
Comprehensive Analysis of Correlations in the Expression of miRNA Genes and Immune Checkpoint Genes in Bladder Cancer Cells

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results
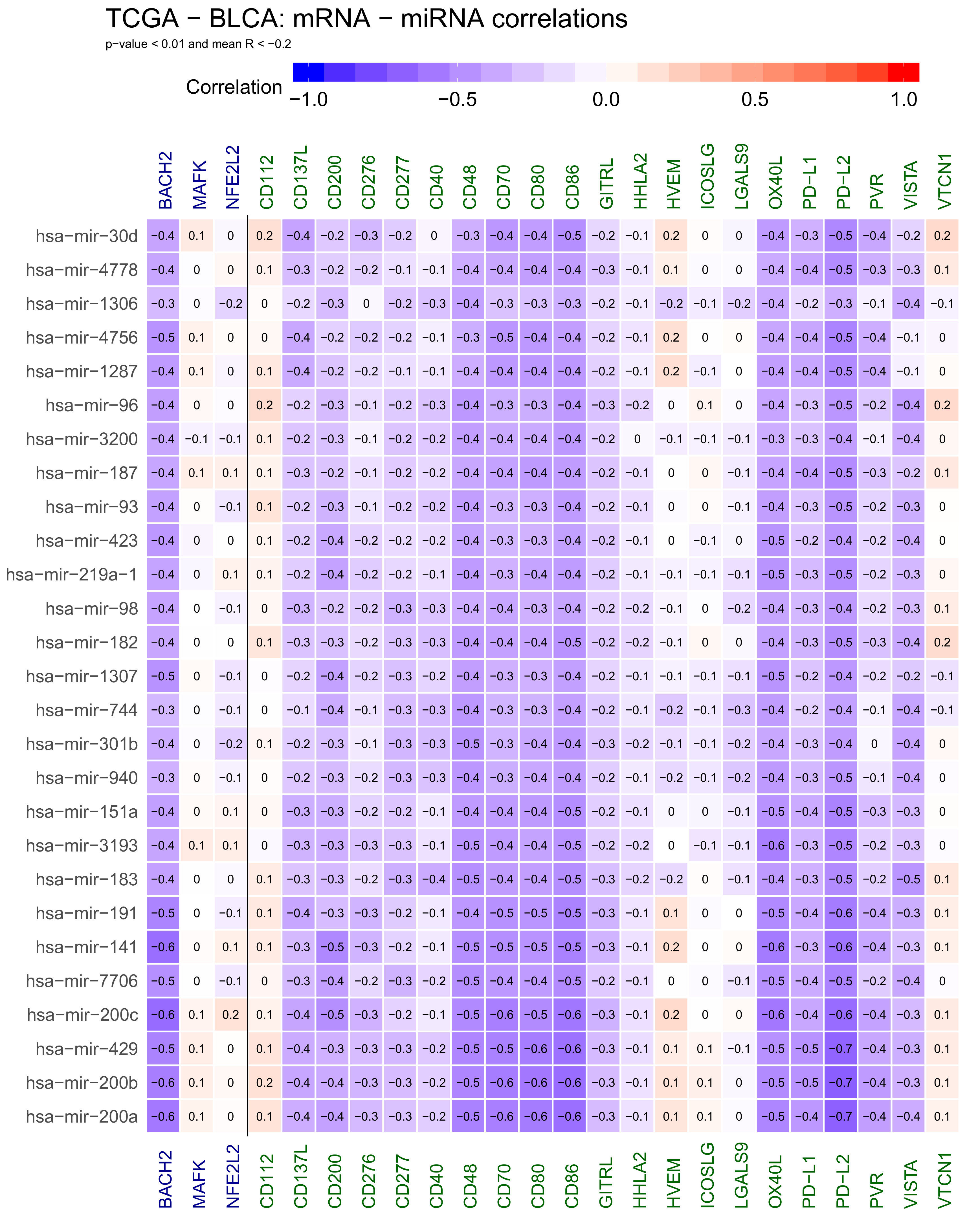
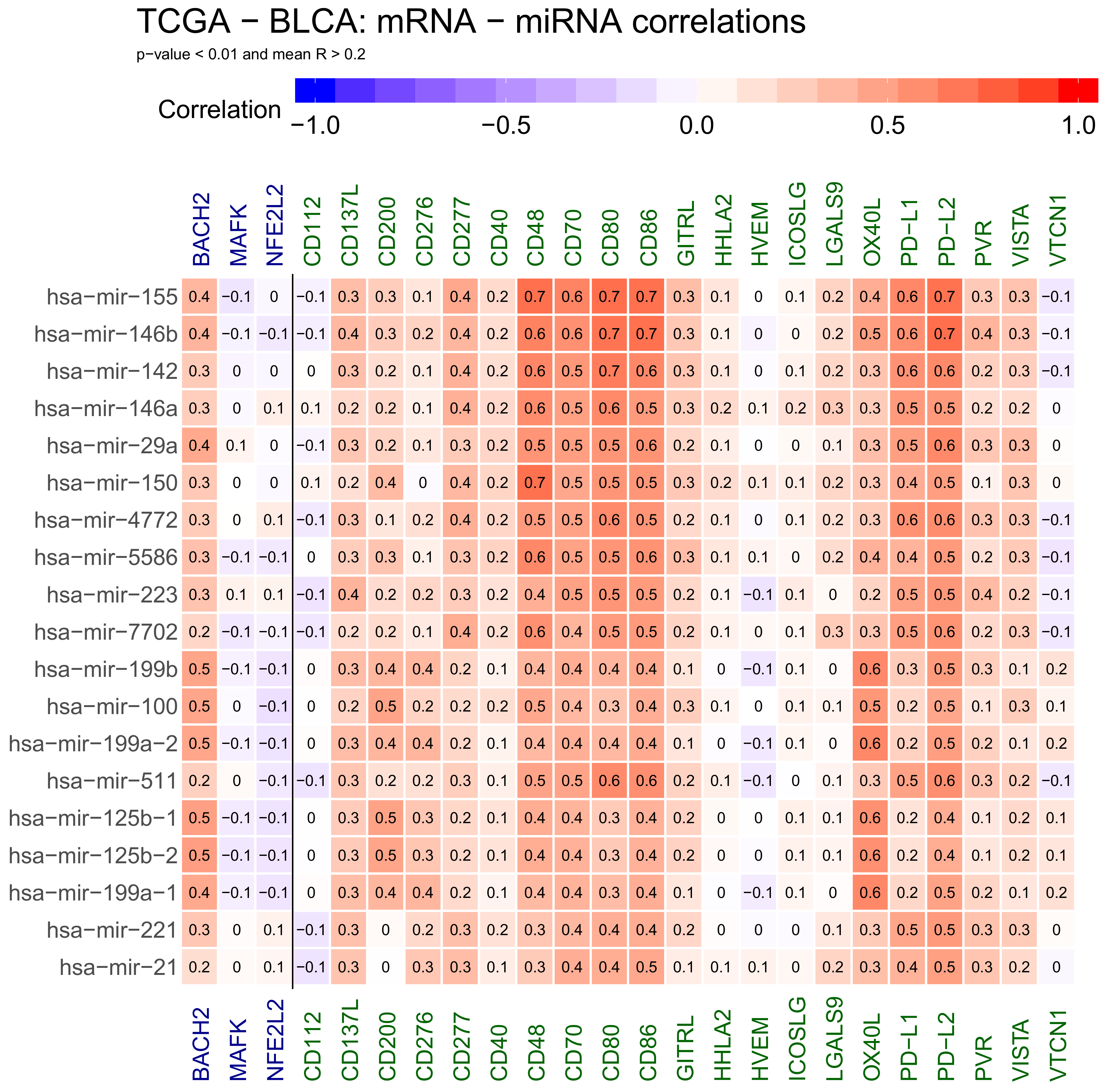
2.1. Correlations Analyses in Bladder Cancer Samples—Genes mRNA to miRNA
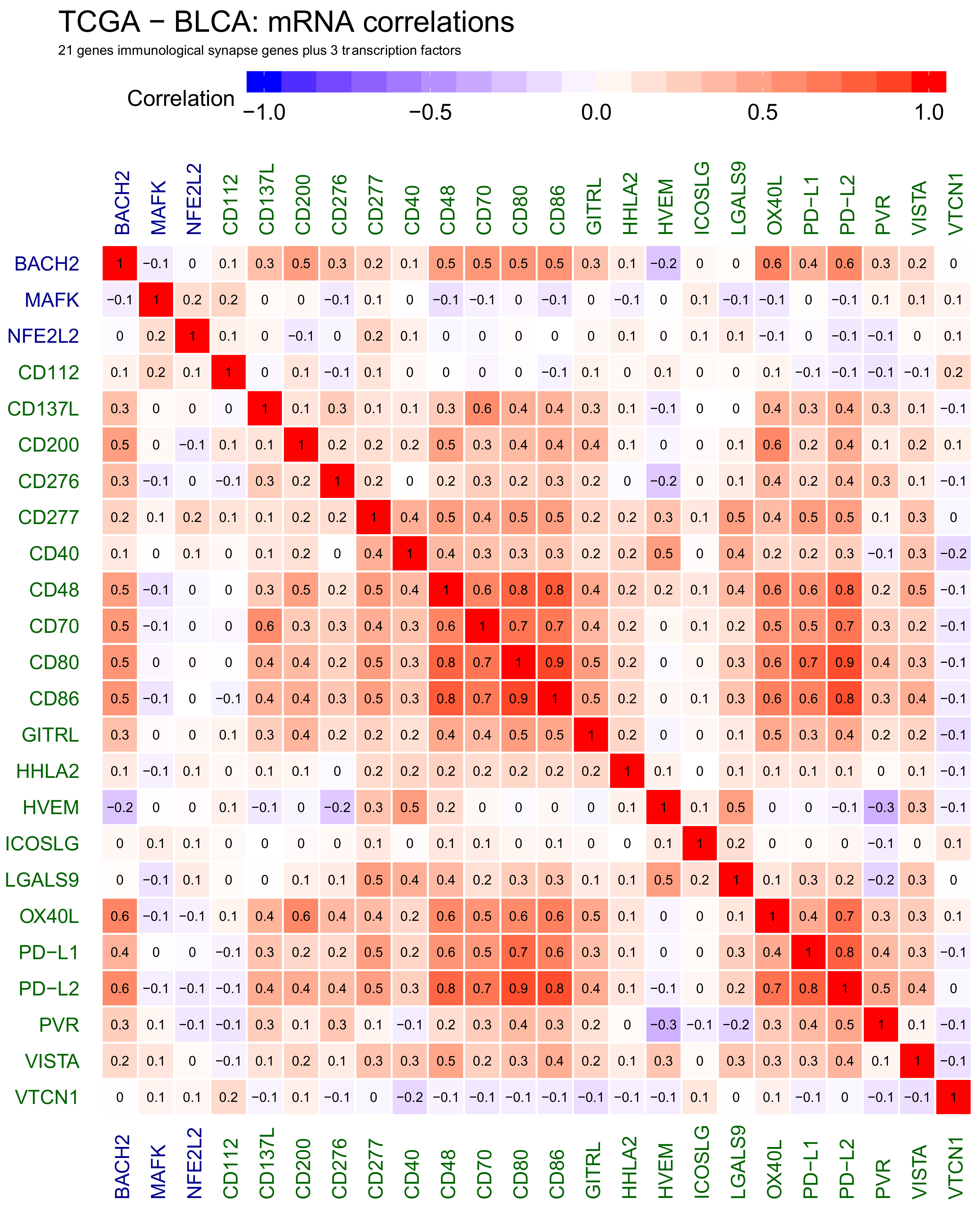
2.2. Correlations Analyses in Bladder Cancer Samples—Genes to Genes
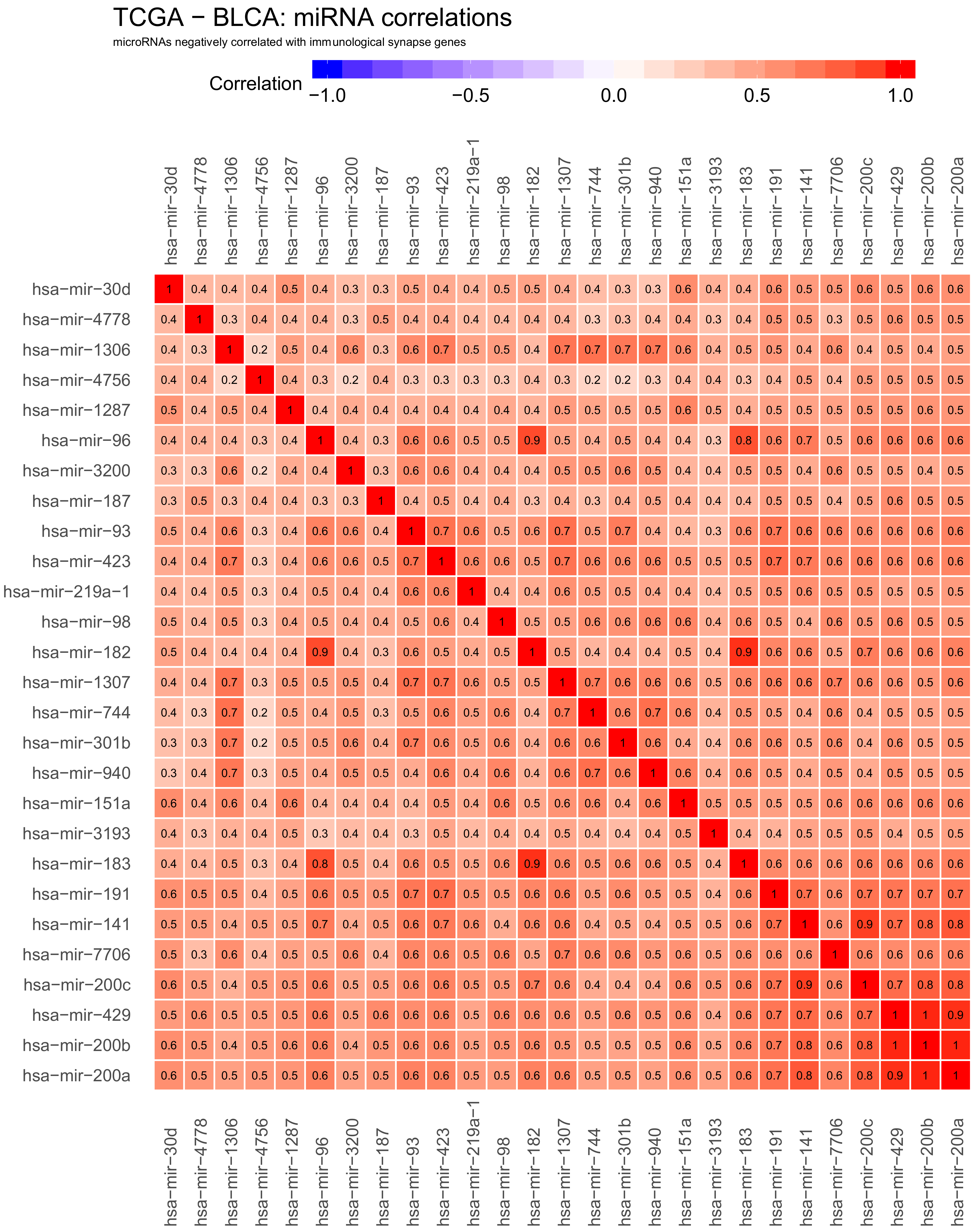
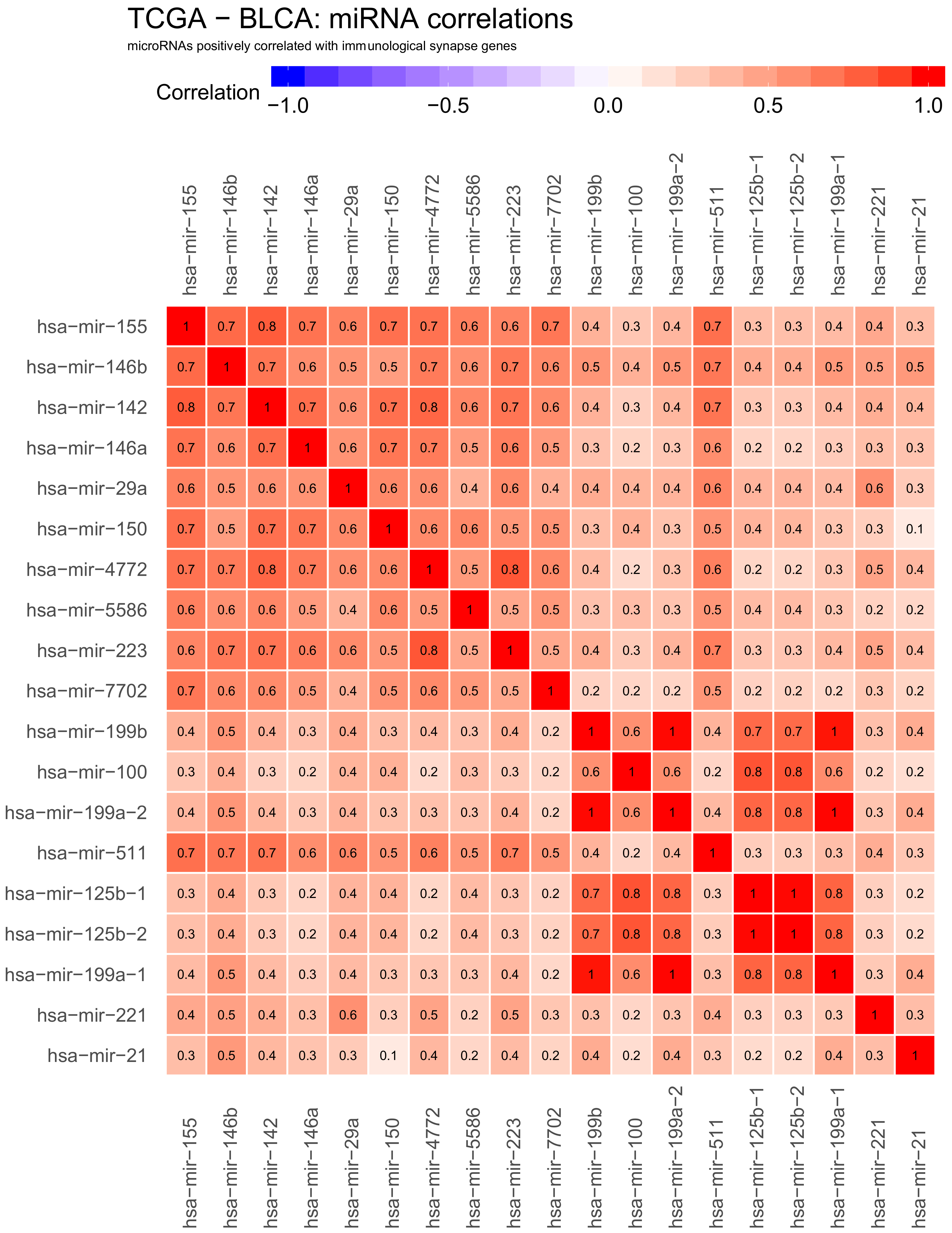
2.3. Correlations Analyses in Bladder Cancer Samples—miRNA to miRNA
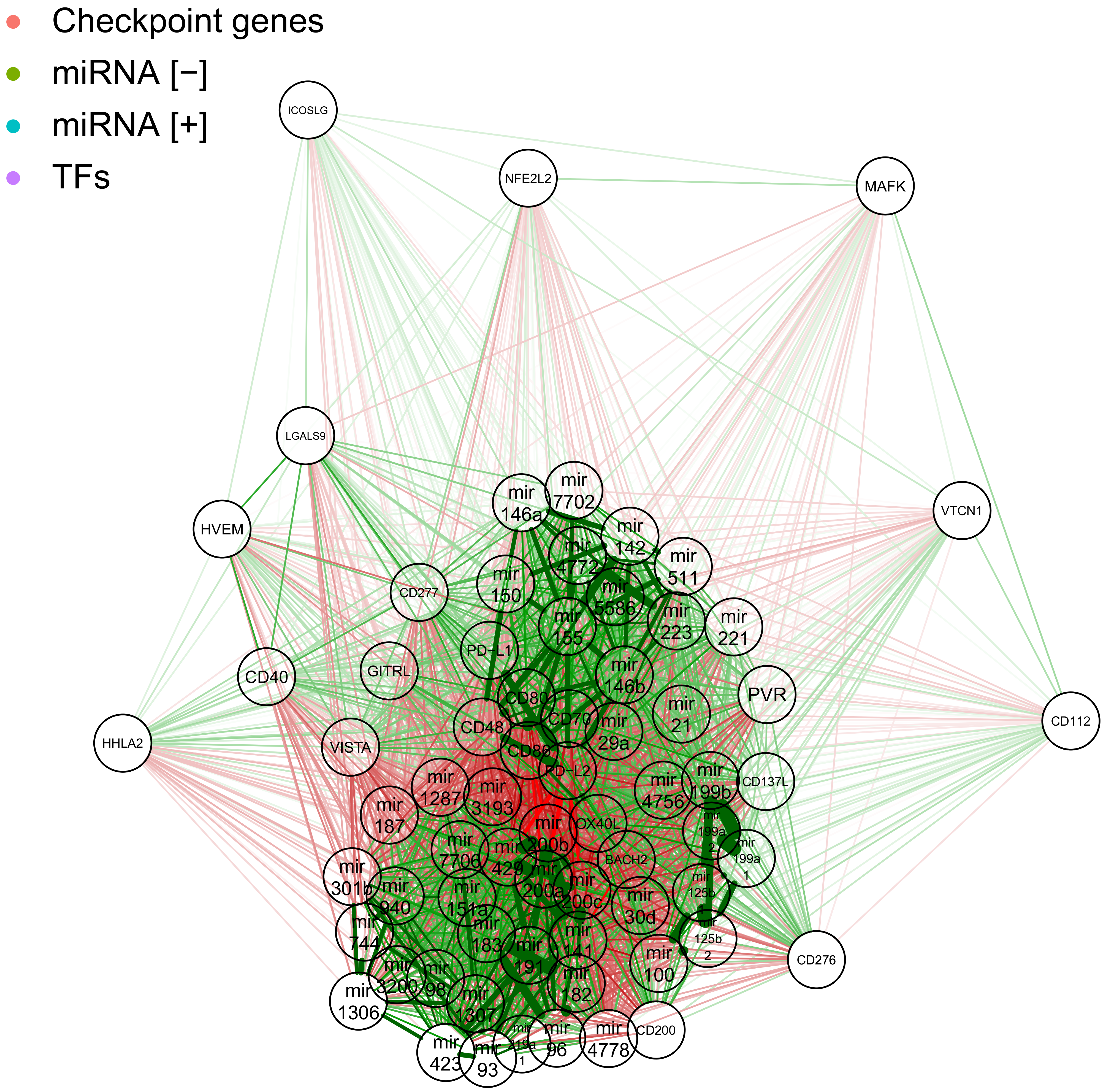
2.4. Network of Correlation Derived Interactions between miRNAs, Checkpoint Genes and TFs
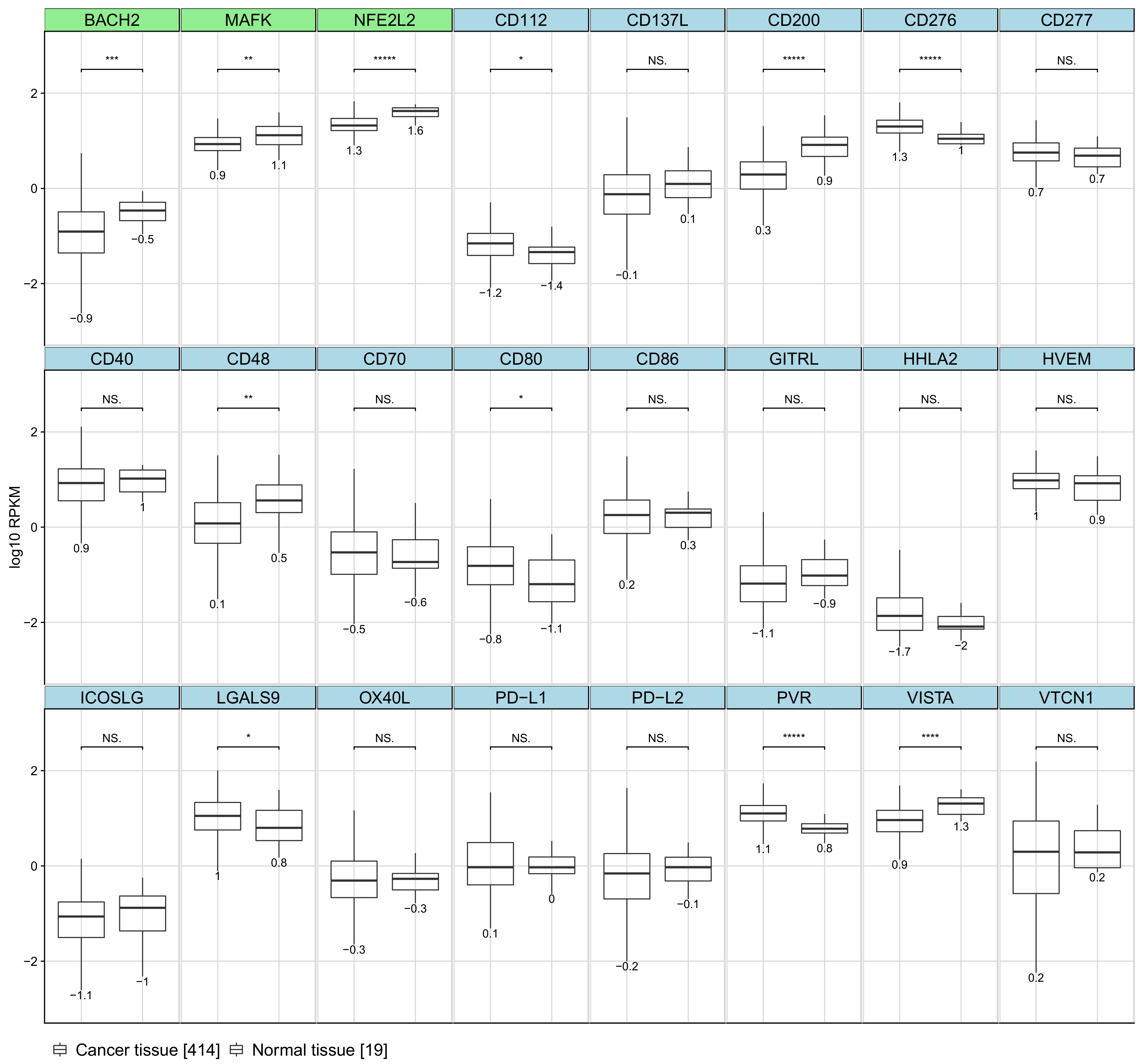
2.5. Immune Synapse Genes Are Deregulated in Breast Cancer
3. Discussion
3.1. The Most Positively Correlated miRNAs
3.2. The Most Anti-Correlated miRNAs
3.3. Other miRNAs Important in Bladder Cancer
3.4. Limitations to the Research Presented
4. Materials and Methods
4.1. Data Acquisition and Pre-Processing
4.2. Correlation Analyses
4.3. Graphical Model Estimation for Correlation Networks
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- MedlinePlus Genetics Home Reference—What Is Precision Medicine? Available online: https://medlineplus.gov/about (accessed on 8 January 2021).
- Smith, R. Stratified, Personalised, or Precision Medicine. Available online: https://0-blogs-bmj-com.brum.beds.ac.uk/bmj/2012/10/15/richard-smith-stratified-personalised-or-precision-medicine (accessed on 8 January 2021).
- O’Shaughnessy, K. Lecture on Stratified Medicine; Cambridge University: Cambridge, UK, 2017. [Google Scholar]
- Nordentoft, I.; Dyrskjøt, L.; Bødker, J.S.; Wild, P.J.; Hartmann, A.; Bertz, S.; Lehmann, J.; Ørntoft, T.F.; Birkenkamp-Demtroder, K. Increased Expression of Transcription Factor TFAP2 Correlates with Chemosensitivity in Advanced Bladder Cancer. BMC Cancer 2011, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faltas, B.M.; Prandi, D.; Tagawa, S.T.; Molina, A.M.; Nanus, D.M.; Sternberg, C.; Rosenberg, J.; Mosquera, J.M.; Robinson, B.; Elemento, O.; et al. Clonal Evolution of Chemotherapy-Resistant Urothelial Carcinoma. Nat. Genet. 2016, 48, 1490–1499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hornstein, E.; Shomron, N. Canalization of Development by MicroRNAs. Nat. Genet. 2006, 38, S20–S24. [Google Scholar] [CrossRef]
- Calin, G.A.; Cimmino, A.; Fabbri, M.; Ferracin, M.; Wojcik, S.E.; Shimizu, M.; Taccioli, C.; Zanesi, N.; Garzon, R.; Aqeilan, R.I.; et al. MiR-15a and MiR-16-1 Cluster Functions in Human Leukemia. Proc. Natl. Acad. Sci. USA 2008, 105, 5166–5171. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Ji, Z.; Xie, Y.; Liu, G.; Li, H. MicroRNA-154 as a Prognostic Factor in Bladder Cancer Inhibits Cellular Malignancy by Targeting RSF1 and RUNX2. Oncol. Rep. 2017, 38, 2727–2734. [Google Scholar] [CrossRef] [Green Version]
- Aqeilan, R.I.; Calin, G.A.; Croce, C.M. MiR-15a and MiR-16-1 in Cancer: Discovery, Function and Future Perspectives. Cell Death Differ. 2009, 17, 215–220. [Google Scholar] [CrossRef] [Green Version]
- Pecorino, L. Molecular Biology of Cancer, 4th ed.; Oxford University Press: Oxford, UK, 2016. [Google Scholar]
- Esquela-Kerscher, A.; Slack, F.J. Oncomirs MicroRNAs with a Role in Cancer. Nat. Rev. Cancer 2006, 6, 259–269. [Google Scholar] [CrossRef]
- Cimmino, A.; Calin, G.A.; Fabbri, M.; Iorio, M.V.; Ferracin, M.; Shimizu, M.; Wojcik, S.E.; Aqeilan, R.I.; Zupo, S.; Dono, M.; et al. MiR-15 and MiR-16 Induce Apoptosis by Targeting BCL2. Proc. Natl. Acad. Sci. USA 2005, 102, 13944–13949. [Google Scholar] [CrossRef] [Green Version]
- Messing, E.M. Checkpoint Inhibitors for Advanced Bladder Cancer. Bladder Cancer 2016, 2, 473–474. [Google Scholar] [CrossRef] [Green Version]
- Knowles, M.A.; Hurst, C.D. Molecular Biology of Bladder Cancer: New Insights into Pathogenesis and Clinical Diversity. Nat. Rev. Cancer 2014, 15, 25–41. [Google Scholar] [CrossRef]
- Shien, K.; Papadimitrakopoulou, V.A.; Wistuba, I.I. Predictive Biomarkers of Response to PD-1/PD-L1 Immune Checkpoint Inhibitors in Nonsmall Cell Lung Cancer. Lung Cancer 2016, 99, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Dobosz, P.; Stempor, P.A.; Roszik, J.; Herman, A.; Layani, A.; Berger, R.; Avni, D.; Sidi, Y.; Leibowitz-Amit, R. Checkpoint Genes at the Cancer Side of the Immunological Synapse in Bladder Cancer. Transl. Oncol. 2020, 13. [Google Scholar] [CrossRef] [PubMed]
- Robertson, E.D.; Wasylyk, C.; Ye, T.; Jung, A.C.; Wasylyk, B. The Oncogenic MicroRNA Hsa-MiR-155-5p Targets the Transcription Factor ELK3 and Links It to the Hypoxia Response. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [Green Version]
- Pasculli, B.; Barbano, R.; Fontana, A.; Biagini, T.; di Viesti, M.P.; Rendina, M.; Valori, V.M.; Morritti, M.; Bravaccini, S.; Ravaioli, S.; et al. Hsa-MiR-155-5p Up-Regulation in Breast Cancer and Its Relevance for Treatment with Poly[ADP-Ribose] Polymerase 1 (PARP-1) Inhibitors. Front. Oncol. 2020, 10. [Google Scholar] [CrossRef]
- Peng, Y.; Dong, W.; Lin, T.; Zhong, G.; Liao, B.; Wang, B.; Gu, P.; Huang, L.; Xie, Y.; Lu, F.; et al. MicroRNA-155 Promotes Bladder Cancer Growth by Repressing the Tumor Suppressor DMTF1. Oncotarget 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Xu, J.; Li, H.; Sun, C.; Yu, L.; Li, Y.; Shi, C.; Zhou, X.; Bian, X.; Ping, Y.; et al. MiR-146b-5p Functions as a Tumor Suppressor by Targeting TRAF6 and Predicts the Prognosis of Human Gliomas. Oncotarget 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Xu, C.; Ruan, L.; Wu, J.; Li, Y.; Zhang, X. MicroRNA-146b Overexpression Promotes Human Bladder Cancer Invasion via Enhancing ETS2-Mediated Mmp2 MRNA Transcription. Mol. Nucleic Acids 2019, 16. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Yang, Y.; Wang, Z.; Zhang, X.; Li, D.; Wei, J. A SNP of MiR-146a Is Involved in Bladder Cancer Relapse by Affecting the Function of Bladder Cancer Stem Cells via the MiR-146a Signallings. J. Cell. Mol. Med. 2020, 24. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lu, H.; Li, T.; Yu, L.; Liu, G.; Peng, X.; Zhao, J. Krüppel-like Factor 8 Promotes Tumorigenic Mammary Stem Cell Induction by Targeting MiR-146a. Am. J. Cancer Res. 2013, 3, 356–373. [Google Scholar] [PubMed]
- Sasaki, H.; Yoshiike, M.; Nozawa, S.; Usuba, W.; Katsuoka, Y.; Aida, K.; Kitajima, K.; Kudo, H.; Hoshikawa, M.; Yoshioka, Y.; et al. Expression Level of Urinary MicroRNA-146a-5p Is Increased in Patients With Bladder Cancer and Decreased in Those After Transurethral Resection. Clin. Genitourin. Cancer 2016, 14. [Google Scholar] [CrossRef] [PubMed]
- Roos, J.; Enlund, E.; Funcke, J.-B.; Tews, D.; Holzmann, K.; Debatin, K.-M.; Wabitsch, M.; Fischer-Posovszky, P. MiR-146a-Mediated Suppression of the Inflammatory Response in Human Adipocytes. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [Green Version]
- Dekkema, G.J.; Bijma, T.; Jellema, P.G.; van den Berg, A.; Kroesen, B.-J.; Stegeman, C.A.; Heeringa, P.; Abdulahad, W.H.; Sanders, J.-S. Increased MiR-142-3p Expression Might Explain Reduced Regulatory T Cell Function in Granulomatosis with Polyangiitis. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Li, W.Q.; Zhao, W.C.; Xin, J.; Niu, T.L.; Chao, Y.F.; Zhou, P.; Zheng, M.H.; Xu, B. MicroRNA-142-3p Suppresses Cell Proliferation and Migration in Bladder Cancer via Rac1. J. Biol. Regul. Homeost. Agents 2020, 34. [Google Scholar] [CrossRef]
- Zhang, W.; Chen, J.; He, G.; Xu, W.; He, G. Impact of Mirna-21 on Survival Prognosis in Patients with Pancreatic Cancer. Medicine 2020, 99. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.-H.; Tian, D.; Yang, Z.-C.; Li, J.-L. Exosomal MiR-21 Promotes Proliferation, Invasion and Therapy Resistance of Colon Adenocarcinoma Cells through Its Target PDCD4. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef]
- Sahraei, M.; Chaube, B.; Liu, Y.; Sun, J.; Kaplan, A.; Price, N.L.; Ding, W.; Oyaghire, S.; García-Milian, R.; Mehta, S.; et al. Suppressing MiR-21 Activity in Tumor-Associated Macrophages Promotes an Antitumor Immune Response. J. Clin. Investig. 2019, 129. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Yuan, L.; Wang, G.; Cao, R.; Peng, J.; Shu, B.; Qian, G.; Wang, X.; Xiao, Y. Identification and Bioinformatics Analysis of MiRNAs Associated with Human Muscle Invasive Bladder Cancer. Mol. Med. Rep. 2017, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavallari, I.; Grassi, A.; del Bianco, P.; Aceti, A.; Zaborra, C.; Sharova, E.; Bertazzolo, I.; D’Agostino, D.M.; Iafrate, M.; Ciminale, V. Prognostic Stratification of Bladder Cancer Patients with a MicroRNA-Based Approach. Cancers 2020, 12, 3133. [Google Scholar] [CrossRef]
- Jiang, H.; Zhang, G.; Wu, J.-H.; Jiang, C.-P. Diverse Roles of MiR-29 in Cancer (Review). Oncol. Rep. 2014, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weissmann-Brenner, A.; Kushnir, M.; Lithwick Yanai, G.; Aharonov, R.; Gibori, H.; Purim, O.; Kundel, Y.; Morgenstern, S.; Halperin, M.; Niv, Y.; et al. Tumor MicroRNA-29a Expression and the Risk of Recurrence in Stage II Colon Cancer. Int. J. Oncol. 2012. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Song, X.; Du, H.; Luo, C.; Wang, X.; Yang, X.; Wang, Y.; Wu, X. Down-Regulation of MiR-29c in Human Bladder Cancer and the Inhibition of Proliferation in T24 Cell via PI3K-AKT Pathway. Med. Oncol. 2014, 31. [Google Scholar] [CrossRef]
- Cai, J.P.; Zhou, X.H.; Yu, H.B.; Li, D.Y.; Zhou, B.X. Study on the Clinical Significance of MiR-100 Expression in the Invasion and Metastasis of Hepatocellular Carcinoma. Chin. J. Hepatol. 2020, 28, 930–935. [Google Scholar]
- Chen, T.; Shao, S.; Li, W.; Liu, Y.; Cao, Y. The Circular RNA Hsa-Circ-0072309 Plays Anti-Tumour Roles by Sponging MiR-100 through the Deactivation of PI3K/AKT and MTOR Pathways in the Renal Carcinoma Cell Lines. Artif. Cells Nanomed. Biotechnol. 2019, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, Y.; Yang, X.; Hu, S.; Kang, M.; Chen, J.; Fang, Y. A Genetic Variant in MiR-100 Is a Protective Factor of Childhood Acute Lymphoblastic Leukemia. Cancer Med. 2019, 8. [Google Scholar] [CrossRef]
- Wu, N.; Lin, X.; Zhao, X.; Zheng, L.; Xiao, L.; Liu, J.; Ge, L.; Cao, S. MiR-125b Acts as an Oncogene in Glioblastoma Cells and Inhibits Cell Apoptosis through P53 and P38MAPK-Independent Pathways. Br. J. Cancer 2013, 109. [Google Scholar] [CrossRef]
- Wu, M.; Tan, X.; Liu, P.; Yang, Y.; Huang, Y.; Liu, X.; Meng, X.; Yu, B.; Wu, Y.; Jin, H. Role of Exosomal MicroRNA-125b-5p in Conferring the Metastatic Phenotype among Pancreatic Cancer Cells with Different Potential of Metastasis. Life Sci. 2020, 255. [Google Scholar] [CrossRef]
- Yu, Z.; Ni, F.; Chen, Y.; Zhang, J.; Cai, J.; Shi, W. MiR-125b Suppresses Cell Proliferation and Metastasis by Targeting HAX-1 in Esophageal Squamous Cell Carcinoma. Pathol. Res. Pract. 2020, 216. [Google Scholar] [CrossRef] [PubMed]
- Rasheed, Z.; Rasheed, N.; Abdulmonem, W.A.; Khan, M.I. MicroRNA-125b-5p Regulates IL-1β Induced Inflammatory Genes via Targeting TRAF6-Mediated MAPKs and NF-KB Signaling in Human Osteoarthritic Chondrocytes. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Wang, J.; Li, J.; Wang, X.; Song, W. MicroRNA-150 Promotes Cell Proliferation, Migration, and Invasion of Cervical Cancer through Targeting PDCD4. Biomed. Pharmacother. 2018, 97. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.; Hu, X.; Li, B.; Peng, M.; Tong, S.; Zu, X.; Wang, Z.; Qi, L.; Chen, M. MiR-150 Modulates Cisplatin Chemosensitivity and Invasiveness of Muscle-Invasive Bladder Cancer Cells via Targeting PDCD4 In Vitro. Med. Sci. Monit. 2014, 20. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.-L.; Lin, C.-H.; Li, A.-L.; Huang, C.-C.; Shen, B.-Y.; Chiang, Y.-R.; Fang, P.-L.; Chang, H.-C.; Li, K.-L.; Yang, W.-C.; et al. Plasma MiRNA Profile Is a Biomarker Associated with Urothelial Carcinoma in Chronic Hemodialysis Patients. Am. J. Physiol. Ren. Physiol. 2019, 316. [Google Scholar] [CrossRef]
- Lian, J.; Lin, S.-H.; Ye, Y.; Chang, D.W.; Huang, M.; Dinney, C.P.; Wu, X. Serum MicroRNAs as Predictors of Risk for Non-Muscle Invasive Bladder Cancer. Oncotarget 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luan, J.; Fu, J.; Chen, C.; Jiao, C.; Kong, W.; Zhang, Y.; Chang, Q.; Wang, Y.; Li, D.; Illei, G.G.; et al. LNA-Anti-MiR-150 Ameliorated Kidney Injury of Lupus Nephritis by Inhibiting Renal Fibrosis and Macrophage Infiltration. Arthritis Res. Ther. 2019, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, D.; Shen, B.; Wang, W.; Zhou, Y.; Yang, X.; Lu, G.; Yang, J.; Shao, Y. MicroRNA-199a-5p Functions as a Tumor Suppressor in Oral Squamous Cell Carcinoma via Targeting the IKKβ/NF-κB Signaling Pathway. Int. J. Mol. Med. 2019. [Google Scholar] [CrossRef] [Green Version]
- Su, W.Z.; Ren, L.F. MiRNA-199 Inhibits Malignant Progression of Lung Cancer through Mediating RGS17. Eur. Rev. Med Pharmacol. Sci. 2019, 23, 3390–3400. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, W.-X.; Huang, X.-G. MicroRNA-199a-3p Inhibits Angiogenesis by Targeting the VEGF/PI3K/AKT Signalling Pathway in an in Vitro Model of Diabetic Retinopathy. Exp. Mol. Pathol. 2020, 116. [Google Scholar] [CrossRef]
- Tan, H.Y.; Zheng, Y.B.; Liu, J. Serum MiR-199a as a Potential Diagnostic Biomarker for Detection of Colorectal Cancer. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 8657–8663. [Google Scholar] [CrossRef]
- Zhang, H.; Qin, D.; Jiang, Z.; Zhang, J. SNHG9/MiR-199a-5p/Wnt2 Axis Regulates Cell Growth and Aerobic Glycolysis in Glioblastoma. J. Neuropathol. Exp. Neurol. 2019, 78. [Google Scholar] [CrossRef] [PubMed]
- Li, G.L.; Yuan, J.H.; Zhuang, G.D.; Wu, D.Q. MiR-199b Exerts Tumor Suppressive Functions in Hepatocellular Carcinoma by Directly Targeting JAG1. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 7679–7687. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.; Wang, F.-B.; Li, Y.; Zeng, X.-T. Down-Regulation of MiR-199b-5p Is Correlated with Poor Prognosis for Breast Cancer Patients. Biomed. Pharmacother. 2016, 84. [Google Scholar] [CrossRef] [PubMed]
- Pan, P.; Weisenberger, D.J.; Zheng, S.; Wolf, M.; Hwang, D.G.; Rose-Nussbaumer, J.R.; Jurkunas, U.V.; Chan, M.F. Aberrant DNA Methylation of MiRNAs in Fuchs Endothelial Corneal Dystrophy. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.-J.; Duan, Y.; Wang, P.; Yin, H.-Q. MiR-199b-5p Promotes Tumor Growth and Metastasis in Cervical Cancer by down-Regulating KLK10. Biochem. Biophys. Res. Commun. 2018, 503. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; He, M.; Zhang, J.; Xu, J. High Expression of MicroRNA 221 Is a Poor Predictor for Glioma. Medicine 2020, 99. [Google Scholar] [CrossRef]
- Zhou, Q.Y.; Peng, P.L.; Xu, Y.H. MiR-221 Affects Proliferation and Apoptosis of Gastric Cancer Cells through Targeting SOCS3. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 9427–9435. [Google Scholar] [CrossRef]
- Yu, W.; Chen, H.; Qu, Y.; Xu, C.; Yang, C.; Liu, Y. MicroRNA-221 Promotes Cisplatin Resistance in Osteosarcoma Cells by Targeting PPP2R2A. Biosci. Rep. 2019, 39. [Google Scholar] [CrossRef] [Green Version]
- Sugita, S.; Yoshino, H.; Yonemori, M.; Miyamoto, K.; Matsushita, R.; Sakaguchi, T.; Itesako, T.; Tatarano, S.; Nakagawa, M.; Enokida, H. Tumor-suppressive MicroRNA-223 Targets WDR62 Directly in Bladder Cancer. Int. J. Oncol. 2019. [Google Scholar] [CrossRef]
- Bradford, M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72. [Google Scholar] [CrossRef]
- Schindelman, G.; Fernandes, J.S.; Bastiani, C.A.; Yook, K.; Sternberg, P.W. Worm Phenotype Ontology: Integrating Phenotype Data within and beyond the C. Elegans Community. BMC Bioinform. 2011, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Cao, R.; Yu, X.; Xiao, Z.; Chen, Z. MicroRNA-223-3p Inhibits Human Bladder Cancer Cell Migration and Invasion. Tumor Biol. 2017, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Feng, Y.; Huang, Y.; Liu, Y.; Wang, Y.; Liang, Y.; Zeng, H.; Qu, H.; Wei, L. MiR-223-3p Regulates Cell Viability, Migration, Invasion, and Apoptosis of Non-Small Cell Lung Cancer Cells by Targeting RHOB. Open Life Sci. 2020, 15. [Google Scholar] [CrossRef]
- Yang, X.; Liu, L.; Zou, H.; Zheng, Y.-W.; Wang, K.-P. CircZFR Promotes Cell Proliferation and Migration by Regulating MiR-511/AKT1 Axis in Hepatocellular Carcinoma. Dig. Liver Dis. 2019, 51. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Yang, Z.; Cai, X.; Zhao, M.; Sun, M.; Li, J.; Feng, G.; Feng, J.; Ye, L.; Niu, J.; et al. MiR-511 Promotes the Proliferation of Human Hepatoma Cells by Targeting the 3′UTR of B Cell Translocation Gene 1 (BTG1) MRNA. Acta Pharmacol. Sin. 2017, 38. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Fan, H.Q.; Zhang, Y.W. MiR-511-5p Functions as a Tumor Suppressor and a Predictive of Prognosis in Colorectal Cancer by Directly Targeting GPR116. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 6119–6130. [Google Scholar] [CrossRef] [PubMed]
- Heinsbroek, S.E.M.; Squadrito, M.L.; Schilderink, R.; Hilbers, F.W.; Verseijden, C.; Hofmann, M.; Helmke, A.; Boon, L.; Wildenberg, M.E.; Roelofs, J.J.T.H.; et al. MiR-511-3p, Embedded in the Macrophage Mannose Receptor Gene, Contributes to Intestinal Inflammation. Mucosal Immunol. 2016, 9. [Google Scholar] [CrossRef]
- Zhang, F.; Wu, Z. Significantly Altered Expression of MiR-511-3p and Its Target AKT3 Has Negative Prognostic Value in Human Prostate Cancer. Biochimie 2017, 140. [Google Scholar] [CrossRef]
- Wang, H.; Peng, R.; Wang, J.; Qin, Z.; Xue, L. Circulating MicroRNAs as Potential Cancer Biomarkers: The Advantage and Disadvantage. Clin. Epigenetics 2018, 10. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Eng, C.; Shen, J.; Lu, Y.; Takata, Y.; Mehdizadeh, A.; Chang, G.J.; Rodriguez-Bigas, M.A.; Li, Y.; Chang, P.; et al. Serum Exosomal MiR-4772-3p Is a Predictor of Tumor Recurrence in Stage II and III Colon Cancer. Oncotarget 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Bai, X.; Lu, D.; Lin, Y.; Lv, Y.; He, L. A Seven-miRNA Expression-based Prognostic Signature and Its Corresponding Potential Competing Endogenous RNA Network in Early Pancreatic Cancer. Exp. Ther. Med. 2019. [Google Scholar] [CrossRef]
- Lim, E.L.; Trinh, D.L.; Scott, D.W.; Chu, A.; Krzywinski, M.; Zhao, Y.; Robertson, A.G.; Mungall, A.J.; Schein, J.; Boyle, M.; et al. Comprehensive MiRNA Sequence Analysis Reveals Survival Differences in Diffuse Large B-Cell Lymphoma Patients. Genome Biol. 2015, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, J.; Zeng, L.; Jiang, X.; Zhang, Z.; Luo, X. Novel Multiple MiRNA-Based Signatures for Predicting Overall Survival and Recurrence-Free Survival of Colorectal Cancer Patients. Med. Sci. Monit. 2019, 25. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Li, D.; Fang, H.; Ning, J. Species-Specific Function of MicroRNA-7702 in Human Colorectal Cancer Cells via Targeting TADA1. Am. J. Transl. Res. 2018, 10, 2579–2589. [Google Scholar]
- Li, H.; Yu, G.; Shi, R.; Lang, B.; Chen, X.; Xia, D.; Xiao, H.; Guo, X.; Guan, W.; Ye, Z.; et al. Correction: Cisplatin-Induced Epigenetic Activation of MiR-34a Sensitizes Bladder Cancer Cells to Chemotherapy. Mol. Cancer 2014, 13, 183. [Google Scholar] [CrossRef] [Green Version]
- Wiklund, E.D.; Bramsen, J.B.; Hulf, T.; Dyrskjøt, L.; Ramanathan, R.; Hansen, T.B.; Villadsen, S.B.; Gao, S.; Ostenfeld, M.S.; Borre, M.; et al. Coordinated Epigenetic Repression of the MiR-200 Family and MiR-205 in Invasive Bladder Cancer. Int. J. Cancer 2011, 128. [Google Scholar] [CrossRef]
- Yang, R.; Xu, J.; Hua, X.; Tian, Z.; Xie, Q.; Li, J.; Jiang, G.; Cohen, M.; Sun, H.; Huang, C. Overexpressed MiR-200a Promotes Bladder Cancer Invasion through Direct Regulating Dicer/MiR-16/JNK2/MMP-2 Axis. Oncogene 2020, 39. [Google Scholar] [CrossRef] [Green Version]
- Pacurari, M.; Addison, J.B.; Bondalapati, N.; Wan, Y.-W.; Luo, D.; Qian, Y.; Castranova, V.; Ivanov, A.V.; Guo, N.L. The MicroRNA-200 Family Targets Multiple Non-Small Cell Lung Cancer Prognostic Markers in H1299 Cells and BEAS-2B Cells. Int. J. Oncol. 2013, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Guan, B.; Liu, J.; Zhang, Z.; He, S.; Zhan, Y.; Su, B.; Han, H.; Zhang, X.; Wang, B.; et al. MicroRNA-200b Is Downregulated and Suppresses Metastasis by Targeting LAMA4 in Renal Cell Carcinoma. EBioMedicine 2019, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braicu, C.; Raduly, L.; Morar-Bolba, G.; Cojocneanu, R.; Jurj, A.; Pop, L.-A.; Pileczki, V.; Ciocan, C.; Moldovan, A.; Irimie, A.; et al. Aberrant MiRNAs Expressed in HER-2 Negative Breast Cancers Patient. J. Exp. Clin. Cancer Res. 2018, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maleki, S.; Cottrill, K.A.; Poujade, F.-A.; Bhattachariya, A.; Bergman, O.; Gådin, J.R.; Simon, N.; Lundströmer, K.; Franco-Cereceda, A.; Björck, H.M.; et al. The Mir-200 Family Regulates Key Pathogenic Events in Ascending Aortas of Individuals with Bicuspid Aortic Valves. J. Intern. Med. 2019, 285. [Google Scholar] [CrossRef] [Green Version]
- Moh-Moh-Aung, A.; Fujisawa, M.; Ito, S.; Katayama, H.; Ohara, T.; Ota, Y.; Yoshimura, T.; Matsukawa, A. Decreased MiR-200b-3p in Cancer Cells Leads to Angiogenesis in HCC by Enhancing Endothelial ERG Expression. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef]
- Gao, Y.; Zhang, W.; Liu, C.; Li, G. MiR-200 Affects Tamoxifen Resistance in Breast Cancer Cells through Regulation of MYB. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef]
- Saleeb, R.; Kim, S.S.; Ding, Q.; Scorilas, A.; Lin, S.; Khella, H.W.; Boulos, C.; Ibrahim, G.; Yousef, G.M. The MiR-200 Family as Prognostic Markers in Clear Cell Renal Cell Carcinoma. Urol. Oncol. Semin. Orig. Investig. 2019, 37. [Google Scholar] [CrossRef]
- Shindo, T.; Niinuma, T.; Nishiyama, N.; Shinkai, N.; Kitajima, H.; Kai, M.; Maruyama, R.; Tokino, T.; Masumori, N.; Suzuki, H. Epigenetic Silencing of MiR-200b Is Associated with Cisplatin Resistance in Bladder Cancer. Oncotarget 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.; Song, C.; Ye, F.; Gao, G.; Ou, X.; Zhang, L.; Xie, X.; Xie, X. MiR-200c Suppresses Stemness and Increases Cellular Sensitivity to Trastuzumab in HER2+ Breast Cancer. J. Cell. Mol. Med. 2019, 23. [Google Scholar] [CrossRef]
- Rogers, T.J.; Christenson, J.L.; Greene, L.I.; O’Neill, K.I.; Williams, M.M.; Gordon, M.A.; Nemkov, T.; D’Alessandro, A.; Degala, G.D.; Shin, J.; et al. Reversal of Triple-Negative Breast Cancer EMT by MiR-200c Decreases Tryptophan Catabolism and a Program of Immunosuppression. Mol. Cancer Res. 2019, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karimi Mazraehshah, M.; Tavangar, S.M.; Saidijam, M.; Amini, R.; Bahreini, F.; Karimi Dermani, F.; Najafi, R. Anticancer Effects of MiR-200c in Colorectal Cancer through BMI1. J. Cell. Biochem. 2018, 119. [Google Scholar] [CrossRef] [PubMed]
- Xie, N.N.; Liu, Z.X.; Wu, C.; Wang, P.L.; Song, G.T.; Chen, Z. MicroRNA-200c Suppresses Tumor Metastasis in Oral Squamous Carcinoma by Inhibiting Epithelial-Mesenchymal Transition. Eur. Rev. Med Pharmacol. Sci. 2018, 22, 3415–3422. [Google Scholar] [CrossRef]
- Oto, J.; Plana, E.; Fernández-Pardo, Á.; Cana, F.; Martínez-Sarmiento, M.; Vera-Donoso, C.D.; España, F.; Medina, P. Identification of MiR-29c-3p as a Robust Normalizer for Urine MicroRNA Studies in Bladder Cancer. Biomedicines 2020, 8, 447. [Google Scholar] [CrossRef]
- Vilella, F.; Moreno-Moya, J.M.; Balaguer, N.; Grasso, A.; Herrero, M.; Martinez, S.; Marcilla, A.; Simon, C. Hsa-MiR-30d, Secreted by the Human Endometrium, Is Taken up by the Pre-Implantation Embryo and Might Modify Its Transcriptome. Development 2015, 142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, S.-F.; Chang, Y.-W.; Andreu-Vieyra, C.; Fang, J.Y.; Yang, Z.; Han, B.; Lee, A.S.; Liang, G. MiR-30d, MiR-181a and MiR-199a-5p Cooperatively Suppress the Endoplasmic Reticulum Chaperone and Signaling Regulator GRP78 in Cancer. Oncogene 2013, 32. [Google Scholar] [CrossRef] [Green Version]
- Zhu, B.; Ekman, M.; Svensson, D.; Lindvall, J.M.; Nilsson, B.-O.; Uvelius, B.; Swärd, K. Array Profiling Reveals Contribution of Cthrc1 to Growth of the Denervated Rat Urinary Bladder. Am. J. Physiol. Ren. Physiol. 2018, 314. [Google Scholar] [CrossRef]
- Liang, L.; Yang, Z.; Deng, Q.; Jiang, Y.; Cheng, Y.; Sun, Y.; Li, L. MiR-30d-5p Suppresses Proliferation and Autophagy by Targeting ATG5 in Renal Cell Carcinoma. FEBS Open Bio 2020. [Google Scholar] [CrossRef]
- Hosseini, S.M.; Soltani, B.M.; Tavallaei, M.; Mowla, S.J.; Tafsiri, E.; Bagheri, A.; Khorshid, H. Clinically Significant Dysregulation of Hsa-MiR-30d-5p and Hsa-Let-7b Expression in Patients with Surgically Resected Non-Small Cell Lung Cancer. Avicenna J. Med. Biotechnol. 2018, 10, 98–104. [Google Scholar]
- Yin, X.-H.; Jin, Y.-H.; Cao, Y.; Wong, Y.; Weng, H.; Sun, C.; Deng, J.-H.; Zeng, X.-T. Development of a 21-MiRNA Signature Associated with the Prognosis of Patients with Bladder Cancer. Front. Oncol. 2019, 9. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Bu, Q.; Zeng, M.; Xia, D.; Wu, A. MicroRNA-93 Promotes Bladder Cancer Proliferation and Invasion by Targeting PEDF. Urol. Oncol. Semin. Orig. Investig. 2019, 37. [Google Scholar] [CrossRef]
- Juracek, J.; Peltanova, B.; Dolezel, J.; Fedorko, M.; Pacik, D.; Radova, L.; Vesela, P.; Svoboda, M.; Slaby, O.; Stanik, M. Genome-Wide Identification of Urinary Cell-Free MicroRNAs for Non-Invasive Detection of Bladder Cancer. J. Cell. Mol. Med. 2018, 22. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Liu, J.; Wang, H.; Wang, Y.; Li, Z.; Pan, Y.; Liu, Q.; Yang, M. Repression of the MiR-93-Enhanced Sensitivity of Bladder Carcinoma to Chemotherapy Involves the Regulation of LASS2. Oncotargets Ther. 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armstrong, D.A.; Green, B.B.; Seigne, J.D.; Schned, A.R.; Marsit, C.J. MicroRNA Molecular Profiling from Matched Tumor and Bio-Fluids in Bladder Cancer. Mol. Cancer 2015, 14. [Google Scholar] [CrossRef] [Green Version]
- Shi, X.; Liu, T.-T.; Yu, X.-N.; Balakrishnan, A.; Zhu, H.-R.; Guo, H.-Y.; Zhang, G.-C.; Bilegsaikhan, E.; Sun, J.-L.; Song, G.-Q.; et al. MicroRNA-93-5p Promotes Hepatocellular Carcinoma Progression via a MicroRNA-93-5p/MAP3K2/c-Jun Positive Feedback Circuit. Oncogene 2020, 39. [Google Scholar] [CrossRef]
- Erdmann, K.; Salomo, K.; Klimova, A.; Heberling, U.; Lohse-Fischer, A.; Fuehrer, R.; Thomas, C.; Roeder, I.; Froehner, M.; Wirth, M.P.; et al. Urinary MicroRNAs as Potential Markers for Non-Invasive Diagnosis of Bladder Cancer. Int. J. Mol. Sci. 2020, 21, 3814. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Zhao, X.; Zhou, J.; Cheng, X.; Ye, Z.; Ji, Z. Long Non-Coding RNA MEG3 Suppresses the Development of Bladder Urothelial Carcinoma by Regulating MiR-96 and TPM1. Cancer Biol. Ther. 2018, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, C.; Zhang, Q.; Gu, R.; Lou, Y.; Liu, W. MiR-96 Regulates Migration and Invasion of Bladder Cancer through Epithelial-Mesenchymal Transition in Response to Transforming Growth Factor-B1. J. Cell. Biochem. 2018, 119. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Du, X.; Hu, J.; Zhu, Y.; Wu, H.; Dai, G.; Shu, Y.; Ouyang, J. Anticancer Effect of MiR-96 Inhibitor in Bladder Cancer Cell Lines. Oncol. Lett. 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.; Liu, K.; Wang, Y.; Xu, Z.; Meng, J.; Gu, S. Upregulation of MicroRNA-96 and Its Oncogenic Functions by Targeting CDKN1A in Bladder Cancer. Cancer Cell Int. 2015, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papadopoulos, E.I.; Scorilas, A. Cisplatin and Paclitaxel Alter the Expression Pattern of MiR-143/145 and MiR-183/96/182 Clusters in T24 Bladder Cancer Cells. Clin. Transl. Sci. 2015, 8. [Google Scholar] [CrossRef] [Green Version]
- Luo, H.; Yang, L.; Liu, C.; Wang, X.; Dong, Q.; Liu, L.; Wei, Q. TMPO-AS1/MiR-98-5p/EBF1 Feedback Loop Contributes to the Progression of Bladder Cancer. Int. J. Biochem. Cell Biol. 2020, 122. [Google Scholar] [CrossRef] [PubMed]
- Fırat, E.; Aybek, Z.; Akgün, Ş.; Küçüker, K.; Akça, H.; Aybek, H. Exploring Biomarkers in the Overactive Bladder: Alterations in MiRNA Levels of a Panel of Genes in Patients with OAB. Neurourol. Urodyn. 2019, 38. [Google Scholar] [CrossRef]
- Luan, T.; Fu, S.; Huang, L.; Zuo, Y.; Ding, M.; Li, N.; Chen, J.; Wang, H.; Wang, J. MicroRNA-98 Promotes Drug Resistance and Regulates Mitochondrial Dynamics by Targeting LASS2 in Bladder Cancer Cells. Exp. Cell Res. 2018, 373. [Google Scholar] [CrossRef]
- Feng, F.; Chen, A.; Huang, J.; Xia, Q.; Chen, Y.; Jin, X. Long Noncoding RNA SNHG16 Contributes to the Development of Bladder Cancer via Regulating MiR-98/STAT3/Wnt/B-catenin Pathway Axis. J. Cell. Biochem. 2018, 119. [Google Scholar] [CrossRef]
- Sun, D.; Luo, X.; Ma, L.; Wang, Y.; Zhang, F. Identifying of MiR-98-5p/IGF1 Axis Contributes Breast Cancer Progression Using Comprehensive Bioinformatic Analyses Methods and Experiments Validation. Life Sci. 2020, 261. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Wang, P. MiR-188-5p and MiR-141-3p Influence Prognosis of Bladder Cancer and Promote Bladder Cancer Synergistically. Pathol. Res. Pract. 2019, 215. [Google Scholar] [CrossRef] [PubMed]
- Nekoohesh, L.; Modarressi, M.H.; Mowla, S.J.; Sadroddiny, E.; Etemadian, M.; Afsharpad, M.; Zolfaghari, F.; Barzegari, M.; Saffari, M.; Oskooei, V.K.; et al. Expression Profile of MiRNAs in Urine Samples of Bladder Cancer Patients. Biomark. Med. 2018, 12. [Google Scholar] [CrossRef] [PubMed]
- Phatak, P.; Noe, M.; Asrani, K.; Chesnick, I.E.; Greenwald, B.D.; Donahue, J.M. MicroRNA-141-3p Regulates Cellular Proliferation, Migration, and Invasion in Esophageal Cancer by Targeting Tuberous Sclerosis Complex 1. Mol. Carcinog. 2020. [Google Scholar] [CrossRef]
- Guan, G.; Niu, X.; Qiao, X.; Wang, X.; Liu, J.; Zhong, M. Upregulation of Neural Cell Adhesion Molecule 1 (NCAM1) by Hsa-MiR-141-3p Suppresses Ameloblastoma Cell Migration. Med. Sci. Monit. 2020, 26. [Google Scholar] [CrossRef]
- Hernández-Rodríguez, R.T.; Amezcua-Guerra, L.M. The Potential Role of MicroRNAs as Biomarkers in Atopic Dermatitis: A Systematic Review. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 11804–11809. [Google Scholar] [CrossRef]
- Zhang, Y.; Gao, T.; Li, X.; Wen, C.-C.; Yan, X.-T.; Peng, C.; Xiao, Y. Circ_0005075 Targeting MiR-151a-3p Promotes Neuropathic Pain in CCI Rats via Inducing NOTCH2 Expression. Gene 2021, 767. [Google Scholar] [CrossRef]
- Liu, H.; Cheng, Y.; Xu, Y.; Xu, H.; Lin, Z.; Fan, J.; Lang, J. The Inhibition of Tumor Protein P53 by MicroRNA-151a-3p Induced Cell Proliferation, Migration and Invasion in Nasopharyngeal Carcinoma. Biosci. Rep. 2019, 39. [Google Scholar] [CrossRef] [PubMed]
- Yue, C.; Chen, X.; Li, J.; Yang, X.; Li, Y.; Wen, Y. MiR-151-3p Inhibits Proliferation and Invasion of Colon Cancer Cell by Targeting Close Homolog of L1. J. Biomed. Nanotechnol. 2020, 16. [Google Scholar] [CrossRef]
- Setti Boubaker, N.; Gurtner, A.; Trabelsi, N.; Manni, I.; Ayed, H.; Saadi, A.; Naimi, Z.; Ksontini, M.; Ayadi, M.; Blel, A.; et al. Uncovering the Expression Patterns and the Clinical Significance of MiR-182, MiR-205, MiR-27a and MiR-369 in Patients with Urinary Bladder Cancer. Mol. Biol. Rep. 2020, 47. [Google Scholar] [CrossRef]
- Gu, C.; Zhao, K.; Zhou, N.; Liu, F.; Xie, F.; Yu, S.; Feng, Y.; Chen, L.; Yang, J.; Tian, F.; et al. UBAC2 Promotes Bladder Cancer Proliferation through BCRC-3/MiRNA-182-5p/P27 Axis. Cell Death Dis. 2020, 11. [Google Scholar] [CrossRef]
- Wang, F.; Wu, D.; Xu, Z.; Chen, J.; Zhang, J.; Li, X.; Chen, S.; He, F.; Xu, J.; Su, L.; et al. MiR-182-5p Affects Human Bladder Cancer Cell Proliferation, Migration and Invasion through Regulating Cofilin 1. Cancer Cell Int. 2019, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.; Zhang, Y. MiR-182 Inhibits Proliferation, Migration, Invasion and Inflammation of Endometrial Stromal Cells through Deactivation of NF-KB Signaling Pathway in Endometriosis. Mol. Cell. Biochem. 2021. [Google Scholar] [CrossRef]
- Lu, B.; Yu, Y.; Xing, X.-L.; Liu, R.-Y. MiR-183/TMSB4Y, a New Potential Signaling Axis, Involving in the Progression of Laryngeal Cancer via Modulating Cell Adhesion. J. Recept. Signal Transduct. 2020. [Google Scholar] [CrossRef]
- Chen, D.; Li, S.G.; Chen, J.Y.; Xiao, M. MiR-183 Maintains Canonical Wnt Signaling Activity and Regulates Growth and Apoptosis in Bladder Cancer via Targeting AXIN2. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 4828–4836. [Google Scholar] [CrossRef]
- Yoshino, H.; Seki, N.; Itesako, T.; Chiyomaru, T.; Nakagawa, M.; Enokida, H. Aberrant Expression of MicroRNAs in Bladder Cancer. Nat. Rev. Urol. 2013, 10. [Google Scholar] [CrossRef]
- Lan, X.; Wu, N.; Wu, L.; Qu, K.; Osoro, E.K.; Guan, D.; Du, X.; Wang, B.; Chen, S.; Miao, J.; et al. The Human Novel Gene LNC-HC Inhibits Hepatocellular Carcinoma Cell Proliferation by Sequestering Hsa-MiR-183-5p. Mol. Ther. Nucleic Acids 2020, 20. [Google Scholar] [CrossRef]
- Li, Z.; Lin, C.; Zhao, L.; Zhou, L.; Pan, X.; Quan, J.; Peng, X.; Li, W.; Li, H.; Xu, J.; et al. Oncogene MiR-187-5p Is Associated with Cellular Proliferation, Migration, Invasion, Apoptosis and an Increased Risk of Recurrence in Bladder Cancer. Biomed. Pharmacother. 2018, 105. [Google Scholar] [CrossRef]
- Huang, G.; Liang, M.; Liu, H.; Huang, J.; Li, P.; Wang, C.; Zhang, Y.; Lin, Y.; Jiang, X. CircRNA Hsa_circRNA_104348 Promotes Hepatocellular Carcinoma Progression through Modulating MiR-187-3p/RTKN2 Axis and Activating Wnt/β-Catenin Pathway. Cell Death Dis. 2020, 11. [Google Scholar] [CrossRef]
- Zheng, Y.; Yang, Z.; Jin, C.; Chen, C.; Wu, N. Hsa-MiR-191-5p Inhibits Replication of Human Immunodeficiency Virus Type 1 by Downregulating the Expression of NUP50. Arch. Virol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Ashirbekov, Y.; Abaildayev, A.; Omarbayeva, N.; Botbayev, D.; Belkozhayev, A.; Askandirova, A.; Neupokoyeva, A.; Utegenova, G.; Sharipov, K.; Aitkhozhina, N. Combination of Circulating MiR-145-5p/MiR-191-5p as Biomarker for Breast Cancer Detection. PeerJ 2020, 8. [Google Scholar] [CrossRef]
- Emami, N.; Mohamadnia, A.; Mirzaei, M.; Bayat, M.; Mohammadi, F.; Bahrami, N. MiR-155, MiR-191, and MiR-494 as Diagnostic Biomarkers for Oral Squamous Cell Carcinoma and the Effects of Avastin on These Biomarkers. J. Korean Assoc. Oral Maxillofac. Surg. 2020, 46. [Google Scholar] [CrossRef]
- Yang, C.; Ota-Kurogi, N.; Ikeda, K.; Okumura, T.; Horie-Inoue, K.; Takeda, S.; Inoue, S. MicroRNA-191 Regulates Endometrial Cancer Cell Growth via TET1-Mediated Epigenetic Modulation of APC. J. Biochem. 2020, 168. [Google Scholar] [CrossRef] [PubMed]
- Pouliot, L.M.; Chen, Y.-C.; Bai, J.; Guha, R.; Martin, S.E.; Gottesman, M.M.; Hall, M.D. Cisplatin Sensitivity Mediated by WEE1 and CHK1 Is Mediated by MiR-155 and the MiR-15 Family. Cancer Res. 2012, 72, 5945–5955. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Wang, G.; Zhu, Y.; Li, X.; Yan, F.; Zhang, C.; Huang, X.; Zhang, Y. Aberrant Regulation of MiR-15b in Human Malignant Tumors and Its Effects on the Hallmarks of Cancer. Tumor Biol. 2015, 37, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Finnerty, J.R.; Wang, W.-X.; Hébert, S.S.; Wilfred, B.R.; Mao, G.; Nelson, P.T. The MiR-15/107 Group of MicroRNA Genes: Evolutionary Biology, Cellular Functions, and Roles in Human Diseases. J. Mol. Biol. 2010, 402, 491–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, K.; Shi, J.; Mo, D.; Yang, Y.; Fu, Q.; Luo, Y. MiR-219a-1 Inhibits Colon Cancer Cells Proliferation and Invasion by Targeting MEMO1. Cancer Biol. Ther. 2020, 21. [Google Scholar] [CrossRef]
- Wei, T.; Cheng, S.; Fu, X.N.; Feng, L.J. MiR-219a-5p Enhances the Radiosensitivity of Non-Small Cell Lung Cancer Cells through Targeting CD164. Biosci. Rep. 2020, 40. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Jin, X.; Liao, C.; Qiao, L.; Zhao, W. MicroRNA-301b-3p Accelerates the Growth of Gastric Cancer Cells by Targeting Zinc Finger and BTB Domain Containing 4. Pathol. Res. Pract. 2019, 215. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Wang, Y.; Liang, J.; Liu, Z.; Sun, X.; Cai, K. MiR-301b Promotes the Proliferation, Mobility, and Epithelial-to-Mesenchymal Transition of Bladder Cancer Cells by Targeting EGR1. Biochem. Cell Biol. 2017, 95. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Chang, M.; Song, X.; Ding, S.; Xie, L.; Song, X. Plasma miR -1247-5p, miR -301b-3p and miR -105-5p as Potential Biomarkers for Early Diagnosis of Non-small Cell Lung Cancer. Thorac. Cancer 2020. [Google Scholar] [CrossRef]
- Dos Santos Arcas, C.; Lin-Wang, H.T.; Umeda, I.I.K.; de Sousa, M.G.; Utiyama, D.M.O.; de Padua Mansur, A.; Macchione, M.; Hirata, M.H.; Nakagawa, N.K. Smoking Load Reduction Is Insufficient to Downregulate MiR-301b, a Lung Cancer Promoter. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef]
- Du, L.; Jiang, X.; Duan, W.; Wang, R.; Wang, L.; Zheng, G.; Yan, K.; Wang, L.; Li, J.; Zhang, X.; et al. Cell-Free MicroRNA Expression Signatures in Urine Serve as Novel Noninvasive Biomarkers for Diagnosis and Recurrence Prediction of Bladder Cancer. Oncotarget 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Yang, X.; Li, P.; Wei, J.; Deng, X.; Cheng, Y.; Qin, C.; Ju, X.; Meng, X.; Li, J.; et al. Identification of Circulating MicroRNA Signatures for Upper Tract Urothelial Carcinoma Detection. Mol. Med. Rep. 2015, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Li, B.; Shi, X.; Zhang, J.; Chen, A.M.; Xu, J.; Wang, W.; Huang, K.; Gao, J.; Zheng, Z.; et al. Cross-Platform Genomic Identification and Clinical Validation of Breast Cancer Diagnostic Biomarkers. Aging 2021. [Google Scholar] [CrossRef]
- Guo, C.M.; Liu, S.Q.; Sun, M.Z. MiR-429 as Biomarker for Diagnosis, Treatment and Prognosis of Cancers and Its Potential Action Mechanisms: A Systematic Literature Review. Neoplasma 2020, 67. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-L.; Ho, J.-Y.; Hung, S.-H.; Yu, D.-S. MiR-429 Expression in Bladder Cancer and Its Correlation with Tumor Behavior and Clinical Outcome. Kaohsiung J. Med. Sci. 2018, 34. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Liu, Y.; He, A.; Liu, Y.; Wu, J.; Liao, X.; Lv, Z.; Wang, F.; Mei, H. Hsa-MiR-429 Promotes Bladder Cancer Cell Proliferation via Inhibiting CDKN2B. Oncotarget 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.-L.; Ho, J.-Y.; Chou, S.-C.; Yu, D.-S. MiR-429 Reverses Epithelial-Mesenchymal Transition by Restoring E-Cadherin Expression in Bladder Cancer. Oncotarget 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhao, W.; Wang, X. Inhibition of Long Non-Coding RNA MALAT1 Elevates MicroRNA-429 to Suppress the Progression of Hypopharyngeal Squamous Cell Carcinoma by Reducing ZEB1. Life Sci. 2020, 262. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Liao, K.; Liu, D. MicroRNA-744-5p Is Downregulated in Colorectal Cancer and Targets SEPT2 to Suppress the Malignant Phenotype. Mol. Med. Rep. 2020, 23. [Google Scholar] [CrossRef]
- Friedrich, M.; Vaxevanis, C.K.; Biehl, K.; Mueller, A.; Seliger, B. Targeting the Coding Sequence: Opposing Roles in Regulating Classical and Non-Classical MHC Class I Molecules by MiR-16 and MiR-744. J. Immunother. Cancer 2020, 8. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, J.; Li, J.; Li, Q. miR-744-5p Inhibits Cellular Proliferation and Invasion via Targeting ARF1 in Epithelial Ovarian Cancer. Kaohsiung J. Med. Sci. 2020, 36. [Google Scholar] [CrossRef] [PubMed]
- Pilson, Q.; Smith, S.; Jefferies, C.A.; Ní Gabhann-Dromgoole, J.; Murphy, C.C. MiR-744-5p Contributes to Ocular Inflammation in Patients with Primary Sjogrens Syndrome. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef]
- Wang, R.; Wu, Y.; Huang, W.; Chen, W. MicroRNA-940 Targets INPP4A or GSK3β and Activates the Wnt/β-Catenin Pathway to Regulate the Malignant Behavior of Bladder Cancer Cells. Oncol. Res. Featur. Preclin. Clin. Cancer Ther. 2018, 26. [Google Scholar] [CrossRef]
- Ahadi, A. A Systematic Review of MicroRNAs as Potential Biomarkers for Diagnosis and Prognosis of Gastric Cancer. Immunogenetics 2021. [Google Scholar] [CrossRef]
- Wu, Y.-P.; Lin, X.-D.; Chen, S.-H.; Ke, Z.-B.; Lin, F.; Chen, D.-N.; Xue, X.-Y.; Wei, Y.; Zheng, Q.-S.; Wen, Y.-A.; et al. Identification of Prostate Cancer-Related Circular RNA Through Bioinformatics Analysis. Front. Genet. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.; Han, W.; Fu, Y.; Chen, Q. The Hsa_circ_0091579/MiR-940/TACR1 Axis Regulates the Development of Hepatocellular Carcinoma. Cancer Manag. Res. 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Li, F.; Zhu, Z.; Ding, A.; Luo, J. CircRNA CDR1as/MiR-1287/Raf1 Axis Modulates Hepatocellular Carcinoma Progression Through MEK/ERK Pathway. Cancer Manag. Res. 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.; Qu, J.; Wang, J.; Liang, X.; Sun, W. Circular RNA Circ_0026134 Regulates Non-Small Cell Lung Cancer Cell Proliferation and Invasion via Sponging MiR-1256 and MiR-1287. Biomed. Pharmacother. 2019, 112. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Tan, Q.; Chen, H.; Chen, K.; Wang, H.; Chen, Z.; Xi, Y.; Yin, H.; Lai, K.; Liu, Y. LncRNA-SNHG7-003 Inhibits the Proliferation, Migration and Invasion of Vascular Smooth Muscle Cells by Targeting the MiR-1306-5p/SIRT7 Signaling Pathway. Int. J. Mol. Med. 2020, 47. [Google Scholar] [CrossRef]
- Zhu, Z.; Rong, Z.; Luo, Z.; Yu, Z.; Zhang, J.; Qiu, Z.; Huang, C. Circular RNA CircNHSL1 Promotes Gastric Cancer Progression through the MiR-1306-3p/SIX1/Vimentin Axis. Mol. Cancer 2019, 18. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Rong, Z.; Luo, Z.; Yu, Z.; Zhang, J.; Qiu, Z.; Huang, C. Correction to: Circular RNA CircNHSL1 Promotes Gastric Cancer Progression through the MiR-1306-3p/SIX1/Vimentin Axis. Mol. Cancer 2020, 19. [Google Scholar] [CrossRef]
- Ma, Y.; Zhou, A.; Song, J. Upregulation of MiR-1307-3p and Its Function in the Clinical Prognosis and Progression of Gastric Cancer. Oncol. Lett. 2020, 21. [Google Scholar] [CrossRef]
- Eun, J.W.; Seo, C.W.; Baek, G.O.; Yoon, M.G.; Ahn, H.R.; Son, J.A.; Sung, S.; Kim, D.W.; Kim, S.S.; Cho, H.J.; et al. Circulating Exosomal MicroRNA-1307-5p as a Predictor for Metastasis in Patients with Hepatocellular Carcinoma. Cancers 2020, 12, 3819. [Google Scholar] [CrossRef]
- Du, X.; Wang, S.; Liu, X.; He, T.; Lin, X.; Wu, S.; Wang, D.; Li, J.; Huang, W.; Yang, H. MiR-1307-5p Targeting TRAF3 Upregulates the MAPK/NF-KB Pathway and Promotes Lung Adenocarcinoma Proliferation. Cancer Cell Int. 2020, 20. [Google Scholar] [CrossRef] [PubMed]
- Stark, M.S.; Tyagi, S.; Nancarrow, D.J.; Boyle, G.M.; Cook, A.L.; Whiteman, D.C.; Parsons, P.G.; Schmidt, C.; Sturm, R.A.; Hayward, N.K. Characterization of the Melanoma MiRNAome by Deep Sequencing. PLoS ONE 2010, 5. [Google Scholar] [CrossRef]
- Wang, S.; Cheng, Y.; Yang, P.; Qin, G. Silencing of Long Noncoding RNA LINC00324 Interacts with MicroRNA-3200-5p to Attenuate the Tumorigenesis of Gastric Cancer via Regulating BCAT1. Gastroenterol. Res. Pract. 2020. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Qin, X.; Shao, Y.; Zhou, Y.; Ye, G.; Xu, S. Circular RNA MTO1 Suppresses Tumorigenesis of Gastric Carcinoma by Sponging MiR-3200-5p and Targeting PEBP1. Mol. Cell. Probes 2020, 52. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chu, X.Q.; Zhang, D.; Kong, D.F. Knockdown of Long Non-Coding RNA PEG10 Inhibits Growth, Migration and Invasion of Gastric Carcinoma Cells by up-Regulating MiR-3200. Neoplasma 2018, 65. [Google Scholar] [CrossRef] [Green Version]
- Gu, Y.; Wang, W.; Wang, X.; Xie, H.; Ye, X.; Shu, P. Integrated Network Analysis Identifies Hsa-MiR-4756-3p as a Regulator of FOXM1 in Triple Negative Breast Cancer. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef] [Green Version]
- Jia, Y.; Zheng, Z.; Yang, Y.; Zou, M.; Li, J.; Wang, L.; Guan, M.; Xue, Y. MiR-4756 Promotes Albumin-induced Renal Tubular Epithelial Cell Epithelial-to-mesenchymal Transition and Endoplasmic Reticulum Stress via Targeting Sestrin2. J. Cell. Physiol. 2019, 234. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, P.; Hu, J.; Zhao, L.N.; Li, J.P.; Ma, R.; Li, W.W.; Shi, M.; Wei, L.C. Role and Mechanism of MiR-4778-3p and Its Targets NR2C2 and Med19 in Cervical Cancer Radioresistance. Biochem. Biophys. Res. Commun. 2019, 508. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Dai, M.; Chen, H.; Li, Y.; Zhang, J.; Zou, Z.; Yang, H. Prognostic Value of Hsa-mir-299 and Hsa-mir-7706 in Hepatocellular Carcinoma. Oncol. Lett. 2018. [Google Scholar] [CrossRef]
- Zhang, Y.; Velez-Delgado, A.; Mathew, E.; Li, D.; Mendez, F.M.; Flannagan, K.; Rhim, A.D.; Simeone, D.M.; Beatty, G.L.; di Magliano, M.P. Myeloid Cells Are Required for PD-1/PD-L1 Checkpoint Activation and the Establishment of an Immunosuppressive Environment in Pancreatic Cancer. Gut 2016, 66, 124–136. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Tao, Z.; Hai, B.; Liang, H.; Shi, Y.; Wang, T.; Song, W.; Chen, Y.; OuYang, J.; Chen, J.; et al. MiR-424(322) Reverses Chemoresistance via T-Cell Immune Response Activation by Blocking the PD-L1 Immune Checkpoint. Nat. Commun. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Weinstein, J.N.; Akbani, R.; Broom, B.M.; Wang, W.; Verhaak, R.G.W.; McConkey, D.; Lerner, S.; Morgan, M.; Creighton, C.J.; Smith, C.; et al. Comprehensive Molecular Characterization of Urothelial Bladder Carcinoma. Nature 2014, 507, 315–322. [Google Scholar] [CrossRef] [Green Version]
- Colaprico, A.; Silva, T.C.; Olsen, C.; Garofano, L.; Cava, C.; Garolini, D.; Sabedot, T.S.; Malta, T.M.; Pagnotta, S.M.; Castiglioni, I.; et al. TCGAbiolinks: An R/Bioconductor Package for Integrative Analysis of TCGA Data. Nucleic Acids Res. 2015, 44, e71. [Google Scholar] [CrossRef]
- TCGA Correlations—Heatmaps. Available online: https://corr.netlify.app (accessed on 8 January 2021).










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA | Function | References |
|---|---|---|
| hsa-miR-21 | Impact on survival and prognosis in patients with pancreatic cancer; exosomal miR-21 promotes proliferation, but also invasion and therapy resistance of colon adenocarcinoma cells via its target PDCD4; nothing is known about its role in bladder cancer; a lack of miR-21 expression in tumor-associated macrophages (TAMs) promotes antitumoral immune response. | [28,29,30] |
| hsa-miR-29a | Downregulated in bladder cancer; shows inhibition of proliferation in bladder cancer cell lines via PI3K-AKT pathway; acts as a tumour suppressor in many cancer types; increased urine levels correlated with shorter event-free survival in most cancer types; high expression of miR-29a was associated with a prolonged disease-free survival. | [31,32,33,34,35] |
| hsa-miR-100 | Suggested role in the invasion and metastasis of hepatocellular carcinoma; involved in the PI3K/AKT and mTOR pathways in renal carcinoma; interestingly, a variant in the miR-100 gene is a protective factor of childhood acute lymphoblastic leukaemia. | [36,37,38] |
| hsa-miR-125b | Suppresses cell proliferation and metastasis by targeting the HAX-1 gene in esophageal squamous cell carcinoma; regulates IL-1β-induced inflammatory genes through targeting TRAF6-mediated MAPKs and NF-κB signaling in human osteoarthritic chondrocytes; acts as an oncogene in glioblastoma cells and inhibits cell apoptosis through p53 and p38MAPK-independent pathways; has a role in conferring the metastatic phenotype among pancreatic cancer cells; unknown function in bladder cancer. | [NO_PRINTED_FORM][39,40,41,42] |
| hsa-miR-142 | Linked to the reduced regulatory T-cell function in granulomatosis; suppresses cell proliferation and cell migration in bladder cancer. | [25,26] |
| hsa-miR-146a | Mediates the suppression of inflammatory response in adipocytes; involved in bladder cancer relapse; important for the maintenance of breast cancer stem cells during EMT; suggested that the urine levels might be possibly used as a prognostic marker for bladder cancer; in bladder cancer, it is usually upregulated, and this event is correlated with the inhibitory effect on the invasion of cancer cells resulting from the reduction of MMP2 expression. | [20,21,22,23,24,25] |
| hsa-miR-150 | Acts as a tumour promoter: promotes cell proliferation, migration and invasion of cancer cells through targeting PDCD4 (programmed cell death 4 protein); modulates cisplatin chemosensitivity and invasiveness of muscle-invasive bladder cancer cells via targeting PDCD4; is suggested as a urinary biomarker for bladder cancer progression; its agonist promotes fibrosis in cultured kidney cells, while its antagonists decrease pro-inflammatory cytokines and pro-fibrotic proteins and increase anti-fibrotic protein SOCS1. | [43,44,45,46,47] |
| hsa-miR-155 | Tumour-promoting and highly oncogenic microRNA that targets ELK3 transcription factor functioning in the hypoxia response; upregulated in breast cancer and linked to PARP-1 inhibitors response; overexpressed in bladder cancer, promoting tumour growth by repressing DMTF1. | [17,18,19] |
| hsa-miR-199a | Functions as a tumour suppressor in oral squamous cell carcinoma, targeting the IKKβ/NF-κB signalling pathway; inhibits malignant progression of lung cancer through mediating RGS17; serum levels were suggested as a potential diagnostic biomarker for detection of colorectal cancer; recently discovered to inhibit angiogenesis by targeting the VEGF/PI3K/AKT signalling pathway in an in vitro model of diabetic retinopathy; can attenuate aerobic glycolysis and cell proliferation in glioblastoma, but the role in bladder cancer remains unrevealed. | [48,49,50,51,52] [NO_PRINTED_FORM] |
| hsa-miR-199b | Downregulated in breast cancer patients; is often associated with malignant clinical characteristics; exerts tumour suppressive functions in hepatocellular carcinoma by targeting JAG1 directly; suppression of miR-199b expressions improves apoptosis and reduces the cell viability in cervical cancer. | [53,54,55,56] |
| hsa-miR-221 | High expression is a poor predictor for glioma; affects proliferation and apoptosis of gastric cancer cells (through targeting SOCS3); promotes cisplatin resistance in osteosarcoma cells by targeting PPP2R2A; the function in bladder cancer is unknown. | [57,58,59] |
| hsa-miR-223 | Tumour-suppressive, but also oncogenic miR in various cancers; targets WDR62 directly in bladder cancer—the knockdown of WDR62 in mice significantly inhibited tumour aggressiveness and induced the apoptosis of bladder cancer cells; it may also inhibit migration and invasion of bladder cancer cells. | [60,61,62,63,64] |
| hsa-miR-511 | circZFR promotes cell proliferation and migration by regulating the miR-511/AKT1 pathway in hepatocellular carcinoma; promotes proliferation of human hepatoma cells; functions as a tumour suppressor and a prognostic marker in colorectal cancer; contributes to intestinal inflammation; significantly altered expression and its target AKT3 have negative prognostic value in prostate cancer, but its function in bladder cancer remains unknown. | [65,66,67,68,69] |
| hsa-miR-4772 | Significance in bladder cancer and immunological surveillance remains unclear, but a high level in serum exosomes derived from stage II and stage III colon cancer patients was negatively associated with the risk of recurrence and the risk of death. | [70,71] |
| hsa-miR-5586 | Downregulated in pancreatic and bladder cancers; high levels linked to good outcomes in diffuse large B-cell lymphoma (DLBCL); significance in bladder cancer and immunological surveillance remains unknown. | [72,73] |
| hsa-miR-7702 | Potentially important in colorectal cancer (CRC) progression, but significance in bladder cancer remains unclear. | [74,75] |
| miRNA | Function | References |
|---|---|---|
| hsa-miR-30d | Involved in suppressing endoplasmic reticulum and chaperone and signalling regulators in several human cancers; suggested as a tumour supressor in lung cancer initiation and progression. | [92,93,94,95,96] |
| hsa-miR-93 | Probably associated with the prognosis of bladder cancer; known to promote bladder cancer cells proliferation and invasion via targeting PEDF gene; involved in sensitivity of bladder cancer to chemotherapy; promotes hepatocellular carcinoma progression, possibly via miRNA-93-5p/MAPK/c-Jun positive feddback loop. | [97,98,99,100,101,102] |
| hsa-miR-96 | Oncogenic miR; potential urinary biomarker in bladder cancer; involved in EMT and migration and invasion of bladder cancer cells via targeting CDKN1A; impacts the response to chemotherapy. | [103,104,105,106,107,108] |
| hsa-miR-98 | Important in the development and progression of bladder cancer due to its involvement in the WNT/β-catenin pathway; promotes drug resistance via targeting the LASS2 gene; axis miR-98/IGF1 contributes to breast cancer progression. | [109,110,111,112,113] |
| hsa-miR-141 | Might be important in the wound-healing process; promotes bladder cancer progression, and thus, has prognostic value; in oesophageal cancer, promotes cell proliferation, migration and invasion; moreover, in ameloblastoma, it has been shown to supress cell migration via upregulation of the NCAM1 molecule. | [114,115,116,117] |
| hsa-miR-151a | Studied in atopic dermatitis, pain transmission and NOTCH2 signalling pathway; targeting CHL1 inhibits proliferation and invasion of colon cancer cells; in nasopharyngeal carcinoma, the inhibition of p53 by miR-151a induced cell proliferation, migration and possibly also invasion. | [118,119,120,121] |
| hsa-miR-182 | Potentially important in the bladder cancer development, proliferation and migration; inhibits inflammation, proliferation and migration of endometrial stromal cells through NFkB pathway deactivation. | [122,123,124,125] |
| hsa-miR-183 | Involved in the cell adhesion modulation and progression of laryngeal cancer; reports in bladder cancer indicated its crucial role in maintaining the canonical WNT signalling pathway, regulating growth and apoptosis; it has also been reported that cisplatin and paclitaxel significantly alter the expression of miR-183; potentially involved in hepatocellular carcinoma cells proliferation by LNC-HC gene inhibition. | [108,126,127,128,129] |
| hsa-miR-187 | Oncogene miR known to be involved in proliferation, migration, invasion and recurrence bladder cancer; regulates the WNT/β-catenin pathway. | [130,131] |
| hsa-miR-191 | Linked to the antiviral response and intracellular mechanisms of viral replication, e.g., may inhibit the replication of human immunodeficiency virus in human cells; circulating miR-191 has been proposed as a biomarker of breast cancer, as well as squamous cell carcinoma; it has been suggested to regulate endometrial cancer cell growth via TET1-mediated epigenetic modulation of APC gene. | [132,133,134,135] |
| hsa-miR-200a | Regulation of EMT; probably predictive for a patient’s prognosis in colorectal cancer; reported to be correlated with early-stage and T1 bladder tumour progression and bladder cancer invasion; when overexpressed, it promotes bladder cancer invasion. | [77,78] |
| hsa-miR-200b | Significant in renal cancer: it is often downregulated and may suppress metastasis by targeting LAMA4 in renal cell carcinoma; aberrant expression has been reported in HER-2 negative breast cancer as well as cardiological pathologies and angiogenesis aberrations; may affect breast cancer cells’ response to tamoxifen, involving MYB; it has been suggested as a prognostic marker in clear cell renal carcinoma; epigenetic silencing of miR-200b is connected with cisplatin resistance in bladder cancer. | [83,84,85,136,137,138] |
| hsa-miR-200c | Important in breast cancer pathogenesis and response to treatment, including trastuzumab use in HER2 positive breast cancer; might act protectively against colorectal cancer through BMI1 gene complex; it suppresses tumour metastasis by inhibiting EMT in oral squamous carcinoma; the role in bladder cancer remains unknown—the level of miR-200c in bladder cancer patients’ urine is significantly different from the level in healthy people, and thus, it has been suggested as a potential biomarker. | [32,87,88,89,90,91] |
| hsa-miR-219a | Inhibits colon cancer progression; enhances the radiosensitivity of lung cancer cells. | [139,140] |
| hsa-miR-301b | Accelerates the growth of gastric cancer; promotes the mobility, proliferation and EMT in bladder cancer by targeting EGR1; plasma levels of miR-301b might be a potential biomarker of early stage non-small-cell lung cancer. | [141,142,143,144] |
| hsa-miR-423 | It has been proposed as a biomarker, indicating early stages of bladder cancer, especially from blood serum and urine; suppression of miR-423 in breast cancer cells inhibited cell proliferation and invasion. | [46,145,146,147] |
| hsa-miR-429 | Known to be involved in the pathogenesis of many cancer types, including bladder; significant role in EMT in bladder cancer; expression levels have been correlated with patient outcomes; it seems to promote proliferation of bladder cancer cells via the inhibition of CDKN2B; elevated miR-429 supresses the progression of hyphopharyngeal squamous cells carcinoma by reducing ZEB1 expression. | [148,149,150,151,152] |
| hsa-miR-744 | Although its function in bladder cancer has to be revealed, it seems important in colorectal cancer, ovarian cancer and heart diseases, targeting the ARF1 gene; interestingly, it has been reported to be involved in regulation of the MHC class I gene expression; contributes to inflammation among patients with Sjorgen Syndrome; promotes proliferation of osteosarcoma cells by targeting PTEN. | [153,154,155,156,157] |
| hsa-miR-940 | Known to impact the aggressiveness of bladder cancer cells via activating the WNT/β-catenin pathway; suggested as a biomarker of gastric cancer; involved in the hepatocellular cancer and prostate cancer development. | [158,159,160,161] |
| hsa-miR-1287 | Probably regulates the MEK/ERK pathway; the role in immune surveillance remains unknown. | [162,163] |
| hsa-miR-1306 | Connected with the SIRT gene family expression; important in gastric cancer; its role in immune surveillance is to be revealed. | [164,165,166] |
| hsa-miR-1307 | Upregulation contributes to the progression of gastric cancer; indicates the metastatic potential of hepatocellular carcinoma; by targeting TRAF3 gene, it also regulates the MAPK/NFkB pathway in lung adenocarcinoma, promoting cancer cells’ proliferation. | [167,168,169] |
| hsa-miR-3193 | Very little is known about this miR; however, its role has been reported in melanoma pathogenesis. | [170] |
| hsa-miR-3200 | Acting in gastric cancer tumorigenesis and progression. | [171,172,173] |
| hsa-miR-4756 | Seems to be particularly important in triple-negative breast cancer pathogenesis; also reported in the EMT process. | [174,175] |
| hsa-miR-4778 | Might be involved in radioresistance development in cervical cancer. | [176] |
| hsa-miR-7706 | Little is known about this miR and its function in cancer pathogenesis; in hepatocellular carcinoma, it seems to have a prognostic value. | [177] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stempor, P.A.; Avni, D.; Leibowitz, R.; Sidi, Y.; Stępień, M.; Dzieciątkowski, T.; Dobosz, P. Comprehensive Analysis of Correlations in the Expression of miRNA Genes and Immune Checkpoint Genes in Bladder Cancer Cells. Int. J. Mol. Sci. 2021, 22, 2553. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052553
Stempor PA, Avni D, Leibowitz R, Sidi Y, Stępień M, Dzieciątkowski T, Dobosz P. Comprehensive Analysis of Correlations in the Expression of miRNA Genes and Immune Checkpoint Genes in Bladder Cancer Cells. International Journal of Molecular Sciences. 2021; 22(5):2553. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052553
Chicago/Turabian StyleStempor, Przemysław A., Dror Avni, Raya Leibowitz, Yechezkel Sidi, Maria Stępień, Tomasz Dzieciątkowski, and Paula Dobosz. 2021. "Comprehensive Analysis of Correlations in the Expression of miRNA Genes and Immune Checkpoint Genes in Bladder Cancer Cells" International Journal of Molecular Sciences 22, no. 5: 2553. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052553