
Trans-Cinnamaldehyde Exhibits Synergy with Conventional Antibiotic against Methicillin-Resistant Staphylococcus aureus


Abstract
:1. Introduction
2. Results
2.1. Synergistic Antibacterial Activity
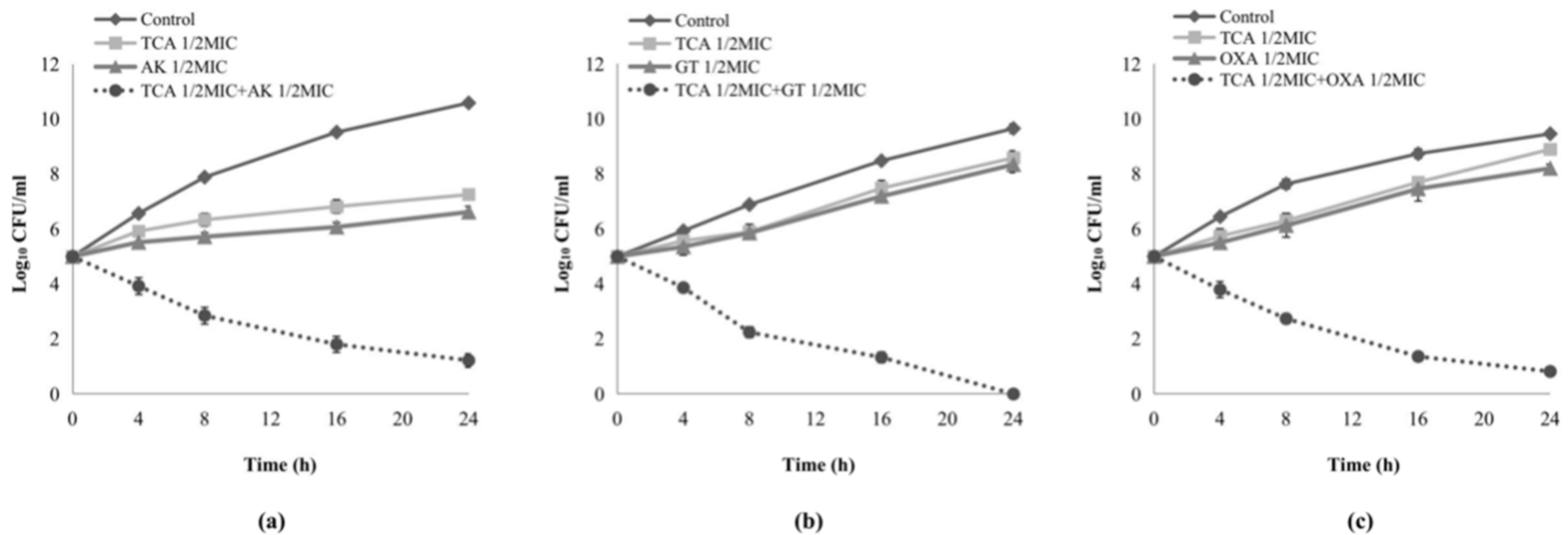
2.2. Time–Kill Assay
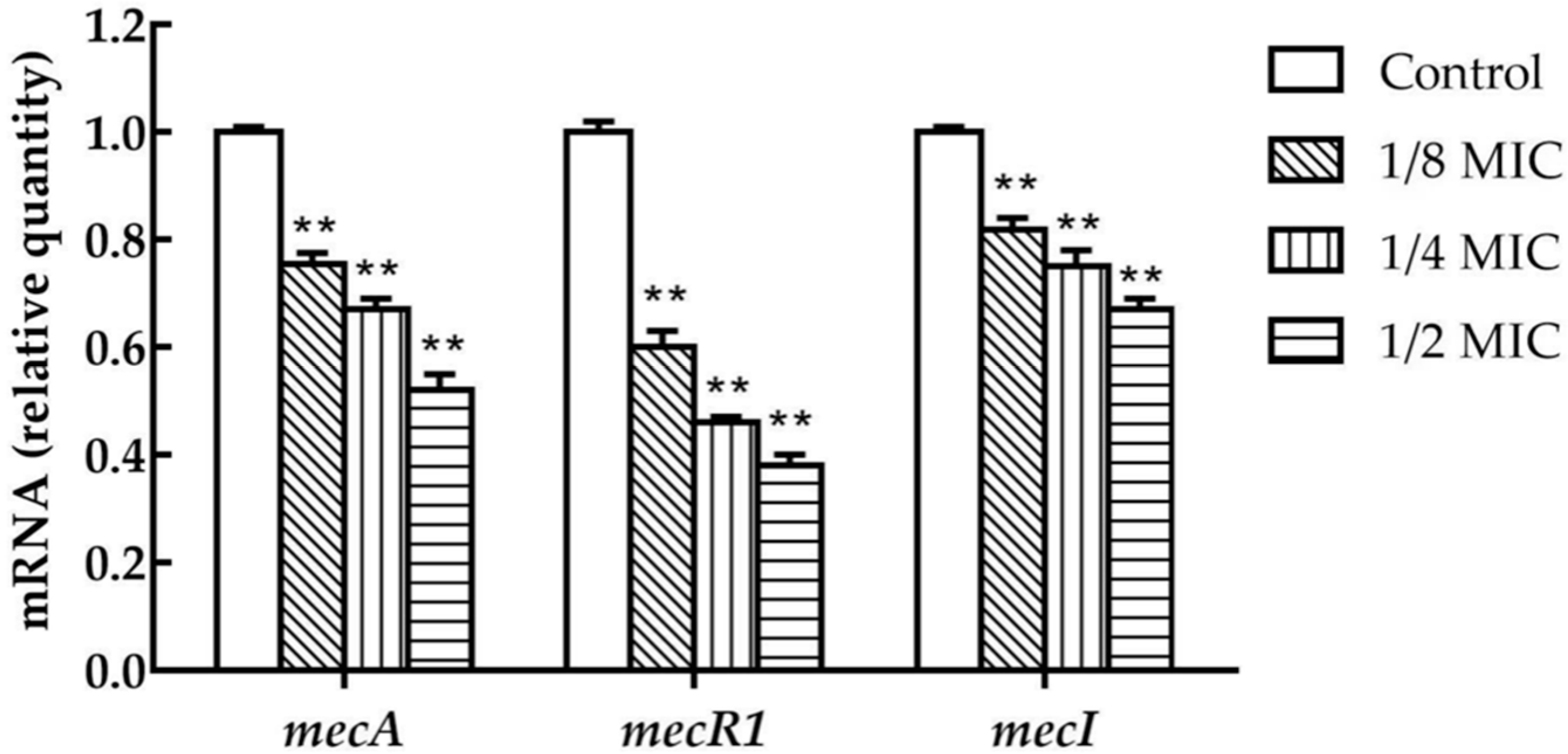
2.3. TCA Represses the Transcription of MecA, MecR1, and MecI in S. aureus
2.4. Expression of PBP2a in MRSA Treated with TCA
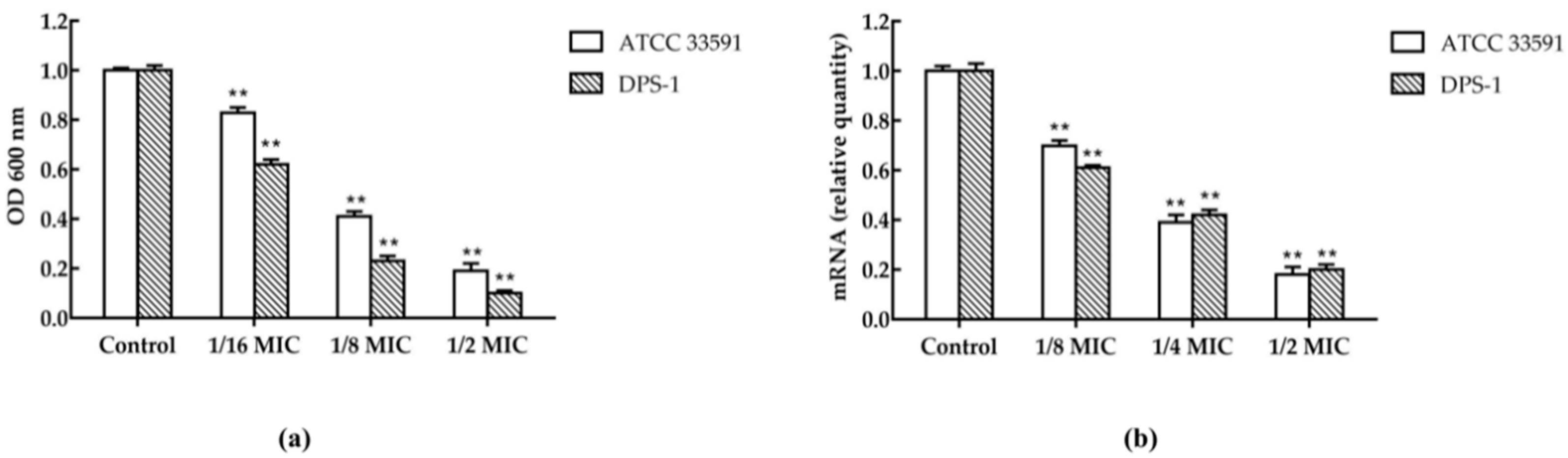
2.5. Biofilm Inhibitory Assay and the Transcription of hld in S. aureus
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Susceptibility Testing of TCA with Conventional Antibiotics
4.3. Bacterial Strains and Conditions
4.4. Time–Kill Assays
4.5. Western Blot Analysis
4.6. qRT-PCR
4.7. Biofilm Inhibition Assay
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Peacock, S.J.; Paterson, G.K. Mechanisms of Methicillin Resistance in Staphylococcus aureus. Annu. Rev. Biochem. 2015, 4, 577–601. [Google Scholar] [CrossRef]
- Jakab, E.; Colcieru, M.; Jakab, R.I.; Gabri, E.E.; Lázár, C.H.; Török, E.; Kremmer, B.; Mészáros, T.J.; Katona, Á.; Fazakas, E.; et al. Screening of mecI Gene in Staphylococcus Strains Isolated in Transylvania Region of Romania. Microb. Drug Resist. 2019, 25, 639–643. [Google Scholar] [CrossRef] [PubMed]
- Yoneda, A.; Thänert, R.; Burnham, C.D.; Dantas, G. In vitro activity of meropenem/piperacillin/tazobactam triple combination therapy against clinical isolates of Staphylococcus aureus, Staphylococcus epidermidis, Staphylococcus pseudintermedius and vancomycin-resistant Enterococcus spp. Int. J. Antimicrob. Agents 2020, 55, 105864. [Google Scholar] [CrossRef]
- Von Dach, E.; Morel, C.M.; Murthy, A.; Pagani, L.; Macedo-Vinas, M.; Olearo, F.; Harbarth, S. Comparing the cost-effectiveness of linezolid to trimethoprim/sulfamethoxazole plus rifampicin for the treatment of methicillin-resistant Staphylococcus aureus infection: A healthcare system perspective. Clin. Microbiol. Infect. 2017, 23, 659–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; El-Hossary, E.M.; Oelschlaeger, T.A.; Donia, M.S.; Quinn, R.J.; Abdelmohsen, U.R. Potential of marine natural products against drug-resistant bacterial infections. Lancet Infect. Dis. 2019, 19, 237–245. [Google Scholar] [CrossRef]
- Mooyottu, S.; Kollanoor-Johny, A.; Flock, G.; Bouillaut, L.; Upadhyay, A.; Sonenshein, A.L.; Venkitanarayanan, K. Carvacrol and trans-cinnamaldehyde reduce Clostridium difficile toxin production and cytotoxicity in vitro. Int. J. Mol. Sci. 2014, 15, 4415–4430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, A.L.; Li, G.H.; Li, Y.R.; Wu, X.Y.; Ren, D.M.; Lou, H.X.; Wang, X.N.; Shen, T. Lignan and flavonoid support the prevention of cinnamon against oxidative stress related diseases. Phytomedicine 2019, 53, 143–153. [Google Scholar] [CrossRef]
- Yemiş, G.P.; Delaquis, P. Natural Compounds with Antibacterial Activity Against Cronobacter spp. in Powdered Infant Formula: A Review. Front. Nutr. 2020, 7, 595964. [Google Scholar] [CrossRef]
- Huang, X.; Wang, P.; Li, T.; Tian, X.; Guo, W.; Xu, B.; Huang, G.; Cai, D.; Zhou, F.; Zhang, H.; et al. Self-Assemblies Based on Traditional Medicine Berberine and Cinnamic Acid for Adhesion-Induced Inhibition Multidrug-Resistant Staphylococcus aureus. ACS Appl. Mater. Interfaces 2020, 12, 227–237. [Google Scholar] [CrossRef]
- Sun, P.; Li, K.; Wang, T.; Ji, J.; Wang, Y.; Chen, K.X.; Jia, Q.; Li, Y.M.; Wang, H.Y. Procyanidin C1, a Component of Cinnamon Extracts, Is a Potential Insulin Sensitizer That Targets Adipocytes. J. Agric. Food Chem. 2019, 32, 8839–8846. [Google Scholar] [CrossRef]
- Yang, G.; Jin, T.; Yin, S.; Guo, D.; Zhang, C.; Xia, X.; Shi, C. trans-Cinnamaldehyde mitigated intestinal inflammation induced by Cronobacter sakazakii in newborn mice. Food Funct. 2019, 10, 2986–2996. [Google Scholar] [CrossRef]
- Narayanan, A.; Muyyarikkandy, M.S.; Mooyottu, S.; Venkitanarayanan, K.; Amalaradjou, M.A. Oral supplementation of trans-cinnamaldehyde reduces uropathogenic Escherichia coli colonization in a mouse model. Lett. Appl. Microbiol. 2017, 64, 192–197. [Google Scholar] [CrossRef] [PubMed]
- Rogiers, G.; Kebede, B.T.; Van Loey, A.; Michiels, C.W. Membrane fatty acid composition as a determinant of Listeria monocytogenes sensitivity to trans-cinnamaldehyde. Res. Microbiol. 2017, 168, 536–546. [Google Scholar] [CrossRef]
- Hakimi Alni, R.; Tavasoli, F.; Barati, A.; Shahrokhi Badarbani, S.; Salimi, Z.; Babaeekhou, L. Synergistic activity of melittin with mupirocin: A study against methicillin-resistant S. Aureus (MRSA) and methicillin-susceptible S. Aureus (MSSA) isolates. Saudi J. Biol. Sci. 2020, 27, 2580–2585. [Google Scholar] [CrossRef] [PubMed]
- Oh, E.; Bae, J.; Kumar, A.; Choi, H.J.; Jeon, B. Antioxidant-based synergistic eradication of methicillin-resistant Staphylococcus aureus (MRSA) biofilms with bacitracin. Int. J. Antimicrob. Agents 2018, 52, 96–99. [Google Scholar] [CrossRef]
- Vasconcelos, N.G.; Croda, J.; Simionatto, S. Antibacterial mechanisms of cinnamon and its constituents: A review. Microb. Pathog. 2018, 120, 198–203. [Google Scholar] [CrossRef]
- Rai, M.; Paralikar, P.; Jogee, P.; Agarkar, G.; Ingle, A.P.; Derita, M.; Zacchino, S. Synergistic antimicrobial potential of essential oils in combination with nanoparticles: Emerging trends and future perspectives. Int. J. Pharm. 2017, 519, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Kluytmans, J.; Harbarth, S. MRSA transmission in the community: Emerging from under the radar. Lancet Infect. Dis. 2020, 20, 147–149. [Google Scholar] [CrossRef]
- Catteau, L.; Reichmann, N.T.; Olson, J.; Pinho, M.G.; Nizet, V.; Van Bambeke, F.; Quetin-Leclercq, J. Synergy between Ursolic and Oleanolic Acids from Vitellaria paradoxa Leaf Extract and β-Lactams against Methicillin-Resistant Staphylococcus aureus: In Vitro and In Vivo Activity and Underlying Mechanisms. Molecules. 2017, 12, 2245. [Google Scholar] [CrossRef] [Green Version]
- Al-Ani, I.; Zimmermann, S.; Reichling, J.; Wink, M. Antimicrobial Activities of European Propolis Collected from Various Geographic Origins Alone and in Combination with Antibiotics. Medicines 2018, 5, 2. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Kim, M.C.; Kang, O.H.; Kwon, D.Y. The Mechanism of Bisdemethoxycurcumin Enhances Conventional Antibiotics against Methicillin-Resistant Staphylococcus aureus. Int. J. Mol. Sci. 2020, 21, 7945. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Luo, J.; Liu, X.Q.; Kang, O.H.; Kwon, D.Y. Antibacterial activity and synergy of antibiotics with sanguisorbigenin isolated from Sanguisorba officinalis L. against methicillin-resistant Staphylococcus aureus. Lett. Appl. Microbiol. 2020, 72, 13417. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotics | ATCC 33591 | DPS-1 | DPS-3 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| MIC (μg/mL) | Fold | FICI | MIC (μg/mL) | Fold | FICI | MIC (μg/mL) | Fold | FICI | |
| Ampicillin | 62.5 | 2 | 1 | 31.3 | 4 | 0.75 | 62.5 | 4 | 0.75 |
| Oxacillin | 62.5 | 4 | 0.75 | 500 | 2 | 1 | 500 | 8 | 0.25 |
| Gentamicin | 3.9 | 4 | 0.75 | 125 | 4 | 0.37 | 250 | 4 | 0.5 |
| Vancomycin | 250 | 8 | 0.25 | 250 | 2 | 1 | 500 | 2 | 1 |
| Amoxicillin | 62.5 | 8 | 0.63 | 125 | 4 | 0.5 | 125 | 8 | 0.25 |
| Ceftazidine | 125 | 2 | 1 | 125 | 2 | 1 | 250 | 4 | 0.75 |
| Amikacin | 31.2 | 4 | 0.38 | 31.2 | 8 | 0.25 | 62.5 | 16 | 0.19 |
| Cefoxitin | 31.2 | 4 | 0.75 | 62.5 | 8 | 0.62 | 250 | 4 | 0.5 |
| Primer | Sequence (5′-3′) |
|---|---|
| 16S | F: ACTCCTACGGGAGGCAGCAG |
| R: ATTACCGCGGCTGCTGG | |
| mecA | F: CAATGCCAAAATCTCAGGTAAAGTG |
| R: AACCATCGTTACGGATTGCTTC | |
| mecR1 | F: GTGCTCGTCTCCACGTTAATTCCA |
| R: GACTAACCGAAGAAGTCGTGTCAG | |
| mecI | F: CGTTATAAGTGTACGAATGGTTTTTG |
| R: TCATCTGCAGAATGGGAAGTT | |
| hld | F: ATTTGTTCACTGTGTCGATAATCC |
| R: GGAGTGATTTCAATGGCACAAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Kang, O.-H.; Kwon, D.-Y. Trans-Cinnamaldehyde Exhibits Synergy with Conventional Antibiotic against Methicillin-Resistant Staphylococcus aureus. Int. J. Mol. Sci. 2021, 22, 2752. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052752
Wang S, Kang O-H, Kwon D-Y. Trans-Cinnamaldehyde Exhibits Synergy with Conventional Antibiotic against Methicillin-Resistant Staphylococcus aureus. International Journal of Molecular Sciences. 2021; 22(5):2752. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052752
Chicago/Turabian StyleWang, Shu, Ok-Hwa Kang, and Dong-Yeul Kwon. 2021. "Trans-Cinnamaldehyde Exhibits Synergy with Conventional Antibiotic against Methicillin-Resistant Staphylococcus aureus" International Journal of Molecular Sciences 22, no. 5: 2752. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052752