
Identification of Predictive Biomarkers of Response to HSP90 Inhibitors in Lung Adenocarcinoma

 ,
, 
Abstract
:1. Introduction
2. Results
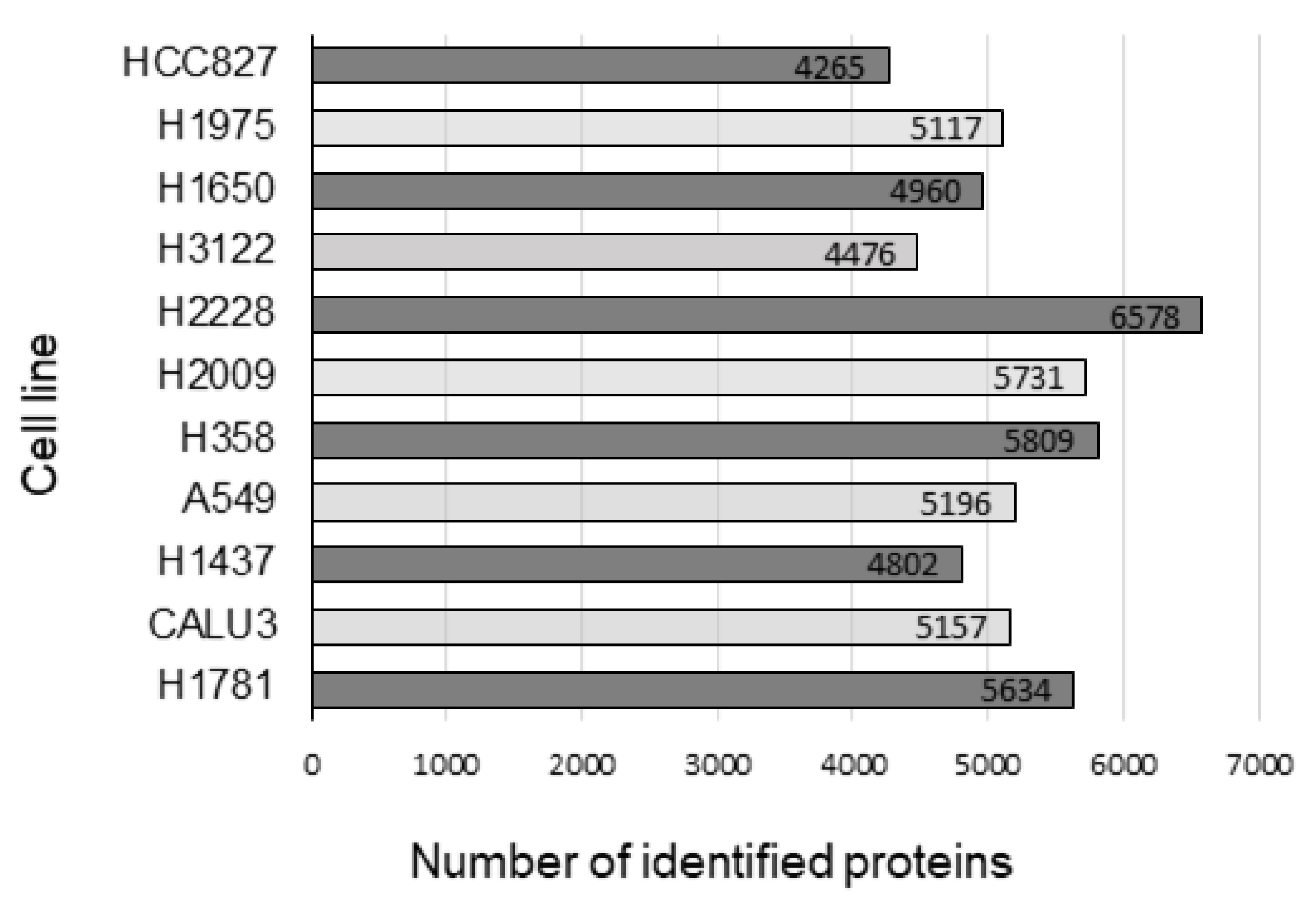
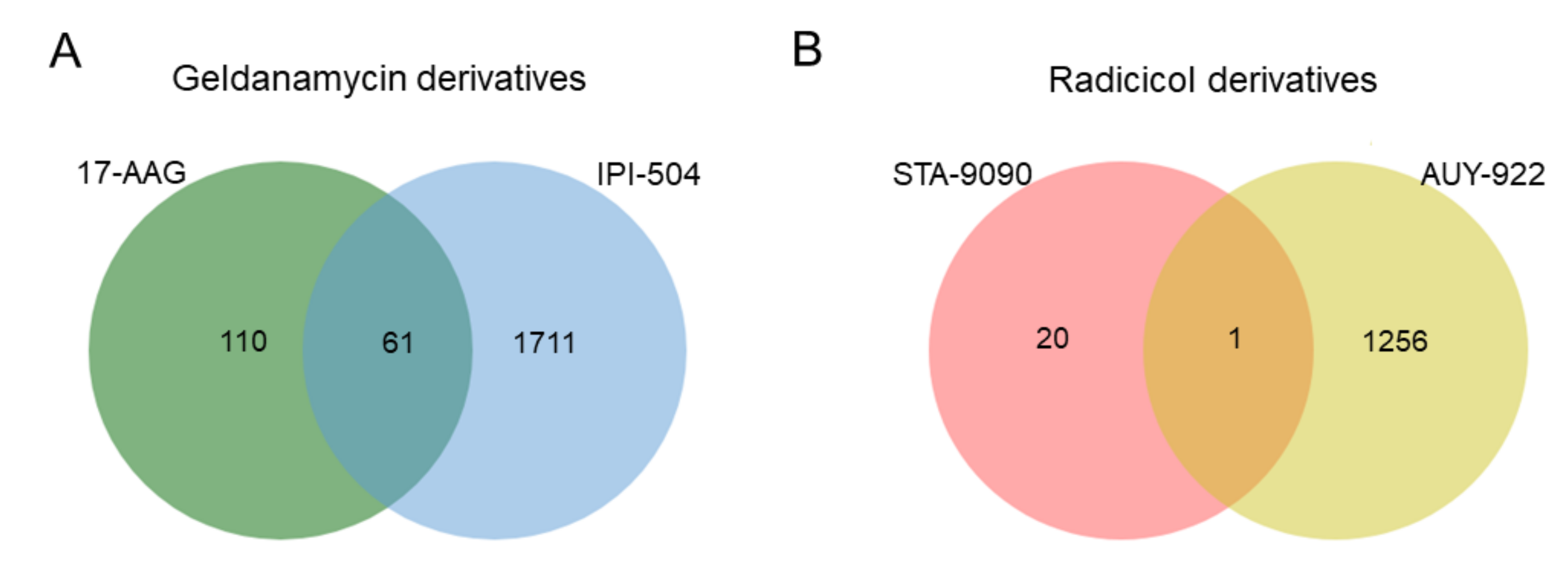
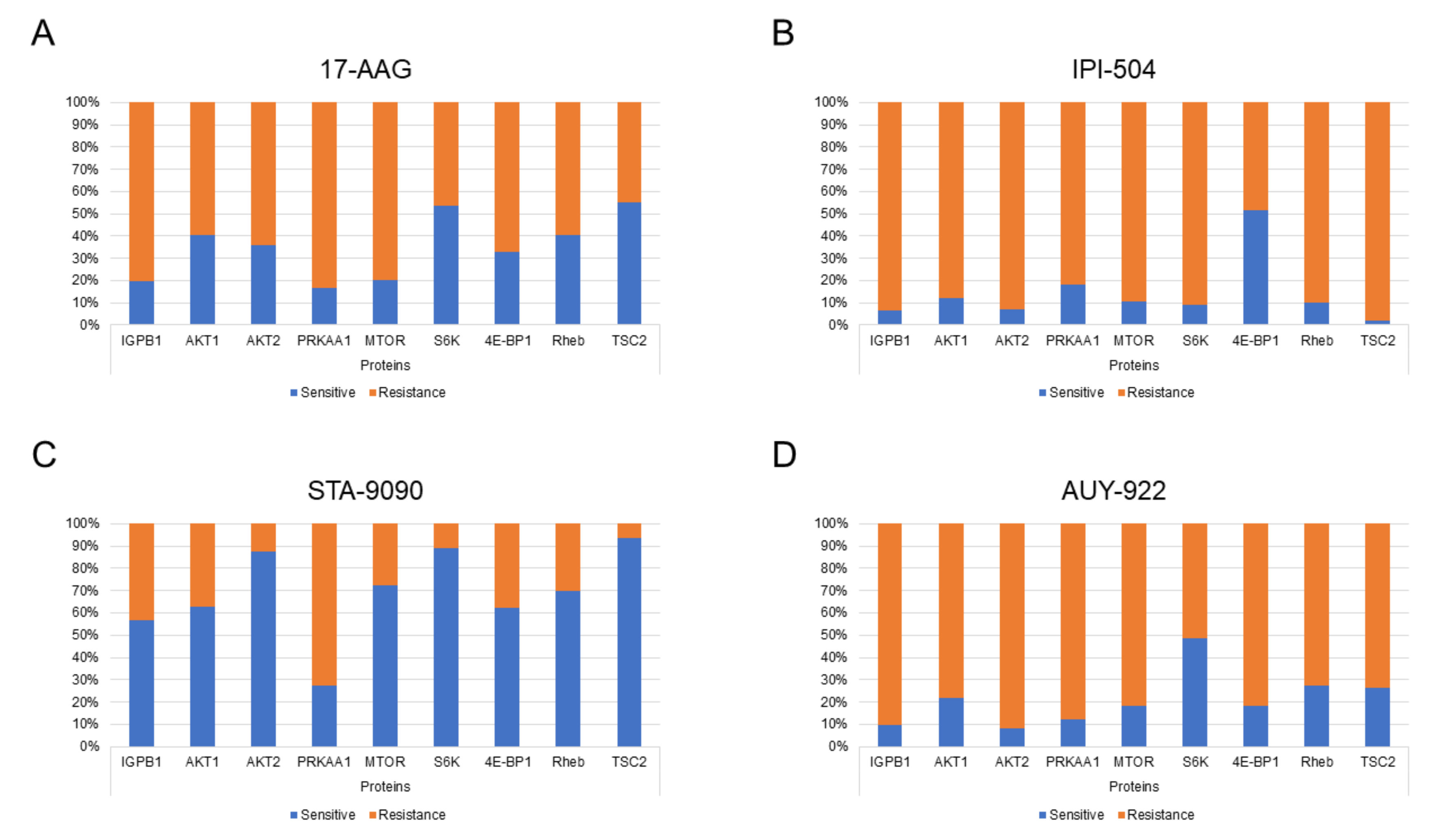
2.1. Determination of Proteomic Profiles Correlated with HSP90 Inhibitor Sensitivity or Resistance
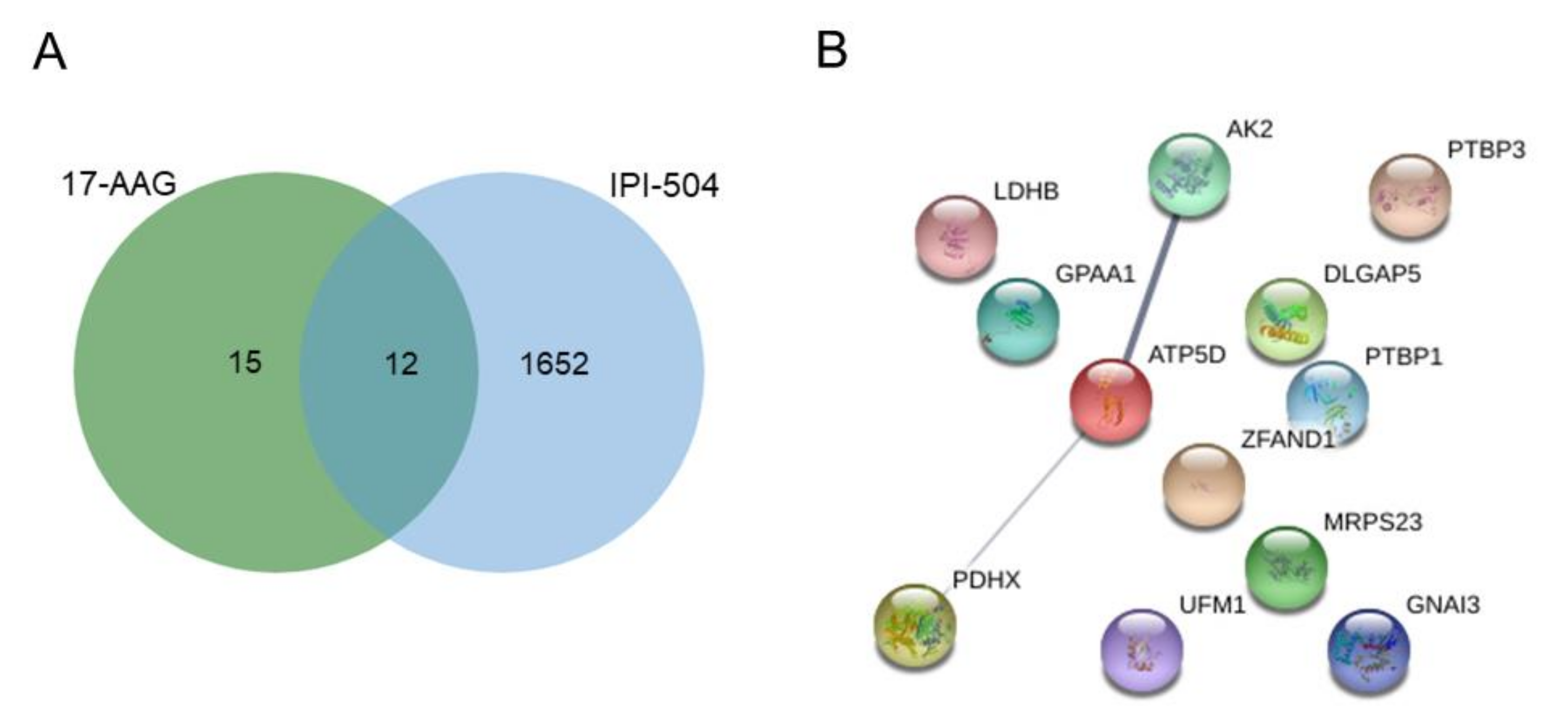
2.2. Identification of Candidate Biomarkers for Predicting Sensitivity to HSP90 Inhibitors in Lung Adenocarcinoma Cell Lines
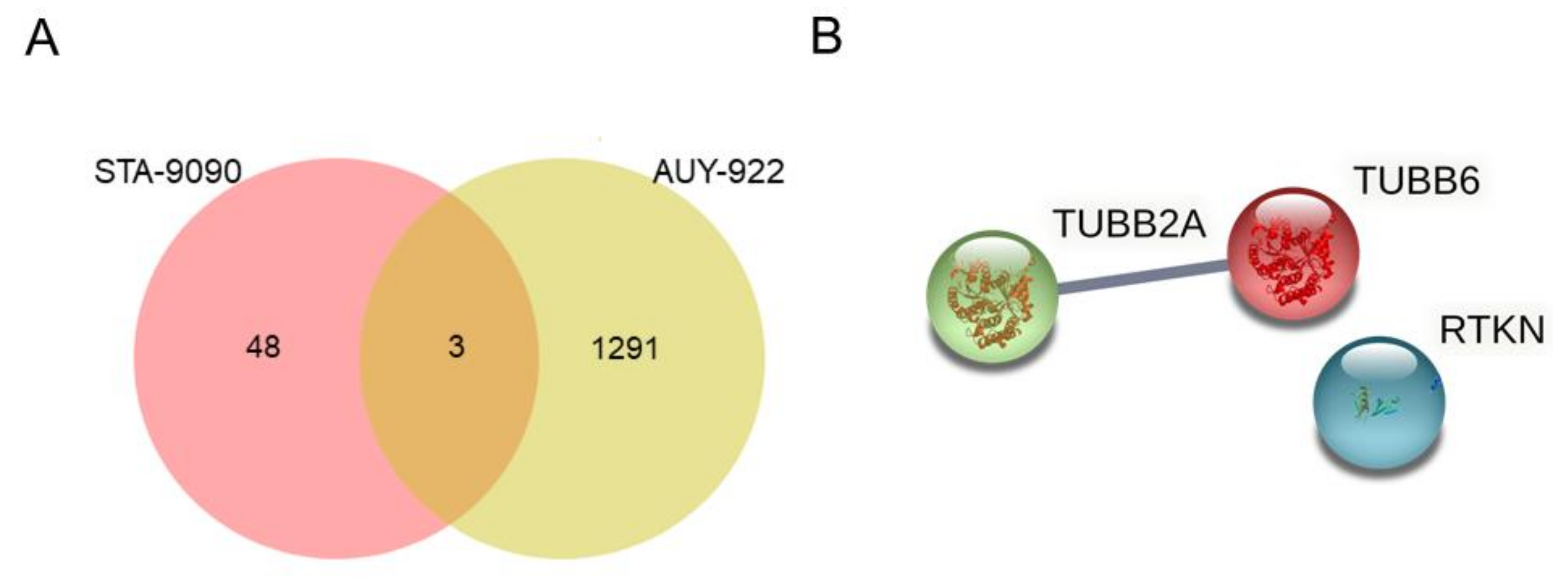
2.3. Detection of Potential Predictive Biomarkers of Resistance to HSP90 Inhibition in Lung Adenocarcinoma Cell Lines
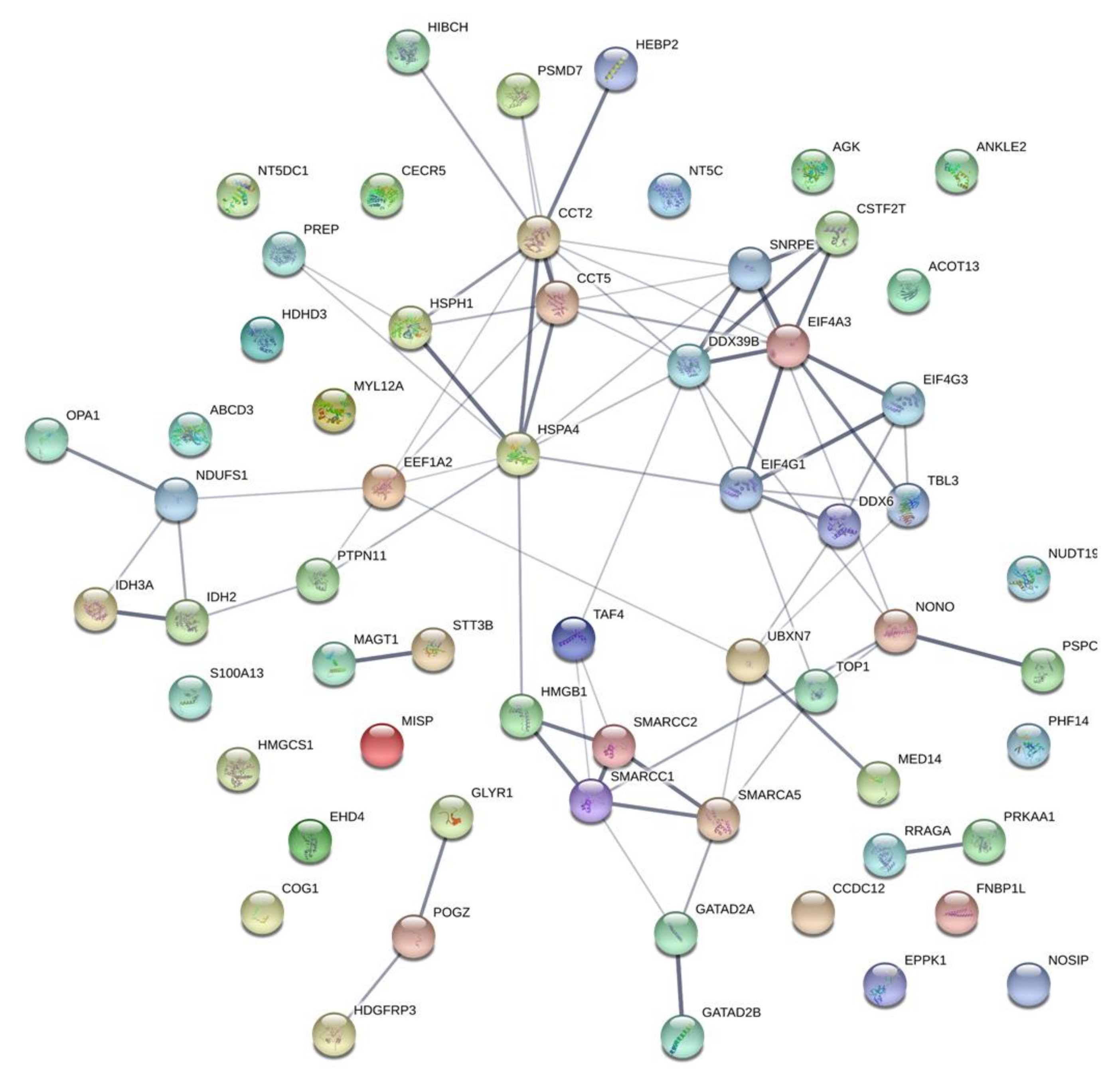
2.4. Candidate Pathways Associated with Susceptibility to HSP90 Inhibition
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Cell Viability
4.3. Proteomic Sample Preparation
4.4. Trypsin Digestion and iTRAQ Labeling
4.5. Cleaning and Fractionation
4.6. NanoLC-MS/MS Analysis
4.7. Protein Identification and Quantification
4.8. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| 17-AAG | Tanespimycin |
| 7PSMD7 | 26S proteasome non-ATPase regulatory subunit |
| 4E-BP1 | Eukaryotic translation initiation factor 4E-binding protein 1 |
| AK2 | Adenylate kinase 2 |
| AKT1 | AKT serine/threonine kinase 1 |
| AKT2 | AKT serine/threonine kinase 2 |
| ALK | Anaplastic lymphoma kinase |
| ATP5D | ATP synthase subunit delta |
| AUY-922 | Luminespib |
| BRAF | B-Raf proto-oncogene: serine/threonine kinase |
| COG1 | Conserved oligomeric Golgi complex subunit 1 |
| DLGAP5 | Disks large-associated protein 5 |
| EGFR | Epidermal growth factor receptor |
| EIF3I | Eukaryotic translation initiation factor 3 subunit I |
| EML4 | Echinoderm microtubule-associated protein-like |
| ERBB2 | Erb-B2 receptor tyrosine kinase 2 |
| GNAI3 | Guanine nucleotide-binding protein G(k) subunit alpha |
| GPAA1 | Glycosylphosphatidylinositol anchor attachment 1 protein |
| HSP | Heat shock protein |
| HSP90 | Heat shock protein 90 |
| IGBP1 | Immunoglobulin binding protein 1 |
| IPI-504 | Retaspimycin hydrochloride |
| LDHB | Lactate dehydrogenase B |
| MET | MET proto-oncogene, receptor tyrosine kinase |
| MYL12A | Myosin regulatory light chain 12A |
| MRPS23 | Mitochondrial ribosomal protein S23 |
| mTOR | Mechanistic target of rapamycin kinase |
| NSCLC | Non-small-cell lung cancer |
| PDHX | Pyruvate dehydrogenase protein X component |
| PDSS2 | Decaprenyl diphosphate synthase subunit 2 |
| PRKAA1 | Protein kinase AMP-activated catalytic subunit alpha 1 |
| PTBP1 | Polypyrimidine tract-binding protein 1 |
| PTBP3 | Polypyrimidine tract-binding protein 3 |
| Rheb | GTP-binding protein Rheb |
| RTKN | Rhotekin |
| S6K | Ribosomal protein S6 kinase beta-1 |
| STA-9090 | Ganetespib |
| TOP1 | DNA topoisomerase 1 |
| TSC2 | Tuberin |
| TUBB2A | Tubulin beta-2A chain |
| TUBB6 | Tubulin beta-6 chain |
| UFM1 | Ubiquitin-fold modifier 1 |
| ZFAND1 | Zinc finger AN1-type containing 1 |
References
- Hoter, A.; EI-Sabban, M.E.; Naim, H.Y. The HSP90 Family: Structure, Regulation, Function, and Implications in Health and Disease. Int. J. Mol. Sci. 2018, 19, 2560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Echeverria, P.C.; Bernthaler, A.; Dupuis, P.; Mayer, B.; Picard, D. An interaction network predicted from public data as a discovery tool: Application to the Hsp90 molecular chaperone machine. PLoS ONE 2011, 6, e26044. [Google Scholar] [CrossRef]
- Tatokoro, M.; Koga, F.; Yoshida, S.; Kihara, K. Heat shock protein 90 targeting therapy: State of the art and future perspective. Excli J. 2015, 14, 48–58. [Google Scholar]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Whitesell, L.; Lindquist, S.L. HSP90 and the chaperoning of cancer. Nat. Rev. Cancer 2005, 5, 761–772. [Google Scholar] [CrossRef]
- Butler, L.M.; Ferraldeschi, R.; Armstrong, H.K.; Centenera, M.M.; Workman, P. Maximizing the Therapeutic Potential of HSP90 Inhibitors. Mol. Cancer Res. 2015, 13, 1445–1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Workman, P.; Burrows, F.; Neckers, L.; Rosen, N. Drugging the cancer chaperone HSP90: Combinatorial therapeutic exploitation of oncogene addiction and tumor stress. Ann. N. Y. Acad. Sci. 2007, 1113, 202–216. [Google Scholar] [CrossRef] [PubMed]
- Neckers, L.; Workman, P. Hsp90 molecular chaperone inhibitors: Are we there yet? Clin. Cancer Res. 2012, 18, 64–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Shimamura, T.; Lowell, A.M.; Engelman, J.A.; Shapiro, G.I. Epidermal growth factor receptors harboring kinase domain mutations associate with the heat shock protein 90 chaperone and are destabilized following exposure to geldanamycins. Cancer Res. 2005, 65, 6401–6408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.; Soga, S.; Beebe, K.; Lee, M.J.; Kim, Y.S.; Trepel, J.; Neckers, L. Sensitivity of epidermal growth factor receptor and ErbB2 exon 20 insertion mutants to Hsp90 inhibition. Br. J. Cancer 2007, 97, 741–744. [Google Scholar] [CrossRef]
- Wang, S.; Pashtan, I.; Tsutsumi, S.; Xu, W.; Neckers, L. Cancer cells harboring MET gene amplification activate alternative signaling pathways to escape MET inhibition but remain sensitive to Hsp90 inhibitors. Cell Cycle 2009, 8, 2050–2056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Rocha Dias, S.; Friedlos, F.; Light, Y.; Springer, C.; Workman, P.; Marais, R. Activated B-RAF is an Hsp90 client protein that is targeted by the anticancer drug 17-allylamino-17-demethoxygeldanamycin. Cancer Res. 2005, 65, 10686–10691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Normant, E.; Paez, G.; West, K.A.; Lim, A.R.; Slocum, K.L.; Tunkey, C.; McDougall, J.; Wylie, A.A.; Robison, K.; Caliri, K.; et al. The Hsp90 inhibitor IPI-504 rapidly lowers EML4-ALK levels and induces tumor regression in ALK-driven NSCLC models. Oncogene 2011, 30, 2581–2586. [Google Scholar] [CrossRef] [Green Version]
- Esfahani, K.; Cohen, V. HSP90 as a novel molecular target in non-small-cell lung cancer. Lung Cancer (Auckl.) 2016, 7, 11–17. [Google Scholar] [PubMed] [Green Version]
- Gallegos Ruiz, M.I.; Floor, K.; Roepman, P.; Rodriguez, J.A.; Meijer, G.A.; Mooi, W.J.; Jassem, E.; Niklinski, J.; Muley, T.; van Zandwijk, N.; et al. Integration of gene dosage and gene expression in non-small cell lung cancer, identification of HSP90 as potential target. PLoS ONE 2008, 3, e0001722. [Google Scholar] [CrossRef] [PubMed]
- Hendriks, L.E.L.; Dingemans, A.C. Heat shock protein antagonists in early stage clinical trials for NSCLC. Expert. Opin. Investig. Drugs 2017, 26, 541–550. [Google Scholar] [CrossRef]
- Proia, D.A.; Sang, J.; He, S.; Smith, D.L.; Sequeira, M.; Zhang, C.; Liu, Y.; Ye, S.; Zhou, D.; Blackman, R.K.; et al. Synergistic activity of the Hsp90 inhibitor ganetespib with taxanes in non-small cell lung cancer models. Invest. New Drugs 2012, 30, 2201–2209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Liu, H.; Diao, L.; Potter, A.; Zhang, J.; Qiao, Y.; Wang, J.; Proia, D.A.; Tailor, R.; Komaki, R.; et al. Hsp90 Inhibitor Ganetespib Sensitizes Non-Small Cell Lung Cancer to Radiation but Has Variable Effects with Chemoradiation. Clin. Cancer Res. 2016, 22, 5876–5886. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.L.; Acquaviva, J.; Sequeira, M.; Jimenez, J.P.; Zhang, C.; Sang, J.; Bates, R.C.; Proia, D.A. The HSP90 inhibitor ganetespib potentiates the antitumor activity of EGFR tyrosine kinase inhibition in mutant and wild-type non-small cell lung cancer. Target. Oncol. 2015, 10, 235–245. [Google Scholar] [CrossRef] [Green Version]
- Sang, J.; Acquaviva, J.; Friedland, J.C.; Smith, D.L.; Sequeira, M.; Zhang, C.; Jiang, Q.; Xue, L.; Lovly, C.M.; Jimenez, J.P.; et al. Targeted inhibition of the molecular chaperone Hsp90 overcomes ALK inhibitor resistance in non-small cell lung cancer. Cancer Discov. 2013, 3, 430–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akram, A.; Khalil, S.; Halim, S.A.; Younas, H.; Iqbal, S.; Mehar, S. Therapeutic Uses of HSP90 Inhibitors in Non-Small Cell Lung Carcinoma (NSCLC). Curr. Drug. Metab. 2018, 19, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Prodromou, C.; Roe, S.M.; O’Brien, R.; Ladbury, J.E.; Piper, P.W.; Pearl, L.H. Identification and structural characterization of the ATP/ADP-binding site in the Hsp90 molecular chaperone. Cell 1997, 90, 65–75. [Google Scholar] [CrossRef] [Green Version]
- Ehrlich, E.S.; Wang, T.; Luo, K.; Xiao, Z.; Niewiadomska, A.M.; Martinez, T.; Xu, W.; Neckers, L.; Yu, X.F. Regulation of Hsp90 client proteins by a Cullin5-RING E3 ubiquitin ligase. Proc. Natl. Acad. Sci. USA 2009, 106, 20330–20335. [Google Scholar] [CrossRef] [Green Version]
- Sequist, L.V.; Gettinger, S.; Senzer, N.N.; Martins, R.G.; Janne, P.A.; Lilenbaum, R.; Gray, J.E.; Iafrate, A.J.; Katayama, R.; Hafeez, N.; et al. Activity of IPI-504, a novel heat-shock protein 90 inhibitor, in patients with molecularly defined non-small-cell lung cancer. J. Clin. Oncol. 2010, 28, 4953–4960. [Google Scholar] [CrossRef] [PubMed]
- Sessa, C.; Shapiro, G.I.; Bhalla, K.N.; Britten, C.; Jacks, K.S.; Mita, M.; Papadimitrakopoulou, V.; Pluard, T.; Samuel, T.A.; Akimov, M.; et al. First-in-human phase I dose-escalation study of the HSP90 inhibitor AUY922 in patients with advanced solid tumors. Clin. Cancer Res. 2013, 19, 3671–3680. [Google Scholar] [CrossRef] [Green Version]
- Felip, E.; Barlesi, F.; Besse, B.; Chu, Q.; Gandhi, L.; Kim, S.W.; Carcereny, E.; Sequist, L.V.; Brunsvig, P.; Chouaid, C.; et al. Phase 2 Study of the HSP-90 Inhibitor AUY922 in Previously Treated and Molecularly Defined Patients with Advanced Non-Small Cell Lung Cancer. J. Thorac. Oncol. 2018, 13, 576–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, M.L.; Yu, H.A.; Hart, E.M.; Weitner, B.B.; Rademaker, A.W.; Patel, J.D.; Kris, M.G.; Riely, G.J. Phase I/II Study of HSP90 Inhibitor AUY922 and Erlotinib for EGFR-Mutant Lung Cancer With Acquired Resistance to Epidermal Growth Factor Receptor Tyrosine Kinase Inhibitors. J. Clin. Oncol. 2015, 33, 1666–1673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramalingam, S.; Goss, G.; Rosell, R.; Schmid-Bindert, G.; Zaric, B.; Andric, Z.; Bondarenko, I.; Komov, D.; Ceric, T.; Khuri, F.; et al. A randomized phase II study of ganetespib, a heat shock protein 90 inhibitor, in combination with docetaxel in second-line therapy of advanced non-small cell lung cancer (GALAXY-1). Ann. Oncol. 2015, 26, 1741–1748. [Google Scholar] [CrossRef]
- Pillai, R.N.; Fennell, D.A.; Kovcin, V.; Ciuleanu, T.E.; Ramlau, R.; Kowalski, D.; Schenker, M.; Yalcin, I.; Teofilovici, F.; Vukovic, V.M.; et al. Randomized Phase III Study of Ganetespib, a Heat Shock Protein 90 Inhibitor, With Docetaxel Versus Docetaxel in Advanced Non-Small-Cell Lung Cancer (GALAXY-2). J. Clin. Oncol. 2020, 38, 613–622. [Google Scholar] [CrossRef]
- Socinski, M.A.; Goldman, J.; El-Hariry, I.; Koczywas, M.; Vukovic, V.; Horn, L.; Paschold, E.; Salgia, R.; West, H.; Sequist, L.V.; et al. A multicenter phase II study of ganetespib monotherapy in patients with genotypically defined advanced non-small cell lung cancer. Clin. Cancer Res. 2013, 19, 3068–3077. [Google Scholar] [CrossRef] [Green Version]
- Rong, B.; Yang, S. Molecular mechanism and targeted therapy of Hsp90 involved in lung cancer: New discoveries and developments (Review). Int. J. Oncol. 2018, 52, 321–336. [Google Scholar] [CrossRef]
- Voruganti, S.; Kline, J.T.; Balch, M.J.; Rogers, J.; Matts, R.L.; Hartson, S.D. Proteomic Profiling of Hsp90 Inhibitors. Methods Mol. Biol. 2018, 1709, 139–162. [Google Scholar]
- Zimmer, A.; Amar-Farkash, S.; Danon, T.; Alon, U. Dynamic proteomics reveals bimodal protein dynamics of cancer cells in response to HSP90 inhibitor. BMC Syst. Biol. 2017, 11, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marrugal, Á.; Ferrer, I.; Pastor, M.D.; Ojeda, L.; Quintanal-Villalonga, Á.; Carnero, A.; Molina-Pinelo, S.; Paz-Ares, L. Impact of Heat Shock Protein 90 Inhibition on the Proteomic Profile of Lung Adenocarcinoma as Measured by Two-Dimensional Electrophoresis Coupled with Mass Spectrometry. Cells 2019, 8, 806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weidenauer, L.; Wang, T.; Joshi, S.; Chiosis, G.; Quadroni, M.R. Proteomic interrogation of HSP90 and insights for medical research. Expert Rev. Proteomics 2017, 14, 1105–1117. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Bhattacharya, S.; Socinski, M.A.; Burns, T.F. HSP90 inhibitors in lung cancer: Promise still unfulfilled. Clin. Adv. Hematol. Oncol. 2016, 14, 346–356. [Google Scholar]
- Mishra, D.; Banerjee, D. Lactate Dehydrogenases as Metabolic Links between Tumor and Stroma in the Tumor Microenvironment. Cancers (Basel) 2019, 11, 750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dennison, J.B.; Molina, J.R.; Mitra, S.; Gonzalez-Angulo, A.M.; Balko, J.M.; Kuba, M.G.; Sanders, M.E.; Pinto, J.A.; Gomez, H.L.; Arteaga, C.L.; et al. Lactate dehydrogenase B: A metabolic marker of response to neoadjuvant chemotherapy in breast cancer. Clin. Cancer Res. 2013, 19, 3703–3713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCleland, M.L.; Adler, A.S.; Deming, L.; Cosino, E.; Lee, L.; Blackwood, E.M.; Solon, M.; Tao, J.; Li, L.; Shames, D.; et al. Lactate dehydrogenase B is required for the growth of KRAS-dependent lung adenocarcinomas. Clin. Cancer Res. 2013, 19, 773–784. [Google Scholar] [CrossRef] [Green Version]
- Hou, G.X.; Liu, P.; Yang, J.; Wen, S. Mining expression and prognosis of topoisomerase isoforms in non-small-cell lung cancer by using Oncomine and Kaplan-Meier plotter. PLoS ONE 2017, 12, e0174515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, B.; Zhang, H.; Zhang, T.; Cai, Y.; Hu, Y.; Zheng, H.; Li, B. Topoisomerase I expression is associated with prognosis in postoperative non-small cell lung cancer patients. Thorac. Cancer 2016, 7, 486–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riedlinger, T.; Bartkuhn, M.; Zimmermann, T.; Hake, S.B.; Nist, A.; Stiewe, T.; Kracht, M.; Schmitz, M.L. Chemotherapeutic Drugs Inhibiting Topoisomerase 1 Activity Impede Cytokine-Induced and NF-kappaB p65-Regulated Gene Expression. Cancers (Basel) 2019, 11, 883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serman, L.; Nikuseva Martic, T.; Serman, A.; Vranic, S. Epigenetic alterations of the Wnt signaling pathway in cancer: A mini review. Bosn. J. Basic Med. Sci. 2014, 14, 191–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Song, X.; Yue, W.; Chen, D.; Yu, J.; Yao, Z.; Zhang, L. Fibulin-5 inhibits Wnt/beta-catenin signaling in lung cancer. Oncotarget 2015, 6, 15022–15034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Remmelink, M.; Mijatovic, T.; Gustin, A.; Mathieu, A.; Rombaut, K.; Kiss, R.; Salmon, I.; Decaestecker, C. Identification by means of cDNA microarray analyses of gene expression modifications in squamous non-small cell lung cancers as compared to normal bronchial epithelial tissue. Int. J. Oncol. 2005, 26, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Iwagami, Y.; Eguchi, H.; Nagano, H.; Akita, H.; Hama, N.; Wada, H.; Kawamoto, K.; Kobayashi, S.; Tomokuni, A.; Tomimaru, Y.; et al. miR-320c regulates gemcitabine-resistance in pancreatic cancer via SMARCC1. Br. J. Cancer 2013, 109, 502–511. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.Q.; Hao, J.J.; Zhang, Z.; Krane, L.S.; Hammerich, K.H.; Sanford, T.; Trepel, J.B.; Neckers, L.; Agarwal, P.K. Proteomic analysis of proteome and histone post-translational modifications in heat shock protein 90 inhibition-mediated bladder cancer therapeutics. Sci. Rep. 2017, 7, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gigek, C.O.; Lisboa, L.C.; Leal, M.F.; Silva, P.N.; Lima, E.M.; Khayat, A.S.; Assumpcao, P.P.; Burbano, R.R.; Smith Mde, A. SMARCA5 methylation and expression in gastric cancer. Cancer Invest. 2011, 29, 162–166. [Google Scholar] [CrossRef]
- Jin, Q.; Mao, X.; Li, B.; Guan, S.; Yao, F.; Jin, F. Overexpression of SMARCA5 correlates with cell proliferation and migration in breast cancer. Tumour. Biol. 2015, 36, 1895–1902. [Google Scholar] [CrossRef]
- Ribeyre, C.; Zellweger, R.; Chauvin, M.; Bec, N.; Larroque, C.; Lopes, M.; Constantinou, A. Nascent DNA Proteomics Reveals a Chromatin Remodeler Required for Topoisomerase I Loading at Replication Forks. Cell Rep. 2016, 15, 300–309. [Google Scholar] [CrossRef] [Green Version]
- Ito, H.; Morishita, R.; Nagata, K.I. Functions of Rhotekin, an Effector of Rho GTPase, and Its Binding Partners in Mammals. Int J. Mol. Sci. 2018, 19, 2121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bishop, A.L.; Hall, A. Rho GTPases and their effector proteins. Biochem. J. 2000, 348, 241–255. [Google Scholar] [CrossRef]
- Memmel, S.; Sisario, D.; Zoller, C.; Fiedler, V.; Katzer, A.; Heiden, R.; Becker, N.; Eing, L.; Ferreira, F.L.R.; Zimmermann, H.; et al. Migration pattern, actin cytoskeleton organization and response to PI3K-, mTOR-, and Hsp90-inhibition of glioblastoma cells with different invasive capacities. Oncotarget 2017, 8, 45298–45310. [Google Scholar] [CrossRef] [Green Version]
- Lopez, L.C.; Schuelke, M.; Quinzii, C.M.; Kanki, T.; Rodenburg, R.J.; Naini, A.; Dimauro, S.; Hirano, M. Leigh syndrome with nephropathy and CoQ10 deficiency due to decaprenyl diphosphate synthase subunit 2 (PDSS2) mutations. Am. J. Hum. Genet. 2006, 79, 1125–1129. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.; Yu, J.; Knecht, J.; Chen, Q. Decrease of PDSS2 expression, a novel tumor suppressor, in non-small cell lung cancer. Cancer. Epidemiol. 2013, 37, 166–171. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Zhang, Y.; Polireddy, K.; Chen, Q. The tumor-suppressing activity of the prenyl diphosphate synthase subunit 2 gene in lung cancer cells. Anticancer Drugs 2014, 25, 790–798. [Google Scholar] [CrossRef] [PubMed]
- Brea-Calvo, G.; Rodriguez-Hernandez, A.; Fernandez-Ayala, D.J.; Navas, P.; Sanchez-Alcazar, J.A. Chemotherapy induces an increase in coenzyme Q10 levels in cancer cell lines. Free Radic. Biol. Med. 2006, 40, 1293–1302. [Google Scholar] [CrossRef]
- HSP90 Interactors. Available online: https://www.picard.ch/downloads/Hsp90interactors.pdf (accessed on 20 May 2020).
- Jahani, M.; Noroznezhad, F.; Mansouri, K. Arginine: Challenges and opportunities of this two-faced molecule in cancer therapy. Biomed. Pharmacother. 2018, 102, 594–601. [Google Scholar] [CrossRef] [PubMed]
- Keshet, R.; Erez, A. Arginine and the metabolic regulation of nitric oxide synthesis in cancer. Dis. Model. Mech 2018, 11. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.J.; Galoforo, S.S.; Berns, C.M.; Chen, J.C.; Davis, B.H.; Sim, J.E.; Corry, P.M.; Spitz, D.R. Glucose deprivation-induced cytotoxicity and alterations in mitogen-activated protein kinase activation are mediated by oxidative stress in multidrug-resistant human breast carcinoma cells. J. Biol. Chem. 1998, 273, 5294–5299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joly, J.H.; Delfarah, A.; Phung, P.S.; Parrish, S.; Graham, N.A. A synthetic lethal drug combination mimics glucose deprivation-induced cancer cell death in the presence of glucose. J. Biol. Chem. 2020, 295, 1350–1365. [Google Scholar] [CrossRef] [PubMed]
- Simons, A.L.; Mattson, D.M.; Dornfeld, K.; Spitz, D.R. Glucose deprivation-induced metabolic oxidative stress and cancer therapy. J. Cancer Res. Ther. 2009, 5, S2–S6. [Google Scholar] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Geldanamycin Derivatives | Radicicol Derivatives | ||
|---|---|---|---|---|
| IC50 17-AAG (nM) | IC50 IPI-504 (nM) | IC50 STA-9090 (nM) | IC50 AUY-922 (nM) | |
| HCC827 | 26.255 | 17.145 | 5.138 | 4.167 |
| H1975 | 1.258 | 12.750 | 4.739 | 2.595 |
| H1650 | 6.554 | 3.764 | 5.659 | 1.472 |
| H3122 | 26.165 | 28.371 | 7.991 | 9.110 |
| H2228 | 10.888 | 46.340 | 4.131 | 4.488 |
| H2009 | 43.198 | 33.833 | 4.659 | 2.477 |
| H358 | 13.066 | 4.662 | 7.740 | 8.105 |
| A549 | 16.296 | 19.492 | 6.310 | 30.733 |
| H1437 | 3.708 | 3.473 | 6.794 | 2.814 |
| CALU-3 | 87.733 | 43.295 | 18.445 | 1740.91 |
| H1781 | 12.345 | 30.975 | 9.954 | 23.787 |
| Treatment | Response | Number of Response-Related Proteins | |
|---|---|---|---|
| Detected | Significant | ||
| 17-AAG | Sensitivity | 3724 | 27 |
| Resistance | 3607 | 171 | |
| IPI-504 | Sensitivity | 3898 | 1664 |
| Resistance | 4395 | 1772 | |
| STA-9090 | Sensitivity | 4497 | 51 |
| Resistance | 3704 | 21 | |
| AUY-922 | Sensitivity | 4149 | 1294 |
| Resistance | 3934 | 1257 | |
| Protein Name | UniProt 1 | Gene 2 |
|---|---|---|
| Spliceosome RNA helicase DDX39B | Q13838 | DDX39B |
| 26S proteasome non-ATPase regulatory subunit 7 | P51665 | PSMD7 |
| 3-hydroxyisobutyryl-CoA hydrolase, mitochondrial | Q6NVY1 | HIBCH |
| 5′(3′)-deoxyribonucleotidase, cytosolic type | Q8TCD5 | NT5C |
| 5′-AMP-activated protein kinase catalytic subunit alpha-1 | Q13131 | PRKAA1 |
| 5′-nucleotidase domain containing 1 | Q5TFE4 | NT5DC1 |
| Acyl-coenzyme A thioesterase 13 | Q9NPJ3 | ACOT13 |
| Acylglycerol kinase, mitochondrial | Q53H12 | AGK |
| Ankyrin repeat and LEM domain-containing protein 2 | Q86XL3 | ANKLE2 |
| ATP-binding cassette sub-family D member 3 | P28288 | ABCD3 |
| Cleavage stimulation factor subunit 2 tau variant | Q9H0L4 | CSTF2T |
| Coiled-coil domain containing 12 | Q8WUD4 | CCDC12 |
| Conserved oligomeric Golgi complex subunit 1 | Q8WTW3 | COG1 |
| DNA topoisomerase 1 | P11387 | TOP1 |
| Dolichyl-diphosphooligosaccharide--protein glycosyltransferase subunit STT3B | Q8TCJ2 | STT3B |
| Dynamin-like 120 kDa protein, mitochondrial | O60313 | OPA1 |
| EH domain-containing protein 4 | Q9H223 | EHD4 |
| Elongation factor 1-alpha 2 | Q05639 | EEF1A2 |
| Epiplakin | P58107 | EPPK1 |
| Eukaryotic initiation factor 4A-III | P38919 | EIF4A3 |
| Eukaryotic translation initiation factor 4 gamma 1 | Q04637 | EIF4G1 |
| Eukaryotic translation initiation factor 4 gamma 3 | O43432 | EIF4G3 |
| Formin-binding protein 1-like | Q5T0N5 | FNBP1L |
| Haloacid dehalogenase like hydrolase domain containing 3 | Q9BSH5 | HDHD3 |
| Haloacid dehalogenase-like hydrolase domain-containing 5 | Q9BXW7 | CECR5 |
| Heat shock protein 105 kDa and 70 kDa proteins | Q92598 | HSPH1 |
| Heat shock protein family A member 4 | P34932 | HSPA4 |
| Heme-binding protein 2 | Q9Y5Z4 | HEBP2 |
| Hepatoma-derived growth factor-related protein 3 | Q9Y3E1 | HDGFRP3 |
| High mobility group protein B1 | P09429 | HMGB1 |
| Hydroxymethylglutaryl-CoA synthase, cytoplasmic | Q01581 | HMGCS1 |
| Isocitrate dehydrogenase [NAD] subunit alpha, mitochondrial | P50213 | IDH3A |
| Isocitrate dehydrogenase [NADP], mitochondrial | P48735 | IDH2 |
| Magnesium transporter protein 1 | Q9H0U3 | MAGT1 |
| Mediator of RNA polymerase II transcription subunit 14 | O60244 | MED14 |
| Mitotic interactor and substrate of PLK1 | Q8IVT2 | MISP |
| Myosin regulatory light chain 12A | P19105 | MYL12A |
| NADH-ubiquinone oxidoreductase 75 kDa subunit, mitochondrial | P28331 | NDUFS1 |
| Nitric oxide synthase-interacting protein | Q9Y314 | NOSIP |
| Non-POU domain-containing octamer-binding protein | Q15233 | NONO |
| Nucleoside diphosphate-linked moiety X motif 19 | A8MXV4 | NUDT19 |
| Paraspeckle component 1 | Q8WXF1 | PSPC1 |
| PHD finger protein 14 | O94880 | PHF14 |
| Pogo transposable element with ZNF domain | Q7Z3K3 | POGZ |
| Probable ATP-dependent RNA helicase DDX6 | P26196 | DDX6 |
| Prolyl endopeptidase | P48147 | PREP |
| Protein S100-A13 | Q99584 | S100A13 |
| Putative oxidoreductase GLYR1 | Q49A26 | GLYR1 |
| Ras-related GTP-binding protein A | Q7L523 | RRAGA |
| Small nuclear ribonucleoprotein E | P62304 | SNRPE |
| SWI/SNF complex subunit SMARCC1 | Q92922 | SMARCC1 |
| SWI/SNF complex subunit SMARCC2 | Q8TAQ2 | SMARCC2 |
| SWI/SNF-related matrix-associated actin-dependent regulator of chromatin subfamily A member 5 | O60264 | SMARCA5 |
| T-complex protein 1 subunit beta; | P78371 | CCT2 |
| T-complex protein 1 subunit epsilon | P48643 | CCT5 |
| Transcription initiation factor TFIID subunit 4 | O00268 | TAF4 |
| Transcriptional repressor p66-alpha | Q86YP4 | GATAD2A |
| Transcriptional repressor p66-beta | Q8WXI9 | GATAD2B |
| Transducin beta-like protein 3; | Q12788 | TBL3 |
| Tyrosine-protein phosphatase non-receptor type 11 | Q06124 | PTPN11 |
| UBX domain-containing protein 7 | O94888 | UBXN7 |
| Sensitivity | Resistance |
|---|---|
| Arginine biosynthesis | Adrenaline and noradrenaline biosynthesis |
| Endogenous cannabinoid signaling | Alzheimer’s disease–amyloid secretase pathway |
| GABA-B receptor II signaling | Blood coagulation |
| Gonadotropin-releasing hormone receptor pathway | Coenzyme A biosynthesis |
| Opioid proenkephalin pathway | General transcription regulation |
| Opioid proopiomelanocortin pathway | Ornithine degradation |
| p53 pathway by glucose deprivation | |
| Purine metabolism | |
| Toll receptor signaling pathway | |
| Vasopressin synthesis |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marrugal, Á.; Ferrer, I.; Gómez-Sánchez, D.; Quintanal-Villalonga, Á.; Pastor, M.D.; Ojeda, L.; Paz-Ares, L.; Molina-Pinelo, S. Identification of Predictive Biomarkers of Response to HSP90 Inhibitors in Lung Adenocarcinoma. Int. J. Mol. Sci. 2021, 22, 2538. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052538
Marrugal Á, Ferrer I, Gómez-Sánchez D, Quintanal-Villalonga Á, Pastor MD, Ojeda L, Paz-Ares L, Molina-Pinelo S. Identification of Predictive Biomarkers of Response to HSP90 Inhibitors in Lung Adenocarcinoma. International Journal of Molecular Sciences. 2021; 22(5):2538. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052538
Chicago/Turabian StyleMarrugal, Ángela, Irene Ferrer, David Gómez-Sánchez, Álvaro Quintanal-Villalonga, María Dolores Pastor, Laura Ojeda, Luis Paz-Ares, and Sonia Molina-Pinelo. 2021. "Identification of Predictive Biomarkers of Response to HSP90 Inhibitors in Lung Adenocarcinoma" International Journal of Molecular Sciences 22, no. 5: 2538. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052538