
Salivary Outer Membrane Vesicles and DNA Methylation of Small Extracellular Vesicles as Biomarkers for Periodontal Status: A Pilot Study


Abstract
:1. Introduction
2. Results
2.1. Participant Characteristics
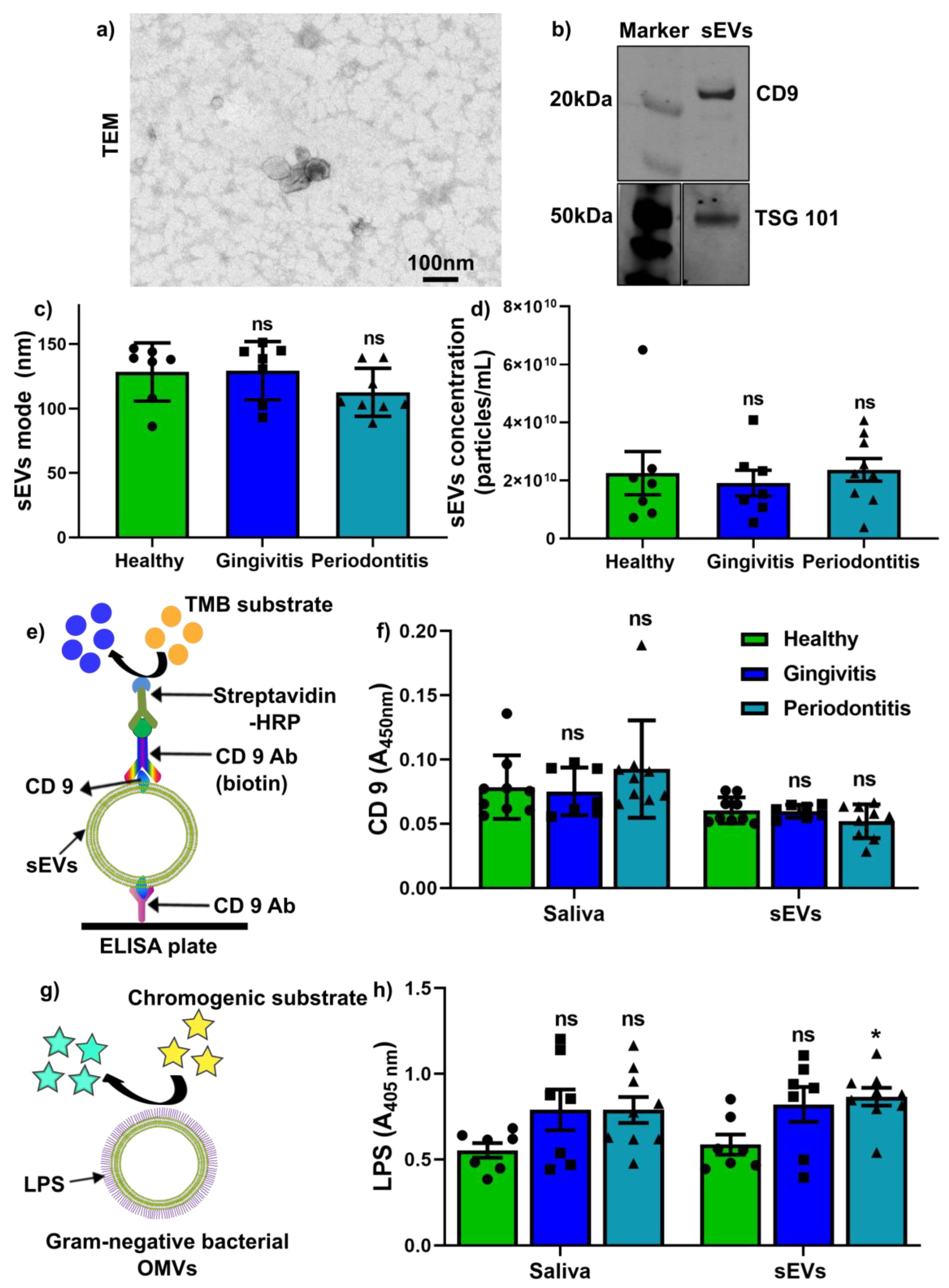
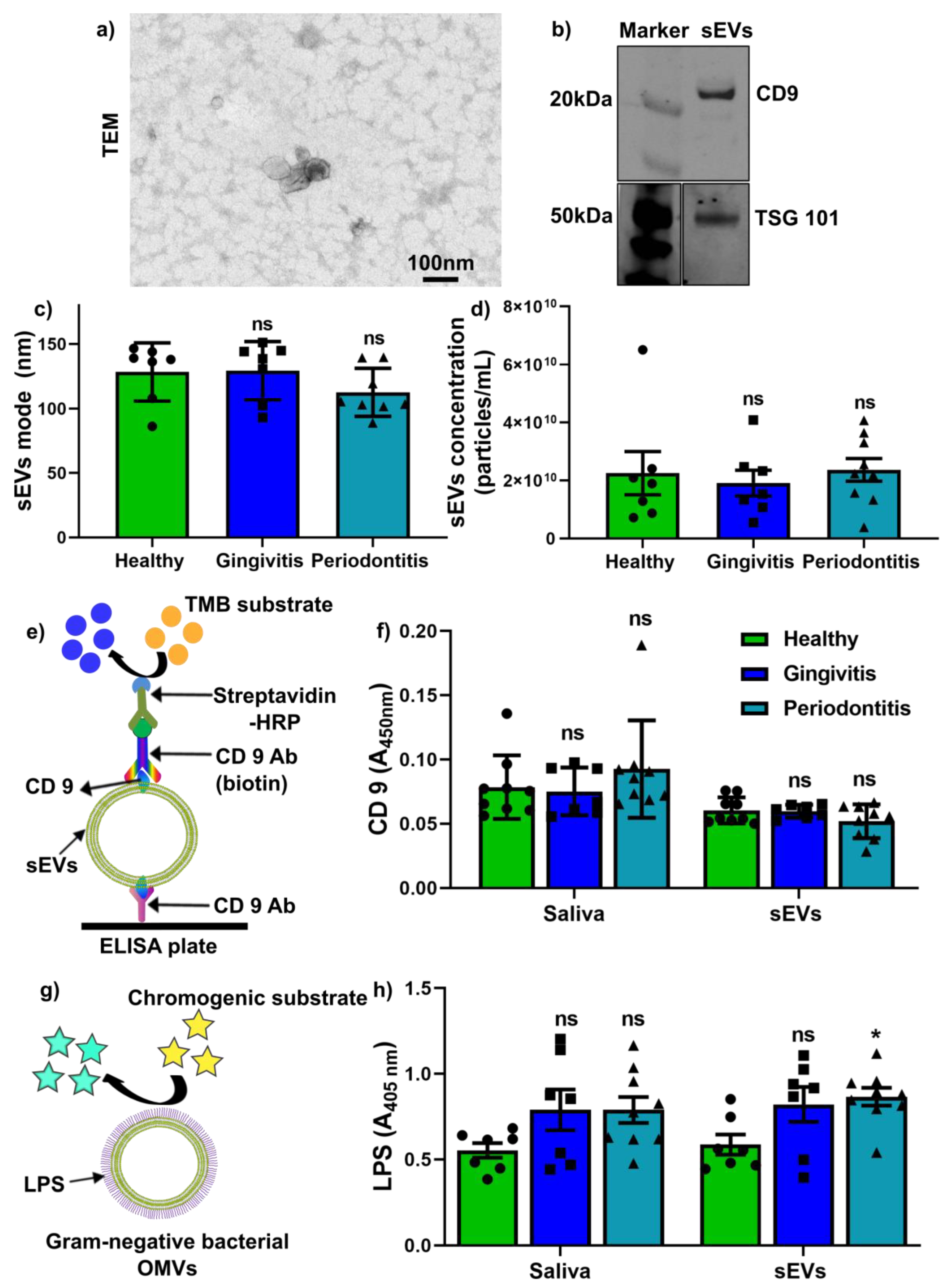
2.2. Salivary sEV Characterisation and Quantification of CD9+ sEVs and LPS+ OMVs
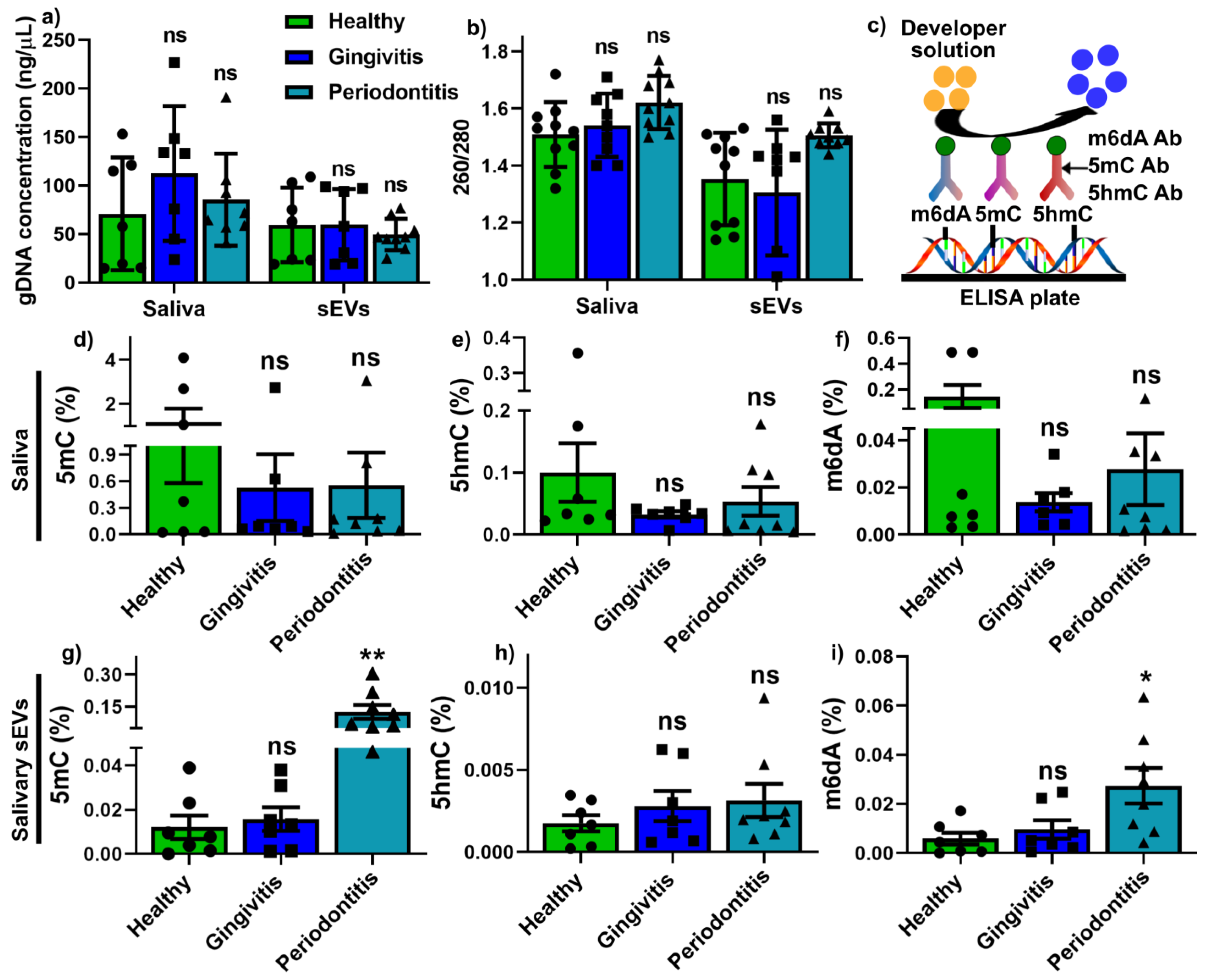
2.3. Global Methylation Profile in Saliva and Salivary sEVs
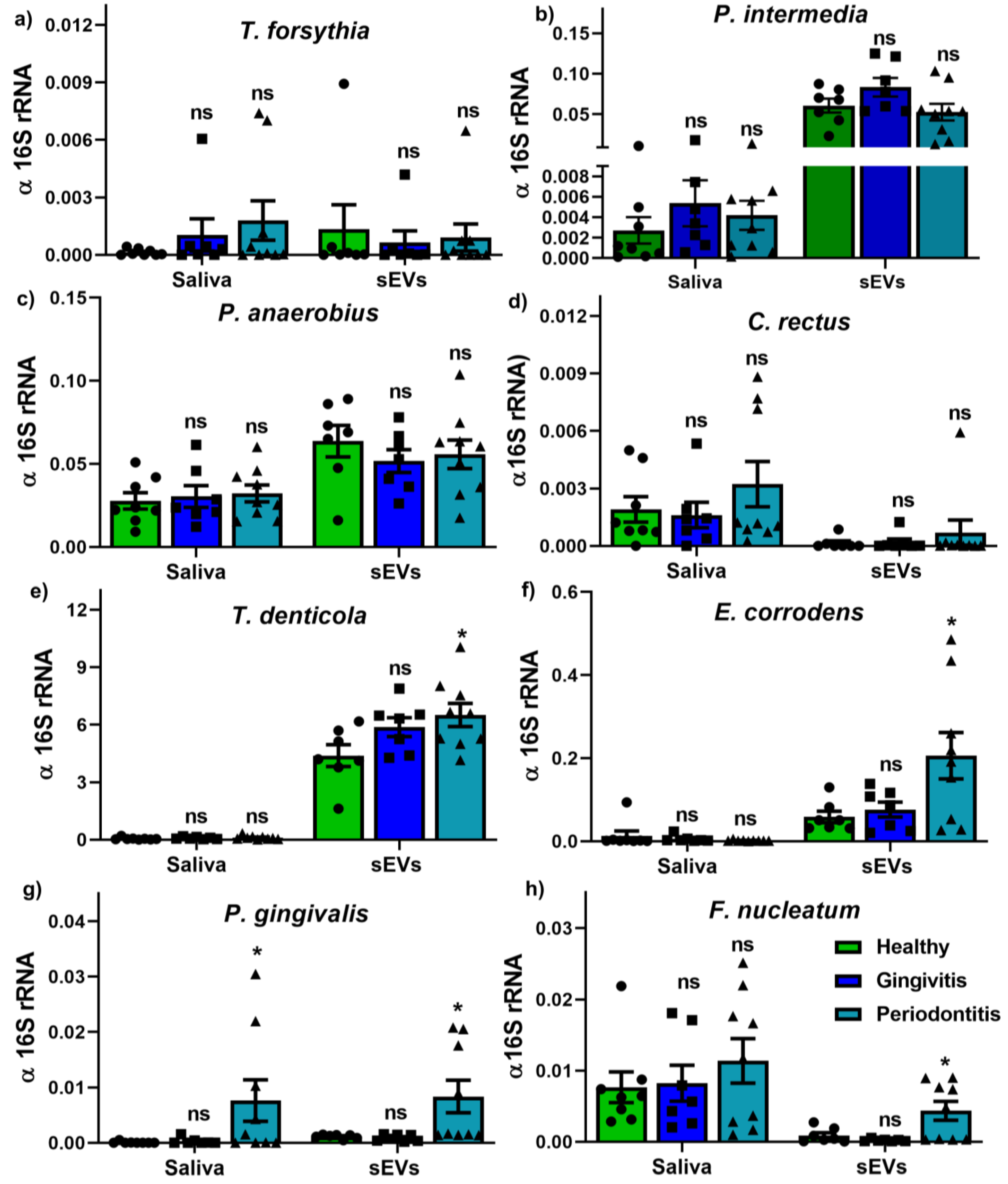
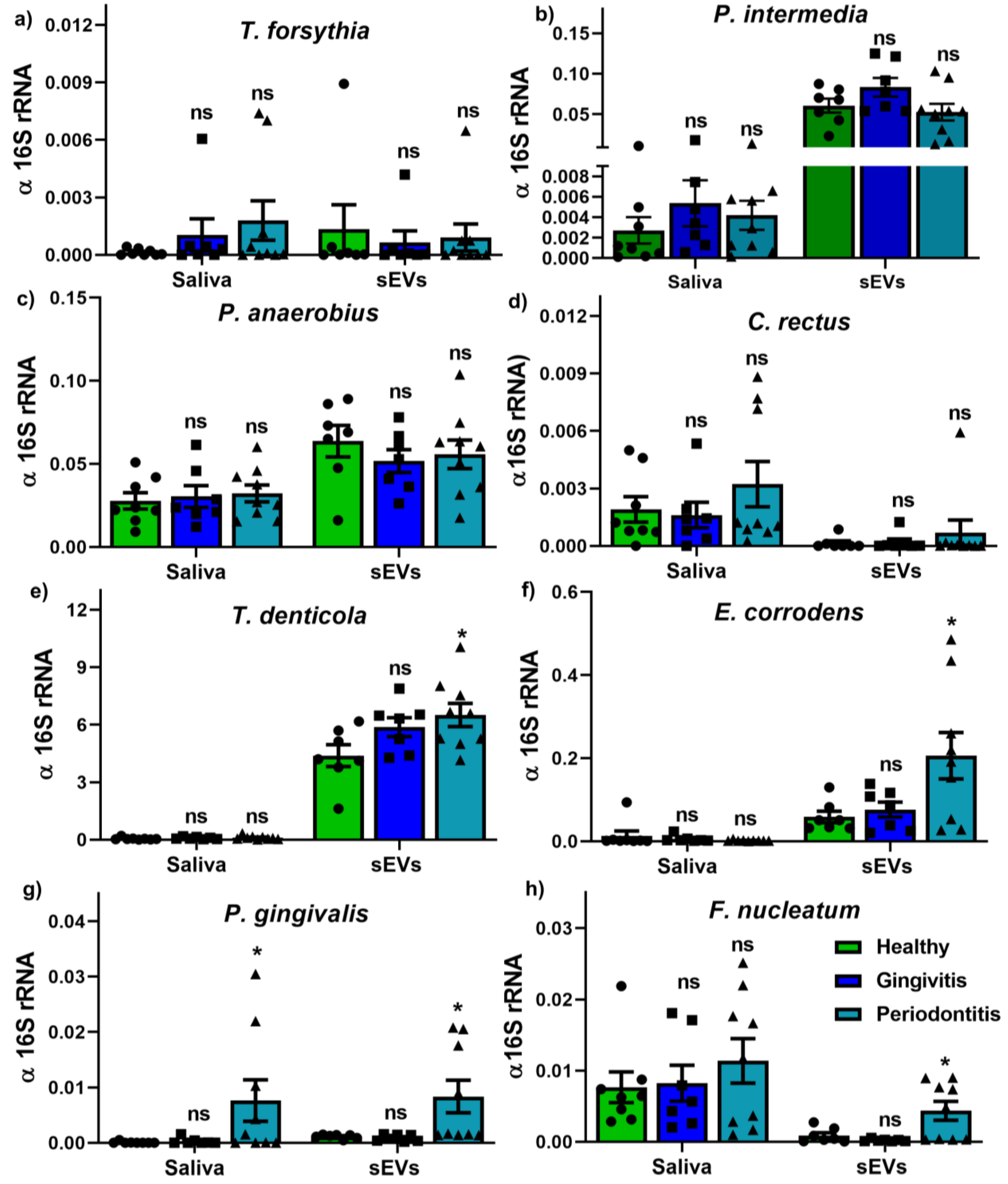
2.4. Bacterium and OMV Detection in Saliva and Salivary sEVs
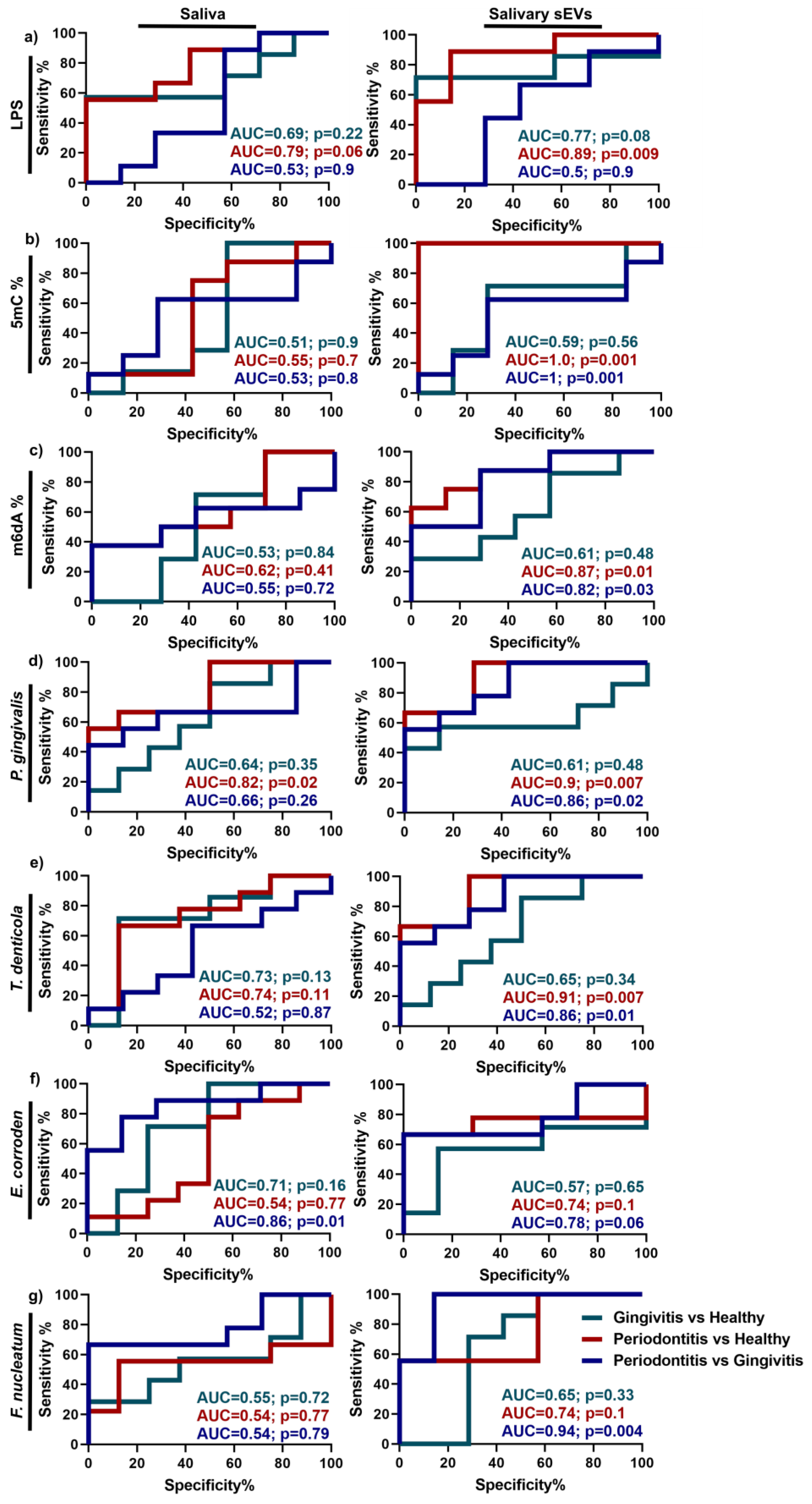
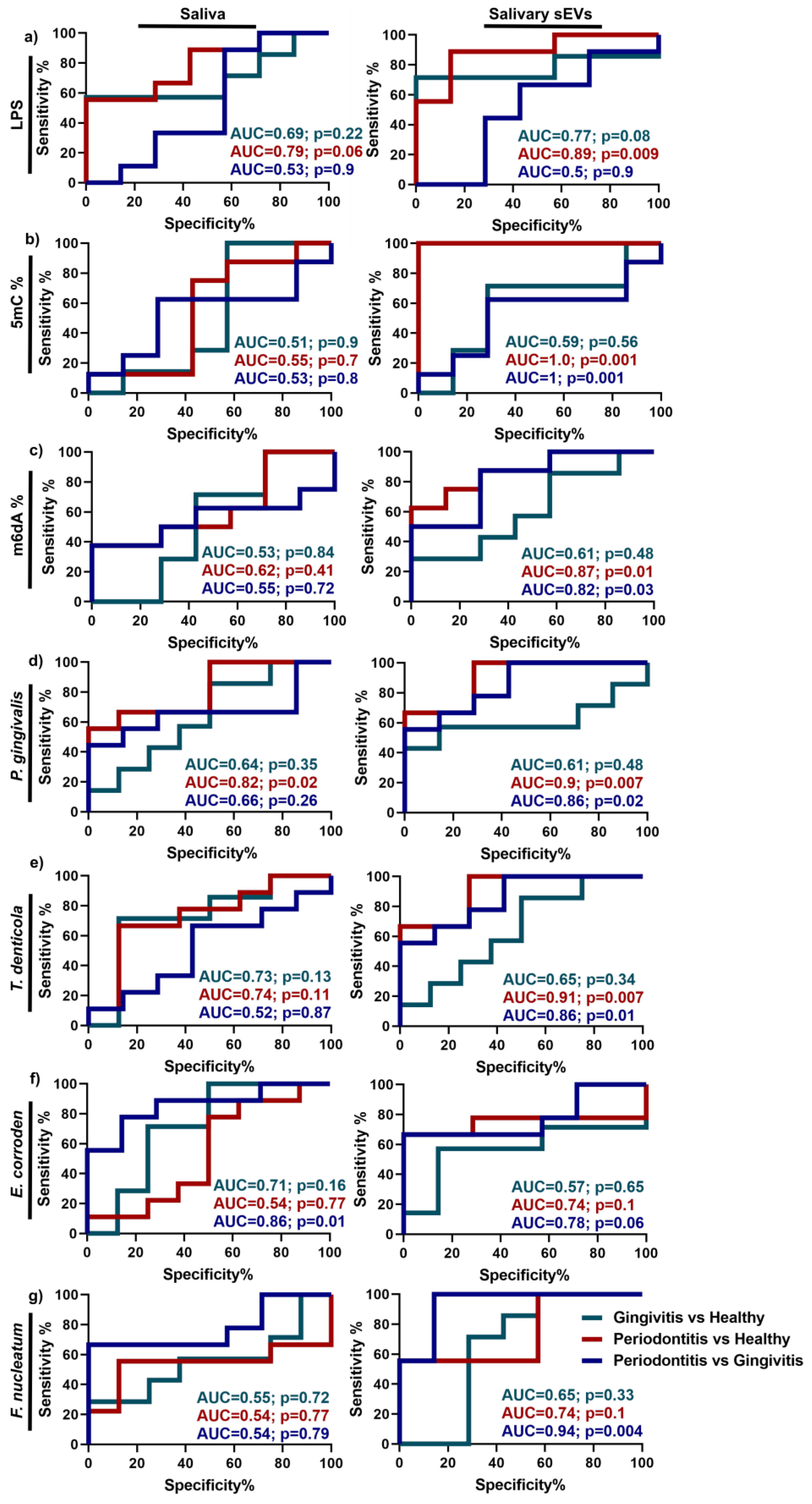
2.5. Discrimination Power of Up-Regulated Parameters in Periodontitis
3. Discussion
4. Materials and Methods
4.1. Participant Recruitment
4.2. Salivary sEV Isolation and Characterisation
4.3. Determination of CD9 and Lipopolysaccharide (LPS) in Saliva and sEVs
4.4. Genomic DNA Isolation and DNA Methylation Landscape Quantification
4.5. gDNA Real-Time Quantitative PCR (qPCR)
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nazir, M.A. Prevalence of periodontal disease, its association with systemic diseases and prevention. Int. J. Health Sci. 2017, 11, 72–80. [Google Scholar]
- Tonetti, M.S.; Greenwell, H.; Kornman, K.S. Staging and grading of periodontitis: Framework and proposal of a new classification and case definition. J. Clin. Periodontol. 2018, 89, 1475. [Google Scholar]
- Bartold, P.M.; Van Dyke, T.E. Periodontitis: A host-mediated disruption of microbial homeostasis. Unlearning learned concepts. Periodontol. 2000 2013, 62, 203–217. [Google Scholar] [CrossRef] [Green Version]
- Preshaw, P.M. Detection and diagnosis of periodontal conditions amenable to prevention. BMC Oral Health 2015, 15, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Pfaffe, T.; Cooper-White, J.; Beyerlein, P.; Kostner, K.; Punyadeera, C. Diagnostic Potential of Saliva: Current State and Future Applications. Clin. Chem. 2011, 57, 675–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almiñana-Pastor, P.J.; Boronat-Catalá, M.; Micó-Martinez, P.; Bellot-Arcís, C.; Lopez-Roldan, A.; Alpiste-Illueca, F.M.; Lopez-Rolda, A. Epigenetics and periodontics: A systematic review. Med. Oral Patol. Oral Cir. Bucal 2019, 24, e659–e672. [Google Scholar] [CrossRef] [PubMed]
- Kc, S.; Wang, X.Z.; Gallagher, J.E.; Sukriti, K. Diagnostic sensitivity and specificity of host-derived salivary biomarkers in periodontal disease amongst adults: Systematic review. J. Clin. Periodontol. 2019, 47, 289–308. [Google Scholar] [CrossRef] [PubMed]
- Arias-Bujanda, N.; Regueira-Iglesias, A.; Balsa-Castro, C.; Nibali, L.; Donos, N.; Tomás, I. Accuracy of single molecular biomarkers in saliva for the diagnosis of periodontitis: A systematic review and meta-analysis. J. Clin. Periodontol. 2020, 47, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Toczewska, J.; Konopka, T.; Zalewska, A.; Maciejczyk, M. Nitrosative Stress Biomarkers in the Non-Stimulated and Stimulated Saliva, as well as Gingival Crevicular Fluid of Patients with Periodontitis: Review and Clinical Study. Antioxidants 2020, 9, 259. [Google Scholar] [CrossRef] [Green Version]
- Toczewska, J.; Maciejczyk, M.; Konopka, T.; Zalewska, A. Total Oxidant and Antioxidant Capacity of Gingival Crevicular Fluid and Saliva in Patients with Periodontitis: Review and Clinical Study. Antioxidants 2020, 9, 450. [Google Scholar] [CrossRef] [PubMed]
- Tóthová, L.; Celec, P. Oxidative Stress and Antioxidants in the Diagnosis and Therapy of Periodontitis. Front. Physiol. 2017, 8, 1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.; El Andaloussi, S.; Wood, M.J. Exosomes and microvesicles: Extracellular vesicles for genetic information transfer and gene therapy. Hum. Mol. Genet. 2012, 21, R125–R134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, P.; Ivanovski, S. Saliva—Friend and Foe in the COVID-19 Outbreak. Diagnostics 2020, 10, 290. [Google Scholar] [CrossRef] [PubMed]
- Tobón-Arroyave, S.I.; Celis-Mejía, N.; Córdoba-Hidalgo, M.P.; Isaza-Guzmán, D.M. Decreased salivary concentration of CD9 and CD81 exosome-related tetraspanins may be associated with the periodontal clinical status. J. Clin. Periodontol. 2019, 46, 470–480. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Lin, Y.; Xiong, X.; Li, K.; Yao, Z.; Dong, H.; Jiang, Z.; Yu, D.; Yeung, S.-C.J.; Zhang, H. Detection of Exosomal PD-L1 RNA in Saliva of Patients with Periodontitis. Front. Genet. 2019, 10, 202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, P.; Bartold, P.M.; Salomon, C.; Ivanovski, S. Salivary Small Extracellular Vesicles Associated miRNAs in Periodontal Status—A Pilot Study. Int. J. Mol. Sci. 2020, 21, 2809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padilla, A.C.; Aracena, L.W.; Fuentes, O.R.; Busquetts, D.A.; Ríos, M.H.; Lobos, V.R.; La Rocca, A.P.; Molina, J.N.; Varas, V.B.; Acuña-Gallardo, S.; et al. Molecular signatures of extracellular vesicles in oral fluids of periodontitis patients. Oral Dis. 2020, 26, 1318–1325. [Google Scholar] [CrossRef] [PubMed]
- Han, P.; Lai, A.; Salomon, C.; Ivanovski, S. Detection of Salivary Small Extracellular Vesicles Associated Inflammatory Cytokines Gene Methylation in Gingivitis. Int. J. Mol. Sci. 2020, 21, 5273. [Google Scholar] [CrossRef]
- Huang, X.; Hu, X.; Zhao, M.; Zhang, Q. Analysis of salivary exosomal proteins in young adults with severe periodontitis. Oral Dis. 2020, 26, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Kamal, N.N.S.N.M.; Awang, R.A.R.; Mohamad, S.; Shahidan, W.N.S. Plasma- and Saliva Exosome Profile Reveals a Distinct MicroRNA Signature in Chronic Peri-odontitis. Front. Physiol. 2020, 11, 1561. [Google Scholar]
- Dewhirst, F.E.; Chen, T.; Izard, J.; Paster, B.J.; Tanner, A.C.R.; Yu, W.-H.; Lakshmanan, A.; Wade, W.G. The Human Oral Microbiome. J. Bacteriol. 2010, 192, 5002–5017. [Google Scholar] [CrossRef] [Green Version]
- Rudney, J.D.; Jagtap, P.D.; Reilly, C.S.; Chen, R.; Markowski, T.W.; Higgins, L.; Johnson, J.E.; Griffin, T.J. Protein relative abundance patterns associated with sucrose-induced dysbiosis are conserved across taxonomically diverse oral microcosm biofilm models of dental caries. Microbiome 2015, 3, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Cecil, J.D.; Sirisaengtaksin, N.; O’Brien-Simpson, N.M.; Krachler, A.M. Outer Membrane Vesicle-Host Cell Interactions. Microbiol. Spectr. 2019, 7, 201–214. [Google Scholar]
- Sharma, A.; Novak, E.K.; Sojar, H.T.; Swank, R.T.; Kuramitsu, H.K.; Genco, R.J. Porphyromonas gingivalis platelet aggregation activity: Outer membrane vesicles are potent activators of murine platelets. Oral Microbiol. Immunol. 2000, 15, 393–396. [Google Scholar] [CrossRef]
- Yoshimura, M.; Ohara, N.; Kondo, Y.; Shoji, M.; Okano, S.; Nakano, Y.; Abiko, Y.; Nakayama, K. Proteome analysis of Porphyromonas gingivalis cells placed in a subcutaneous chamber of mice. Oral Microbiol. Immunol. 2008, 23, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Schübeler, D. Function and information content of DNA methylation. Nature 2015, 517, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Li, E.; Zhang, Y. DNA Methylation in Mammals. Cold Spring Harb. Perspect. Biol. 2014, 6, a019133. [Google Scholar] [CrossRef]
- Li, X.; Zhao, Q.; Wei, W.; Lin, Q.; Magnan, C.; Emami, M.R.; Da Silva, L.E.W.; Viola, T.W.; Marshall, P.R.; Yin, J.; et al. The DNA modification N6-methyl-2′-deoxyadenosine (m6dA) drives activity-induced gene expression and is required for fear extinction. Nat. Neurosci. 2019, 22, 534–544. [Google Scholar] [CrossRef] [PubMed]
- Sina, A.A.I.; Carrascosa, L.G.; Liang, Z.; Grewal, Y.S.; Wardiana, A.; Shiddiky, M.J.; Gardiner, R.A.; Samaratunga, H.; Gandhi, M.K.; Scott, R.J.; et al. Epigenetically reprogrammed methylation landscape drives the DNA self-assembly and serves as a universal cancer biomarker. Nat. Commun. 2018, 9, 4915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zlotogorski-Hurvitz, A.; Dayan, D.; Chaushu, G.; Korvala, J.; Salo, T.; Sormunen, R.; Vered, M. Human saliva-derived exosomes: Comparing methods of isolation. J. Histochem. Cytochem. 2015, 63, 181–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paolicelli, R.C.; Bergamini, G.; Rajendran, L. Cell-to-cell Communication by Extracellular Vesicles: Focus on Microglia. Neuroscience 2019, 405, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Damgaard, C.; Danielsen, A.K.; Enevold, C.; Massarenti, L.; Nielsen, C.H.; Holmstrup, P.; Belstrøm, D. Porphyromonas gingivalis in saliva associates with chronic and aggressive periodontitis. J. Oral Microbiol. 2019, 11, 1653123. [Google Scholar] [CrossRef] [Green Version]
- Maghbooli, Z.; Hossein-Nezhad, A.; Larijani, B.; Amini, M.; Keshtkar, A. Global DNA methylation as a possible biomarker for diabetic retinopathy. Diabetes Metab. Res. Rev. 2014, 31, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Reichetzeder, C.; Putra, S.E.D.; Pfab, T.; Slowinski, T.; Neuber, C.; Kleuser, B.; Hocher, B. Increased global placental DNA methylation levels are associated with gestational diabetes. Clin. Epigenetics 2016, 8, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.; Jia, L.; Zheng, Y.; Li, W. Salivary Exosomes: Emerging Roles in Systemic Disease. Int. J. Biol. Sci. 2018, 14, 633–643. [Google Scholar] [CrossRef] [PubMed]
- Han, P.; Ivanovski, S. Effect of Saliva Collection Methods on the Detection of Periodontium-Related Genetic and Epigenetic Bi-omarkers-A Pilot Study. Int. J. Mol. Sci. 2019, 20, 4729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Théry, C.; Amigorena, S.; Raposo, G.; Clayton, A. Isolation and Characterization of Exosomes from Cell Culture Supernatants and Biological Fluids. Curr. Protoc. Cell Biol. 2006, 30, 3.22.1–3.22.29. [Google Scholar] [CrossRef]
- Han, P.; Wu, C.; Xiao, Y. The effect of silicate ions on proliferation, osteogenic differentiation and cell signalling pathways (WNT and SHH) of bone marrow stromal cells. Biomater. Sci. 2013, 1, 379–392. [Google Scholar] [CrossRef]
- Han, P.; Ivanovski, S.; Crawford, R.; Xiao, Y. Activation of the Canonical Wnt Signaling Pathway Induces Cementum Regeneration. J. Bone Min. Res. 2015, 30, 1160–1174. [Google Scholar] [CrossRef] [Green Version]
- Han, P.; Wu, C.; Chang, J.; Xiao, Y. The cementogenic differentiation of periodontal ligament cells via the activation of Wnt/beta-catenin signalling pathway by Li+ ions released from bioactive scaffolds. Biomaterials 2012, 33, 6370–6379. [Google Scholar] [CrossRef] [PubMed]
- Han, P.; Frith, J.E.; Gomez, G.A.; Yap, A.S.; O’Neill, G.M.; Cooper-White, J.J. Five Piconewtons: The Difference between Osteogenic and Adipogenic Fate Choice in Human Mesenchymal Stem Cells. Acs Nano 2019, 13, 11129–11143. [Google Scholar] [CrossRef] [PubMed]
- Han, P.; Lloyd, T.; Chen, Z.; Xiao, Y. Proinflammatory Cytokines Regulate Cementogenic Differentiation of Periodontal Ligament Cells by Wnt/Ca(2+) Sig-naling Pathway. J. Interferon Cytokine Res. 2016, 36, 328–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Healthy (n = 7) | Gingivitis (n = 7) | Periodontitis (n = 8) | ||
|---|---|---|---|---|
| Gender | Male | 4 (57.1%) | 6 (85.7%) | 5 (62.5%) |
| Female | 3 (42.9%) | 1 (14.3%) | 3 (37.5%) | |
| Age | 35.1 ± 7.8 (24–48) | 30.5 ± 5.7 (28–40) p = 0.57 | 48.2 ± 10.6 (38–66) * p = 0.02 | |
| Smoking habit | Non-smokers | 7 (100%) | 7 (100%) | 6 (75%) |
| Smokers | 0 | 0 | 2 (25%) | |
| Ethnicity | Caucasians | 4 (57.1%) | 2 (28.5%) | 3 (37.5%) |
| Asians | 3 (42.9%) | 5 (71.5%) | 5 (62.5%) | |
| BOP | 11% ± 3.1 (6%–14%) | 48.5 % ± 14.6 (34%–70%) * p = 0.0001 | 47.5 % ± 18 (25%–74%) * p = 0.0001 | |
| PI | 15.1 % ± 6.9 (6%–28%) | 51.5 % ± 27 (9%–89%) * p = 0.08 | 77.3 % ± 16.7 (53%–94%) * p = 0.0006 | |
| No. of deep pockets (≥5 mm) | 24 ± 21.4 (5–55) | |||
| Average PPD (mm) | 4.95 ± 1.4 (2.5–6.4) | |||
| Periodontitis classification | Localised (<30%) | 3 (37.5%) | ||
| Generalised (≥30%) | 5 (62.5%) | |||
| Grade B | 5 (62.5%) | |||
| Grade C | 3 (37.5%) | |||
| Stage III | 7 (85.7%) | |||
| Stage IV | 1 (14.3%) |
| Forward Primer (5′-3′) | Reverse Primer (5′-3′) | |
|---|---|---|
| T. forsythia | GGGTGAGTAACGCGTATGTAACCT | CCCATCCGCAACCAATAAA |
| P. gingivalis | TGCAACTTGCCTTACAGAGGG | ACTCGTATCGCCCGTTATTC |
| T. denticola | TGGTGAGTAACGCGTGGGTGACCT | TTCACCCTCTCAGGCCGGA |
| P. intermedia | CCACATATGGCATCTGACGTG | CACGCTACTTGGCTGGTTCA |
| F. nucleatum | GGATTTATTGGGCGTAAAGC | GGCATTCCTACAAATATCTACGAA |
| C. rectus | TTTCGGAGCGTAAACTCCTTTTC | TGATTCCGAGTAACGCTTGCA |
| P. anaerobius | GGGTGAGTAACGCGTGGGT | TACTGATCGTCGCCTTGGTGG |
| E. corrodens | ACGTCCTACGGGAGAAAGCGG | CCATTGTCCAAAATTCCCCACTG |
| 16S rRNA | TGGAGCATGTGGTTTAATTCGA | TGCGGGACTTAACCCAACA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, P.; Bartold, P.M.; Salomon, C.; Ivanovski, S. Salivary Outer Membrane Vesicles and DNA Methylation of Small Extracellular Vesicles as Biomarkers for Periodontal Status: A Pilot Study. Int. J. Mol. Sci. 2021, 22, 2423. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052423
Han P, Bartold PM, Salomon C, Ivanovski S. Salivary Outer Membrane Vesicles and DNA Methylation of Small Extracellular Vesicles as Biomarkers for Periodontal Status: A Pilot Study. International Journal of Molecular Sciences. 2021; 22(5):2423. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052423
Chicago/Turabian StyleHan, Pingping, Peter Mark Bartold, Carlos Salomon, and Sašo Ivanovski. 2021. "Salivary Outer Membrane Vesicles and DNA Methylation of Small Extracellular Vesicles as Biomarkers for Periodontal Status: A Pilot Study" International Journal of Molecular Sciences 22, no. 5: 2423. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052423