
Shorter P1m Response in Children with Autism Spectrum Disorder without Intellectual Disabilities

 and
and
Abstract
:1. Introduction
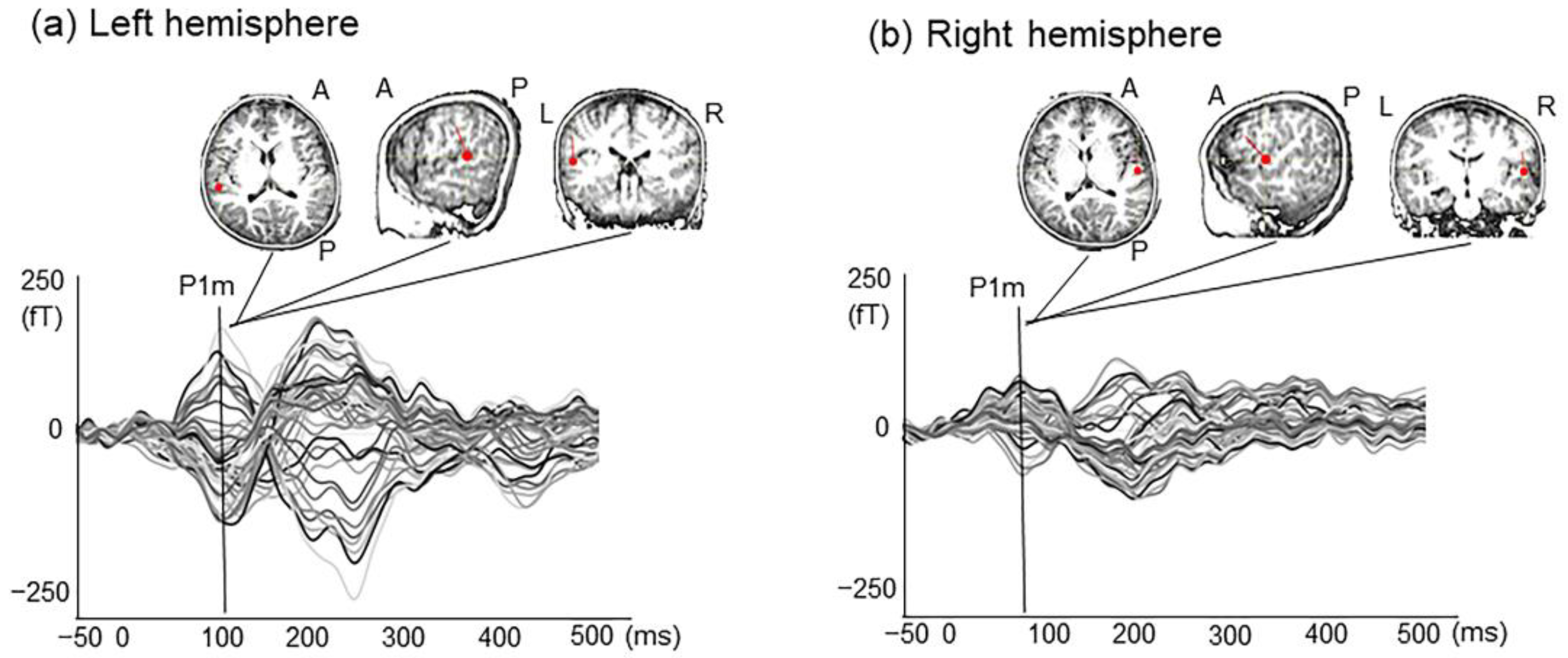
2. Results
2.1. P1m Latency and Intensity
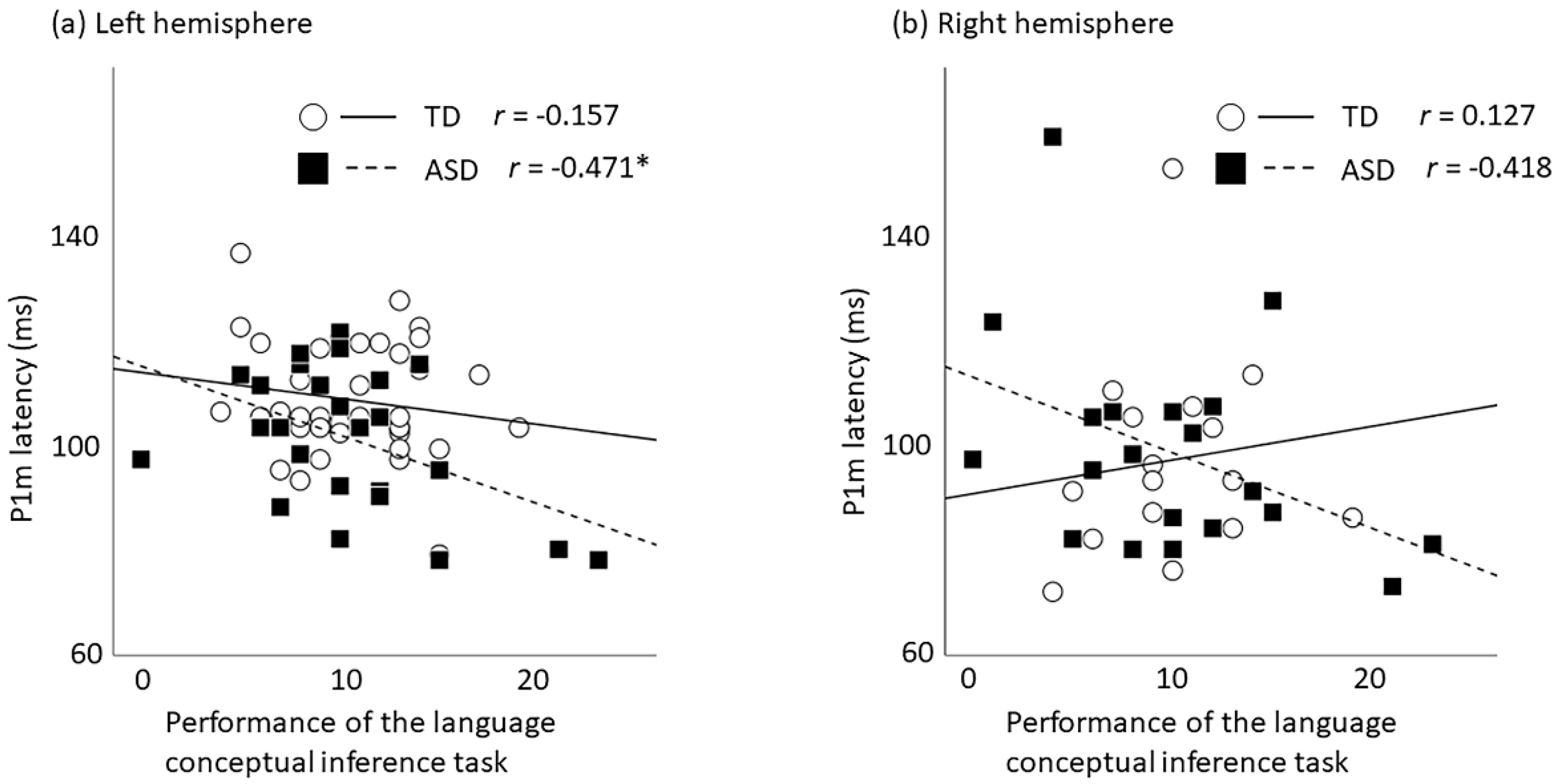
2.2. Correlation between P1m Latency and Language-Related Performances
2.3. Correlation between P1m Intensity and Language-Related Performances
3. Discussion
4. Materials and Methods
4.1. Participants
4.2. Cognitive and Language Performance Measurements
4.3. Auditory Stimuli
4.4. Magnetoencephalography Recording
4.5. Data Analysis
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Association: Philadelphia, PN, USA, 2013. [Google Scholar]
- Osterling, J.; Dawson, G. Early recognition of children with autism: A study of first birthday home videotapes. J. Autism Dev. Disord. 1994, 24, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Carper, R.A.; Moses, P.; Tigue, Z.D.; Courchesne, E. Cerebral lobes in autism: Early hyperplasia and abnormal age effects. Neuroimage 2002, 16, 1038–1051. [Google Scholar] [CrossRef]
- Schumann, C.M.; Bloss, C.S.; Barnes, C.C.; Wideman, G.M.; Carper, R.A.; Akshoomoff, N.; Pierce, K.; Hagler, D.; Schork, N.; Lord, C.; et al. Longitudinal magnetic resonance imaging study of cortical development through early childhood in autism. J. Neurosci. 2010, 30, 4419–4427. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.D.; Nordahl, C.W.; Young, G.S.; Wootton-Gorges, S.L.; Lee, A.; Liston, S.E.; Harrington, K.R.; Ozonoff, S.; Amaral, D.G. Early brain enlargement and elevated extra-axial fluid in infants who develop autism spectrum disorder. Brain 2013, 136, 2825–2835. [Google Scholar] [CrossRef] [Green Version]
- Hazlett, H.C.; Gu, H.; Munsell, B.C.; Kim, S.H.; Styner, M.; Wolff, J.J.; Elison, J.T.; Swanson, M.R.; Zhu, H.; Botteron, K.N.; et al. Early brain development in infants at high risk for autism spectrum disorder. Nature 2017, 542, 348–351. [Google Scholar] [CrossRef]
- Piven, J.; Elison, J.T.; Zylka, M.J. Toward a conceptual framework for early brain and behavior development in autism. Mol. Psychiatry 2017, 22, 1385–1394. [Google Scholar] [CrossRef] [Green Version]
- Cardy, J.E.O.; Flagg, E.J.; Roberts, W.; Roberts, T.P. Auditory evoked fields predict language ability and impairment in children. Int. J. Psychophysiol. 2008, 68, 170–175. [Google Scholar] [CrossRef]
- Matsuzaki, J.; Kagitani-Shimono, K.; Goto, T.; Sanefuji, W.; Yamamoto, T.; Sakai, S.; Uchida, H.; Hirata, M.; Mohri, I.; Yorifuji, S.; et al. Differential responses of primary auditory cortex in autistic spectrum disorder with auditory hypersensitivity. Neuroreport 2012, 23, 113–118. [Google Scholar] [CrossRef]
- Roberts, T.P.L.; Matsuzaki, J.; Blaskey, L.; Bloy, L.; Edgar, J.C.; Kim, M.; Ku, M.; Kuschner, E.S.; Embick, D. Delayed M50/M100 evoked response component latency in minimally verbal/nonverbal children who have autism spectrum disorder. Mol. Autism 2019, 10, 34. [Google Scholar] [CrossRef] [PubMed]
- Cardy, J.E.O.; Ferrari, P.; Flagg, E.J.; Roberts, W.; Roberts, T.P.L. Prominence of M50 auditory evoked response over M100 in childhood and autism. Neuroreport 2004, 15, 1867–1870. [Google Scholar] [CrossRef]
- Orekhova, E.V.; Butorina, A.V.; Tsetlin, M.M.; Novikova, S.I.; Sokolov, P.A.; Elam, M.; Stroganova, T.A. Auditory magnetic response to clicks in children and adults: Its components, hemispheric lateralization and repetition suppression effect. Brain Topogr. 2012, 26, 410–427. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, Y.; Hasegawa, C.; Ikeda, T.; Saito, D.N.; Hiraishi, H.; Takahashi, T.; Kumazaki, H.; Kikuchi, M. The maturation of the P1m component in response to voice from infancy to 3 years of age: A longitudinal study in young children. Brain Behav. 2020, 10, e01706. [Google Scholar] [CrossRef]
- Oram Cardy, J.E.; Flagg, E.J.; Roberts, W.; Brian, J.; Roberts, T.P. Magnetoencephalography identifies rapid temporal processing deficit in autism and language impairment. Neuroreport 2005, 16, 329–332. [Google Scholar] [CrossRef]
- Roberts, T.P.L.; Khan, S.Y.; Rey, M.; Monroe, J.F.; Cannon, K.; Blaskey, L.; Woldoff, S.; Qasmieh, S.; Gandal, M.; Schmidt, G.L.; et al. MEG detection of delayed auditory evoked responses in autism spectrum disorders: Towards an imaging biomarker for autism. Autism Res. 2010, 3, 8–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pihko, E.; Mickos, A.; Kujala, T.; Pihlgren, A.; Westman, M.; Alku, P.; Byring, R.; Korkman, M. Group intervention changes brain activity in bilingual language-impaired children. Cereb. Cortex 2006, 17, 849–858. [Google Scholar] [CrossRef]
- Pihko, E.; Kujala, T.; Mickos, A.; Alku, P.; Byring, R.; Korkman, M. Language impairment is reflected in auditory evoked fields. Int. J. Psychophysiol. 2008, 68, 161–169. [Google Scholar] [CrossRef]
- Tavabi, K.; Obleser, J.; Dobel, C.; Pantev, C. Auditory evoked fields differentially encode speech features: An MEG investigation of the P50m and N100m time courses during syllable processing. Eur. J. Neurosci. 2007, 25, 3155–3162. [Google Scholar] [CrossRef]
- Orekhova, E.V.; Tsetlin, M.M.; Butorina, A.V.; Novikova, S.I.; Gratchev, V.V.; Sokolov, P.A.; Elam, M.; Stroganova, T.A. Auditory cortex responses to clicks and sensory modulation difficulties in children with Autism Spectrum Disorders (ASD). PLoS ONE 2012, 7, e39906. [Google Scholar] [CrossRef] [Green Version]
- Yoshimura, Y.; Kikuchi, M.; Shitamichi, K.; Ueno, S.; Munesue, T.; Ono, Y.; Tsubokawa, T.; Haruta, Y.; Oi, M.; Niida, Y.; et al. Atypical brain lateralisation in the auditory cortex and language performance in 3- to 7-year-old children with high-functioning autism spectrum disorder: A child-customised mag-netoencephalography (MEG) study. Mol. Autism 2013, 4, 38. [Google Scholar] [CrossRef] [Green Version]
- Yoshimura, Y.; Kikuchi, M.; Hiraishi, H.; Hasegawa, C.; Takahashi, T.; Remijn, G.B.; Oi, M.; Munesue, T.; Higashida, H.; Minabe, Y. Synchrony of auditory brain responses predicts behavioral ability to keep still in children with autism spectrum disorder: Auditory-evoked response in children with autism spectrum disorder. NeuroImage Clin. 2016, 12, 300–305. [Google Scholar] [CrossRef] [Green Version]
- Yoshimura, Y.; Kikuchi, M.; Ueno, S.; Shitamichi, K.; Remijn, G.B.; Hiraishi, H.; Hasegawa, C.; Furutani, N.; Oi, M.; Munesue, T.; et al. A longitudinal study of auditory evoked field and language development in young children. Neuroimage 2014, 101, 440–447. [Google Scholar] [CrossRef] [Green Version]
- Edgar, J.C.; Lanza, M.R.; Daina, A.B.; Monroe, J.F.; Khan, S.Y.; Blaskey, L.; Cannon, K.M.; Jenkins, J., III; Qasmieh, S.; Levy, S.E.; et al. Missing and delayed auditory responses in young and older children with autism spectrum disorders. Front. Hum. Neurosci. 2014, 8, 417. [Google Scholar] [CrossRef] [Green Version]
- Kaufman, A.; Kaufman, N. Kaufman Assessment Battery for Children, 2nd ed.; American Guidance Service: Circle Pines, MN, USA, 1983. [Google Scholar]
- Matsuzaki, J.; Ku, M.; DiPiero, M.; Chiang, T.; Saby, J.; Blaskey, L.; Kuschner, E.S.; Kim, M.; Berman, J.I.; Bloy, L.; et al. Delayed Auditory Evoked Responses in Autism Spectrum Disorder across the Life Span. Dev. Neurosci. 2019, 41, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Bloy, L.; Ku, M.; Edgar, J.C.; Miller, J.S.; Blaskey, L.; Ross, J.; Roberts, T.P. Auditory evoked response delays in children with 47,XYY syndrome. Neuroreport 2019, 30, 504–509. [Google Scholar] [CrossRef]
- Williams, Z.J.; Abdelmessih, P.G.; Key, A.P.; Woynaroski, T.G. Cortical auditory processing of simple stimuli is altered in autism: A meta-analysis of auditory evoked responses. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2020. [Google Scholar] [CrossRef] [PubMed]
- Stephen, J.M.; Hill, D.E.; Peters, A.; Flynn, L.; Zhang, T.; Okada, Y. Development of auditory evoked responses in normally developing preschool children and children with autism spectrum disorder. Dev. Neurosci. 2017, 39, 430–441. [Google Scholar] [CrossRef] [PubMed]
- Donkers, F.C.L.; Schipul, S.E.; Baranek, G.T.; Cleary, K.M.; Willoughby, M.T.; Evans, A.M.; Bulluck, J.C.; Lovmo, J.E.; Belger, A. Attenuated Auditory event-related potentials and associations with atypical sensory response patterns in children with autism. J. Autism Dev. Disord. 2015, 45, 506–523. [Google Scholar] [CrossRef] [Green Version]
- Orekhova, E.V.; Stroganova, T.A.; Prokofyev, A.O.; Nygren, G.; Gillberg, C.; Elam, M. Sensory gating in young children with autism: Relation to age, IQ, and EEG gamma oscillations. Neurosci. Lett. 2008, 434, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Roncagliolo, M.; Benítez, J.; Eguibar, J.R. Progressive deterioration of central components of auditory brainstem responses during postnatal development of the myelin mutant taiep rat. Audiol. Neurotol. 2000, 5, 267–275. [Google Scholar] [CrossRef]
- Lee, D.L.; Strathmann, F.G.; Gelein, R.; Walton, J.; Mayer-Proschel, M. Iron deficiency disrupts axon maturation of the developing auditory nerve. J. Neurosci. 2012, 32, 5010–5015. [Google Scholar] [CrossRef] [PubMed]
- Daneshvarfard, F.; Abrishami Moghaddam, H.; Kongolo, G.; Wallois, F.; Mahmoudzadeh, M. Functional and structural cor-relates of the preterm infant’s brain: Relating developmental changes of auditory evoked responses to structural maturation. Brain Struct. Funct. 2020, 225, 2165–2176. [Google Scholar] [CrossRef] [PubMed]
- Yakovlev, P.I.; Lecours, A.R. The Myelogenetic Cycles of Regional Maturation of the Brain. Regional Development of Brain Early Life; Minkocoaki, A., Ed.; Blackwell Scientific: Oxford, UK, 1967. [Google Scholar]
- Kinney, H.C.; Brody, B.A.; Kloman, A.S.; Gilles, F.H. Sequence of central nervous system myelination in human infancy. II. Patterns of myelination in autopsied Infants. J. Neuropathol. Exp. Neurol. 1988, 47, 217–234. [Google Scholar] [CrossRef] [Green Version]
- Roberts, T.P.L.; Khan, S.Y.; Blaskey, L.; Dell, J.; Levy, S.E.; Zarnow, D.M.; Edgar, J.C. Developmental correlation of diffusion anisotropy with auditory-evoked response. Neuroreport 2009, 20, 1586–1591. [Google Scholar] [CrossRef]
- Wolff, J.J.; Gu, H.; Gerig, G.; Elison, J.T.; Styner, M.; Gouttard, S.; Botteron, K.N.; Dager, S.R.; Dawson, G.; Estes, A.M.; et al. Differences in white matter fiber tract development present from 6 to 24 months in infants with autism. Am. J. Psychiatry 2012, 169, 589–600. [Google Scholar] [CrossRef]
- Redcay, E.; Courchesne, E. When is the brain enlarged in autism? A meta-analysis of all brain size reports. Biol. Psychiatry 2005, 58, 1–9. [Google Scholar] [CrossRef]
- Uddin, L.Q.; Supekar, K.; Menon, V. Reconceptualizing functional brain connectivity in autism from a developmental perspective. Front. Hum. Neurosci. 2013, 7, 458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pujol, J.; Soriano-Mas, C.; Ortiz, H.; Sebastian-Galles, N.; Losilla, J.M.; Deus, J. Myelination of language-related areas in the developing brain. Neurology 2006, 66, 339–343. [Google Scholar] [CrossRef]
- Abrams, D.A.; Lynch, C.J.; Cheng, K.M.; Phillips, J.; Supekar, K.; Ryali, S.; Uddin, L.Q.; Menon, V. Underconnectivity between voice-selective cortex and reward circuitry in children with autism. Proc. Natl. Acad. Sci. USA 2013, 110, 12060–12065. [Google Scholar] [CrossRef] [Green Version]
- Lynch, C.J.; Uddin, L.Q.; Supekar, K.; Khouzam, A.; Phillips, J.; Menon, V. Default mode network in childhood autism: Posteromedial cortex heterogeneity and relationship with social deficits. Biol. Psychiatry 2013, 74, 212–219. [Google Scholar] [CrossRef] [Green Version]
- Friederici, A.D.; Rüschemeyer, S.-A.; Hahne, A.; Fiebach, C.J. The Role of Left Inferior Frontal and Superior Temporal Cortex in Sentence Comprehension: Localizing Syntactic and Semantic Processes. Cereb. Cortex 2003, 13, 170–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hickok, G.; Poeppel, D. Dorsal and ventral streams: A framework for understanding aspects of the functional anatomy of language. Cognition 2004, 92, 67–99. [Google Scholar] [CrossRef] [PubMed]
- Catani, M.; Jones, D.K.; Ffytche, D.H. Perisylvian language networks of the human brain. Ann. Neurol. 2005, 57, 8–16. [Google Scholar] [CrossRef]
- Rolheiser, T.; Stamatakis, E.A.; Tyler, L.K. Dynamic processing in the human language system: Synergy between the arcuate fascicle and extreme capsule. J. Neurosci. 2011, 31, 16949–16957. [Google Scholar] [CrossRef] [Green Version]
- Demopoulos, C.; Lewine, J.D. audiometric profiles in autism spectrum disorders: Does Subclinical hearing loss impact communication? Autism Res. 2016, 9, 107–120. [Google Scholar] [CrossRef] [Green Version]
- Lord, C.; Risi, S.; Lambrecht, L.; Cook, E.H.; Leventhal, B.L.; DiLavore, P.C.; Pickles, A.; Rutter, M. The autism diagnostic observation schedule-generic: A standard measure of social and communication deficits associated with the spectrum of autism. J. Autism Dev. Disord. 2000, 30, 205–223. [Google Scholar] [CrossRef]
- Lord, C.; Rutter, M.; DiLavore, P.C. Autism Diagnostic Observation Schedule, 2nd ed.; Western Psychological Services: Gladstone, OR, USA, 2012. [Google Scholar]
- Wing, L.; Leekam, S.R.; Libby, S.J.; Gould, J.; Larcombe, M. The diagnostic interview for social and communication disorders: Background, inter-rater reliability and clinical use. J. Child Psychol. Psychiatry 2002, 43, 307–325. [Google Scholar] [CrossRef] [Green Version]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 4th ed.; American Psychiatric Association: Washington, DC, USA, 1994. [Google Scholar]
- Oldfield, R. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychology 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Yoshimura, Y.; Kikuchi, M.; Ueno, S.; Okumura, E.; Hiraishi, H.; Hasegawa, C.; Remijn, G.B.; Shitamichi, K.; Munesue, T.; Tsubokawa, T.; et al. The brain’s response to the human voice depends on the incidence of autistic traits in the general population. PLoS ONE 2013, 8, e80126. [Google Scholar] [CrossRef] [Green Version]
- Yoshimura, Y.; Kikuchi, M.; Shitamichi, K.; Ueno, S.; Remijn, G.B.; Haruta, Y.; Oi, M.; Munesue, T.; Tsubokawa, T.; Higashida, H.; et al. Language performance and auditory evoked fields in 2- to 5-year-old children. Eur. J. Neurosci. 2012, 35, 644–650. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| TD | ASD | t | p | |
|---|---|---|---|---|
| Latency (ms) | ||||
| Left (N) | 38 | 26 | ||
| 108 (10.8) | 101 (13.0) | 2.390 | 0.020 * | |
| Right (N) | 16 | 20 | ||
| 97 (18.9) | 98 (20.0) | −0.232 | 0.818 | |
| Intensity (nAm) | ||||
| Left (N) | 38 | 26 | ||
| 18.5 (6.5) | 18.4 (7.8) | 0.059 | 0.953 | |
| Right (N) | 16 | 20 | ||
| 13.3 (5.0) | 15.2 (6.8) | −0.966 | 0.341 |
| TD | ASD | |||
|---|---|---|---|---|
| r | p | r | p | |
| Latency | ||||
| Left | −0.157 | 0.346 | −0.471 | 0.015 * |
| Right | 0.127 | 0.640 | −0.418 | 0.067 |
| Intensity | ||||
| Left | 0.205 | 0.217 | −0.102 | 0.619 |
| Right | 0.523 | 0.038 | −0.333 | 0.151 |
| TD | ASD | t | p | |
|---|---|---|---|---|
| Number of participants | 46 | 29 | ||
| Gender (male/female) | 41/5 | 21/8 | n.s. | |
| Chronological age (months) | 70.3 (63–91) | 74.7 (60–98) | n.s. | |
| Handedness (right/left) | 42/4 | 27/2 | n.s. | |
| K-ABC 1–MPS 2 | 104.7 (12.9) | 91.4 (11.7) | 4.488 | <0.001 |
| K-ABC 1–AS 3 | 101.4 (13.9) | 95.2 (13.4) | 1.913 | n.s. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoshimura, Y.; Ikeda, T.; Hasegawa, C.; An, K.-M.; Tanaka, S.; Yaoi, K.; Iwasaki, S.; Saito, D.N.; Kumazaki, H.; Hiraishi, H.; et al. Shorter P1m Response in Children with Autism Spectrum Disorder without Intellectual Disabilities. Int. J. Mol. Sci. 2021, 22, 2611. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052611
Yoshimura Y, Ikeda T, Hasegawa C, An K-M, Tanaka S, Yaoi K, Iwasaki S, Saito DN, Kumazaki H, Hiraishi H, et al. Shorter P1m Response in Children with Autism Spectrum Disorder without Intellectual Disabilities. International Journal of Molecular Sciences. 2021; 22(5):2611. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052611
Chicago/Turabian StyleYoshimura, Yuko, Takashi Ikeda, Chiaki Hasegawa, Kyung-Min An, Sanae Tanaka, Ken Yaoi, Sumie Iwasaki, Daisuke N. Saito, Hirokazu Kumazaki, Hirotoshi Hiraishi, and et al. 2021. "Shorter P1m Response in Children with Autism Spectrum Disorder without Intellectual Disabilities" International Journal of Molecular Sciences 22, no. 5: 2611. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052611