
Nutraceuticals in the Prevention of Neonatal Hypoxia–Ischemia: A Comprehensive Review of their Neuroprotective Properties, Mechanisms of Action and Future Directions

 , , ,
, , , 
Abstract
:1. Introduction
2. Methods
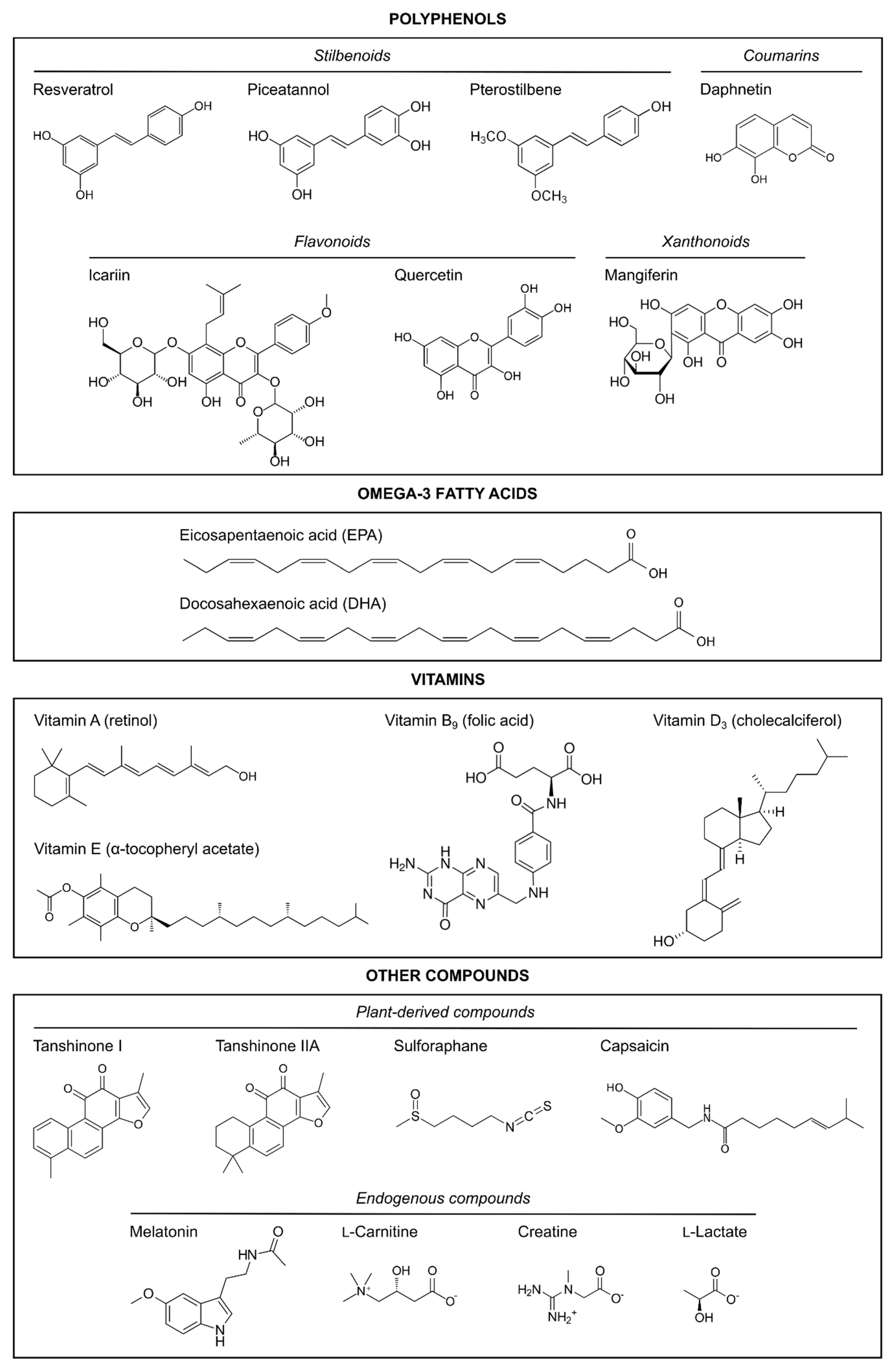
3. Polyphenols
3.1. Resveratrol
3.2. Resveratrol Derivatives: Piceatannol and Pterostilbene
3.2.1. Piceatannol
3.2.2. Pterostilbene
3.3. Quercetin
3.4. Mangiferin
3.5. Pomegranate Juice Polyphenols
3.6. Grape Seed Proanthocyanidin Extract
3.7. Other Polyphenols from Traditional Chinese Medicines: Icariin and Daphnetin
3.7.1. Icariin
3.7.2. Daphnetin
4. Omega-3 Fatty Acids
5. Vitamins
5.1. Vitamin A
5.2. Vitamin B9
5.3. Vitamin D
5.4. Vitamin E
6. Other Neuroprotective Natural Compounds
6.1. Plant-Derived Compounds
6.1.1. Tanshinones
6.1.2. Sulforaphane
6.1.3. Capsaicin
6.2. Endogenous Compounds
6.2.1. Melatonin
6.2.2. l-Carnitine
6.2.3. Creatine
6.2.4. Lactate
7. Conclusions and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ADNES | Advanced neuroprotection strategy |
| ALA | Alpha linoleic acid |
| BBB | Blood–brain barrier |
| CNS | Central nervous system |
| DHA | Docosahexaenoic acid |
| E | Embryonic day |
| EPA | Eicosapentaenoic acid |
| GSPE | Grape seed proanthocyanidin extract |
| HI | Hypoxia–ischemia |
| i.p. | Intraperitoneal |
| IL | Interleukin |
| iNOS | Inducible nitric oxide synthase |
| IVH | Intraventricular hemorrhage |
| n-3 PUFAs | Omega-3 polyunsaturated fatty acids |
| NO | Nitric oxide |
| Nrf/HO-1 | Nuclear factor erythroid 2 related factor 2/heme oxygenase 1 |
| P | Postnatal day |
| PI3K/Akt | Phosphoinositide 3-kinase/protein kinase B |
| RA | Retinoic acid |
| RAR | Retinoic acid receptor |
| ROS | Reactive oxygen species |
| RXR | Retinoic acid X receptor |
| TLR4/NF-κB | Toll-like receptor 4/nuclear factor κB |
| TNF-α | Tumor necrosis factor α |
| TRPV1 | Transient receptor potential vanilloid 1 |
References
- Shaw, O.E.F.; Yager, J.Y. Preventing childhood and lifelong disability: Maternal dietary supplementation for perinatal brain injury. Pharm. Res. 2019, 139, 228–242. [Google Scholar] [CrossRef]
- Rocha-Ferreira, E.; Hristova, M. Antimicrobial peptides and complement in neonatal hypoxia-ischemia induced brain damage. Front. Immunol 2015, 6, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Concepcion, K.; Meng, X.; Zhang, L. Brain-immune interactions in perinatal hypoxic-ischemic brain injury. Prog. Neurobiol. 2017, 159, 50–68. [Google Scholar] [CrossRef]
- Kurinczuk, J.J.; White-Koning, M.; Badawi, N. Epidemiology of neonatal encephalopathy and hypoxic-ischaemic encephalopathy. Early Hum. Dev. 2010, 86, 329–338. [Google Scholar] [CrossRef]
- Jantzie, L.L.; Todd, K.G.; Po-Yin, C. Neonatal ischemic stroke: A hypoxic-ischemic injury to the developing brain. Future Neurol. 2008, 3, 99–102. [Google Scholar] [CrossRef] [Green Version]
- Greco, P.; Nencini, G.; Piva, I.; Scioscia, M.; Volta, C.A.; Spadaro, S.; Neri, M.; Bonaccorsi, G.; Greco, F.; Cocco, I.; et al. Pathophysiology of hypoxic-ischemic encephalopathy: A review of the past and a view on the future. Acta Neurol. Belg. 2020, 120, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Allen, K.A.; Brandon, D.H. Hypoxic ischemic encephalopathy: Pathophysiology and experimental treatments. Newborn Infant Nurs. Rev. 2011, 11, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Lipton, P. Ischemic cell death in brain neurons. Physiol. Rev. 1999, 79, 1431–1568. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Tucker, D.; Dong, Y.; Zhao, N.; Zhuo, X.; Zhang, Q. Role of mitochondria in neonatal hypoxic-ischemic brain injury. J. Neurosci. Rehabil. 2015, 2, 1–14. [Google Scholar] [PubMed]
- Buonocore, G.; Perrone, S.; Bracci, R. Free radicals and brain damage in the newborn. Biol. Neonate. 2001, 79, 180–186. [Google Scholar] [CrossRef]
- Iadecola, C. Bright and dark sides of nitric oxide in ischemic brain injury. Trends Neurosci. 1997, 20, 132–139. [Google Scholar] [CrossRef]
- Kalogeris, T.; Bao, Y.; Korthuis, R.J. Mitochondrial reactive oxygen species: A double edged sword in ischemia/reperfusion vs preconditioning. Redox Biol. 2014, 2, 702–714. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; McCullough, L.D. Inflammatory responses in hypoxic ischemic encephalopathy. Acta Pharm. Sin. 2013, 34, 1121–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moro, M.A.; Cardenas, A.; Hurtado, O.; Leza, J.C.; Lizasoain, I. Role of nitric oxide after brain ischaemia. Cell Calcium 2004, 36, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Roka, A.; Azzopardi, D. Therapeutic hypothermia for neonatal hypoxic ischaemic encephalopathy. Early Hum. Dev. 2010, 86, 361–367. [Google Scholar] [CrossRef]
- Higgins, R.D.; Shankaran, S. Hypothermia: Novel approaches for premature infants. Early Hum. Dev. 2011, 87 (Suppl. S1), S17–S18. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, S.E.; Berg, M.; Hunt, R.; Tarnow-Mordi, W.O.; Inder, T.E.; Davis, P.G. Cooling for newborns with hypoxic ischaemic encephalopathy. Cochrane Database Syst. Rev. 2013, 4, CD003311. [Google Scholar] [CrossRef]
- George, P.M.; Steinberg, G.K. Novel stroke therapeutics: Unraveling stroke pathophysiology and its impact on clinical treatments. Neuron 2015, 87, 297–309. [Google Scholar] [CrossRef] [Green Version]
- Ayuso, M.I.; Montaner, J. Advanced neuroprotection for brain ischemia: An alternative approach to minimize stroke damage. Expert Opin. Investig. Drugs 2015, 24, 1137–1142. [Google Scholar] [CrossRef] [Green Version]
- Ayuso, M.I.; Gonzalo-Gobernado, R.; Montaner, J. Neuroprotective diets for stroke. Neurochem. Int. 2017, 107, 4–10. [Google Scholar] [CrossRef]
- Escudero-Martinez, I.; Mancha, F.; Vega-Salvatierra, A.; Ayuso, M.I.; Ocete, R.F.; Algaba, P.; Lopez-Rueda, A.; Pinero, P.; Fajardo, E.; Fernandez-Engo, J.R.; et al. Mediterranean diet and physical activity protect from silent brain infarcts in a cohort of patients with atrial fibrillation. J. Stroke 2019, 21, 353–355. [Google Scholar] [CrossRef] [Green Version]
- Van Cann, J.; Koskela, E.; Mappes, T.; Sims, A.; Watts, P.C. Intergenerational fitness effects of the early life environment in a wild rodent. J. Anim. Ecol. 2019, 88, 1355–1365. [Google Scholar] [CrossRef]
- Shlobin, N.A.; LoPresti, M.A.; Du, R.Y.; Lam, S. Folate fortification and supplementation in prevention of folate-sensitive neural tube defects: A systematic review of policy. J. Neurosurg. Pediatr. 2020, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Fischer, M.; Stronati, M.; Lanari, M. Mediterranean diet, folic acid, and neural tube defects. Ital. J. Pediatr. 2017, 43, 74. [Google Scholar] [CrossRef] [Green Version]
- Semple, B.D.; Blomgren, K.; Gimlin, K.; Ferriero, D.M.; Noble-Haeusslein, L.J. Brain development in rodents and humans: Identifying benchmarks of maturation and vulnerability to injury across species. Prog. Neurobiol. 2013, 106–107, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Larson, A.C.; Jamrogowicz, J.L.; Kulikowicz, E.; Wang, B.; Yang, Z.J.; Shaffner, D.H.; Koehler, R.C.; Lee, J.K. Cerebrovascular autoregulation after rewarming from hypothermia in a neonatal swine model of asphyxic brain injury. J. Appl Physiol (1985) 2013, 115, 1433–1442. [Google Scholar] [CrossRef] [Green Version]
- Derrick, M.; Luo, N.L.; Bregman, J.C.; Jilling, T.; Ji, X.; Fisher, K.; Gladson, C.L.; Beardsley, D.J.; Murdoch, G.; Back, S.A.; et al. Preterm fetal hypoxia-ischemia causes hypertonia and motor deficits in the neonatal rabbit: A model for human cerebral palsy? J. Neurosci. 2004, 24, 24–34. [Google Scholar] [CrossRef]
- Nitsos, I.; Newnham, J.P.; Rees, S.M.; Harding, R.; Moss, T.J. The impact of chronic intrauterine inflammation on the physiologic and neurodevelopmental consequences of intermittent umbilical cord occlusion in fetal sheep. Reprod. Sci. 2014, 21, 658–670. [Google Scholar] [CrossRef]
- Juul, S.E.; McPherson, R.J.; Farrell, F.X.; Jolliffe, L.; Ness, D.J.; Gleason, C.A. Erytropoietin concentrations in cerebrospinal fluid of nonhuman primates and fetal sheep following high-dose recombinant erythropoietin. Biol. Neonate. 2004, 85, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Rice, J.E., 3rd; Vannucci, R.C.; Brierley, J.B. The influence of immaturity on hypoxic-ischemic brain damage in the rat. Ann. Neurol. 1981, 9, 131–141. [Google Scholar] [CrossRef]
- Levine, S. Anoxic-ischemic encephalopathy in rats. Am. J. Pathol 1960, 36, 1–17. [Google Scholar] [PubMed]
- Vannucci, S.J.; Hagberg, H. Hypoxia-ischemia in the immature brain. J. Exp. Biol. 2004, 207, 3149–3154. [Google Scholar] [CrossRef] [Green Version]
- Wigglesworth, J.S. Effects of varying severity of growth retardation on organ weight and cell population in foetal rats. J. Pathol. 1970, 101, PXII. [Google Scholar] [CrossRef] [Green Version]
- Helmy, M.M.; Ruusuvuori, E.; Watkins, P.V.; Voipio, J.; Kanold, P.O.; Kaila, K. Acid extrusion via blood-brain barrier causes brain alkalosis and seizures after neonatal asphyxia. Brain 2012, 135, 3311–3319. [Google Scholar] [CrossRef] [Green Version]
- Bjelke, B.; Andersson, K.; Ögren, S.O.; Bolme, P. Asphyctic lesion: Proliferation of tyrosine hydroxylase-immunoreactive nerve cell bodies in the rat substantia nigra and functional changes in dopamine neurotransmission. Brain Res. 1991, 543, 1–9. [Google Scholar] [CrossRef]
- Bouslama, M.; Renaud, J.; Olivier, P.; Fontaine, R.H.; Matrot, B.; Gressens, P.; Gallego, J. Melatonin prevents learning disorders in brain-lesioned newborn mice. Neuroscience 2007, 150, 712–719. [Google Scholar] [CrossRef]
- Ikonomidou, C.; Price, M.T.; Mosinger, J.L.; Frierdich, G.; Labruyere, J.; Salles, K.S.; Olney, J.W. Hypobaric-ischemic conditions produce glutamate-like cytopathology in infant rat brain. J. Neurosci. 1989, 9, 1693–1700. [Google Scholar] [CrossRef]
- Vonder Haar, C.; Peterson, T.C.; Martens, K.M.; Hoane, M.R. Vitamins and nutrients as primary treatments in experimental brain injury: Clinical implications for nutraceutical therapies. Brain Res. 2016, 1640, 114–129. [Google Scholar] [CrossRef] [Green Version]
- Sibeko, L.; Johns, T. Global survey of medicinal plants during lactation and postpartum recovery: Evolutionary perspectives and contemporary health implications. J. Ethnopharmacol. 2021, 270, 113812. [Google Scholar] [CrossRef]
- Muñoz Balbontín, Y.; Stewart, D.; Shetty, A.; Fitton, C.A.; McLay, J.S. Herbal medicinal product use during pregnancy and the postnatal period: A systematic review. Obs. Gynecol. 2019, 133, 920–932. [Google Scholar] [CrossRef] [Green Version]
- Matthews, L.G.; Smyser, C.D.; Cherkerzian, S.; Alexopoulos, D.; Kenley, J.; Tuuli, M.G.; Nelson, D.M.; Inder, T.E. Maternal pomegranate juice intake and brain structure and function in infants with intrauterine growth restriction: A randomized controlled pilot study. PLoS ONE 2019, 14, e0219596. [Google Scholar] [CrossRef]
- Speer, M.E.; Blifeld, C.; Rudolph, A.J.; Chadda, P.; Holbein, M.E.; Hittner, H.M. Intraventricular hemorrhage and vitamin E in the very low-birth-weight infant: Evidence for efficacy of early intramuscular vitamin E administration. Pediatrics 1984, 74, 1107–1112. [Google Scholar]
- West, T.; Atzeva, M.; Holtzman, D.M. Pomegranate polyphenols and resveratrol protect the neonatal brain against hypoxic-ischemic injury. Dev. Neurosci. 2007, 29, 363–372. [Google Scholar] [CrossRef] [Green Version]
- Arteaga, O.; Revuelta, M.; Uriguen, L.; Alvarez, A.; Montalvo, H.; Hilario, E. Pretreatment with resveratrol prevents neuronal injury and cognitive deficits induced by perinatal hypoxia-ischemia in rats. PLoS ONE 2015, 10, e0142424. [Google Scholar] [CrossRef] [Green Version]
- Revuelta, M.; Arteaga, O.; Montalvo, H.; Alvarez, A.; Hilario, E.; Martinez-Ibarguen, A. Antioxidant treatments recover the alteration of auditory-evoked potentials and reduce morphological damage in the inferior colliculus after perinatal asphyxia in rat. Brain Pathol. 2016, 26, 186–198. [Google Scholar] [CrossRef]
- Gao, Y.; Fu, R.; Wang, J.; Yang, X.; Wen, L.; Feng, J. Resveratrol mitigates the oxidative stress mediated by hypoxic-ischemic brain injury in neonatal rats via Nrf2/HO-1 pathway. Pharm. Biol. 2018, 56, 440–449. [Google Scholar] [CrossRef] [PubMed]
- Isac, S.; Panaitescu, A.M.; Spataru, A.; Iesanu, M.; Totan, A.; Udriste, A.; Cucu, N.; Peltecu, G.; Zagrean, L.; Zagrean, A.M. Trans-resveratrol enriched maternal diet protects the immature hippocampus from perinatal asphyxia in rats. Neurosci. Lett. 2017, 653, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Dumont, U.; Sanchez, S.; Olivier, B.; Chateil, J.F.; Deffieux, D.; Quideau, S.; Pellerin, L.; Beauvieux, M.C.; Bouzier-Sore, A.K.; Roumes, H. Maternal alcoholism and neonatal hypoxia-ischemia: Neuroprotection by stilbenoid polyphenols. Brain Res. 2020, 1738, 146798. [Google Scholar] [CrossRef]
- Dumont, U.; Sanchez, S.; Olivier, B.; Chateil, J.F.; Pellerin, L.; Beauvieux, M.C.; Bouzier-Sore, A.K.; Roumes, H. Maternal consumption of piceatannol: A nutritional neuroprotective strategy against hypoxia-ischemia in rat neonates. Brain Res. 2019, 1717, 86–94. [Google Scholar] [CrossRef]
- Li, D.; Song, T.; Yang, L.; Wang, X.; Yang, C.; Jiang, Y. Neuroprotective actions of pterostilbene on hypoxic-ischemic brain damage in neonatal rats through upregulation of heme oxygenase-1. Int. J. Dev. Neurosci. 2016, 54, 22–31. [Google Scholar] [CrossRef]
- Blasina, F.; Vaamonde, L.; Silvera, F.; Tedesco, A.C.; Dajas, F. Intravenous nanosomes of quercetin improve brain function and hemodynamic instability after severe hypoxia in newborn piglets. Neurochem. Int. 2015, 89, 149–156. [Google Scholar] [CrossRef]
- Wu, M.; Liu, F.; Guo, Q. Quercetin attenuates hypoxia-ischemia induced brain injury in neonatal rats by inhibiting TLR4/NF-κB signaling pathway. Int. Immunopharmacol. 2019, 74, 105704. [Google Scholar] [CrossRef] [PubMed]
- Xi, J.S.; Wang, Y.F.; Long, X.X.; Ma, Y. Mangiferin potentiates neuroprotection by isoflurane in neonatal hypoxic brain injury by reducing oxidative stress and activation of phosphatidylinositol-3-kinase/Akt/mammalian target of rapamycin (PI3K/Akt/mTOR) signaling. Med. Sci. Monit. 2018, 24, 7459–7468. [Google Scholar] [CrossRef]
- Loren, D.J.; Seeram, N.P.; Schulman, R.N.; Holtzman, D.M. Maternal dietary supplementation with pomegranate juice is neuroprotective in an animal model of neonatal hypoxic-ischemic brain injury. Pediatr Res. 2005, 57, 858–864. [Google Scholar] [CrossRef] [Green Version]
- Tu, X.; Wang, M.; Liu, Y.; Zhao, W.; Ren, X.; Li, Y.; Liu, H.; Gu, Z.; Jia, H.; Liu, J.; et al. Pretreatment of grape seed proanthocyanidin extract exerts neuroprotective effect in murine model of neonatal hypoxic-ischemic brain injury by its antiapoptotic property. Cell Mol. Neurobiol. 2019, 39, 953–961. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Rong, Y.; Luo, L. Neuroprotective effects of icariin in neonatal hypoxia-ischemic brain damage via its anti-apoptotic property. Childs Nerv. Syst. 2021, 37, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Du, G.; Tu, H.; Li, X.; Pei, A.; Chen, J.; Miao, Z.; Li, J.; Wang, C.; Xie, H.; Xu, X.; et al. Daphnetin, a natural coumarin derivative, provides the neuroprotection against glutamate-induced toxicity in HT22 cells and ischemic brain injury. Neurochem. Res. 2014, 39, 269–275. [Google Scholar] [CrossRef]
- De Barros Mucci, D.; Fernandes, F.S.; Souza Ados, S.; Sardinha, F.L.; Soares-Mota, M.; Tavares do Carmo, M.G. Flaxseed mitigates brain mass loss, improving motor hyperactivity and spatial memory, in a rodent model of neonatal hypoxic-ischemic encephalopathy. Prostaglandins Leukot. Essent Fat. Acids 2015, 97, 13–19. [Google Scholar] [CrossRef]
- Decker, M.J.; Jones, K.; Keating, G.L.; Damato, E.G.; Darrah, R. Maternal dietary supplementation with omega-3 polyunsaturated fatty acids confers neuroprotection to the newborn against hypoxia-induced dopamine dysfunction. Sleep. Sci. 2016, 9, 94–99. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.J.; Mayurasakorn, K.; Vannucci, S.J.; Mastropietro, C.; Bazan, N.G.; Ten, V.S.; Deckelbaum, R.J. N-3 fatty acid rich triglyceride emulsions are neuroprotective after cerebral hypoxic-ischemic injury in neonatal mice. PLoS ONE 2013, 8, e56233. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Hu, X.; Yang, W.; Gao, Y.; Chen, J. Omega-3 polyunsaturated fatty acid supplementation confers long-term neuroprotection against neonatal hypoxic-ischemic brain injury through anti-inflammatory actions. Stroke 2010, 41, 2341–2347. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Liu, J.; Hu, X.; Li, P.; Leak, R.K.; Gao, Y.; Chen, J. n-3 Polyunsaturated fatty acids reduce neonatal hypoxic/ischemic brain injury by promoting phosphatidylserine formation and Akt signaling. Stroke 2015, 46, 2943–2950. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Zhang, H.; Mu, H.; Zhu, W.; Jiang, X.; Hu, X.; Shi, Y.; Leak, R.K.; Dong, Q.; Chen, J.; et al. Omega-3 polyunsaturated fatty acids mitigate blood-brain barrier disruption after hypoxic-ischemic brain injury. Neurobiol. Dis. 2016, 91, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Suganuma, H.; Arai, Y.; Kitamura, Y.; Hayashi, M.; Okumura, A.; Shimizu, T. Maternal docosahexaenoic acid-enriched diet prevents neonatal brain injury. Neuropathology 2010, 30, 597–605. [Google Scholar] [CrossRef]
- Berman, D.R.; Mozurkewich, E.; Liu, Y.; Barks, J. Docosahexaenoic acid pretreatment confers neuroprotection in a rat model of perinatal cerebral hypoxia-ischemia. Am. J. Obs. Gynecol 2009, 200, 305.e301–306. [Google Scholar] [CrossRef] [Green Version]
- Berman, D.R.; Liu, Y.Q.; Barks, J.; Mozurkewich, E. Docosahexaenoic acid confers neuroprotection in a rat model of perinatal hypoxia-ischemia potentiated by Escherichia coli lipopolysaccharide-induced systemic inflammation. Am. J. Obs. Gynecol 2010, 202, 469.e1-6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arteaga, O.; Revuelta, M.; Urigüen, L.; Martínez-Millán, L.; Hilario, E.; Álvarez, A. Docosahexaenoic acid reduces cerebral damage and ameliorates long-term cognitive impairments caused by neonatal hypoxia-ischemia in rats. Mol. Neurobiol. 2017, 54, 7137–7155. [Google Scholar] [CrossRef]
- Jiang, W.; Guo, M.; Gong, M.; Chen, L.; Bi, Y.; Zhang, Y.; Shi, Y.; Qu, P.; Liu, Y.; Chen, J.; et al. Vitamin A bio-modulates apoptosis via the mitochondrial pathway after hypoxic-ischemic brain damage. Mol. Brain 2018, 11, 14. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Chen, S.; Yang, M.-L.; Li, S.-Y.; Jiang, W.; Xiao, N. Vitamin A regulates neural stem cell proliferation in rats after hypoxic-ischemic brain damage via RARβ-mediated modulation of the β-catenin pathway. Neurosci. Lett. 2020, 727, 134922. [Google Scholar] [CrossRef]
- Ezquer, M.E.; Valdez, S.R.; Seltzer, A.M.; Jahn, G.A. Reversion by vitamin E treatment of the oxidative damage but not of the advancement in reproductive senescence produced by neonatal hypoxia or hypoxia-ischemia in female rats. Neuroendocrinology 2014, 99, 204–218. [Google Scholar] [CrossRef]
- Deniz, B.F.; Confortim, H.D.; Deckmann, I.; Miguel, P.M.; Bronauth, L.; de Oliveira, B.C.; Barbosa, S.; Cechinel, L.R.; Siqueira, I.R.; Pereira, L.O. Folic acid supplementation during pregnancy prevents cognitive impairments and BDNF imbalance in the hippocampus of the offspring after neonatal hypoxia-ischemia. J. Nutr. Biochem. 2018, 60, 35–46. [Google Scholar] [CrossRef]
- Dai, C.; Liu, Y.; Dong, Z. Tanshinone I alleviates motor and cognitive impairments via suppressing oxidative stress in the neonatal rats after hypoxic-ischemic brain damage. Mol. Brain 2017, 10, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, W.J.; Yang, M.; Fok, T.F.; Li, K.; Chan, W.Y.; Ng, P.C.; Ng, H.K.; Chik, K.W.; Wang, C.C.; Gu, G.J.; et al. Partial neuroprotective effect of pretreatment with tanshinone IIA on neonatal hypoxia-ischemia brain damage. Pediatr. Res. 2005, 58, 784–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ping, Z.; Liu, W.; Kang, Z.; Cai, J.; Wang, Q.; Cheng, N.; Wang, S.; Wang, S.; Zhang, J.H.; Sun, X. Sulforaphane protects brains against hypoxic-ischemic injury through induction of Nrf2-dependent phase 2 enzyme. Brain Res. 2010, 1343, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Black, A.M.; Armstrong, E.A.; Scott, O.; Juurlink, B.J.H.; Yager, J.Y. Broccoli sprout supplementation during pregnancy prevents brain injury in the newborn rat following placental insufficiency. Behav. Brain Res. 2015, 291, 289–298. [Google Scholar] [CrossRef]
- Khatibi, N.H.; Jadhav, V.; Charles, S.; Chiu, J.; Buchholz, J.; Tang, J.; Zhang, J.H. Capsaicin pre-treatment provides neurovascular protection against neonatal hypoxic-ischemic brain injury in rats. Acta Neurochir. Suppl. 2011, 111, 225–230. [Google Scholar] [CrossRef] [Green Version]
- Hutton, L.C.; Abbass, M.; Dickinson, H.; Ireland, Z.; Walker, D.W. Neuroprotective properties of melatonin in a model of birth asphyxia in the spiny mouse (Acomys cahirinus). Dev. Neurosci. 2009, 31, 437–451. [Google Scholar] [CrossRef]
- Watanabe, K.; Hamada, F.; Wakatsuki, A.; Nagai, R.; Shinohara, K.; Hayashi, Y.; Imamura, R.; Fukaya, T. Prophylactic administration of melatonin to the mother throughout pregnancy can protect against oxidative cerebral damage in neonatal rats. J. Matern. Fetal. Neonatal. Med. 2012, 25, 1254–1259. [Google Scholar] [CrossRef] [PubMed]
- Carloni, S.; Perrone, S.; Buonocore, G.; Longini, M.; Proietti, F.; Balduini, W. Melatonin protects from the long-term consequences of a neonatal hypoxic-ischemic brain injury in rats. J. Pineal Res. 2008, 44, 157–164. [Google Scholar] [CrossRef]
- Signorini, C.; Ciccoli, L.; Leoncini, S.; Carloni, S.; Perrone, S.; Comporti, M.; Balduini, W.; Buonocore, G. Free iron, total F-isoprostanes and total F-neuroprostanes in a model of neonatal hypoxic-ischemic encephalopathy: Neuroprotective effect of melatonin. J. Pineal Res. 2009, 46, 148–154. [Google Scholar] [CrossRef]
- Wainwright, M.S.; Mannix, M.K.; Brown, J.; Stumpf, D.A. L-Carnitine reduces brain injury after hypoxia-ischemia in newborn rats. Pediatr. Res. 2003, 54, 688–695. [Google Scholar] [CrossRef] [Green Version]
- Wainwright, M.S.; Kohli, R.; Whitington, P.F.; Chace, D.H. Carnitine treatment inhibits increases in cerebral carnitine esters and glutamate detected by mass spectrometry after hypoxia-ischemia in newborn rats. Stroke 2006, 37, 524–530. [Google Scholar] [CrossRef] [Green Version]
- Türkyilmaz, C.; Türkyilmaz, Z.; Onal, E.; Atalay, Y.; Söylemezoğlu, F.; Celasun, B. L-Carnitine pre-treatment reduces apoptotic cell death in seven-day-old rats hypoxia ischemia. Restor. Neurol. Neurosci. 2010, 28, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Ireland, Z.; Dickinson, H.; Snow, R.; Walker, D.W. Maternal creatine: Does it reach the fetus and improve survival after an acute hypoxic episode in the spiny mouse (Acomys cahirinus)? Am. J. Obs. Gynecol. 2008, 198, e431–e436. [Google Scholar] [CrossRef]
- Ireland, Z.; Castillo-Melendez, M.; Dickinson, H.; Snow, R.; Walker, D.W. A maternal diet supplemented with creatine from mid-pregnancy protects the newborn spiny mouse brain from birth hypoxia. Neuroscience 2011, 194, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Adcock, K.H.; Nedelcu, J.; Loenneker, T.; Martin, E.; Wallimann, T.; Wagner, B.P. Neuroprotection of creatine supplementation in neonatal rats with transient cerebral hypoxia-ischemia. Dev. Neurosci. 2002, 24, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Berger, R.; Middelanis, J.; Vaihinger, H.M.; Mies, G.; Wilken, B.; Jensen, A. Creatine protects the immature brain from hypoxic-ischemic injury. J. Soc. Gynecol. Investig. 2004, 11, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Roumes, H.; Dumont, U.; Sanchez, S.; Mazuel, L.; Blanc, J.; Raffard, G.; Chateil, J.F.; Pellerin, L.; Bouzier-Sore, A.K. Neuroprotective role of lactate in rat neonatal hypoxia-ischemia. J. Cereb. Blood Flow Metab. 2020, 41, 342–358. [Google Scholar] [CrossRef] [Green Version]
- Helmy, M.M.; Tolner, E.A.; Vanhatalo, S.; Voipio, J.; Kaila, K. Brain alkalosis causes birth asphyxia seizures, suggesting therapeutic strategy. Ann. Neurol. 2011, 69, 493–500. [Google Scholar] [CrossRef]
- Wigglesworth, J.S. Fetal growth retardation. Animal model: Uterine vessel ligation in the pregnant rat. Am. J. Pathol. 1974, 77, 347–350. [Google Scholar] [PubMed]
- Tsimogiannis, D.; Oreopoulou, V. Chapter 16. Classification of phenolic compounds in plants. In Polyphenols in Plants, 2nd ed.; Watson, R.R., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 263–284. [Google Scholar] [CrossRef]
- Manach, C.; Scalbert, A.; Morand, C.; Remesy, C.; Jimenez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [Green Version]
- Gu, H.F.; Mao, X.Y.; Du, M. Prevention of breast cancer by dietary polyphenols-role of cancer stem cells. Crit. Rev. Food Sci. Nutr. 2020, 60, 810–825. [Google Scholar] [CrossRef] [PubMed]
- Lall, R.K.; Syed, D.N.; Adhami, V.M.; Khan, M.I.; Mukhtar, H. Dietary polyphenols in prevention and treatment of prostate cancer. Int. J. Mol. Sci. 2015, 16, 3350–3376. [Google Scholar] [CrossRef] [PubMed]
- Dhakal, S.; Kushairi, N.; Phan, C.W.; Adhikari, B.; Sabaratnam, V.; Macreadie, I. Dietary polyphenols: A multifactorial strategy to target Alzheimer’s disease. Int. J. Mol. Sci. 2019, 20, 5090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serra, D.; Almeida, L.M.; Dinis, T.C.P. Polyphenols in the management of brain disorders: Modulation of the microbiota-gut-brain axis. Adv. Food Nutr. Res. 2020, 91, 1–27. [Google Scholar] [CrossRef]
- Guasch-Ferré, M.; Merino, J.; Sun, Q.; Fitó, M.; Salas-Salvadó, J. Dietary polyphenols, mediterranean diet, prediabetes, and type 2 diabetes: A narrative review of the evidence. Oxid. Med. Cell Longev. 2017, 2017, 6723931. [Google Scholar] [CrossRef]
- Tian, B.; Liu, J. Resveratrol: A review of plant sources, synthesis, stability, modification and food application. J. Sci. Food Agric. 2020, 100, 1392–1404. [Google Scholar] [CrossRef]
- Catalgol, B.; Batirel, S.; Taga, Y.; Ozer, N. Resveratrol: French paradox revisited. Front. Pharm. 2012, 3, 141. [Google Scholar] [CrossRef] [Green Version]
- Baur, J.A.; Sinclair, D.A. Therapeutic potential of resveratrol: The in vivo evidence. Nat. Rev. Drug Discov. 2006, 5, 493–506. [Google Scholar] [CrossRef]
- Rauf, A.; Imran, M.; Suleria, H.A.R.; Ahmad, B.; Peters, D.G.; Mubarak, M.S. A comprehensive review of the health perspectives of resveratrol. Food Funct. 2017, 8, 4284–4305. [Google Scholar] [CrossRef] [PubMed]
- Walle, T. Bioavailability of resveratrol. Ann. N. Y. Acad. Sci. 2011, 1215, 9–15. [Google Scholar] [CrossRef]
- Bourque, S.L.; Dolinsky, V.W.; Dyck, J.R.; Davidge, S.T. Maternal resveratrol treatment during pregnancy improves adverse fetal outcomes in a rat model of severe hypoxia. Placenta 2012, 33, 449–452. [Google Scholar] [CrossRef]
- Williams, L.D.; Burdock, G.A.; Edwards, J.A.; Beck, M.; Bausch, J. Safety studies conducted on high-purity trans-resveratrol in experimental animals. Food Chem. Toxicol. 2009, 47, 2170–2182. [Google Scholar] [CrossRef]
- Yu, C.; Shin, Y.G.; Chow, A.; Li, Y.; Kosmeder, J.W.; Lee, Y.S.; Hirschelman, W.H.; Pezzuto, J.M.; Mehta, R.G.; van Breemen, R.B. Human, rat, and mouse metabolism of resveratrol. Pharm. Res. 2002, 19, 1907–1914. [Google Scholar] [CrossRef]
- Raval, A.P.; Dave, K.R.; Pérez-Pinzón, M.A. Resveratrol mimics ischemic preconditioning in the brain. J. Cereb. Blood Flow Metab. 2006, 26, 1141–1147. [Google Scholar] [CrossRef] [Green Version]
- Zhao, B.; Hall, C.A. Composition and antioxidant activity of raisin extracts obtained from various solvents. Food Chem. 2008, 108, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Karalis, F.; Soubasi, V.; Georgiou, T.; Nakas, C.T.; Simeonidou, C.; Guiba-Tziampiri, O.; Spandou, E. Resveratrol ameliorates hypoxia/ischemia-induced behavioral deficits and brain injury in the neonatal rat brain. Brain Res. 2011, 1425, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Bian, H.; Shan, H.; Chen, T. Resveratrol ameliorates hypoxia/ischemia-induced brain injury in the neonatal rat via the miR-96/Bax axis. Childs Nerv. Syst. 2017, 33, 1937–1945. [Google Scholar] [CrossRef] [PubMed]
- Girbovan, C.; Kent, P.; Merali, Z.; Plamondon, H. Dose-related effects of chronic resveratrol administration on neurogenesis, angiogenesis, and corticosterone secretion are associated with improved spatial memory retention following global cerebral ischemia. Nutr. Neurosci. 2016, 19, 352–368. [Google Scholar] [CrossRef] [PubMed]
- Morin, C.; Zini, R.; Albengres, E.; Bertelli, A.A.; Bertelli, A.; Tillement, J.P. Evidence for resveratrol-induced preservation of brain mitochondria functions after hypoxia-reoxygenation. Drugs Exp. Clin. Res. 2003, 29, 227–233. [Google Scholar] [PubMed]
- Piotrowska, H.; Kucinska, M.; Murias, M. Biological activity of piceatannol: Leaving the shadow of resveratrol. Mutat. Res. 2012, 750, 60–82. [Google Scholar] [CrossRef]
- Potter, G.A.; Patterson, L.H.; Wanogho, E.; Perry, P.J.; Butler, P.C.; Ijaz, T.; Ruparelia, K.C.; Lamb, J.H.; Farmer, P.B.; Stanley, L.A.; et al. The cancer preventative agent resveratrol is converted to the anticancer agent piceatannol by the cytochrome P450 enzyme CYP1B1. Br. J. Cancer 2002, 86, 774–778. [Google Scholar] [CrossRef] [PubMed]
- Agundez, J.A. Cytochrome P450 gene polymorphism and cancer. Curr. Drug Metab. 2004, 5, 211–224. [Google Scholar] [CrossRef]
- Setoguchi, Y.; Oritani, Y.; Ito, R.; Inagaki, H.; Maruki-Uchida, H.; Ichiyanagi, T.; Ito, T. Absorption and metabolism of piceatannol in rats. J. Agric. Food Chem. 2014, 62, 2541–2548. [Google Scholar] [CrossRef] [PubMed]
- Murias, M.; Jager, W.; Handler, N.; Erker, T.; Horvath, Z.; Szekeres, T.; Nohl, H.; Gille, L. Antioxidant, prooxidant and cytotoxic activity of hydroxylated resveratrol analogues: Structure-activity relationship. Biochem. Pharm. 2005, 69, 903–912. [Google Scholar] [CrossRef] [PubMed]
- Kosuru, R.; Rai, U.; Prakash, S.; Singh, A.; Singh, S. Promising therapeutic potential of pterostilbene and its mechanistic insight based on preclinical evidence. Eur. J. Pharm. 2016, 789, 229–243. [Google Scholar] [CrossRef] [PubMed]
- McCormack, D.; McFadden, D. A review of pterostilbene antioxidant activity and disease modification. Oxid. Med. Cell Longev. 2013, 2013, 575482. [Google Scholar] [CrossRef] [Green Version]
- Kapetanovic, I.M.; Muzzio, M.; Huang, Z.; Thompson, T.N.; McCormick, D.L. Pharmacokinetics, oral bioavailability, and metabolic profile of resveratrol and its dimethylether analog, pterostilbene, in rats. Cancer Chemother. Pharm. 2011, 68, 593–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, H.S.; Yue, B.D.; Ho, P.C. Determination of pterostilbene in rat plasma by a simple HPLC-UV method and its application in pre-clinical pharmacokinetic study. Biomed. Chromatogr. 2009, 23, 1308–1315. [Google Scholar] [CrossRef]
- Hertog, M.G.; Kromhout, D.; Aravanis, C.; Blackburn, H.; Buzina, R.; Fidanza, F.; Giampaoli, S.; Jansen, A.; Menotti, A.; Nedeljkovic, S.; et al. Flavonoid intake and long-term risk of coronary heart disease and cancer in the seven countries study. Arch. Intern. Med. 1995, 155, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Hertog, M.G.L.; Hollman, P.C.H.; Katan, M.B. Content of potentially anticarcinogenic flavonoids of 28 vegetables and 9 fruits commonly consumed in the Netherlands. J. Agric. Food Chem. 1992, 40, 2379–2383. [Google Scholar] [CrossRef]
- Hertog, M.G.L.; Hollman, P.C.H.; van de Putte, B. Content of potentially anticarcinogenic flavonoids of tea infusions, wines, and fruit juices. J. Agric. Food Chem. 1993, 41, 1242–1246. [Google Scholar] [CrossRef]
- Dajas, F. Life or death: Neuroprotective and anticancer effects of quercetin. J. Ethnopharmacol. 2012, 143, 383–396. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.K.; Patnaik, R.; Muresanu, D.F.; Sharma, A.; Sharma, H.S. Quercetin in hypoxia-induced oxidative stress: Novel target for neuroprotection. Int. Rev. Neurobiol. 2012, 102, 107–146. [Google Scholar] [CrossRef]
- Russo, M.; Spagnuolo, C.; Tedesco, I.; Bilotto, S.; Russo, G.L. The flavonoid quercetin in disease prevention and therapy: Facts and fancies. Biochem. Pharm. 2012, 83, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Dajas, F.; Rivera, F.; Blasina, F.; Arredondo, F.; Echeverry, C.; Lafon, L.; Morquio, A.; Heinzen, H. Cell culture protection and in vivo neuroprotective capacity of flavonoids. Neurotox. Res. 2003, 5, 425–432. [Google Scholar] [CrossRef]
- Rivera, F.; Urbanavicius, J.; Gervaz, E.; Morquio, A.; Dajas, F. Some aspects of the in vivo neuroprotective capacity of flavonoids: Bioavailability and structure-activity relationship. Neurotox. Res. 2004, 6, 543–553. [Google Scholar] [CrossRef]
- Yao, R.Q.; Qi, D.S.; Yu, H.L.; Liu, J.; Yang, L.H.; Wu, X.X. Quercetin attenuates cell apoptosis in focal cerebral ischemia rat brain via activation of BDNF-TrkB-PI3K/Akt signaling pathway. Neurochem. Res. 2012, 37, 2777–2786. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Chang, C.Y.; Lin, S.Y.; Wang, J.D.; Wu, C.C.; Chen, W.Y.; Kuan, Y.H.; Liao, S.L.; Wang, W.Y.; Chen, C.J. Quercetin protects against cerebral ischemia/reperfusion and oxygen glucose deprivation/reoxygenation neurotoxicity. J. Nutr. Biochem. 2020, 83, 108436. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Q.; Yao, R.Q.; Liu, X.; Huang, J.J.; Qi, D.S.; Yang, L.H. Quercetin protects oligodendrocyte precursor cells from oxygen/glucose deprivation injury in vitro via the activation of the PI3K/Akt signaling pathway. Brain Res. Bull. 2011, 86, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Le, K.; Song, Z.; Deng, J.; Peng, X.; Zhang, J.; Wang, L.; Zhou, L.; Bi, H.; Liao, Z.; Feng, Z. Quercetin alleviates neonatal hypoxic-ischemic brain injury by inhibiting microglia-derived oxidative stress and TLR4-mediated inflammation. Inflamm. Res. 2020, 69, 1201–1213. [Google Scholar] [CrossRef]
- Ossola, B.; Kääriäinen, T.M.; Männistö, P.T. The multiple faces of quercetin in neuroprotection. Expert Opin. Drug Saf. 2009, 8, 397–409. [Google Scholar] [CrossRef]
- Huang, J.J.; Liu, X.; Wang, X.Q.; Yang, L.H.; Qi, D.S.; Yao, R.Q. Effects of quercetin on the learning and memory ability of neonatal rats with hypoxic-ischemic brain damage. Zhongguo Dang Dai. Er. Ke. Za. Zhi. 2012, 14, 454–457. [Google Scholar] [PubMed]
- Qu, X.; Qi, D.; Dong, F.; Wang, B.; Guo, R.; Luo, M.; Yao, R. Quercetin improves hypoxia-ischemia induced cognitive deficits via promoting remyelination in neonatal rat. Brain Res. 2014, 1553, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Priprem, A.; Watanatorn, J.; Sutthiparinyanont, S.; Phachonpai, W.; Muchimapura, S. Anxiety and cognitive effects of quercetin liposomes in rats. Nanomedicine 2008, 4, 70–78. [Google Scholar] [CrossRef]
- Masibo, M.; He, Q. Major mango polyphenols and their potential significance to human health. Compr. Rev. Food Sci. Food Saf. 2008, 7, 309–319. [Google Scholar] [CrossRef]
- Luo, F.; Lv, Q.; Zhao, Y.; Hu, G.; Huang, G.; Zhang, J.; Sun, C.; Li, X.; Chen, K. Quantification and purification of mangiferin from Chinese mango (Mangifera indica L.) cultivars and its protective effect on human umbilical vein endothelial cells under H2O2-induced stress. Int. J. Mol. Sci. 2012, 13, 11260–11274. [Google Scholar] [CrossRef] [PubMed]
- Du, S.; Liu, H.; Lei, T.; Xie, X.; Wang, H.; He, X.; Tong, R.; Wang, Y. Mangiferin: An effective therapeutic agent against several disorders (review). Mol. Med. Rep. 2018, 18, 4775–4786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittal, S.; Iqubal, M.K.; Iqbal, B.; Gupta, M.M.; Ali, J.; Baboota, S. A pervasive scientific overview on mangiferin in the prevention and treatment of various diseases with preclinical and clinical updates. J. Complement. Integr. Med. 2020. [Google Scholar] [CrossRef]
- Sosunov, S.A.; Ameer, X.; Niatsetskaya, Z.V.; Utkina-Sosunova, I.; Ratner, V.I.; Ten, V.S. Isoflurane anesthesia initiated at the onset of reperfusion attenuates oxidative and hypoxic-ischemic brain injury. PLoS ONE 2015, 10, e0120456. [Google Scholar] [CrossRef]
- Xu, Y.; Xue, H.; Zhao, P.; Yang, Y.; Ji, G.; Yu, W.; Han, G.; Ding, M.; Wang, F. Isoflurane postconditioning induces concentration- and timing-dependent neuroprotection partly mediated by the GluR2 AMPA receptor in neonatal rats after brain hypoxia-ischemia. J. Anesth. 2016, 30, 427–436. [Google Scholar] [CrossRef]
- Zhao, P.; Peng, L.; Li, L.; Xu, X.; Zuo, Z. Isoflurane preconditioning improves long-term neurologic outcome after hypoxic-ischemic brain injury in neonatal rats. Anesthesiology 2007, 107, 963–970. [Google Scholar] [CrossRef] [Green Version]
- Fahmy, H.; Hegazi, N.; El-Shamy, S.; Farag, M.A. Pomegranate juice as a functional food: A comprehensive review of its polyphenols, therapeutic merits, and recent patents. Food Funct. 2020, 11, 5768–5781. [Google Scholar] [CrossRef] [PubMed]
- Vučić, V.; Grabež, M.; Trchounian, A.; Arsić, A. Composition and potential health benefits of pomegranate: A review. Curr. Pharm. Des. 2019, 25, 1817–1827. [Google Scholar] [CrossRef] [PubMed]
- Forouzanfar, F.; Afkhami Goli, A.; Asadpour, E.; Ghorbani, A.; Sadeghnia, H.R. Protective effect of Punica granatum L. against serum/glucose deprivation-induced PC12 cells injury. Evid. Based Complement. Altern. Med. 2013, 2013, 716730. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.A.; El Morsy, E.M.; Ahmed, A.A. Pomegranate extract protects against cerebral ischemia/reperfusion injury and preserves brain DNA integrity in rats. Life Sci. 2014, 110, 61–69. [Google Scholar] [CrossRef]
- Sarkaki, A.; Rezaiei, M.; Gharib Naseri, M.; Rafieirad, M. Improving active and passive avoidance memories deficits due to permanent cerebral ischemia by pomegranate seed extract in female rats. Malays. J. Med. Sci. 2013, 20, 25–34. [Google Scholar]
- Bellone, J.A.; Murray, J.R.; Jorge, P.; Fogel, T.G.; Kim, M.; Wallace, D.R.; Hartman, R.E. Pomegranate supplementation improves cognitive and functional recovery following ischemic stroke: A randomized trial. Nutr. Neurosci. 2019, 22, 738–743. [Google Scholar] [CrossRef]
- Miller, S.L.; Huppi, P.S.; Mallard, C. The consequences of fetal growth restriction on brain structure and neurodevelopmental outcome. J. Physiol. 2016, 594, 807–823. [Google Scholar] [CrossRef] [Green Version]
- Rauf, A.; Imran, M.; Abu-Izneid, T.; Iahtisham Ul, H.; Patel, S.; Pan, X.; Naz, S.; Sanches Silva, A.; Saeed, F.; Rasul Suleria, H.A. Proanthocyanidins: A comprehensive review. Biomed. Pharm. 2019, 116, 108999. [Google Scholar] [CrossRef]
- Kong, X.; Guan, J.; Gong, S.; Wang, R. Neuroprotective effects of grape seed procyanidin extract on ischemia-reperfusion brain injury. Chin. Med. Sci. J. 2017, 32, 92–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.; Liu, Y.M.; Fratkins, J.D.; LeBlanc, M.H. Grape seed extract suppresses lipid peroxidation and reduces hypoxic ischemic brain injury in neonatal rats. Brain Res. Bull. 2005, 66, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Liu, Y.M.; Leblanc, M.H.; Bhatt, A.J.; Rhodes, P.G. Grape seed extract given three hours after injury suppresses lipid peroxidation and reduces hypoxic-ischemic brain injury in neonatal rats. Pediatr. Res. 2007, 61, 295–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arola-Arnal, A.; Oms-Oliu, G.; Crescenti, A.; del Bas, J.M.; Ras, M.R.; Arola, L.; Caimari, A. Distribution of grape seed flavanols and their metabolites in pregnant rats and their fetuses. Mol. Nutr. Food Res. 2013, 57, 1741–1752. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Wang, H.; Hua, X.; Chen, D.; Huang, C.; Chen, Z. An outline for the pharmacological effect of icariin in the nervous system. Eur. J. Pharm. 2019, 842, 20–32. [Google Scholar] [CrossRef]
- He, C.; Wang, Z.; Shi, J. Pharmacological effects of icariin. Adv. Pharm. 2020, 87, 179–203. [Google Scholar] [CrossRef]
- Xiong, D.; Deng, Y.; Huang, B.; Yin, C.; Liu, B.; Shi, J.; Gong, Q. Icariin attenuates cerebral ischemia-reperfusion injury through inhibition of inflammatory response mediated by NF-κB, PPARα and PPARγ in rats. Int. Immunopharmacol. 2016, 30, 157–162. [Google Scholar] [CrossRef]
- Mo, Z.T.; Li, W.N.; Zhai, Y.R.; Gong, Q.H. Icariin attenuates OGD/R-induced autophagy via Bcl-2-dependent cross talk between apoptosis and autophagy in PC12 cells. Evid. Based Complement. Altern. Med. 2016, 2016, 4343084. [Google Scholar] [CrossRef] [Green Version]
- Han, S.; Li, L.Z.; Song, S.J. Daphne giraldii Nitsche (Thymelaeaceae): Phytochemistry, pharmacology and medicinal uses. Phytochemistry 2020, 171, 112231. [Google Scholar] [CrossRef]
- Zhang, W.; Shen, Y.; Liu, R.; Zhang, C.; Chen, H.; Fu, P.; Shan, L.; Zhang, W. Coumarins from the bark of Daphne marginata. Chem. Nat. Compd. 2007, 43, 317–318. [Google Scholar] [CrossRef]
- Fylaktakidou, K.C.; Hadjipavlou-Litina, D.J.; Litinas, K.E.; Nicolaides, D.N. Natural and synthetic coumarin derivatives with anti-inflammatory/antioxidant activities. Curr. Pharm. Des. 2004, 10, 3813–3833. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Liu, Q.; Zhou, J.; Tan, G.; Deng, X.; Ci, X. Daphnetin-mediated Nrf2 antioxidant signaling pathways ameliorate tert-butyl hydroperoxide (t-BHP)-induced mitochondrial dysfunction and cell death. Free Radic. Biol. Med. 2017, 106, 38–52. [Google Scholar] [CrossRef] [Green Version]
- Tu, L.; Li, S.; Fu, Y.; Yao, R.; Zhang, Z.; Yang, S.; Zeng, X.; Kuang, N. The therapeutic effects of daphnetin in collagen-induced arthritis involve its regulation of Th17 cells. Int. Immunopharmacol. 2012, 13, 417–423. [Google Scholar] [CrossRef]
- Wang, Q.M.; Ni, Y.C.; Xu, Y.Q.; Ha, S.H.; Cai, Y. The schizontocidal activity of daphnetin against malaria parasites in vitro and in vivo. Zhongguo Ji Sheng Chong Xue Yu Ji Sheng Chong Bing Za Zhi 2000, 18, 204–206. [Google Scholar] [PubMed]
- Liu, J.; Chen, Q.; Jian, Z.; Xiong, X.; Shao, L.; Jin, T.; Zhu, X.; Wang, L. Daphnetin protects against cerebral ischemia/reperfusion injury in mice via inhibition of TLR4/NF-κB signaling pathway. Biomed. Res. Int. 2016, 2016, 2816056. [Google Scholar] [CrossRef] [Green Version]
- Zhi, J.; Duan, B.; Pei, J.; Wu, S.; Wei, J. Daphnetin protects hippocampal neurons from oxygen-glucose deprivation-induced injury. J. Cell Biochem. 2019, 120, 4132–4139. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Tian, J.; He, W.; Xie, J.; Hu, Z.; Chen, X. Spectrofluorimetric study of the binding of daphnetin to bovine serum albumin. J. Pharm. Biomed. Anal. 2004, 35, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Qu, S.-Y.; Jiang, X.-L.; Zhao, X.-H.; Pan, D.-B.; Wang, X.-R.; Chen, Y.-L.; Pan, G.; Wu, Y.-J. Antithrombotic effect of daphnetin in the rat. Yao Xue Xue Bao 1986, 21, 498–501. [Google Scholar] [PubMed]
- Scorletti, E.; Byrne, C.D. Omega-3 fatty acids, hepatic lipid metabolism, and nonalcoholic fatty liver disease. Annu. Rev. Nutr. 2013, 33, 231–248. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, J. Docosahexaenoic acid (DHA): An ancient nutrient for the modern human brain. Nutrients 2011, 3, 529–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uauy, R.; Hoffman, D.R.; Peirano, P.; Birch, D.G.; Birch, E.E. Essential fatty acids in visual and brain development. Lipids 2001, 36, 885–895. [Google Scholar] [CrossRef] [PubMed]
- Kidd, P.M. Omega-3 DHA and EPA for cognition, behavior, and mood: Clinical findings and structural-functional synergies with cell membrane phospholipids. Altern. Med. Rev. 2007, 12, 207–227. [Google Scholar]
- Salem, N., Jr.; Litman, B.; Kim, H.Y.; Gawrisch, K. Mechanisms of action of docosahexaenoic acid in the nervous system. Lipids 2001, 36, 945–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fedorova, I.; Hussein, N.; Di Martino, C.; Moriguchi, T.; Hoshiba, J.; Majchrzak, S.; Salem, N., Jr. An n-3 fatty acid deficient diet affects mouse spatial learning in the Barnes circular maze. Prostaglandins Leukot Essent Fat. Acids 2007, 77, 269–277. [Google Scholar] [CrossRef] [Green Version]
- Wurtman, R.J. Synapse formation and cognitive brain development: Effect of docosahexaenoic acid and other dietary constituents. Metabolism 2008, 57 (Suppl. 2), S6–S10. [Google Scholar] [CrossRef] [Green Version]
- Heurteaux, C.; Laigle, C.; Blondeau, N.; Jarretou, G.; Lazdunski, M. Alpha-linolenic acid and riluzole treatment confer cerebral protection and improve survival after focal brain ischemia. Neuroscience 2006, 137, 241–251. [Google Scholar] [CrossRef]
- Blondeau, N.; Nguemeni, C.; Debruyne, D.N.; Piens, M.; Wu, X.; Pan, H.; Hu, X.; Gandin, C.; Lipsky, R.H.; Plumier, J.C.; et al. Subchronic alpha-linolenic acid treatment enhances brain plasticity and exerts an antidepressant effect: A versatile potential therapy for stroke. Neuropsychopharmacology 2009, 34, 2548–2559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourourou, M.; Heurteaux, C.; Blondeau, N. Alpha-linolenic acid given as enteral or parenteral nutritional intervention against sensorimotor and cognitive deficits in a mouse model of ischemic stroke. Neuropharmacology 2016, 108, 60–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguemeni, C.; Delplanque, B.; Rovere, C.; Simon-Rousseau, N.; Gandin, C.; Agnani, G.; Nahon, J.L.; Heurteaux, C.; Blondeau, N. Dietary supplementation of alpha-linolenic acid in an enriched rapeseed oil diet protects from stroke. Pharm. Res. 2010, 61, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Ueda, M.; Inaba, T.; Nito, C.; Kamiya, N.; Katayama, Y. Therapeutic impact of eicosapentaenoic acid on ischemic brain damage following transient focal cerebral ischemia in rats. Brain Res. 2013, 1519, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Okabe, N.; Nakamura, T.; Toyoshima, T.; Miyamoto, O.; Lu, F.; Itano, T. Eicosapentaenoic acid prevents memory impairment after ischemia by inhibiting inflammatory response and oxidative damage. J. Stroke Cereb. Dis. 2011, 20, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.-C.; Kao, T.-K.; Ou, Y.-C.; Yang, D.-Y.; Yen, Y.-J.; Wang, C.-C.; Chuang, Y.-H.; Liao, S.-L.; Raung, S.-L.; Wu, C.-W.; et al. Protective effect of docosahexaenoic acid against brain injury in ischemic rats. J. Nutr Biochem. 2009, 20, 715–725. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.-H.; Khoutorova, L.; Bazan, N.G.; Belayev, L. Docosahexaenoic acid improves behavior and attenuates blood-brain barrier injury induced by focal cerebral ischemia in rats. Exp. Transl. Stroke Med. 2015, 7, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eady, T.N.; Khoutorova, L.; Obenaus, A.; Mohd-Yusof, A.; Bazan, N.G.; Belayev, L. Docosahexaenoic acid complexed to albumin provides neuroprotection after experimental stroke in aged rats. Neurobiol. Dis. 2014, 62, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belayev, L.; Khoutorova, L.; Atkins, K.D.; Bazan, N.G. Robust docosahexaenoic acid-mediated neuroprotection in a rat model of transient, focal cerebral ischemia. Stroke 2009, 40, 3121–3126. [Google Scholar] [CrossRef]
- Arteaga, O.; Alvarez, A.; Revuelta, M.; Santaolalla, F.; Urtasun, A.; Hilario, E. Role of antioxidants in neonatal hypoxic-ischemic brain injury: New therapeutic approaches. Int. J. Mol. Sci. 2017, 18, 265. [Google Scholar] [CrossRef] [Green Version]
- Tanumihardjo, S.A.; Russell, R.M.; Stephensen, C.B.; Gannon, B.M.; Craft, N.E.; Haskell, M.J.; Lietz, G.; Schulze, K.; Raiten, D.J. Biomarkers of nutrition for development (BOND)—Vitamin A review. J. Nutr. 2016, 146, 1816S–1848S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maden, M. Retinoic acid in the development, regeneration and maintenance of the nervous system. Nat. Rev. Neurosci. 2007, 8, 755–765. [Google Scholar] [CrossRef]
- Ghyselinck, N.B.; Duester, G. Retinoic acid signaling pathways. Development 2019, 146, dev167502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, W.; Wen, E.Y.; Gong, M.; Shi, Y.; Chen, L.; Bi, Y.; Zhang, Y.; Liu, Y.F.; Chen, J.; Qu, P.; et al. The pattern of retinoic acid receptor expression and subcellular, anatomic and functional area translocation during the postnatal development of the rat cerebral cortex and white matter. Brain Res. 2011, 1382, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Imbard, A.; Benoist, J.F.; Blom, H.J. Neural tube defects, folic acid and methylation. Int. J. Environ. Res. Public Health 2013, 10, 4352–4389. [Google Scholar] [CrossRef] [Green Version]
- Fenech, M. The role of folic acid and vitamin B12 in genomic stability of human cells. Mutat. Res. 2001, 475, 57–67. [Google Scholar] [CrossRef]
- Mattson, M.P.; Shea, T.B. Folate and homocysteine metabolism in neural plasticity and neurodegenerative disorders. Trends Neurosci. 2003, 26, 137–146. [Google Scholar] [CrossRef]
- Iskandar, B.J.; Nelson, A.; Resnick, D.; Skene, J.H.; Gao, P.; Johnson, C.; Cook, T.D.; Hariharan, N. Folic acid supplementation enhances repair of the adult central nervous system. Ann. Neurol. 2004, 56, 221–227. [Google Scholar] [CrossRef]
- Van Guelpen, B.; Hultdin, J.; Johansson, I.; Stegmayr, B.; Hallmans, G.; Nilsson, T.K.; Weinehall, L.; Witthöft, C.; Palmqvist, R.; Winkvist, A. Folate, vitamin B12, and risk of ischemic and hemorrhagic stroke: A prospective, nested case-referent study of plasma concentrations and dietary intake. Stroke 2005, 36, 1426–1431. [Google Scholar] [CrossRef] [Green Version]
- Deniz, B.F.; Confortim, H.D.; Deckmann, I.; Miguel, P.M.; Bronauth, L.; de Oliveira, B.C.; Vieira, M.C.; Dos Santos, T.M.; Bertó, C.G.; Hartwig, J.; et al. Gestational folic acid supplementation does not affects the maternal behavior and the early development of rats submitted to neonatal hypoxia-ischemia but the high supplementation impairs the dam’s memory and the Na+,K+-ATPase activity in the pup’s hippocampus. Int. J. Dev. Neurosci. 2018, 71, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Carletti, J.V.; Deniz, B.F.; Miguel, P.M.; Rojas, J.J.; Kolling, J.; Scherer, E.B.; de Souza Wyse, A.T.; Netto, C.A.; Pereira, L.O. Folic acid prevents behavioral impairment and Na+,K+-ATPase inhibition caused by neonatal hypoxia-ischemia. Neurochem. Res. 2012, 37, 1624–1630. [Google Scholar] [CrossRef] [PubMed]
- Cesari, M.; Incalzi, R.A.; Zamboni, V.; Pahor, M. Vitamin D hormone: A multitude of actions potentially influencing the physical function decline in older persons. Geriatr. Gerontol. Int. 2011, 11, 133–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holick, M.F. Vitamin D deficiency. N. Engl. J. Med. 2007, 357, 266–281. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.B.; Lin, A.M.; Chiu, T.H. Systemic vitamin D3 attenuated oxidative injuries in the locus coeruleus of rat brain. Ann. N. Y. Acad. Sci. 2003, 993, 313–324, discussion 345–319. [Google Scholar] [CrossRef] [PubMed]
- Thota, C.; Farmer, T.; Garfield, R.E.; Menon, R.; Al-Hendy, A. Vitamin D elicits anti-inflammatory response, inhibits contractile-associated proteins, and modulates Toll-like receptors in human myometrial cells. Reprod. Sci. 2013, 20, 463–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mutlu, M.; Sarıaydın, M.; Aslan, Y.; Kader, Ş.; Dereci, S.; Kart, C.; Yaman, S.; Kural, B. Status of vitamin D, antioxidant enzymes, and antioxidant substances in neonates with neonatal hypoxic-ischemic encephalopathy. J. Matern. Fetal Neonatal. Med. 2016, 29, 2259–2263. [Google Scholar] [CrossRef]
- Galli, F.; Azzi, A.; Birringer, M.; Cook-Mills, J.M.; Eggersdorfer, M.; Frank, J.; Cruciani, G.; Lorkowski, S.; Özer, N.K. Vitamin E: Emerging aspects and new directions. Free Radic. Biol. Med. 2017, 102, 16–36. [Google Scholar] [CrossRef] [PubMed]
- Sozen, E.; Demirel, T.; Ozer, N.K. Vitamin E: Regulatory role in the cardiovascular system. Iubmb. Life 2019, 71, 507–515. [Google Scholar] [CrossRef]
- Yang, C.S.; Luo, P.; Zeng, Z.; Wang, H.; Malafa, M.; Suh, N. Vitamin E and cancer prevention: Studies with different forms of tocopherols and tocotrienols. Mol. Carcinog. 2020, 59, 365–389. [Google Scholar] [CrossRef]
- Ulatowski, L.M.; Manor, D. Vitamin E and neurodegeneration. Neurobiol. Dis. 2015, 84, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Browne, D.; McGuinness, B.; Woodside, J.V.; McKay, G.J. Vitamin E and Alzheimer’s disease: What do we know so far? Clin. Interv. Aging 2019, 14, 1303–1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohsenpour, H.; Pesce, M.; Patruno, A.; Bahrami, A.; Pour, P.M.; Farzaei, M.H. A review of plant extracts and plant-derived natural compounds in the prevention/treatment of neonatal hypoxic-ischemic brain injury. Int. J. Mol. Sci 2021, 22, 833. [Google Scholar] [CrossRef]
- Park, O.K.; Choi, J.H.; Park, J.H.; Kim, I.H.; Yan, B.C.; Ahn, J.H.; Kwon, S.H.; Lee, J.C.; Kim, Y.S.; Kim, M.; et al. Comparison of neuroprotective effects of five major lipophilic diterpenoids from Danshen extract against experimentally induced transient cerebral ischemic damage. Fitoterapia 2012, 83, 1666–1674. [Google Scholar] [CrossRef]
- Gu, M.; Zhang, G.; Su, Z.; Ouyang, F. Identification of major active constituents in the fingerprint of Salvia miltiorrhiza Bunge developed by high-speed counter-current chromatography. J. Chromatogr. A 2004, 1041, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Houghton, P.J.; Hider, R.C.; Howes, M.J. Novel diterpenoid acetylcholinesterase inhibitors from Salvia Miltiorhiza. Planta Med. 2004, 70, 201–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chong, C.M.; Su, H.; Lu, J.J.; Wang, Y. The effects of bioactive components from the rhizome of Salvia Miltiorrhiza (Danshen) on the characteristics of Alzheimer’s disease. Chin. Med. 2019, 14, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.C.; Park, J.H.; Park, O.K.; Kim, I.H.; Yan, B.C.; Ahn, J.H.; Kwon, S.H.; Choi, J.H.; Kim, J.D.; Won, M.H. Neuroprotective effects of tanshinone I from Danshen extract in a mouse model of hypoxia-ischemia. Anat. Cell Biol. 2013, 46, 183–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Zhang, B.; Yin, L.; Cai, B.; Shan, H.; Zhang, L.; Lu, Y.; Bi, Z. Tanshinone IIA prevented brain iron dyshomeostasis in cerebral ischemic rats. Cell Physiol. Biochem. 2011, 27, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.; Han, R.; Xiao, H.; Shen, J.; Luo, Q.; Li, J. Neuroprotective effects of tanshinone IIA and/or tetramethylpyrazine in cerebral ischemic injury in vivo and in vitro. Brain Res. 2012, 1488, 81–91. [Google Scholar] [CrossRef]
- Fang, C.; Xie, L.; Liu, C.; Fu, C.; Ye, W.; Liu, H.; Zhang, B. Tanshinone IIA improves hypoxic ischemic encephalopathy through TLR-4-mediated NF-κB signal pathway. Mol. Med. Rep. 2018, 18, 1899–1908. [Google Scholar] [CrossRef] [Green Version]
- Hei, M.; Luo, Y.; Zhang, X.; Liu, F. Tanshinone IIa alleviates the biochemical changes associated with hypoxic ischemic brain damage in a rat model. Phytother. Res. 2011, 25, 1865–1869. [Google Scholar] [CrossRef]
- Juge, N.; Mithen, R.F.; Traka, M. Molecular basis for chemoprevention by sulforaphane: A comprehensive review. Cell Mol. Life Sci. 2007, 64, 1105–1127. [Google Scholar] [CrossRef] [PubMed]
- Klomparens, E.A.; Ding, Y. The neuroprotective mechanisms and effects of sulforaphane. Brain Circ. 2019, 5, 74–83. [Google Scholar] [CrossRef]
- Townsend, B.E.; Johnson, R.W. Sulforaphane induces Nrf2 target genes and attenuates inflammatory gene expression in microglia from brain of young adult and aged mice. Exp. Gerontol. 2016, 73, 42–48. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.M.; Zheng, Z.H.; Fan, S.H.; Zhang, Z.F.; Chen, G.Q.; Lu, J. Sulforaphane administration alleviates diffuse axonal injury (DAI) via regulation signaling pathway of NRF2 and HO-1. J. Cell Biochem. 2020, 121, 430–442. [Google Scholar] [CrossRef]
- Shinkai, Y.; Sumi, D.; Fukami, I.; Ishii, T.; Kumagai, Y. Sulforaphane, an activator of Nrf2, suppresses cellular accumulation of arsenic and its cytotoxicity in primary mouse hepatocytes. FEBS Lett. 2006, 580, 1771–1774. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Kobori, N.; Aronowski, J.; Dash, P.K. Sulforaphane reduces infarct volume following focal cerebral ischemia in rodents. Neurosci. Lett. 2006, 393, 108–112. [Google Scholar] [CrossRef]
- Yu, C.; He, Q.; Zheng, J.; Li, L.Y.; Hou, Y.H.; Song, F.Z. Sulforaphane improves outcomes and slows cerebral ischemic/reperfusion injury via inhibition of NLRP3 inflammasome activation in rats. Int. Immunopharmacol. 2017, 45, 74–78. [Google Scholar] [CrossRef]
- Ma, L.L.; Xing, G.P.; Yu, Y.; Liang, H.; Yu, T.X.; Zheng, W.H.; Lai, T.B. Sulforaphane exerts neuroprotective effects via suppression of the inflammatory response in a rat model of focal cerebral ischemia. Int. J. Clin. Exp. Med. 2015, 8, 17811–17817. [Google Scholar] [PubMed]
- Wang, B.; Kulikowicz, E.; Lee, J.K.; Koehler, R.C.; Yang, Z.J. Sulforaphane protects piglet brains from neonatal hypoxic-ischemic injury. Dev. Neurosci. 2020, 42, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Caterina, M.J. Transient receptor potential ion channels as participants in thermosensation and thermoregulation. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 292, R64–R76. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Zheng, J. Understand spiciness: Mechanism of TRPV1 channel activation by capsaicin. Protein. Cell 2017, 8, 169–177. [Google Scholar] [CrossRef] [Green Version]
- Dairam, A.; Fogel, R.; Daya, S.; Limson, J.L. Antioxidant and iron-binding properties of curcumin, capsaicin, and S-allylcysteine reduce oxidative stress in rat brain homogenate. J. Agric. Food Chem. 2008, 56, 3350–3356. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.Y.; Yang, G.P.; Jiang, D.J.; Wang, F.; Song, T.; Tan, X.H.; Sun, Z.Q. Protection of capsaicin against hypoxia-reoxygenation-induced apoptosis of rat hippocampal neurons. Can. J. Physiol. Pharm. 2008, 86, 785–792. [Google Scholar] [CrossRef] [PubMed]
- Luqman, S.; Meena, A.; Marler, L.E.; Kondratyuk, T.P.; Pezzuto, J.M. Suppression of tumor necrosis factor-α-induced nuclear factor κB activation and aromatase activity by capsaicin and its analog capsazepine. J. Med. Food 2011, 14, 1344–1351. [Google Scholar] [CrossRef] [Green Version]
- Pegorini, S.; Braida, D.; Verzoni, C.; Guerini-Rocco, C.; Consalez, G.G.; Croci, L.; Sala, M. Capsaicin exhibits neuroprotective effects in a model of transient global cerebral ischemia in Mongolian gerbils. Br. J. Pharm. 2005, 144, 727–735. [Google Scholar] [CrossRef] [Green Version]
- Huang, M.; Cheng, G.; Tan, H.; Qin, R.; Zou, Y.; Wang, Y.; Zhang, Y. Capsaicin protects cortical neurons against ischemia/reperfusion injury via down-regulating NMDA receptors. Exp. Neurol. 2017, 295, 66–76. [Google Scholar] [CrossRef]
- Wu, D.; Shi, J.; Elmadhoun, O.; Duan, Y.; An, H.; Zhang, J.; He, X.; Meng, R.; Liu, X.; Ji, X.; et al. Dihydrocapsaicin (DHC) enhances the hypothermia-induced neuroprotection following ischemic stroke via PI3K/Akt regulation in rat. Brain Res. 2017, 1671, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Zhi, X.; Duan, Y.; Zhang, M.; An, H.; Wei, W.; Dong, K.; Zhang, Y.; Shi, J.; He, X.; et al. Inflammatory cytokines are involved in dihydrocapsaicin (DHC) and regional cooling infusion (RCI)-induced neuroprotection in ischemic rat. Brain Res. 2019, 1710, 173–180. [Google Scholar] [CrossRef]
- Janyou, A.; Wicha, P.; Seechamnanturakit, V.; Bumroongkit, K.; Tocharus, C.; Suksamrarn, A.; Tocharus, J. Dihydrocapsaicin-induced angiogenesis and improved functional recovery after cerebral ischemia and reperfusion in a rat model. J. Pharm. Sci. 2020, 143, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R. Antioxidative protection by melatonin: Multiplicity of mechanisms from radical detoxification to radical avoidance. Endocrine 2005, 27, 119–130. [Google Scholar] [CrossRef]
- Tan, D.X.; Manchester, L.C.; Terron, M.P.; Flores, L.J.; Reiter, R.J. One molecule, many derivatives: A never-ending interaction of melatonin with reactive oxygen and nitrogen species? J. Pineal Res. 2007, 42, 28–42. [Google Scholar] [CrossRef]
- Lee, J.G.; Woo, Y.S.; Park, S.W.; Seog, D.H.; Seo, M.K.; Bahk, W.M. The neuroprotective effects of melatonin: Possible role in the pathophysiology of neuropsychiatric disease. Brain Sci. 2019, 9, 285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomás-Zapico, C.; Coto-Montes, A. A proposed mechanism to explain the stimulatory effect of melatonin on antioxidative enzymes. J. Pineal Res. 2005, 39, 99–104. [Google Scholar] [CrossRef]
- Srinivasan, V.; Pandi-Perumal, S.R.; Cardinali, D.P.; Poeggeler, B.; Hardeland, R. Melatonin in Alzheimer’s disease and other neurodegenerative disorders. Behav. Brain Funct. 2006, 2, 15. [Google Scholar] [CrossRef] [Green Version]
- Cardinali, D.P.; Furio, A.M.; Brusco, L.I. Clinical aspects of melatonin intervention in Alzheimer’s disease progression. Curr. Neuropharmacol. 2010, 8, 218–227. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Cook, A.; Kim, J.; Baranov, S.V.; Jiang, J.; Smith, K.; Cormier, K.; Bennett, E.; Browser, R.P.; Day, A.L.; et al. Melatonin inhibits the caspase-1/cytochrome c/caspase-3 cell death pathway, inhibits MT1 receptor loss and delays disease progression in a mouse model of amyotrophic lateral sclerosis. Neurobiol. Dis. 2013, 55, 26–35. [Google Scholar] [CrossRef] [Green Version]
- Jacob, S.; Poeggeler, B.; Weishaupt, J.H.; Siren, A.L.; Hardeland, R.; Bahr, M.; Ehrenreich, H. Melatonin as a candidate compound for neuroprotection in amyotrophic lateral sclerosis (ALS): High tolerability of daily oral melatonin administration in ALS patients. J. Pineal Res. 2002, 33, 186–187. [Google Scholar] [CrossRef]
- Weishaupt, J.H.; Bartels, C.; Polking, E.; Dietrich, J.; Rohde, G.; Poeggeler, B.; Mertens, N.; Sperling, S.; Bohn, M.; Huther, G.; et al. Reduced oxidative damage in ALS by high-dose enteral melatonin treatment. J. Pineal Res. 2006, 41, 313–323. [Google Scholar] [CrossRef]
- Macleod, M.R.; O’Collins, T.; Horky, L.L.; Howells, D.W.; Donnan, G.A. Systematic review and meta-analysis of the efficacy of melatonin in experimental stroke. J. Pineal Res. 2005, 38, 35–41. [Google Scholar] [CrossRef]
- Sadanandan, N.; Cozene, B.; Cho, J.; Park, Y.J.; Saft, M.; Gonzales-Portillo, B.; Borlongan, C.V. Melatonin—A potent therapeutic for stroke and stroke-related dementia. Antioxidants 2020, 9, 672. [Google Scholar] [CrossRef]
- Hu, Y.; Wang, Z.; Pan, S.; Zhang, H.; Fang, M.; Jiang, H.; Zhang, H.; Gao, Z.; Xu, K.; Li, Z.; et al. Melatonin protects against blood-brain barrier damage by inhibiting the TLR4/ NF-κB signaling pathway after LPS treatment in neonatal rats. Oncotarget 2017, 8, 31638–31654. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.-X.; Lv, Y.; Li, Y.-H.; Ding, X.; Wang, Y.; Han, X.; Liu, M.-H.; Sun, B.; Feng, X. Melatonin alleviates brain and peripheral tissue edema in a neonatal rat model of hypoxic-ischemic brain damage: The involvement of edema related proteins. BMC Pediatrics 2017, 17, 90. [Google Scholar] [CrossRef] [Green Version]
- Carloni, S.; Albertini, M.C.; Galluzzi, L.; Buonocore, G.; Proietti, F.; Balduini, W. Melatonin reduces endoplasmic reticulum stress and preserves sirtuin 1 expression in neuronal cells of newborn rats after hypoxia-ischemia. J. Pineal Res. 2014, 57, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Ozyener, F.; Çetinkaya, M.; Alkan, T.; Gören, B.; Kafa, I.M.; Kurt, M.A.; Koksal, N. Neuroprotective effects of melatonin administered alone or in combination with topiramate in neonatal hypoxic-ischemic rat model. Restor. Neurol. Neurosci. 2012, 30, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Yawno, T.; Mahen, M.; Li, J.; Fahey, M.C.; Jenkin, G.; Miller, S.L. The beneficial effects of melatonin administration following hypoxia-ischemia in preterm fetal sheep. Front. Cell Neurosci. 2017, 11, 296. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, Q.M.; Chishti, A.L.; Waseem, N. Role of melatonin in management of hypoxic ischaemic encephalopathy in newborns: A randomized control trial. J. Pak. Med. Assoc. 2018, 68, 1233–1237. [Google Scholar]
- El Farargy, M.S.; Soliman, N.A. A randomized controlled trial on the use of magnesium sulfate and melatonin in neonatal hypoxic ischemic encephalopathy. J. Neonatal Perinat. Med. 2019, 12, 379–384. [Google Scholar] [CrossRef]
- Aly, H.; Elmahdy, H.; El-Dib, M.; Rowisha, M.; Awny, M.; El-Gohary, T.; Elbatch, M.; Hamisa, M.; El-Mashad, A.R. Melatonin use for neuroprotection in perinatal asphyxia: A randomized controlled pilot study. J. Perinatol. 2015, 35, 186–191. [Google Scholar] [CrossRef]
- Alonso-Alconada, D.; Alvarez, A.; Arteaga, O.; Martínez-Ibargüen, A.; Hilario, E. Neuroprotective effect of melatonin: A novel therapy against perinatal hypoxia-ischemia. Int. J. Mol. Sci. 2013, 14, 9379–9395. [Google Scholar] [CrossRef] [Green Version]
- D’Angelo, G.; Cannavo, L.; Reiter, R.J.; Gitto, E. Melatonin administration from 2000 to 2020 to human newborns with hypoxic-ischemic encephalopathy. Am. J. Perinatol. 2020. [Google Scholar] [CrossRef]
- Okatani, Y.; Okamoto, K.; Hayashi, K.; Wakatsuki, A.; Tamura, S.; Sagara, Y. Maternal-fetal transfer of melatonin in pregnant women near term. J. Pineal Res. 1998, 25, 129–134. [Google Scholar] [CrossRef]
- Ferreira, G.C.; McKenna, M.C. L-Carnitine and acetyl-L-carnitine roles and neuroprotection in developing brain. Neurochem. Res. 2017, 42, 1661–1675. [Google Scholar] [CrossRef]
- Cam, H.; Yildirim, B.; Aydin, A.; Say, A. Carnitine levels in neonatal hypoxia. J. Trop. Pediatr. 2005, 51, 106–108. [Google Scholar] [CrossRef]
- Tang, S.; Xu, S.; Lu, X.; Gullapalli, R.P.; McKenna, M.C.; Waddell, J. Neuroprotective effects of acetyl-L-carnitine on neonatal hypoxia ischemia-induced brain injury in rats. Dev. Neurosci. 2016, 38, 384–396. [Google Scholar] [CrossRef] [Green Version]
- Helton, E.; Darragh, R.; Francis, P.; Fricker, F.J.; Jue, K.; Koch, G.; Mair, D.; Pierpont, M.E.; Prochazka, J.V.; Linn, L.S.; et al. Metabolic aspects of myocardial disease and a role for L-carnitine in the treatment of childhood cardiomyopathy. Pediatrics 2000, 105, 1260–1270. [Google Scholar]
- Wyss, M.; Kaddurah-Daouk, R. Creatine and creatinine metabolism. Physiol. Rev. 2000, 80, 1107–1213. [Google Scholar] [CrossRef]
- Bender, A.; Klopstock, T. Creatine for neuroprotection in neurodegenerative disease: End of story? Amino Acids 2016, 48, 1929–1940. [Google Scholar] [CrossRef]
- Sullivan, P.G.; Geiger, J.D.; Mattson, M.P.; Scheff, S.W. Dietary supplement creatine protects against traumatic brain injury. Ann. Neurol. 2000, 48, 723–729. [Google Scholar] [CrossRef]
- Beal, M.F. Neuroprotective effects of creatine. Amino Acids 2011, 40, 1305–1313. [Google Scholar] [CrossRef] [PubMed]
- Sartini, S.; Lattanzi, D.; Ambrogini, P.; Di Palma, M.; Galati, C.; Savelli, D.; Polidori, E.; Calcabrini, C.; Rocchi, M.B.; Sestili, P.; et al. Maternal creatine supplementation affects the morpho-functional development of hippocampal neurons in rat offspring. Neuroscience 2016, 312, 120–129. [Google Scholar] [CrossRef]
- Sartini, S.; Lattanzi, D.; Di Palma, M.; Savelli, D.; Eusebi, S.; Sestili, P.; Cuppini, R.; Ambrogini, P. Maternal creatine supplementation positively affects male rat hippocampal synaptic plasticity in adult offspring. Nutrients 2019, 11, 2014. [Google Scholar] [CrossRef] [Green Version]
- Ellery, S.J.; Dickinson, H.; McKenzie, M.; Walker, D.W. Dietary interventions designed to protect the perinatal brain from hypoxic-ischemic encephalopathy—Creatine prophylaxis and the need for multi-organ protection. Neurochem. Int. 2016, 95, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Allah Yar, R.; Akbar, A.; Iqbal, F. Creatine monohydrate supplementation for 10 weeks mediates neuroprotection and improves learning/memory following neonatal hypoxia ischemia encephalopathy in female albino mice. Brain Res. 2015, 1595, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, S.; Ali, M.; Iqbal, F. Long term creatine monohydrate supplementation, following neonatal hypoxic ischemic insult, improves neuromuscular coordination and spatial learning in male albino mouse. Brain Res. 2015, 1603, 76–83. [Google Scholar] [CrossRef]
- Iqbal, S.; Ali, M.; Iqbal, F. Effect of creatine monohydrate supplementation on relative serum level of IL-6 and IL-18 following neonatal hypoxia ischemia in male albino mouse. Pak. J. Pharm. Sci. 2015, 28, 2141–2145. [Google Scholar]
- Magistretti, P.J.; Allaman, I. Lactate in the brain: From metabolic end-product to signalling molecule. Nat. Rev. Neurosci. 2018, 19, 235–249. [Google Scholar] [CrossRef]
- Boumezbeur, F.; Petersen, K.F.; Cline, G.W.; Mason, G.F.; Behar, K.L.; Shulman, G.I.; Rothman, D.L. The contribution of blood lactate to brain energy metabolism in humans measured by dynamic 13C nuclear magnetic resonance spectroscopy. J. Neurosci. 2010, 30, 13983. [Google Scholar] [CrossRef] [PubMed]
- Berthet, C.; Lei, H.; Thevenet, J.; Gruetter, R.; Magistretti, P.J.; Hirt, L. Neuroprotective role of lactate after cerebral ischemia. J. Cereb. Blood Flow. Metab. 2009, 29, 1780–1789. [Google Scholar] [CrossRef]
- Jourdain, P.; Allaman, I.; Rothenfusser, K.; Fiumelli, H.; Marquet, P.; Magistretti, P.J. L-Lactate protects neurons against excitotoxicity: Implication of an ATP-mediated signaling cascade. Sci. Rep. 2016, 6, 21250. [Google Scholar] [CrossRef] [Green Version]
- Buscemi, L.; Blochet, C.; Price, M.; Magistretti, P.J.; Lei, H.; Hirt, L. Extended preclinical investigation of lactate for neuroprotection after ischemic stroke. Clin. Transl. Neurosci. 2020. [Google Scholar] [CrossRef] [Green Version]
- Tassinari, I.D.; Andrade, M.K.G.; da Rosa, L.A.; Hoff, M.L.M.; Nunes, R.R.; Vogt, E.L.; Fabres, R.B.; Sanches, E.F.; Netto, C.A.; Paz, A.H.; et al. Lactate administration reduces brain injury and ameliorates behavioral outcomes following neonatal hypoxia-ischemia. Neuroscience 2020, 448, 191–205. [Google Scholar] [CrossRef]
- Bereczki, D.J.; Balla, J.; Bereczki, D. Heme oxygenase-1: Clinical relevance in ischemic stroke. Curr. Pharm. Des. 2018, 24, 2229–2235. [Google Scholar] [CrossRef]
- Yu, P.; Wang, L.; Tang, F.; Zeng, L.; Zhou, L.; Song, X.; Jia, W.; Chen, J.; Yang, Q. Resveratrol pretreatment decreases ischemic injury and improves neurological function via sonic hedgehog signaling after stroke in rats. Mol. Neurobiol. 2017, 54, 212–226. [Google Scholar] [CrossRef]
- Hou, Y.; Wang, K.; Wan, W.; Cheng, Y.; Pu, X.; Ye, X. Resveratrol provides neuroprotection by regulating the JAK2/STAT3/PI3K/AKT/mTOR pathway after stroke in rats. Genes Dis. 2018, 5, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Dou, Z.; Rong, X.; Zhao, E.; Zhang, L.; Lv, Y. Neuroprotection of resveratrol against focal cerebral ischemia/reperfusion injury in mice through a mechanism targeting gut-brain axis. Cell Mol. Neurobiol. 2019, 39, 883–898. [Google Scholar] [CrossRef]
- Shin, J.A.; Lee, H.; Lim, Y.K.; Koh, Y.; Choi, J.H.; Park, E.M. Therapeutic effects of resveratrol during acute periods following experimental ischemic stroke. J. Neuroimmunol. 2010, 227, 93–100. [Google Scholar] [CrossRef]
- Angeloni, C.; Malaguti, M.; Barbalace, M.C.; Hrelia, S. Bioactivity of olive oil phenols in neuroprotection. Int. J. Mol. Sci. 2017, 18, 2230. [Google Scholar] [CrossRef] [Green Version]
- Misirli, G.; Benetou, V.; Lagiou, P.; Bamia, C.; Trichopoulos, D.; Trichopoulou, A. Relation of the traditional Mediterranean diet to cerebrovascular disease in a Mediterranean population. Am. J. Epidemiol. 2012, 176, 1185–1192. [Google Scholar] [CrossRef] [PubMed]
- Mohagheghi, F.; Bigdeli, M.R.; Rasoulian, B.; Zeinanloo, A.A.; Khoshbaten, A. Dietary virgin olive oil reduces blood brain barrier permeability, brain edema, and brain injury in rats subjected to ischemia-reperfusion. Sci. World J. 2010, 10, 1180–1191. [Google Scholar] [CrossRef] [Green Version]
- Rabiei, Z.; Bigdeli, M.R.; Rasoulian, B. Neuroprotection of dietary virgin olive oil on brain lipidomics during stroke. Curr. Neurovasc. Res. 2013, 10, 231–237. [Google Scholar] [CrossRef]
- Bu, Y.; Rho, S.; Kim, J.; Kim, M.Y.; Lee, D.H.; Kim, S.Y.; Choi, H.; Kim, H. Neuroprotective effect of tyrosol on transient focal cerebral ischemia in rats. Neurosci. Lett. 2007, 414, 218–221. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Liu, P.; Tang, H.; Jing, J.; Lv, X.; Chen, L.; Jiang, L.; Xu, J.; Li, J. Oleuropein, a natural extract from plants, offers neuroprotection in focal cerebral ischemia/reperfusion injury in mice. Eur. J. Pharm. 2016, 775, 113–119. [Google Scholar] [CrossRef]
- Netting, M.J.; Middleton, P.F.; Makrides, M. Does maternal diet during pregnancy and lactation affect outcomes in offspring? A systematic review of food-based approaches. Nutrition 2014, 30, 1225–1241. [Google Scholar] [CrossRef] [Green Version]
- Stiemsma, L.T.; Michels, K.B. The role of the microbiome in the developmental origins of health and disease. Pediatrics 2018, 141, e20172437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Du, L.; Shi, R.; Chen, Z.; Zhou, Y.; Li, Z. Early-life food nutrition, microbiota maturation and immune development shape life-long health. Crit. Rev. Food Sci. Nutr. 2019, 59, S30–S38. [Google Scholar] [CrossRef] [PubMed]
- Netto, C.A.; Sanches, E.F.; Odorcyk, F.; Duran-Carabali, L.E.; Sizonenko, S.V. Pregnancy as a valuable period for preventing hypoxia-ischemia brain damage. Int. J. Dev. Neurosci. 2018, 70, 12–24. [Google Scholar] [CrossRef]
- Brannon, P.M.; Taylor, C.L. Iron supplementation during pregnancy and infancy: Uncertainties and implications for research and policy. Nutrients 2017, 9, 1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Compound | Organism | HI Injury Method * | Injury Date | Administration Timing | Administration Duration ¶ | Dose | Administration Mode | Neuroprotective Effects | Ref |
|---|---|---|---|---|---|---|---|---|---|
| POLYPHENOLS | |||||||||
| Resveratrol | Mouse and rat | RV: 8% O2 for 45 min (mouse) or 2.5 h (rat) | P7 | Pups: 24 h or 10 min before H | Single dose | 0.002, 0.2 or 20 mg/kg | i.p. injection | ↓ Tissue loss in hippocampus and striatum, ↓ apoptotic and necrotic cell death | [43] |
| Rat | RV: 8% O2 for 2 h 15 min | P7 | Pups: 10 min before H | Single dose | 20 mg/kg | i.p. injection | ↓ Brain infarct volume, loss of myelination and cell loss in cortex and hippocampus, ↓ ROS production, ↑ maintenance of the mitochondrial inner membrane integrity and transmembrane potential, ↓ long-term cognitive impairments and functional damage | [44] | |
| Rat | RV: 8% O2 for 2 h 15 min | P7 | Pups: 10 min before HI | Single dose | 20 mg/kg | i.p. injection | ↓ Morphological damage and astrogliosis in the inferior colliculus, ↓ loss of myelination, restored the auditory brainstem functional response | [45] | |
| Rat | RV: 8% O2 for 2.5 h | P14 | Pups: P7–P14 | 7 days | 20 or 40 mg/kg/day | i.p. injection | ↓ Brain infarct volume, ↓ cerebral edema, ↓ neuroinflammation, ↓ oxidative stress, ↑ Nrf2/HO-1 signaling pathway | [46] | |
| Rat | Helmy model: 9% O2 + 20% CO2 for 90 min | P6 | Mothers: from end of weaning till pups reached P7 | 88–98 days † | 50 mg/kg/day | Drinking water | ↓ Neuroinflammation in hippocampus | [47] | |
| Rat | RV: 8% O2 for 2 h | P7 | Mothers: E15–P9 | 15 days | 0.15 mg/kg/day | Drinking water | Partially ↓ sensorimotor defects and long-term memory deficits in the context of moderate maternal alcohol consumption | [48] | |
| Piceatannol | Rat | RV: 8% O2 for 2 h | P7 | Mothers: E15–P7 or P0–P7 | 6 or 14 days | 0.15 mg/kg/day | Drinking water | ↓ Brain infarct volume and anatomical brain lesions, ↓ cerebral edema, ↓ neuronal apoptosis, ↑ early reflexes, ↓ sensorimotor defects and long-term cognitive impairments | [49] |
| Rat | RV: 8% O2 for 2 h | P7 | Mothers: E15–P9 | 15 days | 0.15 mg/kg/day | Drinking water | ↓ Sensorimotor defects and cognitive impairments in the context of moderate maternal alcohol consumption | [48] | |
| Pterostilbene | Rat | RV: 8% O2 for 2 h | P7 | Pups: 30 min before HI | Single dose | 50 mg/kg | i.p. injection | ↓ Brain infarct volume, ↓ brain edema, ↓ neuronal apoptosis, ↓ neuroinflammation, ↓ oxidative stress through ↑ HO-1 expression, ↓ motor and memory deficits | [50] |
| Quercetin | Piglet | Transient bilateral carotid ligation + 8% O2 for 40 min | P2 | Pups: 1 h before HI | Single dose | 10 mg/kg | Nanosomes injected intravenously | Improved electroencephalographic amplitude records and neurological functions, restored blood pressure and spontaneous breathing | [51] |
| Rat | RV: 8% O2 for 2.5 h | P7 | Pups: P0–P7 | 7 days | 40 mg/kg/day | Intragastric | ↓ Cortical cell apoptosis, microgliosis, and astrogliosis, ↓ neuroinflammation, ↓ TLR4/NF-κB signaling | [52] | |
| Mangiferin | Rat | RV: 8% O2 for 2 h | P10 | Pups: P3–P12 | 9 days | 50, 100 or 200 mg/kg/day | Oral gavage | ↓ Brain infarct volume, ↓ neuronal apoptosis, ↓ oxidative stress, ↑ PI3K/Akt signaling pathway, ↑ isoflurane’s neuroprotection | [53] |
| Pomegranate juice | Mouse | RV: 8% O2 for 45 min | P7 | Mothers: E14–P8 or E14–P14 | 15 or 21 days | 8, 16, or 32 µmol/day | Drinking water | ↓ Tissue loss in the hippocampus, cortex, and striatum in a dose-dependent manner, ↓ neuronal apoptosis | [54] |
| Mouse and rat | RV: 8% O2 for 45 min (mouse) or 2.5 h (rat) | P7 | Mothers: E0–P8 | 29 days | 4.8 mg/day | Drinking water | ↓ Neuronal apoptosis | [43] | |
| GSPE | Mouse | RV: 8% O2 for 2 h | P7 | Pups: 20 min before HI | Single dose | 30 mg/kg | i.p. injection | ↓ Brain infarct volume, ↓ neuronal apoptosis, improved neurobehavioral outcomes | [55] |
| Icariin | Mouse | RV: 8% O2 for 2 h | P7 | Pups: 20 min before HI | Single dose | 10 mg/kg | i.p. injection | ↓ Brain infarct volume, ↓ neuronal apoptosis, improved neurobehavioral outcomes, ↑ PI3K/Akt signaling pathway | [56] |
| Daphnetin | Rat | RV: 8% O2 for 2.5 h | P7 | Pups: 1 h before H | Single dose | 10 mg/kg | i.p. injection | ↓ Brain infarct volume | [57] |
| OMEGA-3 FATTY ACIDS | |||||||||
| n-3 PUFAs | Rat | RV: 8% O2 for 90 min | P7 | Mothers: E1–P21 | 41 days | 46% w/w total fatty acids | Flaxseed-enriched diet | ↓ Brain tissue loss, ↑ hippocampal n-3 PUFAs content in pups’ brains, ↓ depressive behavior and cognitive impairments | [58] |
| Rat | 3 hypoxic insults per day with 10% O2 + 3% CO2 for 2 h | P7–P12 | Mothers: E1–P12 | 32 days | 3.5–4 g/day | Fish-oil-enriched diet | ↓ Hippocampal apoptosis, preserved striatal dopamine levels | [59] | |
| Mouse | RV: 8% O2 for 15 min | P10 | Pups: postsurgery and post-H | 2 doses | 3 mg/day | i.p. injection | ↓ Brain infarct volume | [60] | |
| DHA and EPA | Rat | RV: 8% O2 for 2.5 h | P7 | Mothers: E2–P14 | 33 days | 1.5% w/w | Fish-oil-enriched diet | ↓ Brain tissue loss, ↑ cortical content of DHA and EPA in pups’ brain, ↓ microgliosis, ↓ neuroinflammation, ↓ long-term sensorimotor and cognitive impairments | [61] |
| Rat | RV: 8% O2 for 2.5 h | P7 | Mothers: E2–P14 | 33 days | Not specified | Fish-oil-enriched diet | ↓ Brain tissue loss, ↓ cortical apoptosis, ↑ phosphatidylserine, DHA and EPA content in pups’ brain, ↑ PI3K/Akt signaling pathway, improved neurological outcomes | [62] | |
| Rat | RV: 8% O2 for 2.5 h | P7 | Mothers: E2–P14 | 33 days | 1.5% w/w | Fish-oil-enriched diet | ↓ Brain edema, ↑ BBB integrity, ↓ matrix metalloproteinase activity | [63] | |
| DHA | Rat | RV: 8% O2 for 90 min | P7 | Mothers: E7 till pups’ sacrifice at P8–P14 | 22–28 days | 10% w/w total fatty acids | Fish-oil-enriched diet | ↑ DHA content in pups’ brain, ↓ hippocampal apoptosis, ↓ oxidative stress | [64] |
| Rat | RV: 8% O2 for 90 min | P7 | Pups: 2.5 h before HI | Single dose | 1, 2.5 or 5 mg/kg | i.p. injection | ↓ Hippocampal tissue loss, improved functional outcomes | [65] | |
| Rat | RV: 8% O2 for 90 min | P7 | Pups: 2.5 h before HI | Single dose | 1 mg/kg | i.p. injection | Improved functional outcomes in the context of E. coli lipopolysaccharide-induced systemic inflammation | [66] | |
| Rat | RV: 8% O2 for 2 h 15 min | P7 | Pups: 10 min before HI | Single dose | 1 mg/kg | i.p. injection | ↓ Brain infarct volume, ↓ loss of myelination, ↓ astrogliosis and microgliosis, ↑ maintenance of the mitochondrial inner membrane integrity and transmembrane potential, ↓ long-term behavioral and cognitive impairments | [67] | |
| Rat | RV: 8% O2 for 2 h 15 min | P7 | Pups: 10 min before HI | Single dose | 1 mg/kg | i.p. injection | ↓ Astrogliosis, ↓ loss of myelination, restored the auditory brainstem functional response | [45] | |
| VITAMINS | |||||||||
| Vitamin A | Rat | RV: 8% O2 for 2.5 h | P7 | Mothers: 4 weeks before pregnancy till pups’ sacrifice | Whole pregnancy and lactation | 300 or 7000 IU/kg/day | Diet | ↓ Hippocampal cell apoptosis by ↑ caspase-3 and caspase-8/Bid pathways, ↑ PI3K/Akt signaling pathway by binding to RARα, improved learning ability, and spatial memory impairments | [68] |
| Rat | RV: 8% O2 for 2.5 h | P7 | Mothers: 4 weeks before pregnancy till pups’ sacrifice | Whole pregnancy and lactation | 300 or 7000 IU/kg/day | Diet | ↑ Neurol stem cell proliferation by ↑ RARα-mediated modulation of β-catenin signaling, improved learning ability, and spatial memory impairments | [69] | |
| Vitamin E | Rat | RV: 8% O2 for 90 min | P7 | Pups: P4, P6 and P8 | 3 doses | 1.5 mg/day | s.c. injection | ↓ Expression of iNOS, nNOS and IGF-related proteins | [70] |
| Folic acid | Rat | RV: 8% O2 for 90 min | P7 | Mothers: E0 till birth | Whole pregnancy | 2 or 20 mg/kg/day | Diet | ↓ Long-term memory impairments, ↓ brain-derived neurotrophic factor imbalance | [71] |
| PLANT-DERIVED COMPOUNDS | |||||||||
| Tanshinone I | Rat | RV: 8% O2 for 2.5 h | P7 | Pups: P6–P12 | 6 days | 5 mg/kg/day | i.p. injection | ↓ Neuronal loss in the hippocampus, ↓ motor and cognitive deficits, ↓ oxidative stress | [72] |
| Tanshinone IIA | Rat | RV: 8% O2 for 2 h | P7 | Pups: P5–14 or P5–21 | 9 or 16 days | 10 mg/kg/day | i.p. injection | ↓ Brain injury, ↓ cortical cell loss, ↑ plasma antioxidant capacity, improved sensorimotor function | [73] |
| Sulforaphane | Rat | RV: 8% O2 for 90 min | P7 | Pups: 30 min before HI | Single dose | 5 mg/kg | i.p. injection | ↓ Brain infarct volume, ↓ apoptosis in cortex and hippocampus, ↓ microgliosis, ↓ lipid peroxidation, ↑ Nrf2/HO-1 signaling pathway | [74] |
| Rat | Wigglesworth model: bilateral uterine artery ligation | E20 | Mothers: E15–P14 | 20 days | 200 mg/day | Diet (dried broccoli sprouts) | ↓ White matter loss and ventricular dilation, ↓ hippocampal cell loss, ↓ loss of myelination, ↓ astrogliosis, improved neurobehavioral outcomes | [75] | |
| Capsaicin | Rat | RV: 8% O2 for 2 h | P10 | Pups: 3 h before HI | Single dose | 0.2 or 2 mg/kg | i.p. injection | ↓ Brain infarct volume, ↑ the middle cerebral artery myogenic tone | [76] |
| ENDOGENOUS COMPOUNDS | |||||||||
| Melatonin | Spiny mouse | Bjelke model: 7.5 min intrauterine ischemia | E37 | Mothers: E29–E37 | 8 days | 0.1 mg/kg/day | Subscapula osmotic pump | ↓ Macrophage infiltration, ↓ microgliosis, ↓ apoptosis in cortex | [77] |
| Rat | Bilateral utero-ovarian artery occlusion for 30 min | E16 | Mothers: E0 till birth | Whole pregnancy | 4 mg/kg/day | Drinking water | ↓ Mitochondrial damage, ↓ degeneration of pyramidal cells in the hippocampus | [78] | |
| Rat | RV: 8% O2 for 2.5 h | P7 | Pups: 30 min before HI | Single dose | 5 or 15 mg/kg | i.p. injection | ↓ Brain tissue loss in a concentration-dependent manner | [79] | |
| Rat | RV: 8% O2 for 2.5 h | P7 | Pups: 30 min before HI | Single dose | 15 mg/kg | i.p. injection | ↓ Brain damage in hippocampus and cortex, ↓ oxidative stress by ↓ the levels of free iron, F2-isoprostanes, and F4-neuroprostanes | [80] | |
| Rat | Excitotoxity injury by ibotenate injection | P5 | Pups: 15 min before ibotenate injection | Single dose | 5 mg/kg | i.p. injection | ↓ White matter lesions, preserved the ability to develop conditioning | [36] | |
| Carnitine | Rat | RV: 8% O2 for 70 min | P7 | Pups: 30 min before H | Single dose | 16 mmol/kg | i.p. injection | ↓ Brain injury, ↓ apoptosis in hippocampus and cortex | [81] |
| Rat | RV: 8% O2 for 70 min | P7 | Pups: 30 min before H | Single dose | 16 mmol/kg | i.p. injection | ↓ Accumulation of acyl-CoA esters in the brain, ↓ superoxide levels, ↓ mitochondrial injury | [82] | |
| Rat | RV: 8% O2 for 1 h | P7 | Pups: just before HI | Single dose | 200 mg/kg | i.p. injection | ↓ Apoptosis in hippocampus and striatum | [83] | |
| Creatine | Spiny mouse | Bjelke model: 7.5 min intrauterine ischemia | E37 or E38 | Mothers: E20 till sacrifice at E37–E38 | 17–18 days | 5% w/w | Enriched diet | ↑ Capacity of the offspring to survive birth asphyxia, improved postnatal growth, ↑ creatine content in brain and other fetal tissues | [84] |
| Spiny mouse | Bjelke model: 7.5 min intrauterine ischemia | E38 | Mothers: E20–E38 | 18 days | 5% w/w | Enriched diet | ↓ Lipid peroxidation, ↓ apoptosis, ↓ mitochondrial damage | [85] | |
| Rat | RV: 8% O2 for 100 min | P7 | Pups: P6–P8 | 3 doses | 3 g/kg/day | s.c. injection | ↓ Brain edema, ↑ energy potential and the levels of phosphocreatine in the brain | [86] | |
| Rat | RV: 8% O2 for 80 min | P7 | Pups: P4–P7 | 4 doses | 3 g/kg/day | s.c. injection | ↓ Brain injury, ↓ neuronal cell damage in the cortex and hippocampus | [87] | |
| Lactate | Rat | RV: 8% O2 for 2 h | P7 | Pups: just before H | Single dose | 2.14 mmol/kg | i.p. injection | ↓ Brain infarct volume | [88] |
| Cell Death | Oxidative Stress | Neuroinflammation | BBB Integrity and Edema | Excito Toxicity | Energy Metabolism | Synaptic Function and Neuroplasticity | |
|---|---|---|---|---|---|---|---|
| POLYPHENOLS | |||||||
| Resveratrol | x | x | x | x | x | ||
| Piceatannol | x | x | |||||
| Pterostilbene | x | x | x | x | |||
| Quercetin | x | x | x | ||||
| Mangiferin | x | x | |||||
| Pomegranate polyphenols | x | ? | |||||
| GSPE | x | ||||||
| Icariin | x | ||||||
| Daphnetin | ? | ||||||
| OMEGA-3 FATTY ACIDS | |||||||
| EPA | x | x | x | ||||
| DHA | x | x | x | x | x | ||
| VITAMINS | |||||||
| Vitamin A | x | x | |||||
| Vitamin B9 | ? | ||||||
| Vitamin D | ? | ||||||
| Vitamin E | x | ||||||
| PLANT-DERIVED COMPOUNDS | |||||||
| Tanshinones | x | x | |||||
| Sulforaphane | x | x | x | ||||
| Capsaicin | ? | ||||||
| ENDOGENOUS COMPOUNDS | |||||||
| Melatonin | x | x | x | ||||
| Carnitine | x | x | x | ||||
| Creatine | x | x | x | x | |||
| Lactate | ? | ? | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reyes-Corral, M.; Sola-Idígora, N.; de la Puerta, R.; Montaner, J.; Ybot-González, P. Nutraceuticals in the Prevention of Neonatal Hypoxia–Ischemia: A Comprehensive Review of their Neuroprotective Properties, Mechanisms of Action and Future Directions. Int. J. Mol. Sci. 2021, 22, 2524. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052524
Reyes-Corral M, Sola-Idígora N, de la Puerta R, Montaner J, Ybot-González P. Nutraceuticals in the Prevention of Neonatal Hypoxia–Ischemia: A Comprehensive Review of their Neuroprotective Properties, Mechanisms of Action and Future Directions. International Journal of Molecular Sciences. 2021; 22(5):2524. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052524
Chicago/Turabian StyleReyes-Corral, Marta, Noelia Sola-Idígora, Rocío de la Puerta, Joan Montaner, and Patricia Ybot-González. 2021. "Nutraceuticals in the Prevention of Neonatal Hypoxia–Ischemia: A Comprehensive Review of their Neuroprotective Properties, Mechanisms of Action and Future Directions" International Journal of Molecular Sciences 22, no. 5: 2524. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052524