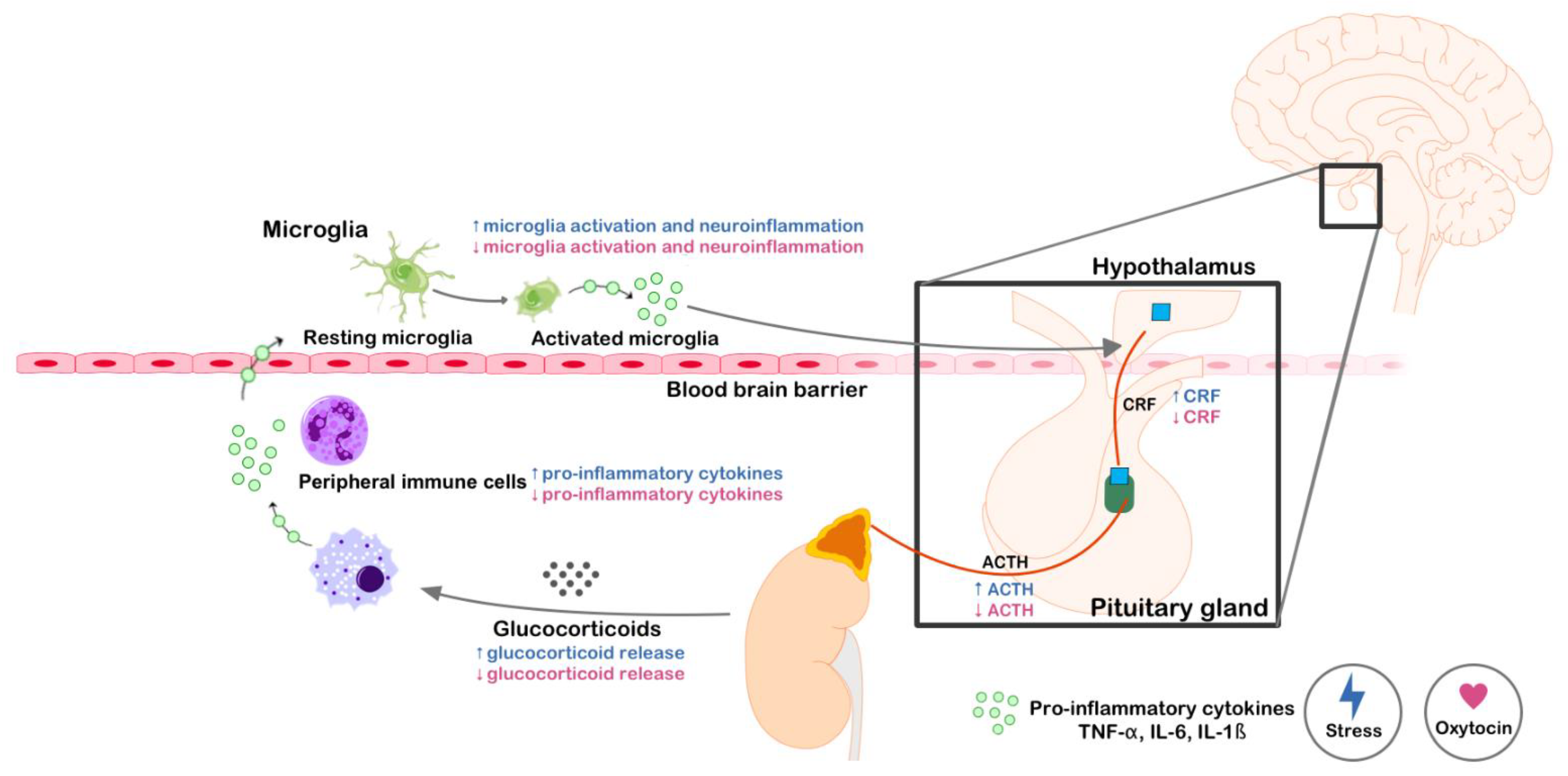
2. The Anti-Stress Potential of Oxytocin
2.1. What Is Social Stress?
2.2. Brain Mechanisms Activated by Social Stress
2.3. Role of Oxytocin in Social Stress
2.4. Exogenous OXT Administration as a Therapeutic Target for Social Stress Disorders
3. Oxytocin Blocks Increased Drug Intake Induced by Social Defeat
4. Final Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ACTH | Adrenocorticotropic hormone |
| BNST | Bed nucleus of the stria terminalis |
| CBP | CREB-binding protein |
| CPP | Conditioned place preference |
| CREB | cAMP response element-binding protein |
| CRF | Corticotropin-releasing factor |
| DA | Dopamine |
| DARPP-32 | Dopamine and cAMP-regulated phosphoprotein of 32 kDa |
| fMRI | Functional magnetic resonance imaging |
| GC | Glucocorticoids |
| HAT | Histone acetyltransferase |
| HDAC | Histone deacetylases |
| HPA | Hypothalamic-pituitary-adrenal |
| LS | Lateral septum |
| MDD | Major depression disorder |
| MeA | Medial amygdala |
| mPFC | Medial prefrontal cortex |
| NAc | Nucleus accumbens |
| OXT | Oxytocin |
| OXTR | Oxytocin receptor |
| PFC | Prefrontal cortex |
| PKA | Protein kinase A |
| PVN | Paraventricular nucleus |
| SA | Self-administration |
| SD | Social defeat |
| SON | Supraoptic nucleus |
| VTA | Ventral tegmental area |
References
- Jurek, B.; Neumann, I.D. The Oxytocin Receptor: From Intracellular Signaling to Behavior. Physiol. Rev. 2018, 98, 1805–1908. [Google Scholar] [CrossRef] [PubMed]
- Gimpl, G.; Fahrenholz, F. The Oxytocin Receptor System: Structure, Function, and Regulation. Physiol. Rev. 2001, 81, 629–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dumais, K.M.; Veenema, A.H. Vasopressin and Oxytocin Receptor Systems in the Brain: Sex Differences and Sex-Specific Regulation of Social Behavior. Front. Neuroendocrinol. 2016, 40, 1–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quintana, D.S.; Guastella, A.J. An Allostatic Theory of Oxytocin. Trends Cogn. Sci. 2020, 24, 515–528. [Google Scholar] [CrossRef]
- Lee, M.R.; Weerts, E.M. Oxytocin for the Treatment of Drug and Alcohol Use Disorders. Behav. Pharmacol. 2016, 27, 640–648. [Google Scholar] [CrossRef] [Green Version]
- King, C.E.; Gano, A.; Becker, H.C. The Role of Oxytocin in Alcohol and Drug Abuse. Brain Res. 2020, 1736, 146761. [Google Scholar] [CrossRef]
- Johns, J.M.; Caldwell, J.D.; Pedersen, C.A. Acute Cocaine Treatment Decreases Oxytocin Levels in the Rat Hippocampus. Neuropeptides 1993, 24, 165–169. [Google Scholar] [CrossRef]
- Baracz, S.J.; Parker, L.M.; Suraev, A.S.; Everett, N.A.; Goodchild, A.K.; McGregor, I.S.; Cornish, J.L. Chronic Methamphetamine Self-Administration Dysregulates Oxytocin Plasma Levels and Oxytocin Receptor Fibre Density in the Nucleus Accumbens Core and Subthalamic Nucleus of the Rat. J. Neuroendocrinol. 2016, 28. [Google Scholar] [CrossRef]
- Georgiou, P.; Zanos, P.; Hourani, S.; Kitchen, I.; Bailey, A. Cocaine Abstinence Induces Emotional Impairment and Brain Region-Specific Upregulation of the Oxytocin Receptor Binding. Eur. J. Neurosci. 2016, 44, 2446–2454. [Google Scholar] [CrossRef] [Green Version]
- McGregor, I.S.; Callaghan, P.D.; Hunt, G.E. From Ultrasocial to Antisocial: A Role for Oxytocin in the Acute Reinforcing Effects and Long-Term Adverse Consequences of Drug Use? Br. J. Pharmacol. 2008, 154, 358–368. [Google Scholar] [CrossRef]
- Clarke, G.; Wood, P.; Merrick, L.; Lincoln, D.W. Opiate Inhibition of Peptide Release from the Neurohumoral Terminals of Hypothalamic Neurones. Nature 1979, 282, 746–748. [Google Scholar] [CrossRef]
- Kovács, G.L.; Laczi, F.; Vecsernyés, M.; Hódi, K.; Telegdy, G.; László, F.A. Limbic Oxytocin and Arginine 8-Vasopressin in Morphine Tolerance and Dependence. Exp. Brain Res. 1987, 65, 307–311. [Google Scholar] [CrossRef]
- Zanos, P.; Georgiou, P.; Weber, C.; Robinson, F.; Kouimtsidis, C.; Niforooshan, R.; Bailey, A. Oxytocin and Opioid Addiction Revisited: Old Drug, New Applications. Br. J. Pharmacol. 2018, 175, 2809–2824. [Google Scholar] [CrossRef]
- Zanos, P.; Georgiou, P.; Wright, S.R.; Hourani, S.M.; Kitchen, I.; Winsky-Sommerer, R.; Bailey, A. The Oxytocin Analogue Carbetocin Prevents Emotional Impairment and Stress-Induced Reinstatement of Opioid-Seeking in Morphine-Abstinent Mice. Neuropsychopharmacology 2014, 39, 855–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansson, A.C.; Koopmann, A.; Uhrig, S.; Bühler, S.; Domi, E.; Kiessling, E.; Ciccocioppo, R.; Froemke, R.C.; Grinevich, V.; Kiefer, F.; et al. Oxytocin Reduces Alcohol Cue-Reactivity in Alcohol-Dependent Rats and Humans. Neuropsychopharmacology 2018, 43, 1235–1246. [Google Scholar] [CrossRef] [Green Version]
- Butovsky, E.; Juknat, A.; Elbaz, J.; Shabat-Simon, M.; Eilam, R.; Zangen, A.; Altstein, M.; Vogel, Z. Chronic Exposure to Delta9-Tetrahydrocannabinol Downregulates Oxytocin and Oxytocin-Associated Neurophysin in Specific Brain Areas. Mol. Cell. Neurosci. 2006, 31, 795–804. [Google Scholar] [CrossRef]
- Mennella, J.A.; Pepino, M.Y.; Teff, K.L. Acute Alcohol Consumption Disrupts the Hormonal Milieu of Lactating Women. J. Clin. Endocrinol. Metab. 2005, 90, 1979–1985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchesi, C.; Chiodera, P.; Brusamonti, E.; Volpi, R.; Coiro, V. Abnormal Plasma Oxytocin and Beta-Endorphin Levels in Alcoholics after Short and Long Term Abstinence. Prog. Neuropsychopharmacol. Biol. Psychiatry 1997, 21, 797–807. [Google Scholar] [CrossRef]
- Lee, M.R.; Schwandt, M.L.; Sankar, V.; Suchankova, P.; Sun, H.; Leggio, L. Effect of Alcohol Use Disorder on Oxytocin Peptide and Receptor MRNA Expression in Human Brain: A Post-Mortem Case-Control Study. Psychoneuroendocrinology 2017, 85, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.R.; Rohn, M.C.H.; Tanda, G.; Leggio, L. Targeting the Oxytocin System to Treat Addictive Disorders: Rationale and Progress to Date. CNS Drugs 2016, 30, 109–123. [Google Scholar] [CrossRef] [Green Version]
- Bowen, M.T.; Neumann, I.D. Rebalancing the Addicted Brain: Oxytocin Interference with the Neural Substrates of Addiction. Trends Neurosci. 2017, 40, 691–708. [Google Scholar] [CrossRef]
- Koob, G.F.; Volkow, N.D. Neurobiology of Addiction: A Neurocircuitry Analysis. Lancet Psychiatry 2016, 3, 760–773. [Google Scholar] [CrossRef]
- Dölen, G.; Darvishzadeh, A.; Huang, K.W.; Malenka, R.C. Social Reward Requires Coordinated Activity of Nucleus Accumbens Oxytocin and Serotonin. Nature 2013, 501, 179–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peris, J.; Steck, M.R.; Krause, E.G. Oxytocin Treatment for Alcoholism: Potential Neurocircuitry Targets. Neuropharmacology 2020, 171, 108091. [Google Scholar] [CrossRef] [PubMed]
- Peters, S.T.; Bowen, M.T.; Bohrer, K.; McGregor, I.S.; Neumann, I.D. Oxytocin Inhibits Ethanol Consumption and Ethanol-Induced Dopamine Release in the Nucleus Accumbens. Addict. Biol. 2017, 22, 702–711. [Google Scholar] [CrossRef] [PubMed]
- Sarnyai, Z.; Szabó, G.; Kovács, G.L.; Telegdy, G. Oxytocin Attenuates the Cocaine-Induced Exploratory Hyperactivity in Mice. Neuroreport 1990, 1, 200–202. [Google Scholar] [CrossRef] [PubMed]
- Carson, D.S.; Cornish, J.L.; Guastella, A.J.; Hunt, G.E.; McGregor, I.S. Oxytocin Decreases Methamphetamine Self-Administration, Methamphetamine Hyperactivity, and Relapse to Methamphetamine-Seeking Behaviour in Rats. Neuropharmacology 2010, 58, 38–43. [Google Scholar] [CrossRef]
- Qi, J.; Yang, J.-Y.; Song, M.; Li, Y.; Wang, F.; Wu, C.-F. Inhibition by Oxytocin of Methamphetamine-Induced Hyperactivity Related to Dopamine Turnover in the Mesolimbic Region in Mice. Naunyn Schmiedebergs Arch. Pharmacol. 2008, 376, 441–448. [Google Scholar] [CrossRef]
- Tops, M.; Koole, S.L.; IJzerman, H.; Buisman-Pijlman, F.T.A. Why Social Attachment and Oxytocin Protect against Addiction and Stress: Insights from the Dynamics between Ventral and Dorsal Corticostriatal Systems. Pharmacol. Biochem. Behav. 2014, 119, 39–48. [Google Scholar] [CrossRef]
- El Rawas, R.; Amaral, I.M.; Hofer, A. Social Interaction Reward: A Resilience Approach to Overcome Vulnerability to Drugs of Abuse. Eur. Neuropsychopharmacol. 2020, 37, 12–28. [Google Scholar] [CrossRef]
- Ballaz, S.J.; Akil, H.; Watson, S.J. Previous Experience Affects Subsequent Anxiety-like Responses in Rats Bred for Novelty Seeking. Behav. Neurosci. 2007, 121, 1113–1118. [Google Scholar] [CrossRef]
- Arenas, M.C.; Aguilar, M.A.; Montagud-Romero, S.; Mateos-García, A.; Navarro-Francés, C.I.; Miñarro, J.; Rodríguez-Arias, M. Influence of the Novelty-Seeking Endophenotype on the Rewarding Effects of Psychostimulant Drugs in Animal Models. Curr. Neuropharmacol. 2016, 14, 87–100. [Google Scholar] [CrossRef] [Green Version]
- Dellu, F.; Piazza, P.V.; Mayo, W.; Le Moal, M.; Simon, H. Novelty-Seeking in Rats—Biobehavioral Characteristics and Possible Relationship with the Sensation-Seeking Trait in Man. Neuropsychobiology 1996, 34, 136–145. [Google Scholar] [CrossRef]
- Qi, J.; Yang, J.-Y.; Wang, F.; Zhao, Y.-N.; Song, M.; Wu, C.-F. Effects of Oxytocin on Methamphetamine-Induced Conditioned Place Preference and the Possible Role of Glutamatergic Neurotransmission in the Medial Prefrontal Cortex of Mice in Reinstatement. Neuropharmacology 2009, 56, 856–865. [Google Scholar] [CrossRef]
- Bowen, M.T.; Carson, D.S.; Spiro, A.; Arnold, J.C.; McGregor, I.S. Adolescent Oxytocin Exposure Causes Persistent Reductions in Anxiety and Alcohol Consumption and Enhances Sociability in Rats. PLoS ONE 2011, 6, e27237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hicks, C.; Cornish, J.L.; Baracz, S.J.; Suraev, A.; McGregor, I.S. Adolescent Pre-Treatment with Oxytocin Protects against Adult Methamphetamine-Seeking Behavior in Female Rats. Addict. Biol. 2016, 21, 304–315. [Google Scholar] [CrossRef] [PubMed]
- McGregor, I.S.; Bowen, M.T. Breaking the Loop: Oxytocin as a Potential Treatment for Drug Addiction. Horm. Behav. 2012, 61, 331–339. [Google Scholar] [CrossRef]
- Venniro, M.; Zhang, M.; Caprioli, D.; Hoots, J.K.; Golden, S.A.; Heins, C.; Morales, M.; Epstein, D.H.; Shaham, Y. Volitional Social Interaction Prevents Drug Addiction in Rat Models. Nat. Neurosci. 2018, 21, 1520–1529. [Google Scholar] [CrossRef] [PubMed]
- El Rawas, R.; Saria, A. The Two Faces of Social Interaction Reward in Animal Models of Drug Dependence. Neurochem. Res. 2016, 41, 492–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sampedro-Piquero, P.; Ávila-Gámiz, F.; Moreno Fernández, R.D.; Castilla-Ortega, E.; Santín, L.J. The Presence of a Social Stimulus Reduces Cocaine-Seeking in a Place Preference Conditioning Paradigm. J. Psychopharmacol. 2019, 33, 1501–1511. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, C.A. Oxytocin, Tolerance, and the Dark Side of Addiction. Int. Rev. Neurobiol. 2017, 136, 239–274. [Google Scholar] [CrossRef]
- Kovács, G.L.; Horváth, Z.; Sarnyai, Z.; Faludi, M.; Telegdy, G. Oxytocin and a C-Terminal Derivative (Z-Prolyl-D-Leucine) Attenuate Tolerance to and Dependence on Morphine and Interact with Dopaminergic Neurotransmission in the Mouse Brain. Neuropharmacology 1985, 24, 413–419. [Google Scholar] [CrossRef]
- Sarnyai, Z.; Viski, S.; Kriván, M.; Szabó, G.; Kovács, G.L.; Telegdy, G. Endogenous Oxytocin Inhibits Morphine Tolerance through Limbic Forebrain Oxytocin Receptors. Brain Res. 1988, 463, 284–288. [Google Scholar] [CrossRef]
- Jodogne, C.; Tirelli, E.; Klingbiel, P.; Legros, J.J. Oxytocin Attenuates Tolerance Not Only to the Hypothermic but Also to the Myorelaxant and Akinesic Effects of Ethanol in Mice. Pharmacol. Biochem. Behav. 1991, 40, 261–265. [Google Scholar] [CrossRef]
- Sarnyai, Z.; Babarczy, E.; Kriván, M.; Szabó, G.; Kovács, G.L.; Barth, T.; Telegdy, G. Selective Attenuation of Cocaine-Induced Stereotyped Behaviour by Oxytocin: Putative Role of Basal Forebrain Target Sites. Neuropeptides 1991, 19, 51–56. [Google Scholar] [CrossRef]
- Sarnyai, Z.; Kovács, G.L. Oxytocin in Learning and Addiction: From Early Discoveries to the Present. Pharmacol. Biochem. Behav. 2014, 119, 3–9. [Google Scholar] [CrossRef]
- Szabó, G.; Kovács, G.L.; Telegdy, G. Effects of Neurohypophyseal Peptide Hormones on Alcohol Dependence and Withdrawal. Alcohol Alcohol. 1987, 22, 71–74. [Google Scholar]
- Pedersen, C.A.; Smedley, K.L.; Leserman, J.; Jarskog, L.F.; Rau, S.W.; Kampov-Polevoi, A.; Casey, R.L.; Fender, T.; Garbutt, J.C. Intranasal Oxytocin Blocks Alcohol Withdrawal in Human Subjects. Alcohol. Clin. Exp. Res. 2013, 37, 484–489. [Google Scholar] [CrossRef] [Green Version]
- Stauffer, C.S.; Musinipally, V.; Suen, A.; Lynch, K.L.; Shapiro, B.; Woolley, J.D. A Two-Week Pilot Study of Intranasal Oxytocin for Cocaine-Dependent Individuals Receiving Methadone Maintenance Treatment for Opioid Use Disorder. Addict. Res. Theory 2016, 24, 490–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, B.M.; Young, A.B.; See, R.E.; Reichel, C.M. Sex Differences in Methamphetamine Seeking in Rats: Impact of Oxytocin. Psychoneuroendocrinology 2013, 38, 2343–2353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, B.M.; Bentzley, B.S.; Regen-Tuero, H.; See, R.E.; Reichel, C.M.; Aston-Jones, G. Oxytocin Acts in Nucleus Accumbens to Attenuate Methamphetamine Seeking and Demand. Biol. Psychiatry 2017, 81, 949–958. [Google Scholar] [CrossRef]
- Miller, M.A.; Bershad, A.; King, A.C.; Lee, R.; de Wit, H. Intranasal Oxytocin Dampens Cue-Elicited Cigarette Craving in Daily Smokers: A Pilot Study. Behav. Pharmacol. 2016, 27, 697–703. [Google Scholar] [CrossRef]
- McClure, E.A.; Baker, N.L.; Gray, K.M.; Hood, C.O.; Tomko, R.L.; Carpenter, M.J.; Ramakrishnan, V.R.; Buchanan, C.J.; Saladin, M.E. The Influence of Gender and Oxytocin on Stress Reactivity, Cigarette Craving, and Smoking in a Randomized, Placebo-Controlled Laboratory Relapse Paradigm. Psychopharmacology 2020, 237, 543–555. [Google Scholar] [CrossRef]
- Van Hedger, K.; Bershad, A.K.; Lee, R.; de Wit, H. Effects of Intranasal Oxytocin on Stress-Induced Cigarette Craving in Daily Smokers. Nicotine Tob. Res. 2020, 22, 89–95. [Google Scholar] [CrossRef]
- McRae-Clark, A.L.; Baker, N.L.; Maria, M.M.-S.; Brady, K.T. Effect of Oxytocin on Craving and Stress Response in Marijuana-Dependent Individuals: A Pilot Study. Psychopharmacology 2013, 228, 623–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reed, S.C.; Haney, M.; Manubay, J.; Campagna, B.R.; Reed, B.; Foltin, R.W.; Evans, S.M. Sex Differences in Stress Reactivity after Intranasal Oxytocin in Recreational Cannabis Users. Pharmacol. Biochem. Behav. 2019, 176, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Koob, G.F.; Schulkin, J. Addiction and Stress: An Allostatic View. Neurosci. Biobehav. Rev. 2019, 106, 245–262. [Google Scholar] [CrossRef]
- Miller, A.H.; Haroon, E.; Felger, J.C. Therapeutic Implications of Brain-Immune Interactions: Treatment in Translation. Neuropsychopharmacology 2017, 42, 334–359. [Google Scholar] [CrossRef]
- Ferrer-Pérez, C.; Castro-Zavala, A.; Luján, M.Á.; Filarowska, J.; Ballestín, R.; Miñarro, J.; Valverde, O.; Rodríguez-Arias, M. Oxytocin Prevents the Increase of Cocaine-Related Responses Produced by Social Defeat. Neuropharmacology 2019, 146, 50–64. [Google Scholar] [CrossRef] [PubMed]
- Heinrichs, M.; Baumgartner, T.; Kirschbaum, C.; Ehlert, U. Social Support and Oxytocin Interact to Suppress Cortisol and Subjective Responses to Psychosocial Stress. Biol. Psychiatry 2003, 54, 1389–1398. [Google Scholar] [CrossRef]
- Reguilón, M.D.; Ferrer-Pérez, C.; Miñarro, J.; Rodríguez-Arias, M. Oxytocin Reverses Ethanol Consumption and Neuroinflammation Induced by Social Defeat in Male Mice. Horm. Behav. 2021, 127, 104875. [Google Scholar] [CrossRef] [PubMed]
- Selye, H. The evolution of the stress concept. Am. Sci. 1973, 61, 692–699. [Google Scholar] [CrossRef] [Green Version]
- Chrousos, G.P. Stress and disorders of the stress system. Nat. Rev. Endocrinol. 2009, 5, 374–381. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S.; Wingfield, J.C. What is in a name? Integrating homeostasis, allostasis and stress. Horm. Behav. 2010, 57, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Panksepp, J. Neuroscience. Feeling the pain of social loss. Science 2003, 302, 237–239. [Google Scholar] [CrossRef] [Green Version]
- Seidel, E.M.; Silani, G.; Metzler, H.; Thaler, H.; Lamm, C.; Gur, R.C.; Kryspin-Exner, I.; Habel, U.; Derntl, B. The impact of social exclusion vs. inclusion on subjective and hormonal reactions in females and males. Psychoneuroendocrinology 2013, 38, 2925–2932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tossani, E. The concept of mental pain. Psychother. Psychosom. 2013, 82, 67–73. [Google Scholar] [CrossRef]
- Bains, J.S.; Wamsteeker Cusulin, J.I.; Inoue, W. Stress-related synaptic plasticity in the hypothalamus. Nat. Rev. Neurosci 2015, 16, 377–388. [Google Scholar] [CrossRef]
- Neumann, I.D. Brain oxytocin: A key regulator of emotional and social behaviours in both females and males. J. Neuroendocrinol. 2008, 20, 858–865. [Google Scholar] [CrossRef] [PubMed]
- Groves, N.J.; Zhou, M.; Jhaveri, D.J.; McGrath, J.J.; Burne, T. Adult vitamin D deficiency exacerbates impairments caused by social stress in BALB/c and C57BL/6 mice. Psychoneuroendocrinology 2017, 86, 53–63. [Google Scholar] [CrossRef] [Green Version]
- Heshmati, M.; Christoffel, D.J.; LeClair, K.; Cathomas, F.; Golden, S.A.; Aleyasin, H.; Turecki, G.; Friedman, A.K.; Han, M.H.; Menard, C.; et al. Depression and Social Defeat Stress Are Associated with Inhibitory Synaptic Changes in the Nucleus Accumbens. J. Neurosci. 2020, 40, 6228–6233. [Google Scholar] [CrossRef]
- Koo, J.W.; Chaudhury, D.; Han, M.H.; Nestler, E.J. Role of Mesolimbic Brain-Derived Neurotrophic Factor in Depression. Biol. Psychiatry 2019, 86, 738–748. [Google Scholar] [CrossRef] [PubMed]
- Macedo, G.C.; Morita, G.M.; Domingues, L.P.; Favoretto, C.A.; Suchecki, D.; Quadros, I. Consequences of continuous social defeat stress on anxiety- and depressive-like behaviors and ethanol reward in mice. Horm. Behav. 2018, 97, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Niraula, A.; Wang, Y.; Godbout, J.P.; Sheridan, J.F. Corticosterone Production during Repeated Social Defeat Causes Monocyte Mobilization from the Bone Marrow, Glucocorticoid Resistance, and Neurovascular Adhesion Molecule Expression. J. Neurosci. 2018, 38, 2328–2340. [Google Scholar] [CrossRef]
- Rodríguez-Arias, M.; Navarrete, F.; Blanco-Gandia, M.C.; Arenas, M.C.; Bartoll-Andrés, A.; Aguilar, M.A.; Rubio, G.; Miñarro, J.; Manzanares, J. Social defeat in adolescent mice increases vulnerability to alcohol consumption. Addict. Biol. 2016, 21, 87–97. [Google Scholar] [CrossRef]
- Tas, C.; Brown, E.C.; Eskikurt, G.; Irmak, S.; Aydın, O.; Esen-Danaci, A.; Brüne, M. Cortisol response to stress in schizophrenia: Associations with oxytocin, social support and social functioning. Psychiatry Res. 2018, 270, 1047–1052. [Google Scholar] [CrossRef]
- Selten, J.P.; van der Ven, E.; Rutten, B.P.; Cantor-Graae, E. The social defeat hypothesis of schizophrenia: An update. Schizophr. Bull. 2013, 39, 1180–1186. [Google Scholar] [CrossRef] [Green Version]
- Tornatzky, W.; Miczek, K.A. Long-term impairment of autonomic circadian rhythms after brief intermittent social stress. Physiol. Behav. 1993, 53, 983–993. [Google Scholar] [CrossRef]
- Hammels, C.; Pishva, E.; De Vry, J.; van den Hove, D.L.; Prickaerts, J.; van Winkel, R.; Selten, J.P.; Lesch, K.P.; Daskalakis, N.P.; Steinbusch, H.W.; et al. Defeat stress in rodents: From behavior to molecules. Neurosci. Biobehav. Rev. 2015, 59, 111–140. [Google Scholar] [CrossRef] [PubMed]
- Neisewander, J.L.; Peartree, N.A.; Pentkowski, N.S. Emotional valence and context of social influences on drug abuse-related behavior in animal models of social stress and prosocial interaction. Psychopharmacology 2012, 224, 33–56. [Google Scholar] [CrossRef]
- Covington, H.E., 3rd; Miczek, K.A. Repeated social-defeat stress, cocaine or morphine. Effects on behavioral sensitization and intravenous cocaine self-administration “binges”. Psychopharmacology 2001, 158, 388–398. [Google Scholar] [CrossRef] [PubMed]
- Miczek, K.A.; Covington, H.E., 3rd; Nikulina, E.M., Jr.; Hammer, R.P. Aggression and defeat: Persistent effects on cocaine self-administration and gene expression in peptidergic and aminergic mesocorticolimbic circuits. Neurosci. Biobehav. Rev. 2004, 27, 787–802. [Google Scholar] [CrossRef]
- Benton, D.; Brain, P.F. Behavioral and adrenocortical reactivity in female mice following individual or group housing. Dev. Psychobiol. 1981, 14, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.J.; Grunberg, N.E. Effects of housing on male and female rats: Crowding stresses male but calm females. Physiol. Behav. 1995, 58, 1085–1089. [Google Scholar] [CrossRef]
- Sial, O.K.; Warren, B.L.; Alcantara, L.F.; Parise, E.M.; Bolaños-Guzmán, C.A. Vicarious social defeat stress: Bridging the gap between physical and emotional stress. J. Neurosci. Methods 2016, 258, 94–103. [Google Scholar] [CrossRef] [Green Version]
- Yohn, C.N.; Ashamalla, S.A.; Bokka, L.; Gergues, M.M.; Garino, A.; Samuels, B.A. Social instability is an effective chronic stress paradigm for both male and female mice. Neuropharmacology 2019, 160, 107780. [Google Scholar] [CrossRef]
- Bao, A.M.; Meynen, G.; Swaab, D.F. The stress system in depression and neurodegeneration: Focus on the human hypothalamus. Brain Res. Rev. 2008, 57, 531–553. [Google Scholar] [CrossRef]
- Leistner, C.; Menke, A. Hypothalamic-pituitary-adrenal axis and stress. Handb. Clin. Neurol. 2020, 175, 55–64. [Google Scholar] [CrossRef]
- Lupien, S.J.; Maheu, F.; Tu, M.; Fiocco, A.; Schramek, T.E. The effects of stress and stress hormones on human cognition: Implications for the field of brain and cognition. Brain Cogn. 2007, 65, 209–237. [Google Scholar] [CrossRef] [Green Version]
- Smeets, T. Autonomic and hypothalamic-pituitary-adrenal stress resilience: Impact of cardiac vagal tone. Biol. Psychol. 2010, 84, 290–295. [Google Scholar] [CrossRef]
- Bhattacharyya, S.; Ratajczak, C.K.; Vogt, S.K.; Kelley, C.; Colonna, M.; Schreiber, R.D.; Muglia, L.J. TAK1 targeting by glucocorticoids determines JNK and IkappaB regulation in Toll-like receptor-stimulated macrophages. Blood 2010, 115, 1921–1931. [Google Scholar] [CrossRef] [Green Version]
- Grad, I.; Picard, D. The glucocorticoid responses are shaped by molecular chaperones. Mol. Cell. Endocrinol. 2007, 275, 2–12. [Google Scholar] [CrossRef]
- Kagoshima, M.; Ito, K.; Cosio, B.; Adcock, I.M. Glucocorticoid suppression of nuclear factor-kappa B: A role for histone modifications. Biochem. Soc. Trans. 2003, 31, 60–65. [Google Scholar] [CrossRef] [Green Version]
- Leung, D.Y.; Bloom, J.W. Update on glucocorticoid action and resistance. J. Allergy Clin. Immunol. 2003, 111, 3–23. [Google Scholar] [CrossRef] [PubMed]
- Stahn, C.; Buttgereit, F. Genomic and nongenomic effects of glucocorticoids. Nat. Clin. Pract. Rheumatol. 2008, 4, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Busillo, J.M.; Cidlowski, J.A. The five Rs of glucocorticoid action during inflammation: Ready, reinforce, repress, resolve, and restore. Trends Endocrinol. Metab. 2013, 24, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Webster, J.C.; Oakley, R.H.; Jewell, C.M.; Cidlowski, J.A. Proinflammatory cytokines regulate human glucocorticoid receptor gene expression and lead to the accumulation of the dominant negative beta isoform: A mechanism for the generation of glucocorticoid resistance. Proc. Natl. Acad. Sci. USA 2001, 98, 6865–6870. [Google Scholar] [CrossRef] [Green Version]
- Jeanneteau, F.D.; Lambert, W.M.; Ismaili, N.; Bath, K.G.; Lee, F.S.; Garabedian, M.J.; Chao, M.V. BDNF and glucocorticoids regulate corticotrophin-releasing hormone (CRH) homeostasis in the hypothalamus. Proc. Natl. Acad. Sci. USA 2012, 109, 1305–1310. [Google Scholar] [CrossRef] [Green Version]
- Logrip, M.L.; Koob, G.F.; Zorrilla, E.P. Role of corticotropin-releasing factor in drug addiction: Potential for pharmacological intervention. CNS Drugs 2011, 25, 271–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francis, T.C.; Chandra, R.; Friend, D.M.; Finkel, E.; Dayrit, G.; Miranda, J.; Brooks, J.M.; Iñiguez, S.D.; O’Donnell, P.; Kravitz, A.; et al. Nucleus accumbens medium spiny neuron subtypes mediate depression-related outcomes to social defeat stress. Biol. Psychiatry 2015, 77, 212–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heshmati, M.; Russo, S.J. Anhedonia and the brain reward circuitry in depression. Curr. Behav. Neurosci. Rep. 2015, 2, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Finnell, J.E.; Moffitt, C.M.; Hesser, L.A.; Harrington, E.; Melson, M.N.; Wood, C.S.; Wood, S.K. The contribution of the locus coeruleus-norepinephrine system in the emergence of defeat-induced inflammatory priming. Brain Behav. Immun. 2019, 79, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Haenisch, B.; Bilkei-Gorzo, A.; Caron, M.G.; Bönisch, H. Knockout of the norepinephrine transporter and pharmacologically diverse antidepressants prevent behavioral and brain neurotrophin alterations in two chronic stress models of depression. J. Neurochem. 2009, 111, 403–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasegawa, S.; Miyake, Y.; Yoshimi, A.; Mouri, A.; Hida, H.; Yamada, K.; Ozaki, N.; Nabeshima, T.; Noda, Y. Dysfunction of Serotonergic and Dopaminergic Neuronal Systems in the Antidepressant-Resistant Impairment of Social Behaviors Induced by Social Defeat Stress Exposure as Juveniles. Int. J. Neuropsychopharmacol. 2018, 21, 837–846. [Google Scholar] [CrossRef]
- Ramírez, L.A.; Pérez-Padilla, E.A.; García-Oscos, F.; Salgado, H.; Atzori, M.; Pineda, J.C. A new theory of depression based on the serotonin/kynurenine relationship and the hypothalamicpituitary-adrenal axis. Biomedica 2018, 38, 437–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, M.A.; Grober, M.S.; Nicholas, C.R.; Huhman, K.L. Aggressive encounters alter the activation of serotonergic neurons and the expression of 5-HT1A mRNA in the hamster dorsal raphe nucleus. Neuroscience 2009, 161, 680–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Fan, Y.; Li, Y.; Zhu, H.; Wang, L.; Zhu, M.Y. Chronic social defeat up-regulates expression of the serotonin transporter in rat dorsal raphe nucleus and projection regions in a glucocorticoid-dependent manner. J. Neurochem. 2012, 123, 1054–1068. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, M.; Yoshida, M.; Emoto, H.; Ishii, H. Noradrenaline systems in the hypothalamus, amygdala and locus coeruleus are involved in the provocation of anxiety: Basic studies. Eur. J. Pharmacol. 2000, 405, 397–406. [Google Scholar] [CrossRef]
- Cohen, S.; Janicki-Deverts, D.; Doyle, W.J.; Miller, G.E.; Frank, E.; Rabin, B.S.; Turner, R.B. Chronic stress, glucocorticoid receptor resistance, inflammation, and disease risk. Proc. Natl. Acad. Sci. USA 2012, 109, 5995–5999. [Google Scholar] [CrossRef] [Green Version]
- Horowitz, M.A.; Cattaneo, A.; Cattane, N.; Lopizzo, N.; Tojo, L.; Bakunina, N.; Musaelyan, K.; Borsini, A.; Zunszain, P.A.; Pariante, C.M. Glucocorticoids prime the inflammatory response of human hippocampal cells through up-regulation of inflammatory pathways. Brain Behav. Immun. 2020, 87, 777–794. [Google Scholar] [CrossRef]
- Johnson, J.D.; Barnard, D.F.; Kulp, A.C.; Mehta, D.M. Neuroendocrine Regulation of Brain Cytokines After Psychological Stress. J. Endocr. Soc. 2019, 3, 1302–1320. [Google Scholar] [CrossRef]
- Takahashi, A.; Flanigan, M.E.; McEwen, B.S.; Russo, S.J. Aggression, Social Stress, and the Immune System in Humans and Animal Models. Front. Behav. Neurosci. 2018, 12, 56. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.J.; Ni, H.Y.; Chen, R.; Chang, L.; Shi, H.J.; Qiu, D.; Zhang, Z.; Wu, D.L.; Jiang, Z.C.; Xin, H.L.; et al. Hippocampal nuclear factor kappa B accounts for stress-induced anxiety behaviors via enhancing neuronal nitric oxide synthase (nNOS)-carboxy-terminal PDZ ligand of nNOS-Dexras1 coupling. J. Neurochem. 2018, 146, 598–612. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.C.; Lin, C.C.; Tzeng, N.S.; Tung, C.S.; Liu, Y.P. Effects of oxytocin on prosocial behavior and the associated profiles of oxytocinergic and corticotropin-releasing hormone receptors in a rodent model of posttraumatic stress disorder. J. Biomed. Sci. 2019, 26, 26. [Google Scholar] [CrossRef] [Green Version]
- Winter, J.; Jurek, B. The interplay between oxytocin and the CRF system: Regulation of the stress response. Cell Tissue Res. 2019, 375, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Jurek, B.; Slattery, D.A.; Hiraoka, Y.; Liu, Y.; Nishimori, K.; Aguilera, G.; Neumann, I.D.; van den Burg, E.H. Oxytocin Regulates Stress-Induced Crf Gene Transcription through CREB-Regulated Transcription Coactivator 3. J. Neurosci. 2015, 35, 12248–12260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gobrogge, K.; Wang, Z. Neuropeptidergic regulation of pair-bonding and stress buffering: Lessons from voles. Horm. Behav. 2015, 76, 91–105. [Google Scholar] [CrossRef] [Green Version]
- Gunnar, M.R.; Hostinar, C.E. The social buffering of the hypothalamic-pituitary-adrenocortical axis in humans: Developmental and experiential determinants. Soc. Neurosci. 2015, 10, 479–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heck, A.L.; Sheng, J.A.; Miller, A.M.; Stover, S.A.; Bales, N.J.; Tan, S.; Daniels, R.M.; Fleury, T.K.; Handa, R.J. Social isolation alters hypothalamic pituitary adrenal axis activity after chronic variable stress in male C57BL/6 mice. Stress 2020, 23, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Engelmann, M.; Landgraf, R.; Wotjak, C.T. The hypothalamic-neurohypophysial system regulates the hypothalamic-pituitary-adrenal axis under stress: An old concept revisited. Front. Neuroendocrinol. 2004, 25, 132–149. [Google Scholar] [CrossRef]
- Neumann, I.D.; Landgraf, R. Balance of brain oxytocin and vasopressin: Implications for anxiety, depression, and social behaviors. Trends Neurosci. 2012, 35, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Quirin, M.; Kuhl, J.; Düsing, R. Oxytocin buffers cortisol responses to stress in individuals with impaired emotion regulation abilities. Psychoneuroendocrinology 2011, 36, 898–904. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.S.; Wang, Z. Hypothalamic oxytocin mediates social buffering of the stress response. Biol. Psychiatry 2014, 76, 281–288. [Google Scholar] [CrossRef] [Green Version]
- Engert, V.; Koester, A.M.; Riepenhausen, A.; Singer, T. Boosting recovery rather than buffering reactivity: Higher stress-induced oxytocin secretion is associated with increased cortisol reactivity and faster vagal recovery after acute psychosocial stress. Psychoneuroendocrinology 2016, 74, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.A.; Cardoso, C.; Ellenbogen, M.A. A meta-analytic review of the correlation between peripheral oxytocin and cortisol concentrations. Front. Neuroendocrinol. 2016, 43, 19–27. [Google Scholar] [CrossRef]
- Jong, T.R.; Menon, R.; Bludau, A.; Grund, T.; Biermeier, V.; Klampfl, S.M.; Jurek, B.; Bosch, O.J.; Hellhammer, J.; Neumann, I.D. Salivary oxytocin concentrations in response to running, sexual self-stimulation, breastfeeding and the TSST: The Regensburg Oxytocin Challenge (ROC) study. Psychoneuroendocrinology 2015, 62, 381–388. [Google Scholar] [CrossRef]
- Litvin, Y.; Murakami, G.; Pfaff, D.W. Effects of chronic social defeat on behavioral and neural correlates of sociality: Vasopressin, oxytocin and the vasopressinergic V1b receptor. Physiol Behav. 2011, 103, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Nasanbuyan, N.; Yoshida, M.; Takayanagi, Y.; Inutsuka, A.; Nishimori, K.; Yamanaka, A.; Onaka, T. Oxytocin-Oxytocin Receptor Systems Facilitate Social Defeat Posture in Male Mice. Endocrinology 2018, 159, 763–775. [Google Scholar] [CrossRef]
- Steinman, M.Q.; Duque-Wilckens, N.; Greenberg, G.D.; Hao, R.; Campi, K.L.; Laredo, S.A.; Laman-Maharg, A.; Manning, C.E.; Doig, I.E.; Lopez, E.M.; et al. Sex-Specific Effects of Stress on Oxytocin Neurons Correspond With Responses to Intranasal Oxytocin. Biol. Psychiatry 2016, 80, 406–414. [Google Scholar] [CrossRef] [Green Version]
- Steinman, M.Q.; Laredo, S.A.; Lopez, E.M.; Manning, C.E.; Hao, R.C.; Doig, I.E.; Campi, K.L.; Flowers, A.E.; Knight, J.K.; Trainor, B.C. Hypothalamic vasopressin systems are more sensitive to the long term effects of social defeat in males versus females. Psychoneuroendocrinology 2015, 51, 122–134. [Google Scholar] [CrossRef] [Green Version]
- Duque-Wilckens, N.; Steinman, M.Q.; Busnelli, M.; Chini, B.; Yokoyama, S.; Pham, M.; Laredo, S.A.; Hao, R.; Perkeybile, A.M.; Minie, V.A.; et al. Oxytocin Receptors in the Anteromedial Bed Nucleus of the Stria Terminalis Promote Stress-Induced Social Avoidance in Female California Mice. Biol. Psychiatry 2018, 83, 203–213. [Google Scholar] [CrossRef]
- Nowacka-Chmielewska, M.M.; Kasprowska-Liśkiewicz, D.; Barski, J.J.; Obuchowicz, E.; Małecki, A. The behavioral and molecular evaluation of effects of social instability stress as a model of stress-related disorders in adult female rats. Stress 2017, 20, 549–561. [Google Scholar] [CrossRef]
- Hou, W.; He, Z.; Yang, Y.; Yuan, W.; Wang, L.; Zhang, J.; Zhang, X.; Cai, W.; Guo, Q.; Tai, F. The involvement of oxytocin in the effects of chronic social defeat stress on emotional behaviours in adult female mandarin voles. Eur. J. Neurosci. 2020, 52, 2853–2872. [Google Scholar] [CrossRef]
- Yuen, K.W.; Garner, J.P.; Carson, D.S.; Keller, J.; Lembke, A.; Hyde, S.A.; Kenna, H.A.; Tennakoon, L.; Schatzberg, A.F.; Parker, K.J. Plasma oxytocin concentrations are lower in depressed vs. healthy control women and are independent of cortisol. J. Psychiatr. Res. 2014, 51, 30–36. [Google Scholar] [CrossRef]
- Jobst, A.; Sabass, L.; Palagyi, A.; Bauriedl-Schmidt, C.; Mauer, M.C.; Sarubin, N.; Buchheim, A.; Renneberg, B.; Falkai, P.; Zill, P.; et al. Effects of social exclusion on emotions and oxytocin and cortisol levels in patients with chronic depression. J. Psychiatr. Res. 2015, 60, 170–177. [Google Scholar] [CrossRef]
- Eckstein, M.; Scheele, D.; Weber, K.; Stoffel-Wagner, B.; Maier, W.; Hurlemann, R. Oxytocin facilitates the sensation of social stress. Hum. Brain Mapp. 2014, 35, 4741–4750. [Google Scholar] [CrossRef]
- Yoon, S.; Kim, Y.K. The Role of the Oxytocin System in Anxiety Disorders. Adv. Exp. Med. Biol. 2020, 1191, 103–120. [Google Scholar] [CrossRef] [PubMed]
- Hoge, E.A.; Lawson, E.A.; Metcalf, C.A.; Keshaviah, A.; Zak, P.J.; Pollack, M.H.; Simon, N.M. Plasma oxytocin immunoreactive products and response to trust in patients with social anxiety disorder. Depress. Anxiety 2012, 29, 924–930. [Google Scholar] [CrossRef] [PubMed]
- Dodhia, S.; Hosanagar, A.; Fitzgerald, D.A.; Labuschagne, I.; Wood, A.G.; Nathan, P.J.; Phan, K.L. Modulation of resting-state amygdala-frontal functional connectivity by oxytocin in generalized social anxiety disorder. Neuropsychopharmacology 2014, 39, 2061–2069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labuschagne, I.; Phan, K.L.; Wood, A.; Angstadt, M.; Chua, P.; Heinrichs, M.; Stout, J.C.; Nathan, P.J. Oxytocin attenuates amygdala reactivity to fear in generalized social anxiety disorder. Neuropsychopharmacology 2010, 35, 2403–2413. [Google Scholar] [CrossRef] [Green Version]
- Labuschagne, I.; Phan, K.L.; Wood, A.; Angstadt, M.; Chua, P.; Heinrichs, M.; Stout, J.C.; Nathan, P.J. Medial frontal hyperactivity to sad faces in generalized social anxiety disorder and modulation by oxytocin. Int. J. Neuropsychopharmacol. 2012, 15, 883–896. [Google Scholar] [CrossRef] [Green Version]
- Luo, L.; Becker, B.; Geng, Y.; Zhao, Z.; Gao, S.; Zhao, W.; Yao, S.; Zheng, X.; Ma, X.; Gao, Z.; et al. Sex-dependent neural effect of oxytocin during subliminal processing of negative emotion faces. Neuroimage 2017, 162, 127–137. [Google Scholar] [CrossRef]
- Fang, A.; Treadway, M.T.; Hofmann, S.G. Working hard for oneself or others: Effects of oxytocin on reward motivation in social anxiety disorder. Biol. Psychol. 2017, 127, 157–162. [Google Scholar] [CrossRef]
- Fan, Y.; Herrera-Melendez, A.L.; Pestke, K.; Feeser, M.; Aust, S.; Otte, C.; Pruessner, J.C.; Böker, H.; Bajbouj, M.; Grimm, S. Early life stress modulates amygdala-prefrontal functional connectivity: Implications for oxytocin effects. Hum. Brain Mapp. 2014, 35, 5328–5339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feeser, M.; Fan, Y.; Weigand, A.; Hahn, A.; Gärtner, M.; Aust, S.; Böker, H.; Bajbouj, M.; Grimm, S. The beneficial effect of oxytocin on avoidance-related facial emotion recognition depends on early life stress experience. Psychopharmacology 2014, 231, 4735–4744. [Google Scholar] [CrossRef] [PubMed]
- Schwaiger, M.; Heinrichs, M.; Kumsta, R. Oxytocin administration and emotion recognition abilities in adults with a history of childhood adversity. Psychoneuroendocrinology 2019, 99, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Nawijn, L.; van Zuiden, M.; Koch, S.B.; Frijling, J.L.; Veltman, D.J.; Olff, M. Intranasal oxytocin enhances neural processing of monetary reward and loss in post-traumatic stress disorder and traumatized controls. Psychoneuroendocrinology 2016, 66, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Nawijn, L.; van Zuiden, M.; Koch, S.B.; Frijling, J.L.; Veltman, D.J.; Olff, M. Intranasal oxytocin increases neural responses to social reward in post-traumatic stress disorder. Soc. Cogn. Affect. Neurosci. 2017, 12, 212–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, S.B.; van Zuiden, M.; Nawijn, L.; Frijling, J.L.; Veltman, D.J.; Olff, M. Intranasal Oxytocin Normalizes Amygdala Functional Connectivity in Posttraumatic Stress Disorder. Neuropsychopharmacology 2016, 41, 2041–2051. [Google Scholar] [CrossRef] [Green Version]
- Koch, S.; van Zuiden, M.; Nawijn, L.; Frijling, J.L.; Veltman, D.J.; Olff, M. Effects of intranasal oxytocin on distraction as emotion regulation strategy in patients with post-traumatic stress disorder. Eur. Neuropsychopharmacol. 2019, 29, 266–277. [Google Scholar] [CrossRef]
- Wang, L.; Hou, W.; He, Z.; Yuan, W.; Yang, J.; Yang, Y.; Jia, R.; Zhu, Z.; Zhou, Y.; Tai, F. Effects of chronic social defeat on social behaviors in adult female mandarin voles (Microtus mandarinus): Involvement of the oxytocin system in the nucleus accumbens. Prog. Neuropsychopharmacol. Biol. Psychiatry 2018, 82, 278–288. [Google Scholar] [CrossRef]
- Li, Q.; Zhang, B.; Cao, H.; Liu, W.; Guo, F.; Shen, F.; Ye, B.; Liu, H.; Li, Y.; Liu, Z. Oxytocin Exerts Antidepressant-like effect by potentiating dopaminergic synaptic transmission in the mPFC. Neuropharmacology 2020, 162, 107836. [Google Scholar] [CrossRef]
- Arena, D.T.; Covington, H.E., 3rd; DeBold, J.F.; Miczek, K.A. Persistent increase of I.V. cocaine self-administration in a subgroup of C57BL/6J male mice after social defeat stress. Psychopharmacology 2019, 236, 2027–2037. [Google Scholar] [CrossRef] [PubMed]
- Boyson, C.O.; Holly, E.N.; Shimamoto, A.; Albrechet-Souza, L.; Weiner, L.A.; DeBold, J.F.; Miczek, K.A. Social stress and CRF-dopamine interactions in the VTA: Role in long-term escalation of cocaine self-administration. J. Neurosci. 2014, 34, 6659–6667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, X.; DeBold, J.F.; Miczek, K.A. Prevention and reversal of social stress-escalated cocaine self-administration in mice by intra-VTA CRFR1 antagonism. Psychopharmacology 2017, 234, 2813–2821. [Google Scholar] [CrossRef] [PubMed]
- Leonard, M.Z.; DeBold, J.F.; Miczek, K.A. Escalated cocaine “binges” in rats: Enduring effects of social defeat stress or intra-VTA CRF. Psychopharmacology 2017, 234, 2823–2836. [Google Scholar] [CrossRef] [PubMed]
- Montagud-Romero, S.; Reguilón, M.D.; Pascual, M.; Blanco-Gandía, M.C.; Guerri, C.; Miñarro, J.; Rodríguez-Arias, M. Critical role of TLR4 in uncovering the increased rewarding effects of cocaine and ethanol induced by social defeat in male mice. Neuropharmacology 2021, 182, 108368. [Google Scholar] [CrossRef]
- Reguilón, M.D.; Ferrer-Pérez, C.; Ballestín, R.; Miñarro, J.; Rodríguez-Arias, M. Voluntary wheel running protects against the increase in ethanol consumption induced by social stress in mice. Drug Alcohol Depend. 2020, 212, 108004. [Google Scholar] [CrossRef]
- Ballestín, R.; Alegre-Zurano, L.; Ferrer-Pérez, C.; Cantacorps, L.; Miñarro, J.; Valverde, O.; Rodríguez-Arias, M. Neuroinflammatory and behavioral susceptibility profile of mice exposed to social stress towards cocaine effects. Prog. Neuropsychopharmacol. Biol. Psychiatry 2021, 105, 110123. [Google Scholar] [CrossRef]
- Ferrer-Pérez, C.; Reguilón, M.D.; Manzanedo, C.; Aguilar, M.A.; Miñarro, J.; Rodríguez-Arias, M. Antagonism of corticotropin-releasing factor CRF1 receptors blocks the enhanced response to cocaine after social stress. Eur. J. Pharmacol. 2018, 823, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Montagud-Romero, S.; Nuñez, C.; Blanco-Gandia, M.C.; Martínez-Laorden, E.; Aguilar, M.A.; Navarro-Zaragoza, J.; Almela, P.; Milanés, M.V.; Laorden, M.L.; Miñarro, J.; et al. Repeated social defeat and the rewarding effects of cocaine in adult and adolescent mice: Dopamine transcription factors, proBDNF signaling pathways, and the TrkB receptor in the mesolimbic system. Psychopharmacology 2017, 234, 2063–2075. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, M.A.; Rodríguez-Arias, M.; Miñarro, J. Neurobiological mechanisms of the reinstatement of drug-conditioned place preference. Brain Res. Rev. 2009, 59, 253–277. [Google Scholar] [CrossRef] [PubMed]
- Tzschentke, T.M. Measuring reward with the conditioned place preference (CPP) paradigm: Update of the last decade. Addict. Biol. 2007, 12, 227–462. [Google Scholar] [CrossRef] [PubMed]
- Miczek, K.A.; Yap, J.J.; Covington, H.E., 3rd. Social stress, therapeutics and drug abuse: Preclinical models of escalated and depressed intake. Pharmacol. Ther. 2008, 120, 102–128. [Google Scholar] [CrossRef] [Green Version]
- Vasconcelos, M.; Stein, D.J.; de Almeida, R.M. Social defeat protocol and relevant biomarkers, implications for stress response physiology, drug abuse, mood disorders and individual stress vulnerability: A systematic review of the last decade. Trends Psychiatry Psychother. 2015, 37, 51–66. [Google Scholar] [CrossRef] [Green Version]
- Holly, E.N.; DeBold, J.F.; Miczek, K.A. Increased mesocorticolimbic dopamine during acute and repeated social defeat stress: Modulation by corticotropin releasing factor receptors in the ventral tegmental area. Psychopharmacology 2015, 232, 4469–4479. [Google Scholar] [CrossRef]
- Hwa, L.S.; Holly, E.N.; DeBold, J.F.; Miczek, K.A. Social stress-escalated intermittent alcohol drinking: Modulation by CRF-R1 in the ventral tegmental area and accumbal dopamine in mice. Psychopharmacology 2016, 233, 681–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yap, J.J.; Chartoff, E.H.; Holly, E.N.; Potter, D.N.; Carlezon, W.A., Jr.; Miczek, K.A. Social defeat stress-induced sensitization and escalated cocaine self-administration: The role of ERK signaling in the rat ventral tegmental area. Psychopharmacology 2015, 232, 1555–1569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsushita, H.; Latt, H.M.; Koga, Y.; Nishiki, T.; Matsui, H. Oxytocin and Stress: Neural Mechanisms, Stress-Related Disorders, and Therapeutic Approaches. Neuroscience 2019, 417, 1–10. [Google Scholar] [CrossRef]
- Lee, H.; Jang, M.; Noh, J. Oxytocin attenuates aversive response to nicotine and anxiety-like behavior in adolescent rats. Neurosci. Res. 2017, 115, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Hung, L.W.; Neuner, S.; Polepalli, J.S.; Beier, K.T.; Wright, M.; Walsh, J.J.; Lewis, E.M.; Luo, L.; Deisseroth, K.; Dölen, G.; et al. Gating of social reward by oxytocin in the ventral tegmental area. Science 2017, 357, 1406–1411. [Google Scholar] [CrossRef] [Green Version]
- Peris, J.; MacFadyen, K.; Smith, J.A.; de Kloet, A.D.; Wang, L.; Krause, E.G. Oxytocin receptors are expressed on dopamine and glutamate neurons in the mouse ventral tegmental area that project to nucleus accumbens and other mesolimbic targets. J. Comp. Neurol. 2017, 525, 1094–1108. [Google Scholar] [CrossRef]
- Xiao, L.; Priest, M.F.; Nasenbeny, J.; Lu, T.; Kozorovitskiy, Y. Biased Oxytocinergic Modulation of Midbrain Dopamine Systems. Neuron 2017, 95, 368–384.e5. [Google Scholar] [CrossRef] [Green Version]
- Ferrer-Pérez, C.; Reguilón, M.D.; Manzanedo, C.; Miñarro, J.; Rodríguez-Arias, M. Social Housing Conditions Modulate the Long-Lasting Increase in Cocaine Reward Induced by Intermittent Social Defeat. Front. Behav. Neurosci. 2019, 13, 148. [Google Scholar] [CrossRef] [Green Version]
- Ferrer-Pérez, C.; Reguilón, M.D.; Miñarro, J.; Rodríguez-Arias, M. Endogenous oxytocin is essential for the buffering effects of pair housing against the increase in cocaine reward induced by social stress. Physiol. Behav. 2020, 221, 112913. [Google Scholar] [CrossRef] [PubMed]
- King, C.E.; Becker, H.C. Oxytocin attenuates stress-induced reinstatement of alcohol seeking behavior in male and female mice. Psychopharmacology 2019, 236, 2613–2622. [Google Scholar] [CrossRef] [PubMed]
- Ferland, C.L.; Reichel, C.M.; McGinty, J.F. Effects of oxytocin on methamphetamine-seeking exacerbated by predator odor pre-exposure in rats. Psychopharmacology 2016, 233, 1015–1024. [Google Scholar] [CrossRef]
- Flanagan, J.C.; Baker, N.L.; McRae-Clark, A.L.; Brady, K.T.; Moran-Santa Maria, M.M. Effects of adverse childhood experiences on the association between intranasal oxytocin and social stress reactivity among individuals with cocaine dependence. Psychiatry Res. 2015, 229, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Flanagan, J.C.; Allan, N.P.; Calhoun, C.D.; Badour, C.L.; Moran-Santa Maria, M.; Brady, K.T.; Back, S.E. Effects of oxytocin on stress reactivity and craving in veterans with co-occurring PTSD and alcohol use disorder. Exp. Clin. Psychopharmacol. 2019, 27, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Hood, C.O.; Tomko, R.L.; Baker, N.L.; Tuck, B.M.; Flanagan, J.C.; Carpenter, M.J.; Gray, K.M.; Saladin, M.E.; McClure, E.A. Examining sex, adverse childhood experiences, and oxytocin on neuroendocrine reactivity in smokers. Psychoneuroendocrinology 2020, 120, 104752. [Google Scholar] [CrossRef]
- Calcia, M.A.; Bonsall, D.R.; Bloomfield, P.S.; Selvaraj, S.; Barichello, T.; Howes, O.D. Stress and neuroinflammation: A systematic review of the effects of stress on microglia and the implications for mental illness. Psychopharmacology 2016, 233, 1637–1650. [Google Scholar] [CrossRef] [Green Version]
- Montagud-Romero, S.; Blanco-Gandía, M.C.; Reguilón, M.D.; Ferrer-Pérez, C.; Ballestín, R.; Miñarro, J.; Rodríguez-Arias, M. Social defeat stress: Mechanisms underlying the increase in rewarding effects of drugs of abuse. Eur. J. Neurosci. 2018, 48, 2948–2970. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Arias, M.; Montagud-Romero, S.; Rubio-Araiz, A.; Aguilar, M.A.; Martín-García, E.; Cabrera, R.; Maldonado, R.; Porcu, F.; Colado, M.I.; Miñarro, J. Effects of repeated social defeat on adolescent mice on cocaine-induced CPP and self-administration in adulthood: Integrity of the blood-brain barrier. Addict. Biol. 2017, 22, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Weber, M.D.; Godbout, J.P.; Sheridan, J.F. Repeated Social Defeat, Neuroinflammation, and Behavior: Monocytes Carry the Signal. Neuropsychopharmacology 2017, 42, 46–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stankiewicz, A.M.; Goscik, J.; Majewska, A.; Swiergiel, A.H.; Juszczak, G.R. The Effect of Acute and Chronic Social Stress on the Hippocampal Transcriptome in Mice. PLoS ONE 2015, 10, e0142195. [Google Scholar] [CrossRef]
- Ferrer-Pérez, C.; Escrivá-Martinez, T.; Montagud-Romero, S.; Ballestín, R.; Reguilón, M.D.; Miñarro, J.; Rodríguez-Arias, M. Indomethacin blocks the increased conditioned rewarding effects of cocaine induced by repeated social defeat. PLoS ONE 2019, 14, e0212397. [Google Scholar] [CrossRef] [Green Version]
- Akman, T.; Akman, L.; Erbas, O.; Terek, M.C.; Taskiran, D.; Ozsaran, A. The preventive effect of oxytocin to Cisplatin-induced neurotoxicity: An experimental rat model. Biomed. Res. Int. 2015, 167235. [Google Scholar] [CrossRef]
- Karelina, K.; Stuller, K.A.; Jarrett, B.; Zhang, N.; Wells, J.; Norman, G.J.; DeVries, A.C. Oxytocin mediates social neuroprotection after cerebral ischemia. Stroke 2011, 42, 3606–3611. [Google Scholar] [CrossRef] [Green Version]
- Yuan, L.; Liu, S.; Bai, X.; Gao, Y.; Liu, G.; Wang, X.; Liu, D.; Li, T.; Hao, A.; Wang, Z. Oxytocin inhibits lipopolysaccharide-induced inflammation in microglial cells and attenuates microglial activation in lipopolysaccharide-treated mice. J. Neuroinflamm. 2016, 13, 77. [Google Scholar] [CrossRef] [Green Version]
- Amini-Khoei, H.; Mohammadi-Asl, A.; Amiri, S.; Hosseini, M.J.; Momeny, M.; Hassanipour, M.; Rastegar, M.; Haj-Mirzaian, A.; Mirzaian, A.H.; Sanjarimoghaddam, H.; et al. Oxytocin mitigated the depressive-like behaviors of maternal separation stress through modulating mitochondrial function and neuroinflammation. Prog. Neuropsychopharmacol. Biol. Psychiatry 2017, 76, 169–178. [Google Scholar] [CrossRef]
- Wang, S.C.; Lin, C.C.; Chen, C.C.; Tzeng, N.S.; Liu, Y.P. Effects of Oxytocin on Fear Memory and Neuroinflammation in a Rodent Model of Posttraumatic Stress Disorder. Int. J. Mol. Sci. 2018, 19, 3848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baracz, S.J.; Everett, N.A.; Cornish, J.L. The impact of early life stress on the central oxytocin system and susceptibility for drug addiction: Applicability of oxytocin as a pharmacotherapy. Neurosci. Biobehav. Rev. 2020, 110, 114–132. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Species | Stress Type | Paradigm/Drug Disorder | Treatment/Dosage | Finding | Reference |
|---|---|---|---|---|---|
| Male mice | Social Defeat | Cocaine CPP | Oxytocin 1 mg/kg; i.p. | Blocked the long-term increase in the conditioned rewarding effects, favored extinction and decreased reinstatment | 59 |
| Cocaine SA | |||||
| Male mice | Social Defeat | Ethanol SA | Oxytocin 1 mg/kg; i.p. | Blocked the long-term increase ethanol consumption and seeking behavior | 61 |
| Male and female mice | Predator odor | Ethanol SA | Oxytocin 1 mg/kg; i.p. | Attenuated alcohol seeking behavior | 176 |
| Male mice | Restraint | Methamphetamine CPP | Oxytocin 0.1, 0.5, 2.5 μg/μl; i.c.v. | Attenuated reinstatment | 34 |
| Male mice | Forced swimming stress | Morphine CPP | Carbetocin * 6.4 mg/kg; i.p. | Attenuatted reinstatment | 14 |
| Male rats | Predator odor | Methamphetamine CPP | Oxytocin 1 mg/kg; i.p. | Decreased CPP acquisition | 177 |
| Male and female rats | Yohimbine | Methamphetamine CPP | Oxytocin 1 mg/kg; i.p. | Decreased CPP acquisition | 50 |
| Male and female humans | Mistreatment during childhood | Cocaine-dependent | Oxytocin 40 IU; intransal | Reduced risk of relapse | 178 |
| Male military veterans humans | Post-traumatic stress disorder | Alcohol use disorder | Oxytocin 40 IU; intransal | Attenuated reactivity of cortisol to stress task, but did not reduce craving | 179 |
| Male and female humans | Adverse experiences in childhood | Tobacco smokers | Oxytocin 40 IU; intransal | Attenuated cortisol response in trier social stress task. Greater anxiolytic effect in men | 180 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferrer-Pérez, C.; Reguilón, M.D.; Miñarro, J.; Rodríguez-Arias, M. Oxytocin Signaling as a Target to Block Social Defeat-Induced Increases in Drug Abuse Reward. Int. J. Mol. Sci. 2021, 22, 2372. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052372
Ferrer-Pérez C, Reguilón MD, Miñarro J, Rodríguez-Arias M. Oxytocin Signaling as a Target to Block Social Defeat-Induced Increases in Drug Abuse Reward. International Journal of Molecular Sciences. 2021; 22(5):2372. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052372
Chicago/Turabian StyleFerrer-Pérez, Carmen, Marina D. Reguilón, José Miñarro, and Marta Rodríguez-Arias. 2021. "Oxytocin Signaling as a Target to Block Social Defeat-Induced Increases in Drug Abuse Reward" International Journal of Molecular Sciences 22, no. 5: 2372. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052372